
Phytate and Butyrate Differently Influence the Proliferation, Apoptosis and Survival Pathways in Human Cancer and Healthy Colonocytes

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Phytate and Butyrate
2.2. Characteristics of Phytate Preparation
2.3. Cell Lines and Growth Conditions
2.4. Experimental Design
2.5. Proliferation Test
2.6. Apoptosis and Cell Cycle Determination with Flow Cytometry
2.7. RNA Isolation and Reverse Transcription
2.8. Gene Expression Analysis
2.9. Protein Extraction and Immunoblotting
2.10. Statistical Analysis
3. Results
3.1. The Composition of Phytate Preparation
3.2. Cell Proliferation
3.3. Cell Cycle Analysis and Apoptosis
3.4. Analysis of Gene and Protein Expression
3.4.1. Basal mRNA Expression in Untreated Cells—Comparison of Cell Lines
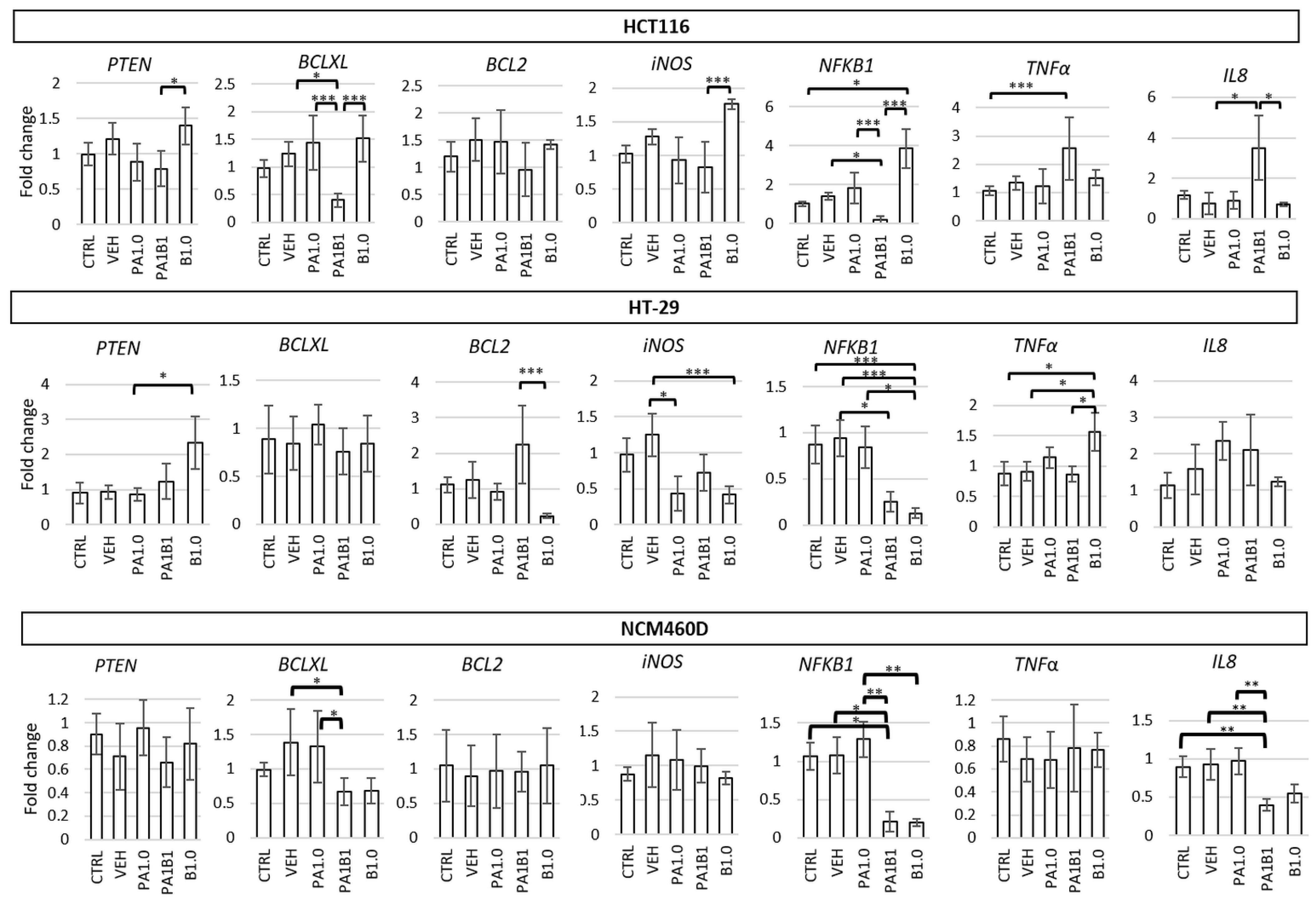
3.4.2. Effect of Phytate and/or Butyrate on Gene and Protein Expression in Treated Colonocytes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zmora, N.; Suez, J.; Elinav, E. You Are What You Eat: Diet, Health and the Gut Microbiota. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 35–56. [Google Scholar] [CrossRef]
- Wilson, A.S.; Koller, K.R.; Ramaboli, M.C.; Nesengani, L.T.; Ocvirk, S.; Chen, C.; Flanagan, C.A.; Sapp, F.R.; Merritt, Z.T.; Bhatti, F.; et al. Diet and the Human Gut Microbiome: An International Review. Dig. Dis. Sci. 2020, 65, 723–740. [Google Scholar] [CrossRef] [Green Version]
- Makki, K.; Deehan, E.C.; Walter, J.; Bäckhed, F. The Impact of Dietary Fiber on Gut Microbiota in Host Health and Disease. Cell Host Microbe 2018, 23, 705–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canani, R.B.; Di Costanzo, M.; Leone, L.; Pedata, M.; Meli, R.; Calignano, A. Potential Beneficial Effects of Butyrate in Intestinal and Extraintestinal Diseases. World J. Gastroenterol. 2011, 17, 1519–1528. [Google Scholar] [CrossRef]
- Liu, H.; Wang, J.; He, T.; Becker, S.; Zhang, G.; Li, D.; Ma, X. Butyrate: A Double-Edged Sword for Health? Adv. Nutr. 2018, 9, 21–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donohoe, D.R.; Curry, K.P.; Bultman, S.J. Microbial Oncotarget: Bacterial-Produced Butyrate, Chemo-Prevention and Warburg Effect Commentary on: The Warburg Effect Dictates the Mechanism of Butyrate-Mediated Histone Acetylation and Cell Proliferation. Oncotarget 2013, 4, 182–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bordonaro, M.; Lazarova, D.L.; Augenlicht, L.H.; Sartorelli, A.C. Cell Type-And Promoter-Dependent Modulation of the Wnt Signaling Pathway by Sodium Butyrate. Int. J. Cancer 2002, 97, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Lazarova, D.L.; Bordonaro, M.; Carbone, R.; Sartorelli, A.C. Linear Relationship between Wnt Activity Levels and Apoptosis in Colorectal Carcinoma Cells Exposed to Butyrate. Int. J. Cancer 2004, 110, 523–531. [Google Scholar] [CrossRef]
- Andriamihaja, M.; Chaumontet, C.; Tome, D.; Blachier, F. Butyrate Metabolism in Human Colon Carcinoma Cells: Implications Concerning Its Growth-Inhibitory Effect. J. Cell. Physiol. 2009, 218, 58–65. [Google Scholar] [CrossRef]
- Donohoe, D.R.; Collins, L.B.; Wali, A.; Bigler, R.; Sun, W.; Bultman, S.J. The Warburg Effect Dictates the Mechanism of Butyrate-Mediated Histone Acetylation and Cell Proliferation. Mol. Cell 2012, 48, 612–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlemmer, U.; Frølich, W.; Prieto, R.M.; Grases, F. Phytate in Foods and Significance for Humans: Food Sources, Intake, Processing, Bioavailability, Protective Role and Analysis. Mol. Nutr. Food Res. 2009, 53, S330–S375. [Google Scholar] [CrossRef]
- Markiewicz, L.H.; Honke, J.; Haros, M.; Światecka, D.; Wróblewska, B. Diet Shapes the Ability of Human Intestinal Microbiota to Degrade Phytate—In vitro Studies. J. Appl. Microbiol. 2013, 115, 247–259. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Woodhouse, L.R.; King, J.C.; Welch, R.M.; Li, S.J.; Paik, H.Y.; Joung, H. Relationships between Faecal Phytate and Mineral Excretion Depend on Dietary Phytate and Age. Br. J. Nutr. 2009, 102, 835–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferry, S.; Matsuda, M.; Yoshida, H.; Hirata, M. Inositol Hexakisphosphate Blocks Tumor Cell Growth by Activating Apoptotic Machinery as Well as by Inhibiting the Akt/NFκB-Mediated Cell Survival Pathway. Carcinogenesis 2002, 23, 2031–2041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapral, M.; Wawszczyk, J.; Jesse, K.; Paul-Samojedny, M.; Kúsmierz, D.; Wȩglarz, L. Inositol Hexaphosphate Inhibits Proliferation and Induces Apoptosis of Colon Cancer Cells by Suppressing the AKT/MTOR Signaling Pathway. Molecules 2017, 22, 1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinicola, S.; Minini, M.; Unfer, V.; Verna, R.; Cucina, A.; Bizzarri, M. Nutritional and Acquired Deficiencies in Inositol Bioavailability. Correlations with Metabolic Disorders. Int. J. Mol. Sci. 2017, 18, 2187. [Google Scholar] [CrossRef] [PubMed]
- Bizzarri, M.; Dinicola, S.; Bevilacqua, A.; Cucina, A. Broad Spectrum Anticancer Activity of Myo-Inositol and Inositol Hexakisphosphate. Int. J. Endocrinol. 2016, 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nissar, J.; Ahad, T.; Naik, H.R.; Hussain, S.Z. A Review Phytic Acid: As Antinutrient or Nutraceutical. J. Pharmacogn. Phytochem. 2017, 6, 1554–1560. [Google Scholar]
- Sakamoto, K.; Venkatraman, G.; Shamsuddin, A.M. Growth Inhibition and Differentiation of HT-29 Cells in Vitro by Inositol Hexaphosphate (Phytic Acid). Carcinogenesis 1993, 14, 1815–1819. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Yang, F.; Liu, C.; Cui, L.; Fu, M.; Song, Y. Inositol Hexaphosphate Hydrolysate Competitively Binds to AKT to Inhibit the Proliferation of Colon Carcinoma. Oncol. Rep. 2017, 38, 2901–2910. [Google Scholar] [CrossRef] [Green Version]
- Keum, N.N.; Giovannucci, E. Global Burden of Colorectal Cancer: Emerging Trends, Risk Factors and Prevention Strategies. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 713–732. [Google Scholar] [CrossRef]
- Wu, X.; Wu, Y.; He, L.; Wu, L.; Wang, X.; Liu, Z. Effects of the Intestinal Microbial Metabolite Butyrate on the Development of Colorectal Cancer. J. Cancer 2018, 9, 2510–2517. [Google Scholar] [CrossRef] [PubMed]
- Ericson, K.; Gan, C.; Cheong, I.; Rago, C.; Samuels, Y.; Velculescu, V.E.; Kinzler, K.W.; Huso, D.L.; Vogelstein, B.; Papadopoulos, N. Genetic Inactivation of AKT1, AKT2, and PDPK1 in Human Colorectal Cancer Cells Clarifies Their Roles in Tumor Growth Regulation. Proc. Natl. Acad. Sci. USA 2010, 107, 2598–2603. [Google Scholar] [CrossRef] [Green Version]
- Rieger, A.M.; Nelson, K.L.; Konowalchuk, J.D.; Barreda, D.R. Modified Annexin V/Propidium Iodide Apoptosis Assay for Accurate Assessment of Cell Death. J. Vis. Exp. 2011. [Google Scholar] [CrossRef] [PubMed]
- Boersma, H.H.; Kietselaer, B.L.J.H.; Stolk, L.M.L.; Bennaghmouch, A.; Hofstra, L.; Narula, J.; Heidendal, G.A.K.; Reutelingsperger, C.P.M. Past, Present, and Future of Annexin A5: From Protein Discovery to Clinical Applications. J. Nucl. Med. 2005, 46, 2035–2050. [Google Scholar]
- Laparra, J.M.; Alegría, A.; Barberá, R.; Farré, R. Antioxidant Effect of Casein Phosphopeptides Compared with Fruit Beverages Supplemented with Skimmed Milk against H2O2-Induced Oxidative Stress in Caco-2 Cells. Food Res. Int. 2008, 41, 773–779. [Google Scholar] [CrossRef]
- Fotschki, J.; Szyc, A.M.; Laparra, J.M.; Wróblewska, B. Mare’s and Cow’s Milk: Promote Similar Metabolic Effects and Expression of Innate Markers in Caco-2 Cells? Food Res. Int. 2015, 72, 184–190. [Google Scholar] [CrossRef]
- Markiewicz, L.H.; Szymkiewicz, A.; Szyc, A.; Wróblewska, B. A Simultaneous Two-Colour Detection Method of Human IgG- and IgE-Reactive Proteins from Lactic Acid Bacteria. J. Microbiol. Methods 2016, 126, 72–75. [Google Scholar] [CrossRef]
- Swiatecka, D.; Markiewicz, L.H.; Wroblewska, B. Pea Protein Hydrolysate as a Factor Modulating the Adhesion of Bacteria to Enterocytes, Epithelial Proliferation and Cytokine Secretion—An in vitro Study. Cent. Eur. J. Immunol. 2012, 37, 227–231. [Google Scholar] [CrossRef] [Green Version]
- Fiedorowicz, E.; Markiewicz, L.H.; Sidor, K.; Świątecka, D.; Cieślińska, A.; Matysiewicz, M.; Piskorz-Ogórek, K.; Sienkiewicz-Szłapka, E.; Teodorowicz, M.; Świątecki, A.; et al. The Influence of Breast Milk and Infant Formulae Hydrolysates on Bacterial Adhesion and Caco-2 Cells Functioning. Food Res. Int. 2016, 89, 679–688. [Google Scholar] [CrossRef]
- Goretsky, T.; Bradford, E.M.; Ye, Q.; Lamping, O.F.; Vanagunas, T.; Moyer, M.P.; Keller, P.C.; Sinh, P.; Llovet, J.M.; Gao, T.; et al. Beta-Catenin Cleavage Enhances Transcriptional Activation. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Schröterova, L.; Ježkova, A.; Rudolf, E.; Caltova, K.; Králova, V.; Hanušova, V. Inositol Hexaphosphate Limits the Migration and the Invasiveness of Colorectal Carcinoma Cells in Vitro. Int. J. Oncol. 2018, 53, 1625–1632. [Google Scholar] [CrossRef]
- Hope, C.; Planutis, K.; Planutiene, M.; Moyer, M.P.; Johal, K.S.; Woo, J.; Santoso, C.; Hanson, J.A.; Holcombe, R.F. Low Concentrations of Resveratrol Inhibit Wnt Signal Throughput in Colon-Derived Cells: Implications for Colon Cancer Prevention. Mol. Nutr. Food Res. 2008, 52, S52–S61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, S.X.; Shen, J.; Cheng, A.S.L.; Lu, L.; Chan, R.L.Y.; Li, Z.J.; Wang, X.J.; Wong, C.C.M.; Zhang, L.; Ng, S.S.M.; et al. FK-16 Derived from the Anticancer Peptide LL-37 Induces Caspase-Independent Apoptosis and Autophagic Cell Death in Colon Cancer Cells. PLoS ONE 2013, 8, e0063641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, Y.F.; Zou, Z.Z.; Chen, P.J.; Huang, G.B.; Li, B.; Zheng, D.Q.; Yu, X.R.; Luo, X.Y. Combination of Gefitinib and DNA Methylation Inhibitor Decitabine Exerts Synergistic Anti-Cancer Activity in Colon Cancer Cells. PLoS ONE 2014, 9, e0097719. [Google Scholar] [CrossRef]
- Zhang, J.; Yi, M.; Zha, L.; Chen, S.; Li, Z.; Li, C.; Gong, M.; Deng, H.; Chu, X.; Chen, J.; et al. Sodium Butyrate Induces Endoplasmic Reticulum Stress and Autophagy in Colorectal Cells: Implications for Apoptosis. PLoS ONE 2016, 11, e0147218. [Google Scholar] [CrossRef]
- Zeng, H.; Taussig, D.P.; Cheng, W.H.; Johnson, L.A.K.; Hakkak, R. Butyrate Inhibits Cancerous HCT116 Colon Cell Proliferation but to a Lesser Extent in Noncancerous NCM460 Colon Cells. Nutrients 2017, 9, 25. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Song, Y.; Cui, L.; Wen, Z.; Lu, X. Inositol Hexaphosphate Suppresses Growth and Induces Apoptosis in HT-29 Colorectal Cancer Cells in Culture: PI3K/Akt Pathway as a Potential Target. Int. J. Clin. Exp. Pathol. 2015, 8, 1402–1410. [Google Scholar]
- Ellerhorst, J.A.; Ekmekcioglu, S.; Johnson, M.K.; Cooke, C.P.; Johnson, M.M.; Grimm, E.A. Regulation of INOS by the P44/42 Mitogen-Activated Protein Kinase Pathway in Human Melanoma. Oncogene 2006, 25, 3956–3962. [Google Scholar] [CrossRef] [Green Version]
- El-Sherbiny, Y.M.; Cox, M.C.; Ismail, Z.A.; Shamsuddin, A.M.; Vucenik, I. G0/G1 Arrest and S Phase Inhibition of Human Cancer Cell Lines by Inositol Hexaphosphate (IP6). Anticancer Res. 2001, 21, 2393–2404. [Google Scholar]
- Shen, A.; Liu, L.; Chen, H.; Qi, F.; Huang, Y.; Lin, J.; Sferra, T.J.; Sankararaman, S.; Wei, L.; Chu, J.; et al. Cell Division Cycle Associated 5 Promotes Colorectal Cancer Progression by Activating the ERK Signaling Pathway. Oncogenesis 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Takeda, H.; Kiyokawa, E. Activation of Erk in Ileal Epithelial Cells Engaged in Ischemic-Injury Repair. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Zeng, H.; Trujillo, O.N.; Moyer, M.P.; Botnen, J.H. Prolonged Sulforaphane Treatment Activates Survival Signaling in Nontumorigenic NCM460 Colon Cells but Apoptotic Signaling in Tumorigenic HCT116 Colon Cells. Nutr. Cancer 2011, 63, 248–255. [Google Scholar] [CrossRef]
- Li, J.; Huang, L.; Zhao, H.; Yan, Y.; Lu, J. The Role of Interleukins in Colorectal Cancer. Int. J. Biol. Sci. 2020, 16, 2323–2339. [Google Scholar] [CrossRef] [PubMed]
- Cotton, J.A.; Platnich, J.M.; Muruve, D.A.; Jijon, H.; Buret, A.G.; Beck, P.L. Interleukin-8 in Gastrointestinal Inflammation and Malignancy: Induction and Clinical Consequences. Int. J. Interferon Cytokine Mediat. Res. 2016, 8, 13–34. [Google Scholar] [CrossRef] [Green Version]
- Ruder, B.; Atreya, R.; Becker, C. Tumour Necrosis Factor Alpha in Intestinal Homeostasis and Gut Related Diseases. Int. J. Mol. Sci. 2019, 20, 1887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.R.; Choi, H.G.; Jeon, C.K.; Lim, S.J.; Kim, S.H. Butyrate-Mediated Acquisition of Chemoresistance by Human Colon Cancer Cells. Oncol. Rep. 2016, 36, 1119–1126. [Google Scholar] [CrossRef] [Green Version]
- Bordonaro, M.; Tewari, S.; Cicco, C.E.; Atamna, W.; Lazarova, D.L. A Switch from Canonical to Noncanonical Wnt Signaling Mediates Drug Resistance in Colon Cancer Cells. PLoS ONE 2011, 6, e0027308. [Google Scholar] [CrossRef]
- Dabkeviciene, D.; Jonusiene, V.; Zitkute, V.; Zalyte, E.; Grigaitis, P.; Kirveliene, V.; Sasnauskiene, A. The Role of Interleukin-8 (CXCL8) and CXCR2 in Acquired Chemoresistance of Human Colorectal Carcinoma Cells HCT116. Med. Oncol. 2015, 32. [Google Scholar] [CrossRef]
- Bahrami, F.; Kukulski, F.; Lecka, J.; Tremblay, A.; Pelletier, J.; Rockenbach, L.; Sévigny, J. Purine-Metabolizing Ectoenzymes Control IL-8 Production in Human Colon HT-29 Cells. Mediat. Inflamm. 2014, 2014. [Google Scholar] [CrossRef]
- Murakami, A. Chemoprevention with Phytochemicals Targeting Inducible Nitric Oxide Synthase. Forum Nutr. 2009, 61, 193–203. [Google Scholar] [CrossRef]
- Fujiki, H.; Sueoka, E.; Watanabe, T.; Suganuma, M. Synergistic Enhancement of Anticancer Effects on Numerous Human Cancer Cell Lines Treated with the Combination of EGCG, Other Green Tea Catechins, and Anticancer Compounds. J. Cancer Res. Clin. Oncol. 2015, 141, 1511–1522. [Google Scholar] [CrossRef] [PubMed]
- Drago, E.; Bordonaro, M.; Lee, S.; Atamna, W.; Lazarova, D.L. Propolis Augments Apoptosis Induced by Butyrate via Targeting Cell Survival Pathways. PLoS ONE 2013, 8, e0073151. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inositol Phosphates 1 | Content | |
|---|---|---|
| µg/mL | % | |
| 1-IP1 | 1.272 | 0.018 |
| 1,2-IP2 | 11.75 | 0.168 |
| 4,5-IP2 | 11.75 | 0.168 |
| 1,4,5-IP3 | 5.380 | 0.077 |
| 1,2,6-IP3 | 1.871 | 0.027 |
| 1,5,6-IP3 | 23.70 | 0.338 |
| 1,2,5,6-IP4 | 50.20 | 0.716 |
| 1,2,3,6-IP4 | 29.35 | 0.418 |
| 1,2,3,5,6-IP5 | 2525 | 36.01 |
| IP6 | 4352 | 62.06 |
| Total | 7012 | 100 |
| Tested Factor | Cellular Processes 1 | Cancer Colonocytes | Healthy Colonocytes | |
|---|---|---|---|---|
| HCT116 | HT-29 | NCM460D | ||
| 1 mM phytate | Proliferation | ↓ | ↔ | ↔ |
| Apoptosis/cell death | ↑/↑ | ↔/↔ | ↔/↑ | |
| Inflammation | ↔ | ↔ | ↓ | |
| Survival | ↔ | ↓↓ | ↑ | |
| 1 mM butyrate | Proliferation | ↔ | ↔ | ↑↑ |
| Apoptosis/cell death | ↑↑↑/↑↑ | ↑/↑ | ↑/↑ | |
| Inflammation | ↑ | ↔ | ↓ | |
| Survival | ↔ | ↓↓ | ↔ | |
| 1 mM phytate + 1 mM butyrate | Proliferation | ↓↓ | ↔ | ↔ |
| Apoptosis/cell death | ↑↑/↑↑↑ | ↑↑↑/↑ | ↑↑/↑↑ | |
| Inflammation | ↑↑ | ↔ | ↓ | |
| Survival | ↔ | ↓↓ | ↑ | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Markiewicz, L.H.; Ogrodowczyk, A.M.; Wiczkowski, W.; Wróblewska, B. Phytate and Butyrate Differently Influence the Proliferation, Apoptosis and Survival Pathways in Human Cancer and Healthy Colonocytes. Nutrients 2021, 13, 1887. https://doi.org/10.3390/nu13061887
Markiewicz LH, Ogrodowczyk AM, Wiczkowski W, Wróblewska B. Phytate and Butyrate Differently Influence the Proliferation, Apoptosis and Survival Pathways in Human Cancer and Healthy Colonocytes. Nutrients. 2021; 13(6):1887. https://doi.org/10.3390/nu13061887
Chicago/Turabian StyleMarkiewicz, Lidia Hanna, Anna Maria Ogrodowczyk, Wiesław Wiczkowski, and Barbara Wróblewska. 2021. "Phytate and Butyrate Differently Influence the Proliferation, Apoptosis and Survival Pathways in Human Cancer and Healthy Colonocytes" Nutrients 13, no. 6: 1887. https://doi.org/10.3390/nu13061887
APA StyleMarkiewicz, L. H., Ogrodowczyk, A. M., Wiczkowski, W., & Wróblewska, B. (2021). Phytate and Butyrate Differently Influence the Proliferation, Apoptosis and Survival Pathways in Human Cancer and Healthy Colonocytes. Nutrients, 13(6), 1887. https://doi.org/10.3390/nu13061887