
The Pro-Health Benefits of Morusin Administration—An Update Review



Abstract
:1. Introduction
2. Sources and Metabolism of Morusin
2.1. Morusin Identified in Plants
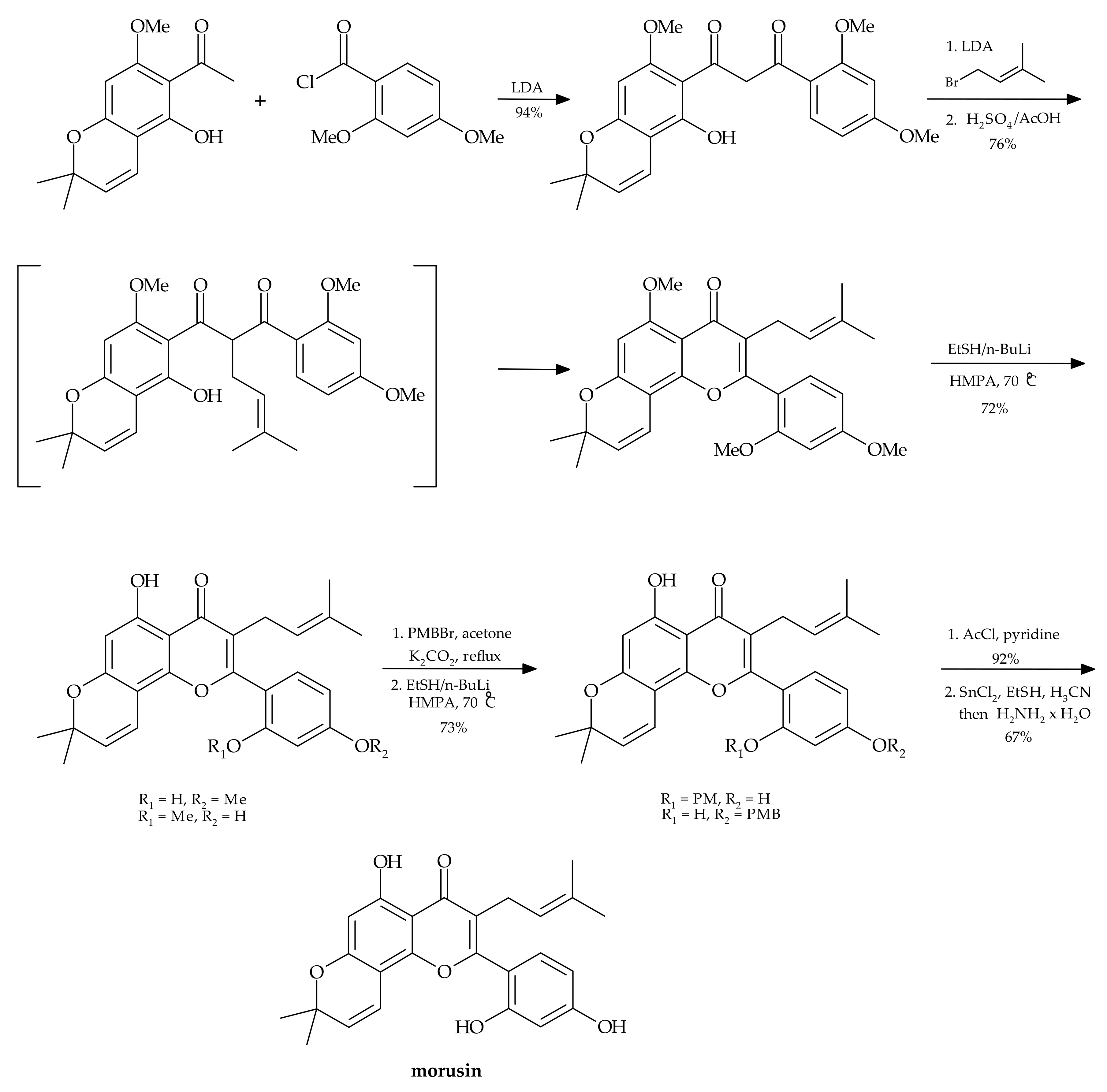
2.2. Chemical Synthesis of Morusin
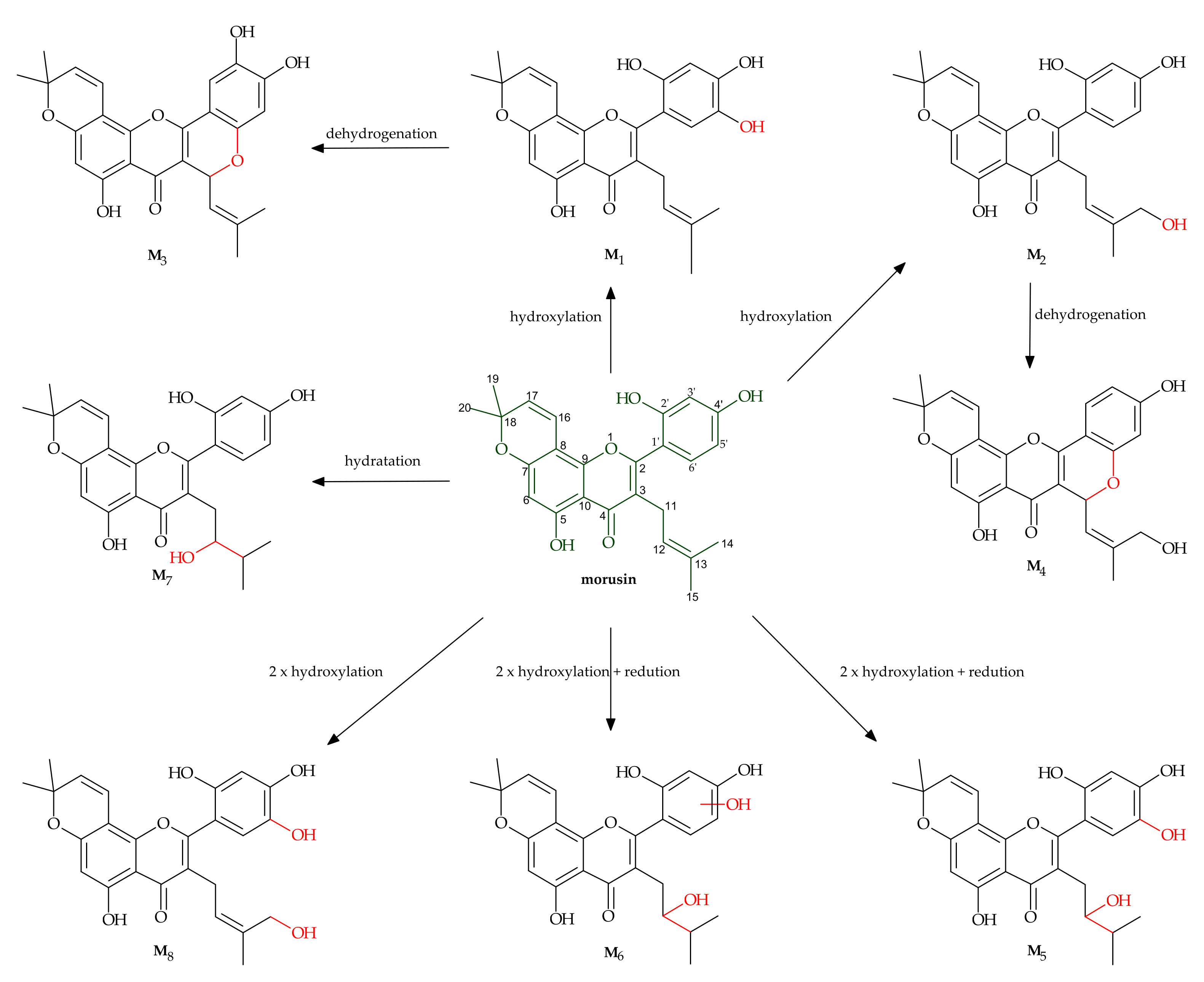
2.3. Metabolism of Morusin
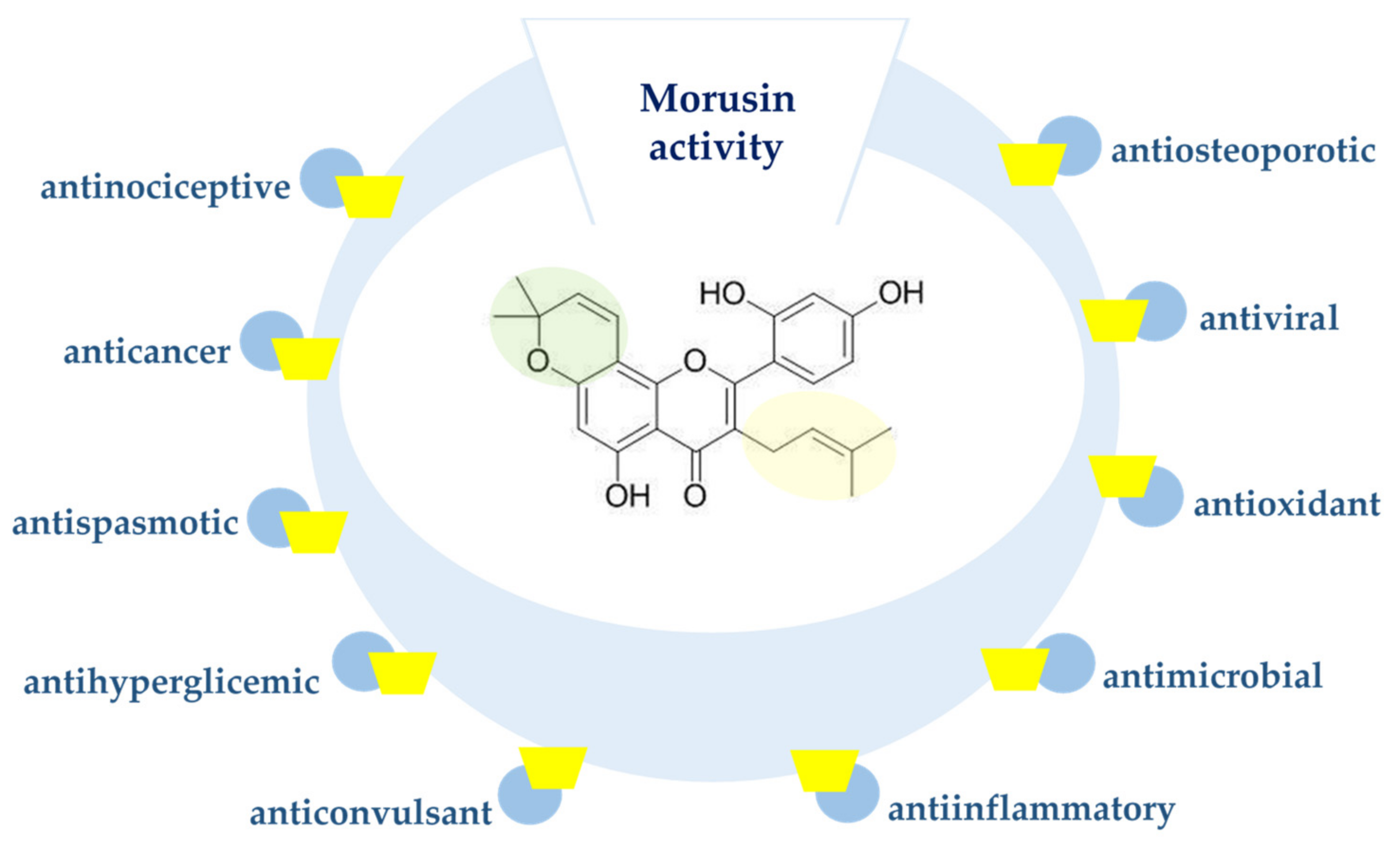
3. Biological Effects of Morusin and Its Derivatives
3.1. Anticancer Properties
3.2. Antioxidant Activity
3.3. Antibacterial and Antiprotozoal Activity
3.4. Anti-Inflammatory Activity
3.5. Anticonvulsant Activity
3.6. Application of Morusin in Memory Disorders
3.7. Anti-Hiperglycemic Action
3.8. Morusin Effect on Fat Metabolism
3.9. Antispasmodic Activity
3.10. Antiviral Activity
3.11. Antiosteoporotic Activity
3.12. Antinociceptive Activity
3.13. Enzyme-Inhibiting Activity
3.14. Antinephritis Activity of Morusin
4. New Systems for Delivery of Morusin to Cells
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| APP | amyloid precursor protein |
| ATBS | 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) |
| BACE-1 | β-amyloid cleaving enzyme 1 |
| CCA | cellular antioxidant assay |
| COX | cyclooxygenase |
| CTX | chlorotoxin |
| FDA | Food and Drug Administration |
| GABA | gamma-aminobutyric acid |
| GPDH | glycerol-3-phosphate dehydrogenase |
| GSH | glutathione |
| HIV | the human immunodeficiency viruses |
| HSV-1 | herpes simplex virus 1 |
| L-DOPA | L 3,4 dihydroxyphenylalanine |
| LOX | lipoxygenase |
| LPS | lipopolysaccharide |
| MIC | minimum inhibitory concentration |
| NFκB | nuclear factor kappa B |
| PLGA | poly(lactic-co-glycolic acid) |
| ROS | reactive oxygen species |
| STAT3 | signal transducer and activator of transcription 3 |
| TEM | transmission electron microscopy |
| TG | triglycerides |
| TRAIL | TNF-related apoptosis-inducing ligand |
| TRAP | tartrate-resistant acid phosphatase |
References
- Stompor-Gorący, M.; Bajek-Bil, A.; Machaczka, M. Chrysin: Perspectives on contemporary status and future possibilities as pro-health agent. Nutrients 2021, 13, 2038. [Google Scholar] [CrossRef]
- Turdo, A.; Glaviano, A.; Pepe, G.; Calapà, F.; Raimondo, S.; Fiori, M.; Carbone, D.; Basilicata, M.; Di Sarno, V.; Ostacolo, C.; et al. Nobiletin and xanthohumol sensitize colorectal cancer stem cells to standard chemotherapy. Cancers 2021, 13, 3927. [Google Scholar] [CrossRef]
- Koosha, S.; Mohamed, Z.; Sinniah, A.; Ibrahim, Z.A.; Seyedan, A.; Alshawsh, M.A. Antiproliferative and apoptotic activities of 8-prenylnaringenin against human colon cancer cells. Life Sci. 2019, 232, 116633. [Google Scholar] [CrossRef]
- Agarwal, S.; Mohamed, M.S.; Mizuki, T.; Maekawa, T.; Kumar, D.S. Chlorotoxin modified morusin–PLGA nanoparticles for targeted glioblastoma therapy. J. Mater. Chem. B 2019, 7, 5896–5919. [Google Scholar] [CrossRef]
- Park, H.-J.; Min, T.-R.; Chi, G.-Y.; Choi, Y.-H.; Park, S.-H. Induction of apoptosis by morusin in human non-small cell lung cancer cells by suppression of EGFR/STAT3 activation. Biochem. Biophys. Res. Commun. 2018, 505, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, X.; Ye, M. Morusin inhibits the growth of human colorectal cancer HCT116-derived sphere-forming cells via the inactivation of Akt pathway. Int. J. Mol. Med. 2021, 47, 1. [Google Scholar] [CrossRef] [PubMed]
- Martins, B.D.A.; Sande, D.; Solares, M.D.; Takahashi, J.A. Antioxidant role of morusin and mulberrofuran B in ethanol extract of Morus alba roots. Nat. Prod. Res. 2020, 1–4. [Google Scholar] [CrossRef]
- Jia, Y.; He, W.; Zhang, H.; He, L.; Wang, Y.; Zhang, T.; Peng, J.; Sun, P.; Qian, Y. Morusin ameliorates IL-1β-induced chondrocyte inflammation and osteoarthritis via NF-ĸB signal pathway. Drug Des Develop Ther. 2020, 14, 1227–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, D.; Liao, S.; Wang, W.; Mu, L.; Li, E.; Shen, W.; Liu, F.; Zou, Y. Destruction of the cell membrane and inhibition of cell phosphatidic acid biosynthesis in Staphylococcus aureus: An explanation for the antibacterial mechanism of morusin. Food Funct. 2019, 10, 6438–6446. [Google Scholar] [CrossRef]
- Syahdi, R.R.; Mun’Im, A.; Suhartanto, H.; Yanuar, A. Virtual screening of Indonesian herbal database as HIV-1 reverse transcriptase inhibitor. Bioinformation 2012, 8, 1206–1210. [Google Scholar] [CrossRef]
- Kim, T.I.; Kwon, E.B.; Oh, Y.C.; Go, Y.; Choi, J.G. Mori rhamulus and its major component morusin inhibit herpes simplex virus type 1 replication and the virus induced reactive oxygen species. Am. J. Chin. Med. 2021, 49, 163–179. [Google Scholar] [CrossRef]
- Zoofishan, Z.; Hohmann, J.; Hunyadi, A. Phenolic antioxidant of Morus nigra roots, and antitumor potential of morusin. Phytochem Rev. 2018, 17, 1031–1045. [Google Scholar] [CrossRef]
- Li, M.; Wu, X.; Wang, X.; Shen, T.; Ren, D. Two novel compounds from the root bark of Morus alba L. Nat. Prod. Res. 2017, 32, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Zhen, P.; Ni, G.; Chen, X.G.; Chen, R.Y.; Yang, H.Z.; Yu, D.Q. Chemical constituent form Morus notabilis and their cytotoxic effect. Yao Xue Xue Bao 2015, 50, 579–582. [Google Scholar] [PubMed]
- Lee, H.J.; Lyu da, H.; Koo, U.; Nam, K.W.; Hong, S.S.; Kim, K.O.; Kim, K.H.; Lee, D.; Mar, W. Protection of prenylated flavonoids from Mori Cortex Radicis (Moraceae) against nitric oxide-induced cell death in neuroblastoma SH-SY5Y cells. Arch Pharm Res. 2012, 35, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.-P.; Qiao, X.; Pan, S.; Shen, H.; Zhu, G.-F.; Hou, A.-J. New isoprenylated flavonoids and cytotoxic constituents from Artocarpus tonkinensis. J. Asian Nat. Prod. Res. 2010, 12, 586–592. [Google Scholar] [CrossRef] [PubMed]
- Shamaun, S.S.; Rahmani, M.; Hashim, N.M.; Ismail, H.B.M.; Sukari, M.A.; Lian, G.E.C.; Go, R. Prenylated flavones from Artocarpus altilis. J. Nat. Med. 2010, 64, 478–481. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Lee, W.S.; Kim, Y.S.; Curtis-Long, M.J.; Lee, B.W.; Ryu, Y.B.; Park, K.H. Isolation of cholinesterase-inhibiting flavonoids from Morus lhou. J. Agric. Food Chem. 2011, 59, 4589–4596. [Google Scholar] [CrossRef] [PubMed]
- Syah, Y.M.; Juliawaty, L.D.; Achmad, S.A.; Hakim, E.H.; Takayama, H.; Said, I.M.; Latip, J. Phenolic constituents from the wood of Morus australis with cytotoxic activity. Z Naturforsch C J Biosci. 2008, 63, 35–39. [Google Scholar]
- Ko, H.-H.; Wang, J.-J.; Lin, H.-C.; Wang, J.-P.; Lin, C.-N. Chemistry and biological activities of constituents from Morus australis. Biochim. et Biophys. Acta (BBA)-Gen. Subj. 1999, 1428, 293–299. [Google Scholar] [CrossRef]
- Wan, L.Z.; Ma, B.; Zhang, Y.Q. Preparation of morusin from Ramulus mori and its effects on mice with transplanted H22 hepatocarcinoma. Biofactors 2014, 40, 636–645. [Google Scholar] [CrossRef]
- Boonphong, S.; Baramee, A.; Kittakoop, P. Antitubercular and antiplasmodial prenylated flavones from the roots of Artocarpus altilis. Chiang Mai J. Sci. 2007, 34, 339–344. [Google Scholar]
- Tseng, T.-H.; Chuang, S.-K.; Hu, C.-C.; Chang, C.-F.; Huang, Y.-C.; Lin, C.-W.; Lee, Y.-J. The synthesis of morusin as a potent antitumor agent. Tetrahedron 2010, 66, 1335–1340. [Google Scholar] [CrossRef]
- Deng, Z.; Sun, X.; Yang, S.; Zhang, L.; Sun, P.; Teng, X.; Zhong, H. Quantification of morusin using LC-MS in rat plasma: Application to a pharmacokinetic study. Biomed. Chromatogr. 2017, 31, e4021. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Mackie, B.; Zhang, G.; Yang, S.; Song, Y.; Su, D.; Liu, Y.; Shan, L. Identification of the metabolic enzyme involved Morusin metabolism and characterization of its metabolites by ultraperformance liquid chromatography quadrupole time-of-flight mass spectrometry (UPLC/Q-TOF-MS/MS). Evid.-Based Complement. Altern. Med. 2016, 2016, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Yang, S.; Zhang, G.; Song, Y.; Su, D.; Liu, Y.; Guo, F.; Shan, L.; Cai, J. The different metabolism of morusin in various species and its potent inhibition against UDP-glucuronosyltransferase (UGT) and cytochrome p450 (CYP450) enzymes. Xenobiotica 2015, 46, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Stompor, M.; Żarowska, B. Antimicrobial activity of xanthohumol and its selected structural analogues. Molecules 2016, 21, 608. [Google Scholar] [CrossRef] [PubMed]
- Stompor, M.; Świtalska, M.; Wietrzyk, J. The influence of a single and double biotinylation of xanthohumol on its anticancer activity. Acta Biochim. Pol. 2019, 66, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Dat, N.T.; Binh, P.T.X.; Quynh, L.T.P.; Van Minh, C.; Huong, H.T.; Lee, J.J. Cytotoxic prenylated flavonoids from Morus alba. Fitoterapia 2010, 81, 1224–1227. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.-L.; Park, S.-Y.; Kang, S.; Park, D.; Kim, S.-H.; Um, J.-Y.; Jang, H.-J.; Lee, J.-H.; Jeong, C.-H.; Jang, J.-H.; et al. Morusin induces cell death through inactivating STAT3 signaling in prostate cancer cells. Am. J. Cancer Res. 2014, 5, 289–299. [Google Scholar]
- Park, Y.-J.; Choi, D.W.; Cho, S.W.; Han, J.; Yang, S.; Choi, C.Y. Stress granule formation attenuates RACK1-mediated apoptotic cell death induced by Morusin. Int. J. Mol. Sci. 2020, 21, 5360. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, Q.; Dong, L.; Liu, C.; Sun, Z.; Gao, L.; Wang, X. Morusin suppresses breast cancer cell growth in vitro and in vivo through C/EBPβ and PPARγ mediated lipoapoptosis. J. Exp. Clin. Cancer Res. 2015, 34, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.C.; Won, S.J.; Chao, C.L.; Wu, F.L.; Liu, H.S.; Ling, P.; Lin, C.N.; Su, C.L. Morusin induces apoptosis and suppresses NF-kappaB activity in human colorectal cancer HT-29 cells. Biochem. Biophys. Res. Commun. 2008, 372, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Lv, Y.; Wang, S.; Zhang, Y.-Q. Morusin suppresses A549 cell migration and induces cell apoptosis by downregulating the expression of COX-2 and VEGF genes. Oncol. Rep. 2018, 40, 504–510. [Google Scholar] [CrossRef]
- Cho, S.W.; Na, W.; Choi, M.; Kang, S.J.; Lee, S.-G.; Choi, C.Y. Autophagy inhibits cell death induced by the anti-cancer drug morusin. Am. J. Cancer Res. 2017, 7, 518–530. [Google Scholar] [PubMed]
- Kang, S.; Kim, E.-O.; Kim, S.-H.; Lee, J.; Ahn, K.S.; Yun, M.; Lee, S.-G. Morusin induces apoptosis by regulating expression of Bax and Survivin in human breast cancer cells. Oncol. Lett. 2017, 13, 4558–4562. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Wang, L.; Sun, Z.; Li, H.; Wang, Q.; Yi, C.; Wang, X. Morusin shows potent antitumor activity for human hepatocellular carcinoma in vitro and in vivo through apoptosis induction and angiogenesis inhibition. Drug Des. Dev. Ther. 2017, ume 11, 1789–1802. [Google Scholar] [CrossRef] [Green Version]
- Park, D.; Ha, I.J.; Park, S.-Y.; Choi, M.; Lim, S.-L.; Kim, S.-H.; Lee, J.-H.; Ahn, K.S.; Yun, M.; Lee, S.-G. Morusin induces TRAIL sensitization by regulating EGFR and DR5 in human glioblastoma Cells. J. Nat. Prod. 2016, 79, 317–323. [Google Scholar] [CrossRef]
- Kim, C.; Kim, J.H.; Oh, E.Y.; Nam, D.; Lee, S.G.; Lee, J.; Kim, S.-H.; Shim, B.S.; Ahn, K.S. Blockage of STAT3 signaling pathway by morusin induces apoptosis and inhibits invasion in human pancreatic tumor cells. Pancreas 2016, 45, 409–419. [Google Scholar] [CrossRef]
- Lin, W.L.; Lai, D.Y.; Lee, Y.J.; Chen, N.F.; Tseng, T.H. Antitumor progression potential of morusin suppressing STAT3 and NFĸB in human hepatoma SK-Hep1 cells. Toxicol. Lett. 2015, 232, 490–498. [Google Scholar] [CrossRef]
- Wang, L.; Guo, H.; Yang, L.; Dong, L.; Lin, C.; Zhang, J.; Lin, P.; Wang, X. Morusin inhibits human cervical cancer stem cell growth and migration through attenuation of NF-κB activity and apoptosis induction. Mol. Cell Biochem. 2013, 379, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Ding, B.; Lv, Y.; Zhang, Y.-Q. Anti-tumor effect of morusin from the branch bark of cultivated mulberry in Bel-7402 cells via the MAPK pathway. RSC Adv. 2016, 6, 17396–17404. [Google Scholar] [CrossRef]
- Xue, J.; Li, R.; Zhao, X.; Ma, C.; Lv, X.; Liu, L.; Liu, P. Morusin induces paraptosis-like cell death through mitochondrial calcium overload and dysfunction in epithelial ovarian cancer. Chem. Interact. 2018, 283, 59–74. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Liu, C.; Yang, L.; Dong, L.; Wang, L.; Wang, Q.; Li, H.; Zhang, J.; Lin, P.; Wang, X. Morusin inhibits glioblastoma stem cell growth in vitro and in vivo through stemness attenuation, adipocyte transdifferentiation, and apoptosis induction. Mol. Carcinog. 2014, 55, 77–89. [Google Scholar] [CrossRef]
- Yang, Z.G.; Matsuzaki, K.; Takamatsu, S.; Kitanaka, S. Inhibitory effects of constituents from Morus alba var. multicaulis on differentiation of 3T30L1 cells and nitric oxide production in RAW264.7 cells. Molecules 2011, 16, 6010–6022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Zhang, D.; Mao, J.; Ke, X.-X.; Zhang, R.; Yin, C.; Gao, N.; Cui, H. Morusin inhibits cell proliferation and tumor growth by down-regulating c-Myc in human gastric cancer. Oncotarget 2017, 8, 57187–57200. [Google Scholar] [CrossRef] [Green Version]
- You, S.; Kim, G.-H. Protective effect of Mori Cortex radicis extract against high glucose-induced oxidative stress in PC12 cells. Biosci. Biotechnol. Biochem. 2019, 83, 1893–1900. [Google Scholar] [CrossRef]
- Abbas, G.M.; Fatma, M.; Bar, A.; Baraka, H.N.; Gohar, A.; Lahloub, M.F. A new antioxidant stilbene and other constituents form the stem bark of Morus nigra L. Nat. Prod. Res. 2014, 28, 952–959. [Google Scholar] [CrossRef]
- Cheng, P.-S.; Hu, C.-C.; Wang, C.-J.; Lee, Y.-J.; Chung, W.-C.; Tseng, T.-H. Involvement of the antioxidative property of morusin in blocking phorbol ester–induced malignant transformation of JB6 P+ mouse epidermal cells. Chem. Interact. 2017, 264, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Treml, J.; Večeřová, P.; Herczogová, P.; Šmejkal, K. Direct and indirect antioxidant effects of selected plant phenolics in cell-based assays. Molecules 2021, 26, 2534. [Google Scholar] [CrossRef] [PubMed]
- Mazimba, O.; Majinda, R.R.T.; Motlhanka, D. Antioxidant and antibacterial constituents from Morus nigra. Afr. J. Pharm. Pharmacol. 2011, 5, 751–754. [Google Scholar] [CrossRef] [Green Version]
- Sohn, H.-Y.; Son, K.; Kwon, C.-S.; Kang, S. Antimicrobial and cytotoxic activity of 18 prenylated flavonoids isolated from medicinal plants: Morus alba L., Morus mongolica Schneider, Broussnetia papyrifera (L.) Vent, Sophora flavescens Ait and Echinosophora koreensis Nakai. Phytomedicine 2004, 11, 666–672. [Google Scholar] [CrossRef]
- Wu, S.C.; Han, F.; Song, M.R.; Chen, S.; Li, Q.; Zhang, Q.; Zhu, K.; Shen, J.Z. Natural flavones from Morus alba against methicillin-resistant Staphylococcus aureus via targeting the proton motive force and membrane permeability. J. Agric. Food Chem. 2019, 67, 10222–10234. [Google Scholar] [CrossRef]
- Yang, X.; Jiang, Y.; Yang, J.; He, J.; Sun, J.; Chen, F.; Zhang, M.; Yang, B. Prenylated flavonoids, promising nutraceuticals with impressive biological activities. Trends Food Sci. Technol. 2015, 44, 93–104. [Google Scholar] [CrossRef]
- Aelenei, P.; Rimbu, C.M.; Horhogea, C.E.; Lobiuc, A.; Neagu, A.-N.; Dunca, S.I.; Motrescu, I.; Dimitriu, G.; Aprotosoaie, A.C.; Miron, A. Prenylated phenolics as promising candidates for combination antibacterial therapy: Morusin and kuwanon G. Saudi Pharm. J. 2020, 28, 1172–1181. [Google Scholar] [CrossRef] [PubMed]
- Chi, Y.S.; Jong, H.G.; Son, K.H.; Chang, H.W.; Kang, S.S.; Kim, H.P. Effects of naturally occurring prenylated flavonoids on enzymes metabolizing arachidonic acid: Cyclooxygenases and lipoxygenases. Biochem. Pharmacol. 2001, 62, 1185–1191. [Google Scholar] [CrossRef]
- Reddy, G.R.; Ueda, N.; Hada, T.; Sackeyfio, A.C.; Yamamoto, S.; Hano, Y.; Aida, M.; Nomura, T. A prenylflavone, artonin E, as arachidonate 5-lipoxygenase inhibitor. Biochem. Pharmacol. 1991, 41, 115–118. [Google Scholar] [CrossRef]
- Cheon, B.S.; Kim, Y.H.; Son, K.S.; Chang, H.W.; Kang, S.S.; Kim, H.P. Effects of prenylated flavonoids and biflavonoids on lipopolysaccharide-induced nitric oxide production from the mouse macrophage cell line RAW 264.7. Planta Med. 2000, 66, 596–600. [Google Scholar] [CrossRef]
- Tseng, T.H.; Lin, W.L.; Zhang, C.K.; Lee, K.C.; Tung, S.Y.; Kuo, H.C. Protective effects of Morus Root extract (MRE) against lipopolysaccharide activated RAW264.7 cells and CCl4-induced mouse hepatic damage. Cell Physiol. Biochem. 2018, 51, 1376–1388. [Google Scholar] [CrossRef]
- Jin, S.E.; Ha, H.; Shin, H.-K.; Seo, C.-S. Anti-allergic and anti-inflammatory effects of kuwanon G and morusin on MC/9 mast cells and HaCaT keratinocytes. Molecules 2019, 24, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Wang, J.; Chen, J.; Zhou, L.; Wang, H.; Chen, J.; Xu, Z.; Zhu, S.; Liu, W.; Yu, R.; et al. Morusin alleviates mycoplasma pneumonia via the inhibition of Wnt/β-catenin and NF-κB signaling. Biosci. Rep. 2019, 39, BSR20190190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, C.; Cestero, J.J.; Rodriguez-Galdón, B.; Macias, P. Xanthohumol, a prenylated flavonoid from hops (Humulus lupulus L.) protects rat tissues against oxidative damage after acute ethanol administration. Toxicol. Rep. 2014, 1, 726–733. [Google Scholar] [CrossRef] [Green Version]
- Benkherouf, A.Y.; Soini, S.L.; Stompor, M.; Uusi-Oukari, M. Positive allosteric modulation of native and recombinant GABAA receptors by hops prenylflavonoids. Eur. J. Pharmacol. 2019, 852, 34–41. [Google Scholar] [CrossRef]
- Gupta, G.; Dua, K.; Kazmi, I.; Anwar, F. Anticonvulsant activity of morusin isolated from Morus alba: Modulation of GABA receptor. Biomed. Aging Pathol. 2014, 4, 29–32. [Google Scholar] [CrossRef]
- Gupta, G.; Chellappan, D.K.; Agarwal, M.; Ashwathanarayana, M.; Nammi, S.; Pabreja, K.; Dua, K. Pharmacological eveluation of the recuperative effect of morusin against aluminium trichloride (AlCl3)-induced memory impairment in rats. Cent. Nerv. Syst. Agents Med. Chem. 2017, 17, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.K.; Ryu, Y.B.; Curtis-Long, M.J.; Kim, J.Y.; Kim, D.; Lee, S.; Lee, W.S.; Park, K.H. Inhibition and structural reliability of prenylated flavones from the stem bark of Morus lhou on β-secretase (BACE-1). Bioorganic Med. Chem. Lett. 2011, 21, 2945–2948. [Google Scholar] [CrossRef]
- Milella, L.; Milazzo, S.; Se Leo, M.; Vera Saltos, M.B.; Faraone, I.; Tuccinardi, T.; Lapillo, M.; De Tommasi, N.; Braca, A. α-Glucosidase and α-amylase inhibitors from Arcytophyllum thymifolium. J. Nat. Prod. 2014, 79, 2104–2112. [Google Scholar] [CrossRef]
- Nakashima, K.; Miyashita, H.; Yoshimitsu, H.; Fujiwara, Y.; Nagai, R.; Ikeda, T. Two new prenylflavonois from Epimedii herba and their inhibitory effects on advanced glycation end-products. J. Nat. Med. 2016, 70, 290–295. [Google Scholar] [CrossRef]
- Seliger, J.M.; Misuri, L.; Maser, E.; Hintzpeter, J. The hop-derived compounds xanthohumol, isoxanthohumol and 8-prenylnaringenin are toght-binding inhibitors of human aldo-keto reductases 1B1 and 1B10. J. Enzym. Inhib. Med. Chem. 2018, 33, 607–614. [Google Scholar] [CrossRef] [Green Version]
- Rao, A.R.; Shyam, P.; Veeresham, C.; Asres, K. Aldose reductase inhibitory and antiglycation activities of four medicinal plant standarized extracts and their main constituents for the prevention of diabetic complications. Ethiop. Pharm. J. 2015, 31, 1–14. [Google Scholar] [CrossRef]
- Chen, Z.; Du, X.; Yang, Y.; Cui, X.; Zhang, Z.; Li, Y. Comparative study of chemical composition and active components against α -glucosidase of various medicinal parts of Morus alba L. Biomed. Chromatogr. 2018, 32, e4328. [Google Scholar] [CrossRef] [PubMed]
- Yimam, M.; Jiao, P.; Hong, M.; Brownell, L.; Lee, Y.C.; Hyun, E.J.; Kim, H.J.; Kim, T.W.; Nam, J.B.; Kim, M.R.; et al. Appetite suppression and antiobesity effect of a botanical composition composed of Morus alba, Yerba mate, and Magnolia officinalis. J. Obes. 2016, 2016, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.R.; Kim, J.E.; Choi, J.Y.; Park, J.J.; Kim, H.R.; Song, B.R.; Park, J.W.; Kang, M.J.; Choi, Y.W.; Kim, K.M.; et al. Morusin functions as a lipogenesis inhibitor as well as a lipolysis stimulator in differentiated 3T3-L1 and primary adipocytes. Molecules 2018, 23, 2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frölich, S.; Schubert, C.; Bienzle, U.; Jenett-Siems, K. In vitro antiplasmodial activity of prenylated chalcone derivatives of hops (Humulus lupulus) and their interaction with haemin. J. Antimicrob. Chemother. 2005, 55, 883–887. [Google Scholar] [CrossRef] [PubMed]
- Mendel, M.; Chłopecka, M.; Dziekan, N.; Karlik, W. Antispasmodic effect of selected citrus flavonoids on rat isolated jejunum specimens. Eur. J. Pharmacol. 2016, 791, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Thabti, I.; Albert, Q.; Philippot, S.; Dupire, F.; Westerhuis, B.; Fontanay, S.; Risler, A.; Kassab, T.; Elfalleh, W.; Aferchichi, A.; et al. Advances on Antiviral Activity of Morus spp. Plant Extracts: Human coronavirus and virus-related respiratory tract infections in the spotlight. Molecules 2020, 25, 1876. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zeng, L.; Xie, J.; Yue, Z.; Deng, H.; Ma, X.; Zheng, C.; Wu, X.; Luo, J.; Liu, M. Inhibition of osteoclastogenesis and bone resorption in vitro and in vivo by a prenylflavonoid xanthohumol from hops. Sci. Rep. 2015, 5, 17605. [Google Scholar] [CrossRef] [Green Version]
- Lim, R.; Li, L.; Yong, E.; Chew, N. STAT-3 regulation of CXCR4 is necessary for the prenylflavonoid icaritin to enhance mesenchymal stem cell proliferation, migration and osteogenic differentiation. Biochim. et Biophys. Acta (BBA)-Gen. Subj. 2018, 1862, 1680–1692. [Google Scholar] [CrossRef]
- Chen, M.; Han, H.; Zhou, S.; Wen, Y.; Chen, L. Morusin induces osteogenic differentiation of bone marrow mesenchymal stem cells by canonical Wnt/β-catenin pathway and prevents bone loss in an ovariectomized rat model. Stem Cell Res. Ther. 2021, 12, 1–11. [Google Scholar] [CrossRef]
- Lin, B.; Huang, J.-F.; Liu, X.-W.; Ma, X.-T.; Lu, Y.; Zhou, Y.; Guo, F.-M.; Feng, T.-T. Rapid, microwave-accelerated synthesis and anti-osteoporosis activities evaluation of morusin scaffolds and morusignin L scaffolds. Bioorganic Med. Chem. Lett. 2017, 27, 2389–2396. [Google Scholar] [CrossRef]
- de Souza, M.M.; Bittar, M.; Cechinel-Filho, V.; Yunes, R.A.; Messana, I.; Monache, F.D.; Ferrari, F. Antinociceptive properties of morusin, a prenylflavonoid isolated from Morus nigra root bark. Z Naturforsch. C J. Biosci. 2000, 55, 256–260. [Google Scholar] [CrossRef] [Green Version]
- Chaita, E.; Lambrinidis, G.; Cheimonidi, C.; Agalou, A.; Beis, D.; Trougakos, I.; Mikros, E.; Skaltsounis, A.-L.; Aligiannis, N. Anti-melanogenic properties of Greek plants. A novel depigmenting agent from Morus alba wood. Molecules 2017, 22, 514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Tao, G.; Chen, J.; Zheng, Z.-P. Characterization of a new flavone and tyrosinase inhibition constituents from the twigs of Morus alba L. Molecules 2016, 21, 1130. [Google Scholar] [CrossRef] [Green Version]
- Fukai, T.; Satoh, K.; Nomura, T.; Sakagami, H. Antinephritis and radical scavenging activity of prenylflavonoids. Fitoterapia 2003, 74, 720–724. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.; Mohamed, M.S.; Raveendran, S.; Rochani, A.K.; Maekawa, T.; Kumar, D.S. Formulation, characterization and evaluation of morusin loaded niosomes for potentiation of anticancer therapy. RSC Adv. 2018, 8, 32621–32636. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Plant | Part of Plant | Ref. |
|---|---|---|
| Morus nigra | root bark | [12] |
| Morus alba | root bark | [13] |
| Morus notabilis | twigs | [14] |
| Mori Cortex Radicis | - | [15] |
| Artocarpus tonkinensis | roots | [16] |
| Artocarpus altilis | stem bark | [17] |
| Morus lhou | root bark | [18] |
| Morus australis | cortex | [19,20] |
| Ramulus mori | mulberry Husang-32 | [21] |
| Artocarpus altilis | root | [22] |
| Metabolite | Chemical Formula | Species | |||||
|---|---|---|---|---|---|---|---|
| Rat | Monkey | Pigs | Rabbits | Human | Dog | ||
| morusin | C25H24O6 | + | + | + | + | + | + |
| M1 | C25H24O7 | + | + | + | + | + | + |
| M2 | C25H24O7 | + | + | + | + | + | + |
| M3 | C25H22O7 | + | + | + | + | + | - |
| M4 | C25H22O7 | + | + | + | - | - | + |
| M5 | C25H26O8 | + | + | + | + | + | + |
| M6 | C25H26O8 | + | + | + | - | + | + |
| M7 | C25H26O7 | + | + | + | + | + | + |
| M8 | C25H24O8 | + | + | + | + | - | - |
| Cell Line | Test | Time | IC50 (µM) | Ref. | |
|---|---|---|---|---|---|
| MDA-MB-231 | breast adenocarcinoma | MTT | 24 h | 3.2 | [23] |
| MCF-7 | breast cancer | 3.4 | |||
| A549 | lung cancer | 3.1 | |||
| DU145 | prostate cancer | MTT | 24 h | 26.27 | [30] |
| M2182 | 22.19 | ||||
| PC3 | 19.97 | ||||
| LNCaP | 21.89 | ||||
| RWPE-1 | 43.48 | ||||
| HeLa | cervical cancer | MTT | 72 h | 0.64 | [29] |
| Hep-3B | hepatocellular carcinoma | 9.21 | |||
| MCF-7 | breast cancer | 7.88 |
| Antioxidant Assays | Effect | Ref. |
|---|---|---|
| ABTS | IC50 (µM): 297.83 | [4] |
| DPPH | IC50 (µM): 1819.83 | |
| iron reducting power (%) | IC50 (µM): 13.54 | |
| ABTS | % inhibition: 61.34 | [44] |
| CAA | no effect | [50] |
| DPPH | ED50 (µg mL−1): 100.31 (30 min), 85.24 (1 h) 70.6 (3 h) 68.29 (6 h) | [51] |
| Strains | MIC (µg/mL) | Ref. |
|---|---|---|
| MSSA ATCC 29123 | 8 | [53] |
| MRSA T144 | 8 | |
| B. subtilis ATCC 6051 | 4 | |
| E. faecalis VRE1010798 | 8 | |
| E. coli ATCC 25922 | >128 | |
| E. coli B2 | >128 | |
| P. aeruginosa 14 | >128 | |
| K. pneumoniae WNX-1 | >128 | |
| S. aureus ATCC6538 | 6.3 | [9] |
| S. aureus ATCC25923 | 6.3 | |
| Salmonella ATCC9120 | 200 | |
| Salmonella DSM4224 | 250 | |
| C. albicans | >60 | [52] |
| S. typhimurium KCTC1926 | >100 | |
| S. epidermis ATCC 12228 | 20 | |
| S. aureus | 25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panek-Krzyśko, A.; Stompor-Gorący, M. The Pro-Health Benefits of Morusin Administration—An Update Review. Nutrients 2021, 13, 3043. https://doi.org/10.3390/nu13093043
Panek-Krzyśko A, Stompor-Gorący M. The Pro-Health Benefits of Morusin Administration—An Update Review. Nutrients. 2021; 13(9):3043. https://doi.org/10.3390/nu13093043
Chicago/Turabian StylePanek-Krzyśko, Anita, and Monika Stompor-Gorący. 2021. "The Pro-Health Benefits of Morusin Administration—An Update Review" Nutrients 13, no. 9: 3043. https://doi.org/10.3390/nu13093043
APA StylePanek-Krzyśko, A., & Stompor-Gorący, M. (2021). The Pro-Health Benefits of Morusin Administration—An Update Review. Nutrients, 13(9), 3043. https://doi.org/10.3390/nu13093043