
Impact of 2′-Fucosyllactose on Gut Microbiota Composition in Adults with Chronic Gastrointestinal Conditions: Batch Culture Fermentation Model and Pilot Clinical Trial Findings



Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro Experiments
2.1.1. 2′-FL and Probiotic Strains
2.1.2. Batch Cultures
2.1.3. Flow Cytometry-Fluorescence In Situ Hybridization for Bacterial Enumerations

{kind=link}
{kind=link}
| Probe Name | Sequence (5′ to 3′) | Targeted Bacterial Group | Reference |
|---|---|---|---|
| Non Eub | ACTCCTACGGGAGGCAGC | Negative control | Wallner et al., 1993 [44] |
| Eub338 | GCTGCCTCCCGTAGGAGT | Total bacteria | Daims et al., 1999 [45] |
| Eub338II | GCAGCCACCCGTAGGTGT | Total bacteria | Daims et al., 1999 [45] |
| Eub338III | GCTGCCACCCGTAGGTGT | Total bacteria | Daims et al., 1999 [45] |
| Ato291 | GGTCGGTCTCTCAACCC | Atopobium cluster | Harmsen et al., 2000 [46] |
| Bac303 | CCAATGTGGGGGACCTT | Bacteroides/Prevotella | Manz et al., 1996 [47] |
| Bif164 | CATCCGGCATTACCACCC | Bifidobacterium spp. | Langendijk et al., 1995 [48] |
| Chis150 | TTATGCGGTATTAATCTYCCTTT | Clostridium histolyticum (Clostridium cluster I and II) | Franks et al., 1998 [49] |
| Prop853 | ATTGCGTTAACTCCGGCAC | Clostridium cluster IX | Walker et al., 2005 [50] |
| DSV687 | TACGGATTTCACTCCT | Desulfovibrio spp. | Hold et al., 2003 [51] |
| Erec482 | GCTTCTTAGTCARGTACCG | Eubacterium rectale/Clostridium coccoides (Clostridium cluster IVXa and IVXb) | Franks et al., 1998 [49] |
| Fprau655 | CGCCTACCTCTGCACTAC | Faecalibacterium prausnitzii | Devereux et al., 1992 [52] |
| Lab158 | GGTATTAGCAYCTGTTTCCA | Lactobacillus/Enterococcus | Harmsen et al., 1999 [53] |
| Rrec584 | TCAGACTTGCCGYACCGC | Roseburia spp. | Walker et al., 2005 [50] |
2.1.4. In Vitro SCFA Analysis
2.1.5. Statistical Analysis of In Vitro Data
2.2. Pilot Clinical Trial
2.2.1. Clinical Trial Design
2.2.2. Clinical Trial Participants and Recruitment
2.2.3. Clinical Trial Intervention
2.2.4. Quality of Life Assessment and Stool Sample Analysis
2.2.5. Statistical Analysis of Clinical Trial Data
3. Results
3.1. In Vitro Study
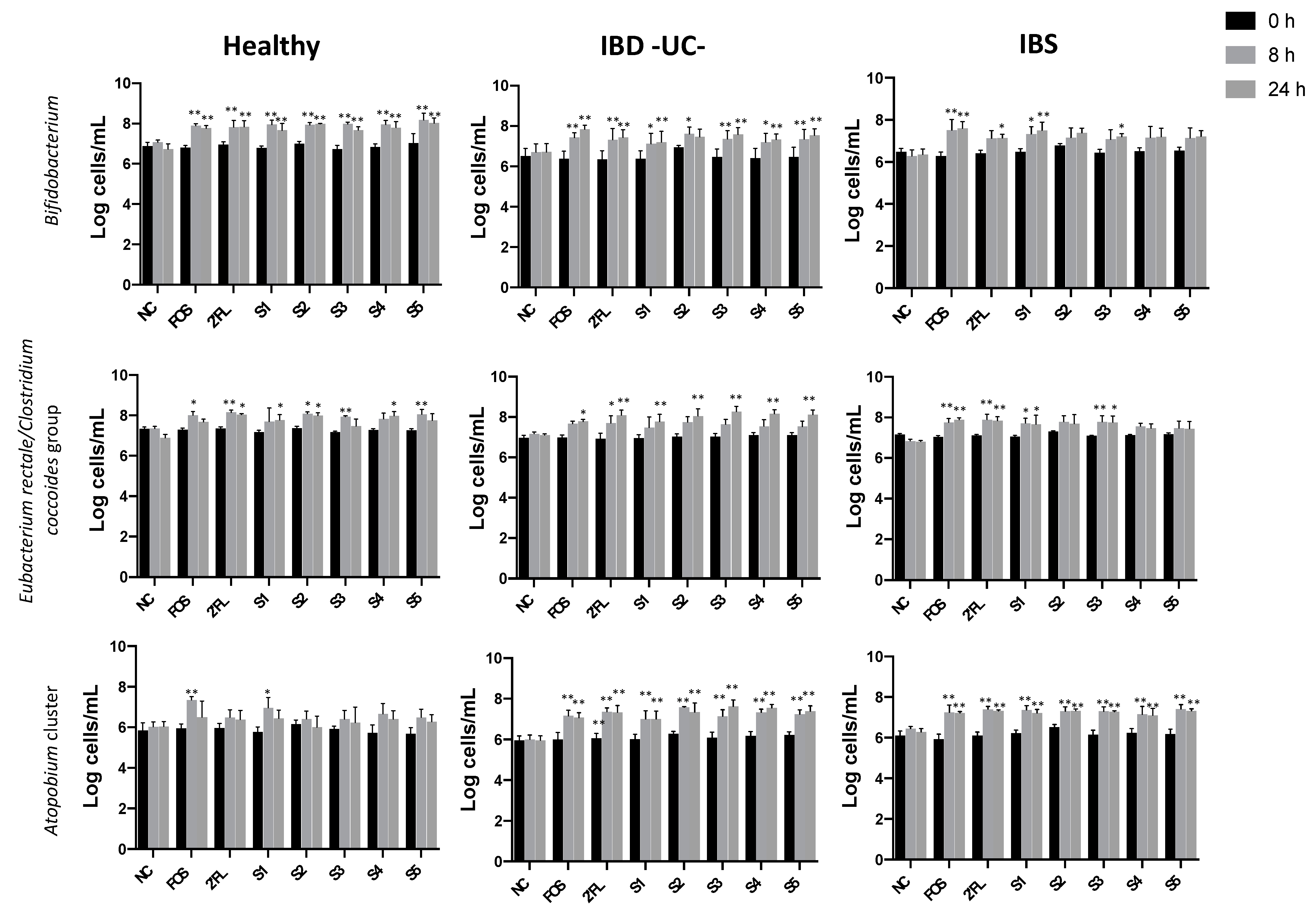
3.1.1. Effect of Fermentation on Bacterial Populations
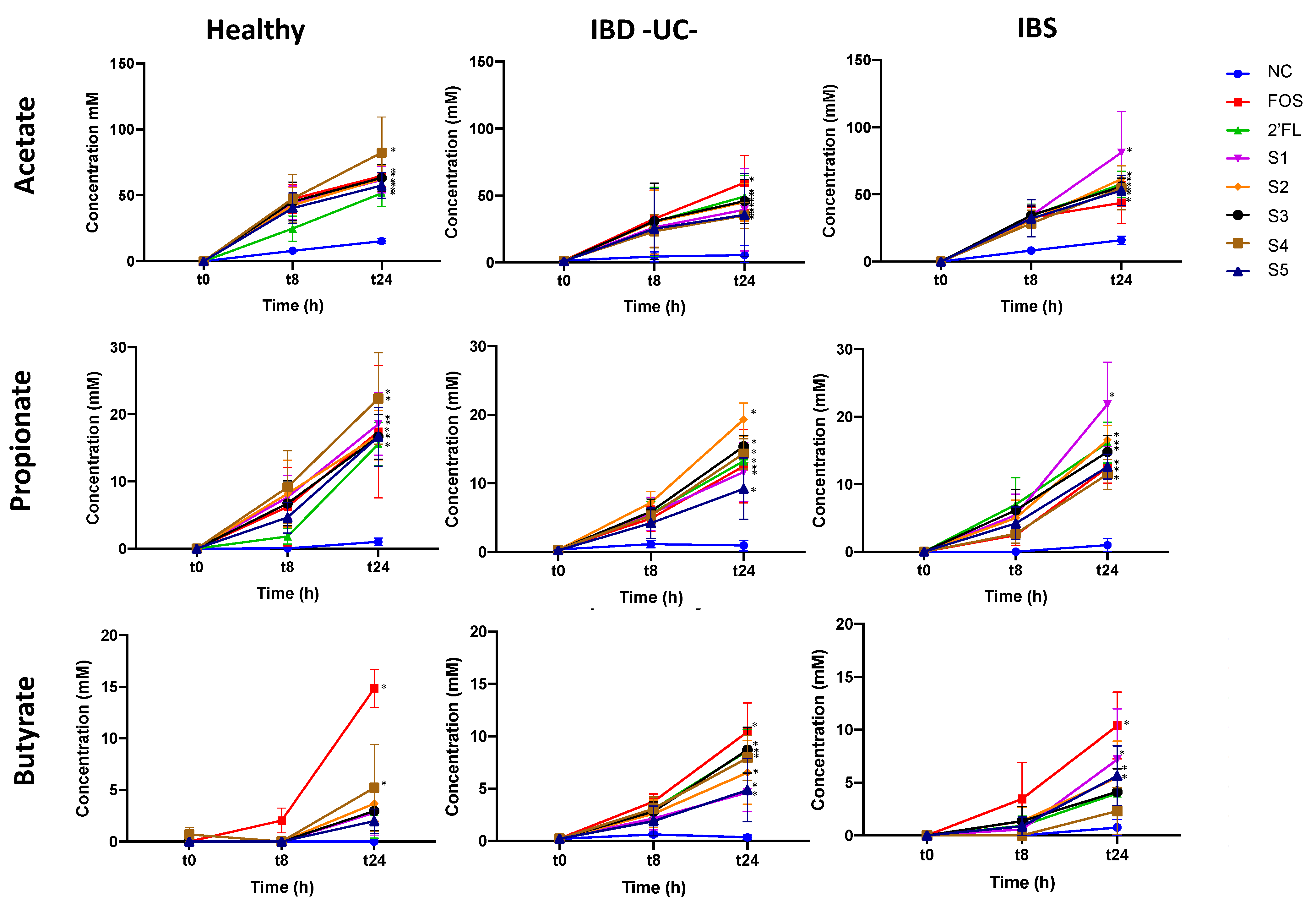
3.1.2. Effect of Fermentation on Short-Chain Fatty Acid (SCFA) Production
3.2. Pilot Clinical Trial
3.2.1. Participant Characteristics
3.2.2. Impact on Gastrointestinal Quality of Life
3.2.3. Impact on Stool Bacteria Counts
3.2.4. Impact on Stool Short-Chain Fatty Acids (SCFAs)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saha, L. Irritable bowel syndrome: Pathogenesis, diagnosis, treatment, and evidence-based medicine. World J. Gastroenterol. 2014, 20, 6759–6773. [Google Scholar] [CrossRef]
- Zhang, Y.-Z.; Li, Y.-Y. Inflammatory bowel disease: Pathogenesis. World J. Gastroenterol. 2014, 7, 91–99. [Google Scholar] [CrossRef]
- Gujral, N.; Freeman, H.J.; Thomson, A.B.R. Celiac disease: Prevalence, diagnosis, pathogenesis and treatment. World J. Gastroenterol. 2012, 18, 6036–6059. [Google Scholar] [CrossRef]
- Barbara, G.; De Giorgio, R.; Stanghellini, V.; Cremon, C.; Corinaldesi, R. A role for inflammation in irritable bowel syndrome? Gut 2002, 51 (Suppl. 1), i41–i44. [Google Scholar] [CrossRef]
- Ng, Q.X.; Soh, A.Y.S.; Loke, W.; Lim, D.Y.; Yeo, W.-S. The role of inflammation in irritable bowel syndrome (IBS). J. Inflamm. Res. 2018, 11, 345–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascual, V.; Dieli-Crimi, R.; López-Palacios, N.; Bodas, A.; Medrano, L.M.; Núñez, C. Inflammatory bowel disease and celiac disease: Overlaps and differences. World J. Gastroenterol. 2014, 20, 4846–4856. [Google Scholar] [CrossRef] [PubMed]
- Petersen, C.; Round, J.L. Defining dysbiosis and its influence on host immunity and disease. Cell. Microbiol. 2014, 16, 1024–1033. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, M.; Lembo, A. Microbiome and Its Role in Irritable Bowel Syndrome. Dig. Dis. Sci. 2020, 65, 829–839. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.-N.; Wu, H.; Chen, Y.-Z.; Chen, Y.-J.; Shen, X.-Z.; Liu, T.-T. Altered molecular signature of intestinal microbiota in irritable bowel syndrome patients compared with healthy controls: A systematic review and meta-analysis. Dig. Liver Dis. 2017, 49, 331–337. [Google Scholar] [CrossRef]
- Kostic, A.D.; Xavier, R.J.; Gevers, D. The Microbiome in Inflammatory Bowel Disease: Current Status and the Future Ahead. Gastroenterology 2014, 146, 1489–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Siles, M.; Duncan, S.H.; Garcia-Gil, L.J.; Martinez-Medina, M. Faecalibacterium prausnitzii: From microbiology to diagnostics and prognostics. ISME J. 2017, 11, 841–852. [Google Scholar] [CrossRef]
- Valitutti, F.; Cucchiara, S.; Fasano, A. Celiac Disease and the Microbiome. Nutrients 2019, 11, 2403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanson, K.S.; Gibson, G.R.; Hutkins, R.; Reimer, R.A.; Reid, G.; Verbeke, K.; Scott, K.P.; Holscher, H.D.; Azad, M.B.; Delzenne, N.M.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of synbiotics. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 687–701. [Google Scholar] [CrossRef] [PubMed]
- Staudacher, H.M.; Whelan, K. Altered gastrointestinal microbiota in irritable bowel syndrome and its modification by diet: Probiotics, prebiotics and the low FODMAP diet. Proc. Nutr. Soc. 2016, 75, 306–318. [Google Scholar] [CrossRef]
- Didari, T.; Mozaffari, S.; Nikfar, S.; Abdollahi, M. Effectiveness of probiotics in irritable bowel syndrome: Updated systematic review with meta-analysis. World J. Gastroenterol. 2015, 21, 3072–3084. [Google Scholar] [CrossRef]
- Moayyedi, P.; Ford, A.C.; Talley, N.J.; Cremonini, F.; Foxx-Orenstein, A.E.; Brandt, L.J.; Quigley, E.M.M. The efficacy of probiotics in the treatment of irritable bowel syndrome: A systematic review. Gut 2008, 59, 325–332. [Google Scholar] [CrossRef]
- Eom, T.; Kim, Y.S.; Choi, C.H.; Sadowsky, M.J.; Unno, T. Current understanding of microbiota and dietary-therapies for treating inflammatory bowel disease. J. Microbiol. 2018, 56, 189–198. [Google Scholar] [CrossRef]
- Ganji-Arjenaki, M.; Rafieian-Kopaei, M. Probiotics are a good choice in remission of inflammatory bowel diseases: A meta analysis and systematic review. J. Cell. Physiol. 2018, 233, 2091–2103. [Google Scholar] [CrossRef]
- Marasco, G.; Cirota, G.G.; Rossini, B.; Lungaro, L.; Di Biase, A.R.; Colecchia, A.; Volta, U.; de Giorgio, R.; Festi, D.; Caio, G. Probiotics, Prebiotics and Other Dietary Supplements for Gut Microbiota Modulation in Celiac Disease Patients. Nutrients 2020, 12, 2674. [Google Scholar] [CrossRef]
- Pryde, S.E.; Duncan, S.H.; Hold, G.L.; Stewart, C.S.; Flint, H.J. The microbiology of butyrate formation in the human colon. FEMS Microbiol Lett. 2002, 217, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Hamer, H.; Jonkers, D.; Venema, K.; Vanhoutvin, S.; Troost, F.J.; Brummer, R. Review article: The role of butyrate on colonic function. Aliment. Pharmacol. Ther. 2007, 27, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Canani, R.B.; Costanzo MDi Leone, L.; Pedata, M.; Meli, R.; Calignano, A. Potential beneficial effects of butyrate in intestinal and extraintestinal diseases. World J. Gastroenterol. 2011, 17, 1519–1528. [Google Scholar] [CrossRef]
- Żółkiewicz, J.; Marzec, A.; Ruszczyński, M.; Feleszko, W. Postbiotics—A step beyond pre- and probiotics. Nutrients 2020, 12, 2189. [Google Scholar] [CrossRef]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, K.P.; Antoine, J.-M.; Midtvedt, T.; Van Hemert, S. Manipulating the gut microbiota to maintain health and treat disease. Microb. Ecol. Health Dis. 2015, 26, 25877. [Google Scholar] [CrossRef]
- De Vuyst, L.; Leroy, F. Cross-feeding between bifidobacteria and butyrate-producing colon bacteria explains bifdobacterial competitiveness, butyrate production, and gas production. Int. J. Food Microbiol. 2011, 149, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Schwab, C.; Ruscheweyh, H.; Bunesova, V.; Pham, V.T.; Schwab, C. Trophic interactions of infant bifidobacteria and Eubacterium hallii during L-fucose and fucosyllactose degradation bacterial strains and culture conditions. Front. Microbiol. 2017, 8, 1–14. [Google Scholar] [CrossRef]
- Castanys-Muñoz, E.; Martin, M.J.; Prieto, P.A. 2′-fucosyllactose: An abundant, genetically determined soluble glycan present in human milk. Nutr. Rev. 2013, 71, 773–789. [Google Scholar] [CrossRef]
- Salminen, S.; Stahl, B.; Vinderola, G.; Szajewska, H. Infant Formula Supplemented with Biotics: Current Knowledge and Future Perspectives. Nutrients 2020, 12, 1952. [Google Scholar] [CrossRef] [PubMed]
- Bode, L. Human milk oligosaccharides: Every baby needs a sugar mama. Glycobiology 2012, 22, 1147–1162. [Google Scholar] [CrossRef] [Green Version]
- Garrido, D.; Barile, D.; Mills, D.A. A Molecular Basis for Bifidobacterial Enrichment in the Infant Gastrointestinal Tract. Adv. Nutr. 2012, 3, 415S–421S. [Google Scholar] [CrossRef]
- Coppa, G.V.; Bruni, S.; Morelli, L.; Soldi, S.; Gabrielli, O. The first prebiotics in humans: Human milk oligosaccharides. J. Clin. Gastroenterol. 2004, 38, 80–83. [Google Scholar] [CrossRef]
- Musilova, S.; Rada, V.; Vlkova, E.; Bunesova, V. Beneficial effects of human milk oligosaccharides on gut microbiota. Benef. Microbes 2014, 5, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Triantis, V.; Bode, L.; Van Neerven, R.J.J. Immunological Effects of Human Milk Oligosaccharides. Front. Pediatr. 2018, 6, 190. [Google Scholar] [CrossRef] [PubMed]
- Nolan, L.S.; Rimer, J.M.; Good, M. The Role of Human Milk Oligosaccharides and Probiotics on the Neonatal Microbiome and Risk of Necrotizing Enterocolitis: A Narrative Review. Nutrients 2020, 12, 3052. [Google Scholar] [CrossRef]
- Zuurveld, M.; Van Witzenburg, N.P.; Garssen, J.; Folkerts, G.; Stahl, B.; Land, B.V.; Willemsen, L.E.M. Immunomodulation by Human Milk Oligosaccharides: The Potential Role in Prevention of Allergic Diseases. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef]
- Reverri, E.J.; Devitt, A.A.; Kajzer, J.A.; Baggs, G.E.; Borschel, M.W. Review of the Clinical Experiences of Feeding Infants Formula Containing the Human Milk Oligosaccharide 2′-Fucosyllactose. Nutrients 2018, 10, 1346. [Google Scholar] [CrossRef] [Green Version]
- Vandenplas, Y.; Berger, B.; Carnielli, V.P.; Ksiazyk, J.; Lagström, H.; Luna, M.S.; Migacheva, N.; Mosselmans, J.-M.; Picaud, J.-C.; Possner, M.; et al. Human Milk Oligosaccharides: 2′-Fucosyllactose (2′-FL) and Lacto-N-Neotetraose (LNnT) in Infant Formula. Nutrients 2018, 10, 1161. [Google Scholar] [CrossRef] [Green Version]
- Elison, E.; Vigsnaes, L.K.; Krogsgaard, L.R.; Rasmussen, J.; Sørensen, N.; McConnell, B.; Hennet, T.; Sommer, M.O.A.; Bytzer, P. Oral supplementation of healthy adults with 2′-O-fucosyllactose and lacto-N-neotetraose is well tolerated and shifts the intestinal microbiota. Br. J. Nutr. 2016, 116, 1356–1368. [Google Scholar] [CrossRef] [Green Version]
- Iribarren, C.; Törnblom, H.; Aziz, I.; Magnusson, M.K.; Sundin, J.; Vigsnæs, L.K.; Amundsen, I.D.; McConnell, B.; Seitzberg, D.; Öhman, L.; et al. Human milk oligosaccharide supplementation in irritable bowel syndrome patients: A parallel, randomized, double-blind, placebo-controlled study. Neurogastroenterol. Motil. 2020, 32, e13920. [Google Scholar] [CrossRef]
- Islam, S.U. Clinical Uses of Probiotics. Medicine 2016, 95, e2658. [Google Scholar] [CrossRef] [PubMed]
- Wallner, G.; Amann, R.; Beisker, W. Optimizing fluorescent in situ hybridization with rRNA-targeted oligonucleotide probes for flow cytometric identification of microorganisms. Cytometry 1993, 14, 136–143. [Google Scholar] [CrossRef]
- Daims, H.; Brühl, A.; Amann, R.; Schleifer, K.-H.; Wagner, M. The Domain-specific Probe EUB338 is Insufficient for the Detection of all Bacteria: Development and Evaluation of a more Comprehensive Probe Set. Syst. Appl. Microbiol. 1999, 22, 434–444. [Google Scholar] [CrossRef]
- Harmsen, H.J.; Wildeboer-Veloo, A.C.; Grijpstra, J.; Knol, J.; Degener, J.E.; Welling, G.W. Development of 16S rRNA-based probes for the Coriobacterium group and the Atopobium cluster and their application for enumeration of Coriobacteriaceae in human feces from volunteers of different age groups. Appl. Environ. Microbiol. 2000, 66, 4523–4527. [Google Scholar] [CrossRef] [Green Version]
- Manz, W.; Amann, R.; Ludwig, W.; Vancanneyt, M.; Schleifer, K.H. Application of a suite of 16S rRNA-specific oligonucleotide probes designed to investigate bacteria of the phylum cytophaga-flavobacter-bacteroides in the natural environment. Microbiology 1996, 142, 1097–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langendijk, P.S.; Schut, F.; Jansen, G.J.; Raangs, G.C.; Kamphuis, G.R.; Wilkinson, M.H.; Welling, G.W. Quantitative fluorescence In Situ hybridization of Bifidobacterium spp. with genus-specific 16S rRNA-targeted probes and its application in fecal samples. Appl. Environ. Microbiol. 1995, 61, 3069–3075. [Google Scholar] [CrossRef] [Green Version]
- Franks, A.H.; Harmsen, H.J.M.; Raangs, G.C.; Jansen, G.J.; Schut, F.; Welling, G.W. Variations of bacterial populations in human feces measured by fluorescent In Situ hybridization with group-specific 16S rRNA-targeted oligonucleotide probes. Appl. Environ. Microbiol. 1998, 64, 3336–3345. [Google Scholar] [CrossRef] [Green Version]
- Walker, A.W.; Duncan, S.H.; Leitch, E.C.M.; Child, M.W.; Flint, H.J. pH and Peptide Supply Can Radically Alter Bacterial Populations and Short-Chain Fatty Acid Ratios within Microbial Communities from the Human Colon. Appl. Environ. Microbiol. 2005, 71, 3692–3700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hold, G.L.; Schwiertz, A.; Aminov, R.I.; Blaut, M.; Flint, H.J. Oligonucleotide Probes That Detect Quantitatively Significant Groups of Butyrate-Producing Bacteria in Human Feces. Appl. Environ. Microbiol. 2003, 69, 4320–4324. [Google Scholar] [CrossRef] [Green Version]
- Devereux, R.; Kane, M.D.; Winfrey, J.; Stahl, D.A. Genus- and Group-Specific Hybridization Probes for Determinative and Environmental Studies of Sulfate-Reducing Bacteria. Syst. Appl. Microbiol. 1992, 15, 601–609. [Google Scholar] [CrossRef]
- Harmsen, H.J.M.; Elfferich, P.; Schut, F.; Welling, G.W. A 16S rRNA-targeted probe for detection of lactobacilli and enterococci in faecal samples by fluorescent In Situ hybridization. Microb. Ecol. Health Dis. 1999, 11, 3–12. [Google Scholar]
- Ferreira-Lazarte, A.; Kachrimanidou, V.; Villamiel, M.; Rastall, R.A.; Moreno, F.J. In Vitro fermentation properties of pectins and enzymatic-modified pectins obtained from different renewable bioresources. Carbohydr. Polym. 2018, 199, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Eypasch, E.; Williams, J.I.; Wood-Dauphinee, S.; Ure, B.M.; Schmulling, C.; Neugebauer, E.; Troidl, H. Gastrointestinal Quality of Life Index: Development, validation and application of a new instrument. Br. J. Nutr. 1995, 82, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Azpiroz, F.; Guyonnet, D.; Donazzolo, Y.; Gendre, D.; Tanguy, J.; Guarner, F. Digestive Symptoms in Healthy People and Subjects With Irritable Bowel Syndrome. J. Clin. Gastroenterol. 2015, 49, e64–e70. [Google Scholar] [CrossRef]
- Guyatt, G.; Mitchell, A.; Irvine, E.J.; Singer, J.; Williams, N.; Goodacre, R.; Tompkins, C. A new measure of health status for clinical trials in inflammatory bowel disease. Gastroenterolohy 1989, 96, 804–810. [Google Scholar] [CrossRef]
- Chen, L.; Reynolds, C.; David, R.; Brewer, A.P. Development of an Index Score for Intestinal Inflammation-Associated Dysbiosis Using Real-World Stool Test Results. Dig. Dis. Sci. 2020, 65, 1111–1124. [Google Scholar] [CrossRef] [Green Version]
- Šuligoj, T.; Vigsnæs, L.K.; Abbeele, P.V.D.; Apostolou, A.; Karalis, K.; Savva, G.M.; McConnell, B.; Juge, N. Effects of Human Milk Oligosaccharides on the Adult Gut Microbiota and Barrier Function. Nutrients 2020, 12, 2808. [Google Scholar] [CrossRef]
- Jayadevappa, R.; Cook, R.; Chhatre, S. Minimal important difference to infer changes in health-related quality of life—A systematic review. J. Clin. Epidemiology 2017, 89, 188–198. [Google Scholar] [CrossRef]
- Coffin, B.; Dapoigny, M.; Cloarec, D.; Comet, D.; Dyard, F. Relationship between severity of symptoms and quality of life in 858 patients with irritable bowel syndrome. Gastroenterol. Clin. Biol. 2004, 28, 11–15. [Google Scholar] [CrossRef]
- Borgaonkar, M.; Irvine, E. Quality of life measurement in gastrointestinal and liver disorders. Gut 2000, 47, 444–454. [Google Scholar] [CrossRef] [Green Version]
- Casellas, L.R.F.; Rodrigo, L.; Vivancos, S.R.P.; Riestra, S.; Pantiga, C.; Baudet, J.S.; Junquera, F.; Diví, V.P.; Abadia, C.; Papo, M.; et al. Factors that impact health-related quality of life in adults with celiac disease: A multicenter study. World J. Gastroenterol. 2008, 14, 46–52. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.-T.; Chen, C.; Kling, D.E.; Liu, B.; McCoy, J.M.; Merighi, M.; Heidtman, M.; Newburg, D.S. The principal fucosylated oligosaccharides of human milk exhibit prebiotic properties on cultured infant microbiota. Glycobiology 2013, 23, 169–177. [Google Scholar] [CrossRef]
- Yu, Z.-T.; Chen, C.; Newburg, D.S. Utilization of major fucosylated and sialylated human milk oligosaccharides by isolated human gut microbes. Glycobiology 2013, 23, 1281–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Moyano, S.; Totten, S.M.; Garrido, D.A.; Smilowitz, J.T.; German, J.B.; Lebrilla, C.B.; Mills, D.A. Variation in Consumption of Human Milk Oligosaccharides by Infant Gut-Associated Strains of Bifidobacterium breve. Appl. Environ. Microbiol. 2013, 79, 6040–6049. [Google Scholar] [CrossRef] [Green Version]
- Bunesova, V.; Lacroix, C.; Schwab, C. Fucosyllactose and L-fucose utilization of infant Bifidobacterium longum and Bifidobacterium kashiwanohense. BMC Microbiol. 2016, 16, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín, R.; Bermúdez-Humarán, L.G.; Langella, P. Searching for the Bacterial Effector: The Example of the Multi-Skilled Commensal Bacterium Faecalibacterium prausnitzii. Front. Microbiol. 2018, 9, 346. [Google Scholar] [CrossRef]
- Rajilić–Stojanović, M.; Biagi, E.; Heilig, H.G.; Kajander, K.; Kekkonen, R.A.; Tims, S.; de Vos, W.M. Global and Deep Molecular Analysis of Microbiota Signatures in Fecal Samples From Patients with Irritable Bowel Syndrome. Gastroenterology 2011, 141, 1792–1801. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Shen, J.; Ran, Z.H. Association between Faecalibacterium prausnitzii Reduction and Inflammatory Bowel Disease: A Meta-Analysis and Systematic Review of the Literature. Gastroenterol. Res. Pr. 2014, 2014, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Sokol, H.; Seksik, P.; Furet, J.P.; Firmesse, O.; Nion-Larmurier, I.; Beaugerie, L.; Cosnes, J.; Corthier, G.; Marteau, P.; Doré, J. Low counts of Faecalibacterium prausnitzii in colitis microbiota. Inflamm. Bowel Dis. 2009, 15, 1183–1189. [Google Scholar] [CrossRef] [PubMed]
- De Palma, G.; Nadal, I.; Medina, M.; Donat, E.; Ribes-koninckx, C.; Calabuig, M.; Sanz, Y. Intestinal dysbiosis and reduced immunoglobulin-coated bacteria associated with coeliac disease in children. BMC Microbiol. 2010, 10, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Verhoog, S.; Taneri, P.E.; Díaz, Z.M.R.; Marques-Vidal, P.; Troup, J.P.; Bally, L.; Franco, O.H.; Glisic, M.; Muka, T. Dietary Factors and Modulation of Bacteria Strains of Akkermansia muciniphila and Faecalibacterium prausnitzii: A Systematic Review. Nutrients 2019, 11, 1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira-Halder, C.V.; Faria, A.V.D.S.; Andrade, S.S. Action and function of Faecalibacterium prausnitzii in health and disease. Best Pr. Res. Clin. Gastroenterol. 2017, 31, 643–648. [Google Scholar] [CrossRef]
- Miquel, S.; Martin, R.; Lashermes, A.; Gillet, M.; Meleine, M.; Gelot, A.; Eschalier, A.; Ardid, D.; Bermúdez-Humarán, L.G.; Sokol, H.; et al. Anti-nociceptive effect of Faecalibacterium prausnitzii in non-inflammatory IBS-like models. Sci. Rep. 2016, 6, 19399. [Google Scholar] [CrossRef] [Green Version]
- Fang, S.; Wu, D.; Yang, Z.; Jiao, N.; Lin, Y.; Li, X.; Xiao, J.; Yuan, M.; Zhu, R.; Zhu, L. Keystone species in the pathogenic process of NAFLD. FASEB 2019, 33, 496.40. [Google Scholar]
- Trosvik, P.; De Muinck, E.J. Ecology of bacteria in the human gastrointestinal tract—Identification of keystone and foundation taxa. Microbiome 2015, 3, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.; Kiewiet, M.B.G.; Logtenberg, M.J.; Groeneveld, A.; Nauta, A.; Schols, H.A.; Walvoort, M.T.C.; Harmsen, H.J.M.; De Vos, P. Effects of Different Human Milk Oligosaccharides on Growth of Bifidobacteria in Monoculture and Co-culture with Faecalibacterium prausnitzii. Front. Microbiol. 2020, 11, 569700. [Google Scholar] [CrossRef]
- Dewulf, E.M.; Cani, P.; Claus, S.P.; Fuentes, S.; Puylaert, P.G.B.; Neyrinck, A.M.; Bindels, L.B.; De Vos, W.M.; Gibson, G.R.; Thissen, J.-P.; et al. Insight into the prebiotic concept: Lessons from an exploratory, double blind intervention study with inulin-type fructans in obese women. Gut 2012, 62, 1112–1121. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-farias, C.; Slezak, K.; Fuller, Z.; Duncan, A.; Holtrop, G.; Louis, P. Effect of inulin on the human gut microbiota: Stimulation of Bifidobacterium adolescentis and Faecalibacterium prausnitzii. Br. J. Nutr. 2009, 101, 541–550. [Google Scholar] [CrossRef] [Green Version]
- Van den Abbeele, P.; Duysburgh, C.; Vazquez, E.; Chow, J.; Buck, R.; Marzorati, M. 2′-Fucosyllactose alters the composition and activity of gut microbiota from formula-fed infants receiving complementary feeding in a validated intestinal model. J. Funct. Foods 2019, 61. [Google Scholar] [CrossRef]
- Li, M.; Bauer, L.L.; Chen, X.; Wang, M.; Kuhlenschmidt, T.B.; Kuhlenschmidt, M.S.; Fahey, G.C.; Donovan, S.M. Microbial Composition and In Vitro Fermentation Patterns of Human Milk Oligosaccharides and Prebiotics Differ between Formula-Fed and Sow-Reared Piglets. J. Nutr. 2012, 142, 681–689. [Google Scholar] [CrossRef] [Green Version]
- Azagra-Boronat, I.; Massot-Cladera, M.; Mayneris-Perxachs, J.; Knipping, K.; Land, B.V.; Tims, S.; Stahl, B.; Garssen, J.; Franch, À.; Castell, M.; et al. Immunomodulatory and Prebiotic Effects of 2′-Fucosyllactose in Suckling Rats. Front. Immunol. 2019, 10, 1773. [Google Scholar] [CrossRef] [Green Version]
- Kumari, R.; Ahuja, V.; Paul, J. Fluctuations in butyrate-producing bacteria in ulcerative colitis patients of North India. World J. Gastroenterol. WJG 2013, 19, 3404–3414. [Google Scholar] [CrossRef] [PubMed]
- Polansky, O.; Sekelova, Z.; Faldynova, M.; Sebkova, A.; Sisak, F.; Rychlik, I. Important Metabolic Pathways and Biological Processes Expressed by Chicken Cecal Microbiota. Appl. Environ. Microbiol. 2016, 82, 1569–1576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, P.A.; Song, Y.; Liu, C.; Molitoris, D.R.; Vaisanen, M.-L.; Collins, M.D.; Finegold, S.M. Anaerotruncus colihominis gen. nov., sp. nov., from human faeces. Int. J. Syst. Evol. Microbiol. 2004, 54, 413–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, P.; Flint, H.J. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol. Lett. 2009, 294, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Vergara, D.M.; Gonz, E. Short chain fatty acids (butyric acid) and intestinal diseases. Nutr. Hosp. 2017, 34, 58–61. [Google Scholar]
- Borycka-Kiciak, K.; Banasiewicz, T.; Rydzewska, G. Butyric acid—A well-known molecule revisited. Gastroenterol. Rev. 2017, 2, 83–89. [Google Scholar] [CrossRef]
- Załęski, A.; Banaszkiewicz, A.; Walkowiak, J. Butyric acid in irritable bowel syndrome. Gastroenterol. Rev. 2013, 8, 350–353. [Google Scholar] [CrossRef] [Green Version]
- Soldavini, J.; Kaunitz, J.D. Pathobiology and potential therapeutic value of intestinal short-chain fatty acids in gut inflammation and obesity. Dig. Dis. Sci. 2013, 58, 2756–2766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín, R.; Miquel, S.; Benevides, L.; Bridonneau, C.; Robert, V.; Hudault, S.; Chain, F.; Berteau, O.; Azevedo, V.; Chatel, J.M.; et al. Functional Characterization of Novel Faecalibacterium prausnitzii Strains Isolated from Healthy Volunteers: A Step Forward in the Use of F. prausnitzii as a Next-Generation Probiotic. Front. Microbiol. 2017, 8, 1226. [Google Scholar] [CrossRef] [PubMed]
- Forbes, A.; Escher, J.; Hébuterne, X.; Kłęk, S.; Krznaric, Z.; Schneider, S.; Shamir, R.; Stardelova, K.; Wierdsma, N.; Wiskin, A.E.; et al. ESPEN guideline: Clinical nutrition in inflammatory bowel disease. Clin. Nutr. 2017, 36, 321–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullin, G.E. Micronutrients and Inflammatory Bowel Disease. Nutr. Clin. Pract. 2012, 27, 136–137. [Google Scholar] [CrossRef]


| Synbiotic 1 (S1) | 2′-FL + Lactobacillus acidophilus NCFM (ATCC 5221) + Bifidobacterium animalis subsp. lactis Bi-07 (ATCC 5220) |
| Synbiotic 2 (S2) | 2′-FL + L. acidophilus NCFM + B. animalis subsp. lactis Bi-07 + Bifidobacterium animalis subsp. lactis Bl-04 (ATCC 5219) + Lactobacillus paracasei Lpc-37 (ATCC 5275) |
| Synbiotic 3 (S3) | 2′-FL + L. acidophilus NCFM + B. animalis subsp. lactis Bi-07 + Lactobacillus rhamnosus GG (ATCC 7017) |
| Synbiotic 4 (S4) | 2′-FL + L. rhamnosus GG |
| Synbiotic 5 (S5) | 2′-FL + Lactobacillus salivarius UCC118 |
| Mean ± SD or n (%) | |
|---|---|
| Age (years) | 31.4 ± 10.5 |
| Sex | |
| Male | 7 (58%) |
| Female | 5 (42%) |
| Body Mass Index (kg/m2) | 23.8 ± 3.4 |
| Race (Ethnicity) | |
| White (not Hispanic/Latino) | 8 (66.7%) |
| White/Native American (not Hispanic/Latino) | 2 (16.7%) |
| White (Hispanic/Latino) | 1 (8.3%) |
| Black (not Hispanic/Latino) | 1 (8.3%) |
| Gastrointestinal Condition | |
| Irritable bowel syndrome (IBS) | 7 (58.3%) |
| Ulcerative colitis (UC) | 4 (33.3%) |
| Celiac disease | 1 (8.3%) |
| Score Range | Baseline | Week 6 | Δ Over 6 Weeks | Mean % Δ | p1 | Cohen’s D | |||
|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | ||||||
| GIQLI Total Score | 0–144 | 94.3 | 25.5 | 109.4 | 19.2 | 15.1 | 20.8% | 0.020 | 0.59 |
| GIQLI-Gastrointestinal Symptoms | 0–76 | 53.3 | 10.3 | 61.4 | 7.7 | 8.1 | 18.1% | 0.022 | 0.79 |
| GIQLI-Physical Function | 0–28 | 15.6 | 7.4 | 17.8 | 6.1 | 2.2 | 36.5% | 0.164 | 0.30 |
| GIQLI-Social Function | 0–16 | 10.7 | 3.8 | 12.3 | 3.7 | 1.6 | 18.4% | 0.004 | 0.42 |
| GIQLI-Emotional Function | 0–20 | 12.0 | 5.8 | 14.7 | 4.5 | 2.7 | 46.5% | 0.139 | 0.47 |
| IBDQ Total Score | 32–224 | 134.0 | 52.4 | 179.0 | 36.1 | 45.0 | 43.6% | 0.078 | 0.86 |
| IBDQ-Bowel Symptoms | 10–70 | 43.0 | 19.4 | 56.3 | 11.7 | 13.3 | 44.3% | 0.165 | 0.69 |
| IBDQ-Systemic Systems | 5–35 | 16.8 | 6.2 | 27.0 | 6.3 | 10.2 | 66.9% | 0.000 | 1.65 |
| IBDQ-Social Function | 5–35 | 22.0 | 9.9 | 27.8 | 7.5 | 5.8 | 41.5% | 0.239 | 0.59 |
| IBDQ-Emotion Health | 12–84 | 51.5 | 18.7 | 68.0 | 11.7 | 16.5 | 43.3% | 0.098 | 0.88 |
| DSFQ Total Score | 0–16 | 9.0 | 3.8 | 8.4 | 5.0 | −0.6 | −10.3% | 0.522 | −0.16 |
| Phylum | Microbiota (CFU/g Stool) | Log Transformed Data | ||||||
|---|---|---|---|---|---|---|---|---|
| Baseline | Week 6 | Geometric Mean % Δ | p1 | Cohen’s D | ||||
| Mean | SD | Mean | SD | |||||
| Actinobacteria | Bifidobacterium spp. | 20.1 | 1.6 | 21.6 | 1.7 | 356.3% | * 0.026 | 0.95 |
| Bifidobacterium longum | 17.4 | 2.0 | 19.4 | 1.6 | 650.0% | * 0.003 | 0.99 | |
| Collinsella aerofaciens | 17.6 | 3.0 | 18.1 | 3.5 | 65.7% | 0.392 | 0.17 | |
| Bacteroidetes | Bacteroides-Prevotella group | 20.2 | 0.9 | 21.2 | 1.3 | 169.0% | 0.101 | 1.09 |
| Bacteroides vulgatus | 19.8 | 2.5 | 20.9 | 2.6 | 190.4% | 0.081 | 0.43 | |
| Barnesiella spp. | 16.0 | 2.0 | 15.4 | 2.9 | −46.1% | 0.468 | −0.31 | |
| Odoribacter spp. | 15.6 | 2.5 | 14.8 | 3.6 | −54.9% | 0.369 | −0.32 | |
| Prevotella spp. | 14.1 | 2.0 | 14.9 | 1.8 | 116.2% | 0.301 | 0.39 | |
| Euryarchaeota | Methanobrevibacter smithii | 13.1 | 2.0 | 14.0 | 2.1 | 134.7% | 0.290 | 0.42 |
| Firmicutes | Anaerotruncus colihominis | 14.5 | 1.6 | 16.1 | 1.1 | 402.6% | * 0.018 | 1.00 |
| Butyrivibrio crossotus | 9.9 | 1.5 | 10.7 | 3.1 | 111.7% | 0.422 | 0.51 | |
| Clostridium spp. | 21.0 | 1.6 | 21.9 | 1.9 | 156.9% | 0.125 | 0.58 | |
| Coprococcus eutactus | 14.1 | 2.3 | 14.1 | 2.7 | −2.4% | 0.983 | −0.01 | |
| Faecalibacterium prausnitzii | 20.6 | 2.2 | 22.1 | 1.8 | 353.7% | * 0.029 | 0.67 | |
| Lactobacillus spp. | 19.5 | 2.5 | 20.5 | 2.4 | 160.7% | 0.252 | 0.39 | |
| Pseudoflavonifractor spp. | 18.0 | 1.5 | 19.2 | 1.2 | 226.4% | * 0.016 | 0.80 | |
| Roseburia spp. | 20.7 | 1.8 | 21.9 | 1.5 | 252.8% | 0.062 | 0.68 | |
| Ruminococcus spp. | 17.7 | 2.0 | 18.4 | 2.1 | 97.5% | 0.273 | 0.34 | |
| Veillonella spp. | 16.1 | 3.0 | 16.9 | 1.7 | 118.8% | 0.331 | 0.26 | |
| Fusobacteria | Fusobacterium spp. | 9.6 | 2.3 | 10.6 | 2.0 | 171.8% | 0.111 | 0.44 |
| Proteobacteria | Desulfovibrio piger | 10.3 | 2.1 | 10.5 | 2.0 | 12.2% | 0.877 | 0.06 |
| Escherichia coli | 16.5 | 3.0 | 17.0 | 1.7 | 67.9% | 0.607 | 0.17 | |
| Oxalobacter formigenes | 14.1 | 1.7 | 13.9 | 2.5 | −16.9% | 0.833 | −0.11 | |
| Verrucomicrobia | Akkermansia muciniphila | 12.6 | 2.0 | 12.9 | 2.0 | 33.6% | 0.702 | 0.15 |
| SCFA (Micromol/g Stool) | Baseline | Week 6 | Mean % Δ | p1 | Cohen’s D | ||
|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | ||||
| Total SCFA | 46.30 | 13.30 | 76.43 | 37.04 | 72.2% | 0.026 | 2.27 |
| Butyrate | 8.12 | 4.76 | 16.71 | 9.63 | 594.0% | 0.040 | 1.80 |
| Acetate | 28.24 | 7.28 | 45.65 | 23.90 | 64.3% | 0.035 | 2.39 |
| Propionate | 10.03 | 4.71 | 14.06 | 8.39 | 42.4% | 0.074 | 0.85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryan, J.J.; Monteagudo-Mera, A.; Contractor, N.; Gibson, G.R. Impact of 2′-Fucosyllactose on Gut Microbiota Composition in Adults with Chronic Gastrointestinal Conditions: Batch Culture Fermentation Model and Pilot Clinical Trial Findings. Nutrients 2021, 13, 938. https://doi.org/10.3390/nu13030938
Ryan JJ, Monteagudo-Mera A, Contractor N, Gibson GR. Impact of 2′-Fucosyllactose on Gut Microbiota Composition in Adults with Chronic Gastrointestinal Conditions: Batch Culture Fermentation Model and Pilot Clinical Trial Findings. Nutrients. 2021; 13(3):938. https://doi.org/10.3390/nu13030938
Chicago/Turabian StyleRyan, Jennifer Joan, Andrea Monteagudo-Mera, Nikhat Contractor, and Glenn R. Gibson. 2021. "Impact of 2′-Fucosyllactose on Gut Microbiota Composition in Adults with Chronic Gastrointestinal Conditions: Batch Culture Fermentation Model and Pilot Clinical Trial Findings" Nutrients 13, no. 3: 938. https://doi.org/10.3390/nu13030938
APA StyleRyan, J. J., Monteagudo-Mera, A., Contractor, N., & Gibson, G. R. (2021). Impact of 2′-Fucosyllactose on Gut Microbiota Composition in Adults with Chronic Gastrointestinal Conditions: Batch Culture Fermentation Model and Pilot Clinical Trial Findings. Nutrients, 13(3), 938. https://doi.org/10.3390/nu13030938