
Effects of Dietary Fibers on Short-Chain Fatty Acids and Gut Microbiota Composition in Healthy Adults: A Systematic Review

 ,
,  ,
,  ,
,  ,
,  ,
,  , , ,
, , ,  ,
,  , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
- (a)
- Development of a comprehensive database of human intervention studies investigating the role of confirmed and candidate prebiotics (Phase 1);
- (b)
- Extraction of all the human intervention studies performed using DFs from the database described above (Phase 2).
2.1. Literature Search Strategy and Study Selection for the Database Development
2.2. Selection of Studies Related to Dietary Fiber Intervention
2.3. Data Extraction and Presentation and Quality Assessment
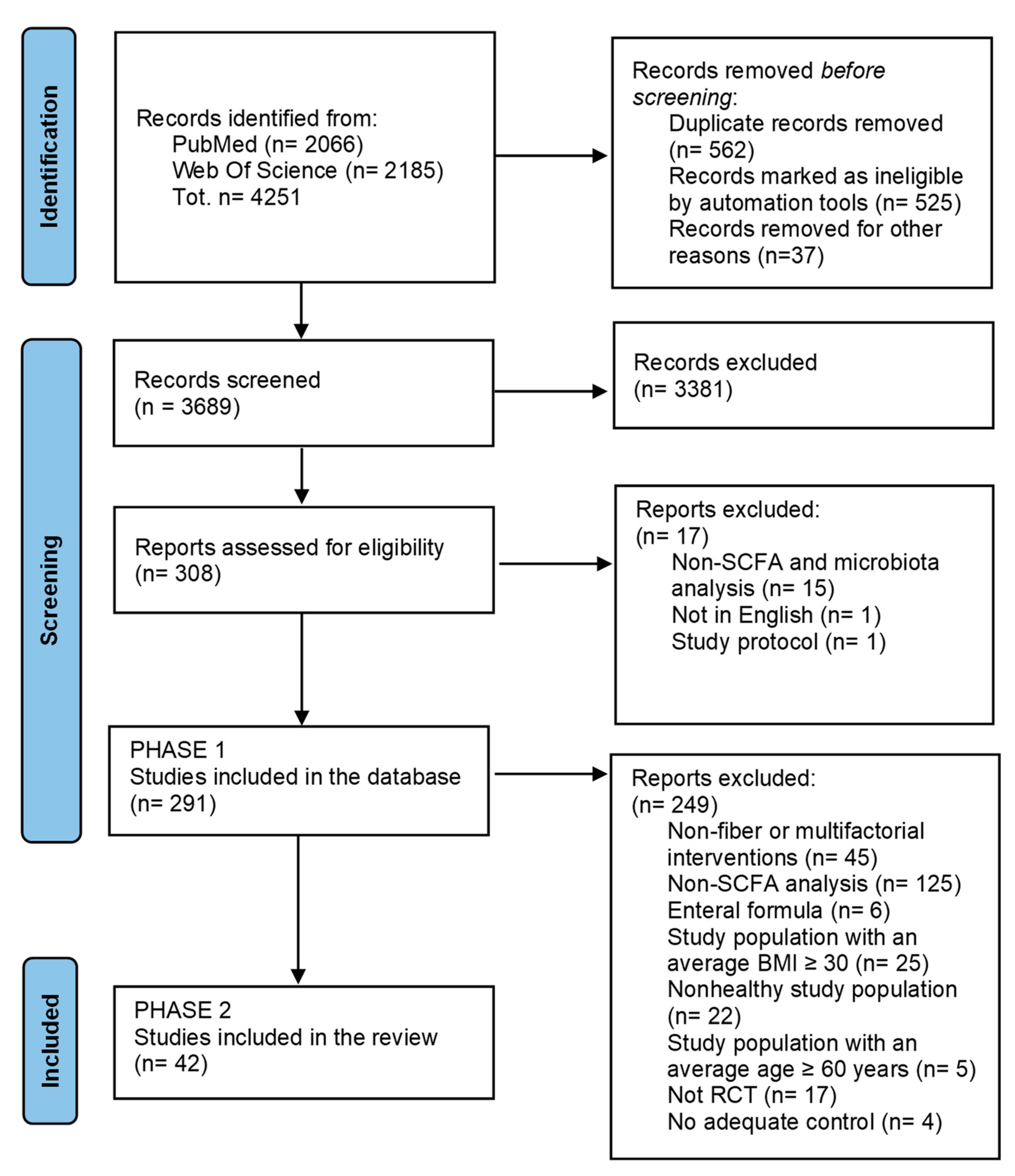
3. Results
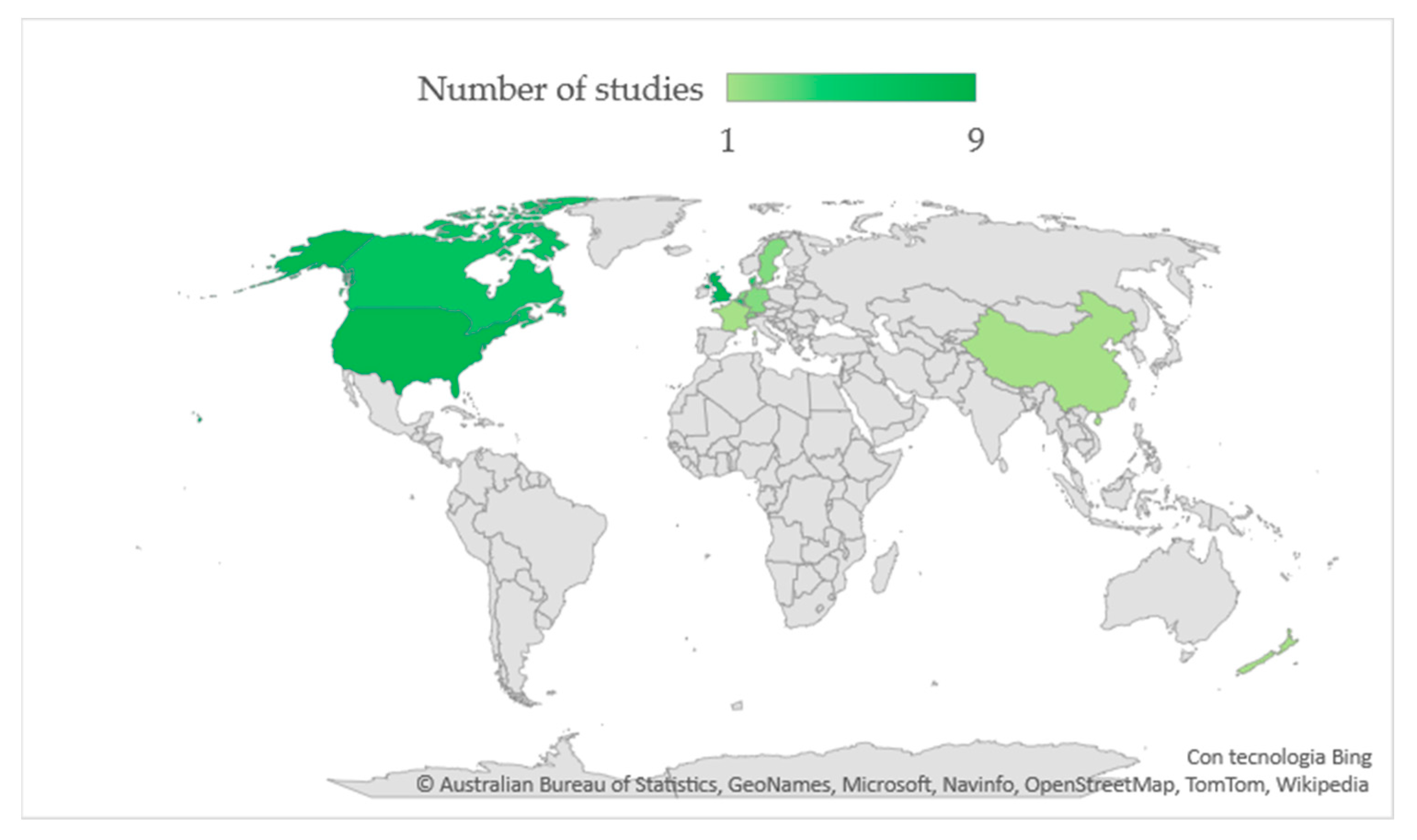
3.1. Description of Selected Trials and Risk of Bias
3.1.1. Study Design and Interventions Tested
3.1.2. Subject Characteristics
3.2. Main Findings
3.2.1. Analysis and Main Effects of Treatments on SCFAs
3.2.2. Analysis and Main Effects of Treatments on Gut Microbiota Composition
Overall Considerations
3.3. Detailed Effects of DF Interventions on the Gut Microbial Ecosystem
3.3.1. Arabinogalactan
3.3.2. Arabinoxylan Oligosaccharides (AXOS)
3.3.3. Galacto-Oligosaccharides (GOS)
3.3.4. Inulin (IN)
3.3.5. Lupin Kernel Fiber (LF) and Citrus Fiber (CF)
3.3.6. L-Rhamnose
3.3.7. Oligofructose (OF)
3.3.8. O-Fucosyllactose (2′FL) and/or Lacto-N-Neotetraose (LNnT) and Their Mix
3.3.9. Polydextrose (PDX)
3.3.10. Raffinose
3.3.11. Resistant Dextrin
3.3.12. Resistant Starch (RS)
3.3.13. Xylo-Oligosaccharide (XOS)
3.3.14. Wheat Bran Extract (WBE)
3.3.15. Fiber Mix
3.3.16. Specific Foods and Diets
3.3.17. Total Fiber Intake during Specific DF Interventions
3.4. Link between 16S rRNA Gene Amplicon Sequencing Data and SCFA Levels
3.5. Other Health-Related Findings Considered in Selected Studies
4. Discussion
5. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, B.; Grant, L.; Gidley, M.; Mikkelsen, D. Gut Fermentation of Dietary Fibres: Physico-Chemistry of Plant Cell Walls and Implications for Health. Int. J. Mol. Sci. 2017, 18, 2203. [Google Scholar] [CrossRef] [Green Version]
- Deehan, E.C.; Duar, R.M.; Armet, A.M.; Perez-Muñoz, M.E.; Jin, M.; Walter, J. Modulation of the Gastrointestinal Microbiome with Nondigestible Fermentable Carbohydrates to Improve Human Health. Microbiol. Spectr. 2017, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Hamaker, B.R.; Tuncil, Y.E. A Perspective on the Complexity of Dietary Fiber Structures and Their Potential Effect on the Gut Microbiota. J. Mol. Biol. 2014, 426, 3838–3850. [Google Scholar] [CrossRef] [PubMed]
- Holscher, H.D. Dietary Fiber and Prebiotics and the Gastrointestinal Microbiota. Gut Microbes 2017, 8, 172–184. [Google Scholar] [CrossRef]
- Koh, A.; de Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [Green Version]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The Role of Short-Chain Fatty Acids in the Interplay between Diet, Gut Microbiota, and Host Energy Metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- Duvallet, C.; Gibbons, S.M.; Gurry, T.; Irizarry, R.A.; Alm, E.J. Meta-Analysis of Gut Microbiome Studies Identifies Disease-Specific and Shared Responses. Nat. Commun. 2017, 8, 1784. [Google Scholar] [CrossRef] [Green Version]
- Brandl, B.; Lee, Y.-M.; Dunkel, A.; Hofmann, T.; Hauner, H.; Skurk, T. Effects of Extrinsic Wheat Fiber Supplementation on Fecal Weight; A Randomized Controlled Trial. Nutrients 2020, 12, 298. [Google Scholar] [CrossRef] [Green Version]
- Reimer, R.A.; Soto-Vaca, A.; Nicolucci, A.C.; Mayengbam, S.; Park, H.; Madsen, K.L.; Menon, R.; Vaughan, E.E. Effect of Chicory Inulin-Type Fructan–Containing Snack Bars on the Human Gut Microbiota in Low Dietary Fiber Consumers in a Randomized Crossover Trial. Am. J. Clin. Nutr. 2020, 111, 1286–1296. [Google Scholar] [CrossRef]
- Vester Boler, B.M.; Rossoni Serao, M.C.; Bauer, L.L.; Staeger, M.A.; Boileau, T.W.; Swanson, K.S.; Fahey, G.C. Digestive Physiological Outcomes Related to Polydextrose and Soluble Maize Fibre Consumption by Healthy Adult Men. Br. J. Nutr. 2011, 106, 1864–1871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernando, W.; Hill, J.; Zello, G.; Tyler, R.; Dahl, W.; van Kessel, A. Diets Supplemented with Chickpea or Its Main Oligosaccharide Component Raffinose Modify Faecal Microbial Composition in Healthy Adults. Benef. Microbes 2010, 1, 197–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooda, S.; Boler, B.M.V.; Serao, M.C.R.; Brulc, J.M.; Staeger, M.A.; Boileau, T.W.; Dowd, S.E.; Fahey, G.C.; Swanson, K.S. 454 Pyrosequencing Reveals a Shift in Fecal Microbiota of Healthy Adult Men Consuming Polydextrose or Soluble Corn Fiber. J. Nutr. 2012, 142, 1259–1265. [Google Scholar] [CrossRef] [PubMed]
- Slavin, J.; Feirtag, J. Chicory Inulin Does Not Increase Stool Weight or Speed up Intestinal Transit Time in Healthy Male Subjects. Food Funct. 2011, 2, 72–77. [Google Scholar] [CrossRef]
- Clarke, S.T.; Green-Johnson, J.M.; Brooks, S.P.J.; Ramdath, D.D.; Bercik, P.; Avila, C.; Inglis, G.D.; Green, J.; Yanke, L.J.; Selinger, L.B.; et al. β 2-1 Fructan Supplementation Alters Host Immune Responses in a Manner Consistent with Increased Exposure to Microbial Components: Results from a Double-Blinded, Randomised, Cross-over Study in Healthy Adults. Br. J. Nutr. 2016, 115, 1748–1759. [Google Scholar] [CrossRef] [Green Version]
- Costabile, A.; Kolida, S.; Klinder, A.; Gietl, E.; Bäuerlein, M.; Frohberg, C.; Landschütze, V.; Gibson, G.R. A Double-Blind, Placebo-Controlled, Cross-over Study to Establish the Bifidogenic Effect of a Very-Long-Chain Inulin Extracted from Globe Artichoke (Cynara scolymus) in Healthy Human Subjects. Br. J. Nutr. 2010, 104, 1007–1017. [Google Scholar] [CrossRef] [Green Version]
- Darzi, J.; Frost, G.S.; Swann, J.R.; Costabile, A.; Robertson, M.D. L-Rhamnose as a Source of Colonic Propionate Inhibits Insulin Secretion but Does Not Influence Measures of Appetite or Food Intake. Appetite 2016, 98, 142–149. [Google Scholar] [CrossRef] [Green Version]
- Deroover, L.; Verspreet, J.; Luypaerts, A.; Vandermeulen, G.; Courtin, C.; Verbeke, K. Wheat Bran Does Not Affect Postprandial Plasma Short-Chain Fatty Acids from 13C-Inulin Fermentation in Healthy Subjects. Nutrients 2017, 9, 83. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, J.; Vogt, J.; Wolever, T.M.S. Inulin Increases Short-Term Markers for Colonic Fermentation Similarly in Healthy and Hyperinsulinaemic Humans. Eur. J. Clin. Nutr. 2011, 65, 1279–1286. [Google Scholar] [CrossRef] [Green Version]
- Healey, G.; Murphy, R.; Butts, C.; Brough, L.; Whelan, K.; Coad, J. Habitual Dietary Fibre Intake Influences Gut Microbiota Response to an Inulin-Type Fructan Prebiotic: A Randomised, Double-Blind, Placebo-Controlled, Cross-over, Human Intervention Study. Br. J. Nutr. 2018, 119, 176–189. [Google Scholar] [CrossRef]
- Holscher, H.D.; Bauer, L.L.; Gourineni, V.; Pelkman, C.L.; Fahey, G.C.; Swanson, K.S. Agave Inulin Supplementation Affects the Fecal Microbiota of Healthy Adults Participating in a Randomized, Double-Blind, Placebo-Controlled, Crossover Trial. J. Nutr. 2015, 145, 2025–2032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecerf, J.-M.; Dépeint, F.; Clerc, E.; Dugenet, Y.; Niamba, C.N.; Rhazi, L.; Cayzeele, A.; Abdelnour, G.; Jaruga, A.; Younes, H.; et al. Xylo-Oligosaccharide (XOS) in Combination with Inulin Modulates Both the Intestinal Environment and Immune Status in Healthy Subjects, While XOS Alone Only Shows Prebiotic Properties. Br. J. Nutr. 2012, 108, 1847–1858. [Google Scholar] [CrossRef] [PubMed]
- Petry, N.; Egli, I.; Chassard, C.; Lacroix, C.; Hurrell, R. Inulin Modifies the Bifidobacteria Population, Fecal Lactate Concentration, and Fecal PH but Does Not Influence Iron Absorption in Women with Low Iron Status. Am. J. Clin. Nutr. 2012, 96, 325–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, O.; Sudakaran, S.; Blonquist, T.; Mah, E.; Durkee, S.; Bellamine, A. Effect of Arabinogalactan on the Gut Microbiome: A Randomized, Double-Blind, Placebo-Controlled, Crossover Trial in Healthy Adults. Nutrition 2021, 90, 111273. [Google Scholar] [CrossRef] [PubMed]
- Finegold, S.M.; Li, Z.; Summanen, P.H.; Downes, J.; Thames, G.; Corbett, K.; Dowd, S.; Krak, M.; Heber, D. Xylooligosaccharide Increases Bifidobacteria but Not Lactobacilli in Human Gut Microbiota. Food Funct. 2014, 5, 436. [Google Scholar] [CrossRef]
- Trimigno, A.; Khakimov, B.; Mejia, J.L.C.; Mikkelsen, M.S.; Kristensen, M.; Jespersen, B.M.; Engelsen, S.B. Identification of Weak and Gender Specific Effects in a Short 3 Weeks Intervention Study Using Barley and Oat Mixed Linkage β-Glucan Dietary Supplements: A Human Fecal Metabolome Study by GC-MS. Metabolomics 2017, 13, 108. [Google Scholar] [CrossRef] [Green Version]
- Lefranc-Millot, C.; Guérin-Deremaux, L.; Wils, D.; Neut, C.; Miller, L.E.; Saniez-Degrave, M.H. Impact of a Resistant Dextrin on Intestinal Ecology: How Altering the Digestive Ecosystem with NUTRIOSE®, a Soluble Fibre with Prebiotic Properties, May Be Beneficial for Health. J. Int. Med. Res. 2012, 40, 211–224. [Google Scholar] [CrossRef]
- Alfa, M.J.; Strang, D.; Tappia, P.S.; Graham, M.; van Domselaar, G.; Forbes, J.D.; Laminman, V.; Olson, N.; DeGagne, P.; Bray, D.; et al. A Randomized Trial to Determine the Impact of a Digestion Resistant Starch Composition on the Gut Microbiome in Older and Mid-Age Adults. Clin. Nutr. 2018, 37, 797–807. [Google Scholar] [CrossRef] [Green Version]
- Deehan, E.C.; Yang, C.; Perez-Muñoz, M.E.; Nguyen, N.K.; Cheng, C.C.; Triador, L.; Zhang, Z.; Bakal, J.A.; Walter, J. Precision Microbiome Modulation with Discrete Dietary Fiber Structures Directs Short-Chain Fatty Acid Production. Cell Host Microbe 2020, 27, 389–404.e6. [Google Scholar] [CrossRef]
- Zhang, L.; Ouyang, Y.; Li, H.; Shen, L.; Ni, Y.; Fang, Q.; Wu, G.; Qian, L.; Xiao, Y.; Zhang, J.; et al. Metabolic Phenotypes and the Gut Microbiota in Response to Dietary Resistant Starch Type 2 in Normal-Weight Subjects: A Randomized Crossover Trial. Sci. Rep. 2019, 9, 4736. [Google Scholar] [CrossRef]
- Boll, E.V.J.; Ekström, L.M.N.K.; Courtin, C.M.; Delcour, J.A.; Nilsson, A.C.; Björck, I.M.E.; Östman, E.M. Effects of Wheat Bran Extract Rich in Arabinoxylan Oligosaccharides and Resistant Starch on Overnight Glucose Tolerance and Markers of Gut Fermentation in Healthy Young Adults. Eur. J. Nutr. 2016, 55, 1661–1670. [Google Scholar] [CrossRef]
- Wilms, E.; An, R.; Smolinska, A.; Stevens, Y.; Weseler, A.R.; Elizalde, M.; Drittij, M.J.; Ioannou, A.; van Schooten, F.J.; Smidt, H.; et al. Galacto-Oligosaccharides Supplementation in Prefrail Older and Healthy Adults Increased Faecal Bifidobacteria, but Did Not Impact Immune Function and Oxidative Stress. Clin. Nutr. 2021, 40, 3019–3031. [Google Scholar] [CrossRef] [PubMed]
- Costabile, A.; Fava, F.; Röytiö, H.; Forssten, S.D.; Olli, K.; Klievink, J.; Rowland, I.R.; Ouwehand, A.C.; Rastall, R.A.; Gibson, G.R.; et al. Impact of Polydextrose on the Faecal Microbiota: A Double-Blind, Crossover, Placebo-Controlled Feeding Study in Healthy Human Subjects. Br. J. Nutr. 2012, 108, 471–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamichhane, S.; Yde, C.C.; Forssten, S.; Ouwehand, A.C.; Saarinen, M.; Jensen, H.M.; Gibson, G.R.; Rastall, R.; Fava, F.; Bertram, H.C. Impact of Dietary Polydextrose Fiber on the Human Gut Metabolome. J. Agric. Food Chem. 2014, 62, 9944–9951. [Google Scholar] [CrossRef] [PubMed]
- Sloan, T.J.; Jalanka, J.; Major, G.A.D.; Krishnasamy, S.; Pritchard, S.; Abdelrazig, S.; Korpela, K.; Singh, G.; Mulvenna, C.; Hoad, C.L.; et al. A Low FODMAP Diet Is Associated with Changes in the Microbiota and Reduction in Breath Hydrogen but Not Colonic Volume in Healthy Subjects. PLoS ONE 2018, 13, e0201410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Windey, K.; François, I.; Broekaert, W.; de Preter, V.; Delcour, J.A.; Louat, T.; Herman, J.; Verbeke, K. High Dose of Prebiotics Reduces Fecal Water Cytotoxicity in Healthy Subjects. Mol. Nutr. Food Res. 2014, 58, 2206–2218. [Google Scholar] [CrossRef] [PubMed]
- François, I.E.J.A.; Lescroart, O.; Veraverbeke, W.S.; Marzorati, M.; Possemiers, S.; Evenepoel, P.; Hamer, H.; Houben, E.; Windey, K.; Welling, G.W.; et al. Effects of a Wheat Bran Extract Containing Arabinoxylan Oligosaccharides on Gastrointestinal Health Parameters in Healthy Adult Human Volunteers: A Double-Blind, Randomised, Placebo-Controlled, Cross-over Trial. Br. J. Nutr. 2012, 108, 2229–2242. [Google Scholar] [CrossRef] [Green Version]
- Müller, M.; Hermes, G.D.A.; Emanuel, E.C.; Holst, J.J.; Zoetendal, E.G.; Smidt, H.; Troost, F.; Schaap, F.G.; Damink, S.O.; Jocken, J.W.E.; et al. Effect of Wheat Bran Derived Prebiotic Supplementation on Gastrointestinal Transit, Gut Microbiota, and Metabolic Health: A Randomized Controlled Trial in Healthy Adults with a Slow Gut Transit. Gut Microbes 2020, 12, 1704141. [Google Scholar] [CrossRef] [Green Version]
- Elison, E.; Vigsnaes, L.K.; Rindom Krogsgaard, L.; Rasmussen, J.; Sørensen, N.; McConnell, B.; Hennet, T.; Sommer, M.O.A.; Bytzer, P. Oral Supplementation of Healthy Adults with 2′-O-Fucosyllactose and Lacto-N-Neotetraose Is Well Tolerated and Shifts the Intestinal Microbiota. Br. J. Nutr. 2016, 116, 1356–1368. [Google Scholar] [CrossRef] [Green Version]
- Fechner, A.; Kiehntopf, M.; Jahreis, G. The Formation of Short-Chain Fatty Acids Is Positively Associated with the Blood Lipid–Lowering Effect of Lupin Kernel Fiber in Moderately Hypercholesterolemic Adults. J. Nutr. 2014, 144, 599–607. [Google Scholar] [CrossRef]
- Damen, B.; Cloetens, L.; Broekaert, W.F.; François, I.; Lescroart, O.; Trogh, I.; Arnaut, F.; Welling, G.W.; Wijffels, J.; Delcour, J.A.; et al. Consumption of Breads Containing In Situ–Produced Arabinoxylan Oligosaccharides Alters Gastrointestinal Effects in Healthy Volunteers. J. Nutr. 2012, 142, 470–477. [Google Scholar] [CrossRef] [Green Version]
- Walton, G.E.; Lu, C.; Trogh, I.; Arnaut, F.; Gibson, G.R. A Randomised, Double-Blind, Placebo Controlled Cross-over Study to Determine the Gastrointestinal Effects of Consumption of Arabinoxylan-Oligosaccharides Enriched Bread in Healthy Volunteers. Nutr. J. 2012, 11, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ampatzoglou, A.; Atwal, K.K.; Maidens, C.M.; Williams, C.L.; Ross, A.B.; Thielecke, F.; Jonnalagadda, S.S.; Kennedy, O.B.; Yaqoob, P. Increased Whole Grain Consumption Does Not Affect Blood Biochemistry, Body Composition, or Gut Microbiology in Healthy, Low-Habitual Whole Grain Consumers. J. Nutr. 2015, 145, 215–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, A.B.; Pere-Trépat, E.; Montoliu, I.; Martin, F.-P.J.; Collino, S.; Moco, S.; Godin, J.-P.; Cléroux, M.; Guy, P.A.; Breton, I.; et al. A Whole-Grain-Rich Diet Reduces Urinary Excretion of Markers of Protein Catabolism and Gut Microbiota Metabolism in Healthy Men after One Week 1,2. Proteom. Metab. J. Nutr 2013, 143, 766–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanegas, S.M.; Meydani, M.; Barnett, J.B.; Goldin, B.; Kane, A.; Rasmussen, H.; Brown, C.; Vangay, P.; Knights, D.; Jonnalagadda, S.; et al. Substituting Whole Grains for Refined Grains in a 6-Wk Randomized Trial Has a Modest Effect on Gut Microbiota and Immune and Inflammatory Markers of Healthy Adults. Am. J. Clin. Nutr. 2017, 105, 635–650. [Google Scholar] [CrossRef] [Green Version]
- Vuholm, S.; Nielsen, D.S.; Iversen, K.N.; Suhr, J.; Westermann, P.; Krych, L.; Andersen, J.R.; Kristensen, M. Whole-Grain Rye and Wheat Affect Some Markers of Gut Health without Altering the Fecal Microbiota in Healthy Overweight Adults: A 6-Week Randomized Trial. J. Nutr. 2017, 147, 2067–2075. [Google Scholar] [CrossRef] [Green Version]
- Carvalho-Wells, A.L.; Helmolz, K.; Nodet, C.; Molzer, C.; Leonard, C.; McKevith, B.; Thielecke, F.; Jackson, K.G.; Tuohy, K.M. Determination of the in Vivo Prebiotic Potential of a Maize-Based Whole Grain Breakfast Cereal: A Human Feeding Study. Br. J. Nutr. 2010, 104, 1353–1356. [Google Scholar] [CrossRef] [Green Version]
- Connolly, M.L.; Tzounis, X.; Tuohy, K.M.; Lovegrove, J.A. Hypocholesterolemic and Prebiotic Effects of a Whole-Grain Oat-Based Granola Breakfast Cereal in a Cardio-Metabolic “At Risk” Population. Front. Microbiol. 2016, 7, 1675. [Google Scholar] [CrossRef] [Green Version]
- Duysburgh, C.; van den Abbeele, P.; Kamil, A.; Fleige, L.; de Chavez, P.J.; Chu, Y.; Barton, W.; O’Sullivan, O.; Cotter, P.D.; Quilter, K.; et al. In Vitro–in Vivo Validation of Stimulatory Effect of Oat Ingredients on Lactobacilli. Pathogens 2021, 10, 235. [Google Scholar] [CrossRef]
- Nilsson, A.C.; Östman, E.M.; Knudsen, K.E.B.; Holst, J.J.; Björck, I.M. A Cereal-Based Evening Meal Rich in Indigestible Carbohydrates Increases Plasma Butyrate the Next Morning 1,2. J. Nutr. 2010, 140, 1932–1936. [Google Scholar] [CrossRef]
- Rahat-Rozenbloom, S.; Fernandes, J.; Gloor, G.B.; Wolever, T.M.S. Evidence for Greater Production of Colonic Short-Chain Fatty Acids in Overweight than Lean Humans. Int. J. Obes. 2014, 38, 1525–1531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priyadarshini, M.; Thomas, A.; Reisetter, A.C.; Scholtens, D.M.; Wolever, T.M.S.; Josefson, J.L.; Layden, B.T. Maternal Short-Chain Fatty Acids Are Associated with Metabolic Parameters in Mothers and Newborns. Transl. Res. 2014, 164, 153–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, J.; Kulemeier, J.; Lembcke, B.; Caspary, W.F. Simple and Rapid Method for Determination of Short-Chain Fatty Acids in Biological Materials by High-Performance Liquid Chromatography with Ultraviolet Detection. J. Chromatogr. B Biomed. Sci. Appl. 1992, 576, 53–61. [Google Scholar] [CrossRef]
- Kotani, A.; Miyaguchi, Y.; Kohama, M.; Ohtsuka, T.; Shiratori, T.; Kusu, F. Determination of Short-Chain Fatty Acids in Rat and Human Feces by High-Performance Liquid Chromatography with Electrochemical Detection. Anal. Sci. 2009, 25, 1007–1011. [Google Scholar] [CrossRef] [Green Version]
- van Eijk, H.M.H.; Bloemen, J.G.; Dejong, C.H.C. Application of Liquid Chromatography–Mass Spectrometry to Measure Short Chain Fatty Acids in Blood. J. Chromatogr. B 2009, 877, 719–724. [Google Scholar] [CrossRef]
- Han, J.; Lin, K.; Sequeira, C.; Borchers, C.H. An Isotope-Labeled Chemical Derivatization Method for the Quantitation of Short-Chain Fatty Acids in Human Feces by Liquid Chromatography–Tandem Mass Spectrometry. Anal. Chim. Acta 2015, 854, 86–94. [Google Scholar] [CrossRef]
- Zhao, G.; Liu, J.F.; Nyman, M.; Jönsson, J.Å. Determination of Short-Chain Fatty Acids in Serum by Hollow Fiber Supported Liquid Membrane Extraction Coupled with Gas Chromatography. J. Chromatogr. B 2007, 846, 202–208. [Google Scholar] [CrossRef]
- García-Villalba, R.; Giménez-Bastida, J.A.; García-Conesa, M.T.; Tomás-Barberán, F.A.; Carlos Espín, J.; Larrosa, M. Alternative Method for Gas Chromatography-Mass Spectrometry Analysis of Short-Chain Fatty Acids in Faecal Samples. J. Sep. Sci. 2012, 35, 1906–1913. [Google Scholar] [CrossRef]
- Garcia, A.; Olmo, B.; Lopez-Gonzalvez, A.; Cornejo, L.; Rupérez, F.J.; Barbas, C. Capillary Electrophoresis for Short Chain Organic Acids in Faeces: Reference Values in a Mediterranean Elderly Population. J. Pharm. Biomed. Anal. 2008, 46, 356–361. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, H.; Zhu, M.J. A Sensitive GC/MS Detection Method for Analyzing Microbial Metabolites Short Chain Fatty Acids in Fecal and Serum Samples. Talanta 2019, 196, 249–254. [Google Scholar] [CrossRef]
- Ahn, Y.G.; Jeon, S.H.; Lim, H.B.; Choi, N.R.; Hwang, G.S.; Kim, Y.P.; Lee, J.Y. Analysis of Polycyclic Aromatic Hydrocarbons in Ambient Aerosols by Using One-Dimensional and Comprehensive Two-Dimensional Gas Chromatography Combined with Mass Spectrometric Method: A Comparative Study. J. Anal. Methods Chem. 2018, 2018, 8341630. [Google Scholar] [CrossRef] [PubMed]
- Misra, B.B.; Bassey, E.; Olivier, M. Comparison of a GC-Orbitrap-MS with Parallel GC-FID Capabilities for Metabolomics of Human Serum. bioRxiv 2019, 740795. [Google Scholar] [CrossRef] [Green Version]
- Park, N.H.; Kim, M.S.; Lee, W.; Lee, M.E.; Hong, J. An in Situ Extraction and Derivatization Method for Rapid Analysis of Short-Chain Fatty Acids in Rat Fecal Samples by Gas Chromatography Tandem Mass Spectrometry. Anal. Methods 2017, 9, 2351–2356. [Google Scholar] [CrossRef]
- Moreau, N.M.; Goupry, S.M.; Antignac, J.P.; Monteau, F.J.; le Bizec, B.J.; Champ, M.M.; Martin, L.J.; Dumon, H.J. Simultaneous Measurement of Plasma Concentrations and 13C-Enrichment of Short-Chain Fatty Acids, Lactic Acid and Ketone Bodies by Gas Chromatography Coupled to Mass Spectrometry. J. Chromatogr. B 2003, 784, 395–403. [Google Scholar] [CrossRef]
- Pouteau, E.; Meirim, I.; Métairon, S.; Fay, L.B. Acetate, Propionate and Butyrate in Plasma: Determination of the Concentration and Isotopic Enrichment by Gas Chromatography/Mass Spectrometry with Positive Chemical Ionization. J. Mass Spectrom. 2001, 36, 798–805. [Google Scholar] [CrossRef]
- Cai, J.; Zhang, J.; Tian, Y.; Zhang, L.; Hatzakis, E.; Krausz, K.W.; Smith, P.B.; Gonzalez, F.J.; Patterson, A.D. Orthogonal Comparison of GC-MS and 1H NMR Spectroscopy for Short Chain Fatty Acid Quantitation. Anal. Chem. 2017, 89, 7900–7906. [Google Scholar] [CrossRef]
- Huart, J.; Leenders, J.; Taminiau, B.; Descy, J.; Saint-Remy, A.; Daube, G.; Krzesinski, J.M.; Melin, P.; de Tullio, P.; Jouret, F. Gut Microbiota and Fecal Levels of Short-Chain Fatty Acids Differ Upon 24-Hour Blood Pressure Levels in Men. Hypertension 2019, 74, 1005–1013. [Google Scholar] [CrossRef]
- Dei Cas, M.; Paroni, R.; Saccardo, A.; Casagni, E.; Arnoldi, S.; Gambaro, V.; Saresella, M.; Mario, C.; la Rosa, F.; Marventano, I.; et al. A Straightforward LC-MS/MS Analysis to Study Serum Profile of Short and Medium Chain Fatty Acids. J. Chromatogr. B 2020, 1154, 121982. [Google Scholar] [CrossRef]
- Schäfer, K. Analysis of Short Chain Fatty Acids from Different Intestinal Samples by Capillary Gas Chromatography. Chromatographia 1995, 40, 550–556. [Google Scholar] [CrossRef]
- Taverniti, V.; Guglielmetti, S. Methodological Issues in the Study of Intestinal Microbiota in Irritable Bowel Syndrome. World J. Gastroenterol. 2014, 20, 8821–8836. [Google Scholar] [CrossRef]
- Flint, H.J.; Bayer, E.A.; Rincon, M.T.; Lamed, R.; White, B.A. Polysaccharide Utilization by Gut Bacteria: Potential for New Insights from Genomic Analysis. Nat. Rev. Microbiol. 2008, 6, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Tannock, G.W.; Liu, Y. Guided Dietary Fibre Intake as a Means of Directing Short-Chain Fatty Acid Production by the Gut Microbiota. J. R. Soc. N. Z. 2019, 50, 434–455. [Google Scholar] [CrossRef]
- Smith, N.W.; Shorten, P.R.; Altermann, E.; Roy, N.C.; McNabb, W.C. The Classification and Evolution of Bacterial Cross-Feeding. Front. Ecol. Evol. 2019, 7, 153. [Google Scholar] [CrossRef] [Green Version]
- Peterson, C.T.; Perez Santiago, J.; Iablokov, S.N.; Chopra, D.; Rodionov, D.A.; Peterson, S.N. Short-Chain Fatty Acids Modulate Healthy Gut Microbiota Composition and Functional Potential. Curr. Microbiol. 2022, 79, 128. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Unno, T.; Kim, B.Y.; Park, M.S. Sex Differences in Gut Microbiota. World J. Men’s Health 2020, 38, 48–60. [Google Scholar] [CrossRef]
- Arner, P. Insulin Resistance in Type 2 Diabetes: Role of Fatty Acids. Diabetes/Metab. Res. Rev. 2002, 18, S5–S9. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, A.; Granfeldt, Y.; Östman, E.; Preston, T.; Björck, I. Effects of GI and Content of Indigestible Carbohydrates of Cereal-Based Evening Meals on Glucose Tolerance at a Subsequent Standardised Breakfast. Eur. J. Clin. Nutr. 2006, 60, 1092–1099. [Google Scholar] [CrossRef]
- Nilsson, A.C.; Östman, E.M.; Holst, J.J.; Björck, I.M.E. Including Indigestible Carbohydrates in the Evening Meal of Healthy Subjects Improves Glucose Tolerance, Lowers Inflammatory Markers, and Increases Satiety after a Subsequent Standardized Breakfast. J. Nutr. 2008, 138, 732–739. [Google Scholar] [CrossRef] [Green Version]
- Wolever, T.M.S.; Bentum-Williams, A.; Jenkins, D.J. Physiological Modulation of Plasma Free Fatty Acid Concentrations by Diet: Metabolic Implications in Nondiabetic Subjects. Diabetes Care 1995, 18, 962–970. [Google Scholar] [CrossRef]
- Chambers, E.S.; Morrison, D.J.; Frost, G. Control of Appetite and Energy Intake by SCFA: What Are the Potential Underlying Mechanisms? Proc. Nutr. Soc. 2015, 74, 328–336. [Google Scholar] [CrossRef]
- De Vadder, F.; Kovatcheva-Datchary, P.; Goncalves, D.; Vinera, J.; Zitoun, C.; Duchampt, A.; Bäckhed, F.; Mithieux, G. Microbiota-Generated Metabolites Promote Metabolic Benefits via Gut-Brain Neural Circuits. Cell 2014, 156, 84–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Psichas, A.; Sleeth, M.L.; Murphy, K.G.; Brooks, L.; Bewick, G.A.; Hanyaloglu, A.C.; Ghatei, M.A.; Bloom, S.R.; Frost, G. The Short Chain Fatty Acid Propionate Stimulates GLP-1 and PYY Secretion via Free Fatty Acid Receptor 2 in Rodents. Int. J. Obes. 2014, 39, 424–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, J.C.; Allen, C.R.; Cushman, S.A. Operationalizing Ecological Resilience Concepts for Managing Species and Ecosystems at Risk. Front. Ecol. Evol. 2019, 7, 241. [Google Scholar] [CrossRef] [Green Version]
- Bernardi, S.; del Bo’, C.; Marino, M.; Gargari, G.; Cherubini, A.; Andrés-Lacueva, C.; Hidalgo-Liberona, N.; Peron, G.; González-Dominguez, R.; Kroon, P.; et al. Polyphenols and Intestinal Permeability: Rationale and Future Perspectives. J. Agric. Food Chem. 2020, 68, 1816–1829. [Google Scholar] [CrossRef] [PubMed]
- Peron, G.; Gargari, G.; Meroño, T.; Miñarro, A.; Lozano, E.V.; Escuder, P.C.; González-Domínguez, R.; Hidalgo-Liberona, N.; del Bo’, C.; Bernardi, S.; et al. Crosstalk among Intestinal Barrier, Gut Microbiota and Serum Metabolome after a Polyphenol-Rich Diet in Older Subjects with “Leaky Gut”: The MaPLE Trial. Clin. Nutr. 2021, 40, 5288–5297. [Google Scholar] [CrossRef]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; deRoos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites Produced by Commensal Bacteria Promote Peripheral Regulatory T-Cell Generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef]
- Rivera-Chávez, F.; Zhang, L.F.; Faber, F.; Lopez, C.A.; Byndloss, M.X.; Olsan, E.E.; Xu, G.; Velazquez, E.M.; Lebrilla, C.B.; Winter, S.E.; et al. Depletion of Butyrate-Producing Clostridia from the Gut Microbiota Drives an Aerobic Luminal Expansion of Salmonella. Cell Host Microbe 2016, 19, 443–454. [Google Scholar] [CrossRef] [Green Version]
- van der Beek, C.M.; Dejong, C.H.C.; Troost, F.J.; Masclee, A.A.M.; Lenaerts, K. Role of Short-Chain Fatty Acids in Colonic Inflammation, Carcinogenesis, and Mucosal Protection and Healing. Nutr. Rev. 2017, 75, 286–305. [Google Scholar] [CrossRef] [Green Version]
- Barber, T.M.; Kabisch, S.; Pfeiffer, A.F.H.; Weickert, M.O. The Health Benefits of Dietary Fibre. Nutrients 2020, 12, 3209. [Google Scholar] [CrossRef]
- Sakata, T. Pitfalls in Short-Chain Fatty Acid Research: A Methodological Review. Anim. Sci. J. 2019, 90, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Primec, M.; Mičetić-Turk, D.; Langerholc, T. Analysis of Short-Chain Fatty Acids in Human Feces: A Scoping Review. Anal. Biochem. 2017, 526, 9–21. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Parameter | Inclusion Criteria |
|---|---|
| Population | Age 18 to 60 years and BMI < 30 kg/m2 |
| Intervention | Dietary intervention studies involving the consumption of dietary fiber, supplied with food or supplements, without probiotics and/or synbiotics |
| Comparison | Control group |
| Outcome | At least SCFA profile |
| Study design | Human clinical and randomized controlled trials |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vinelli, V.; Biscotti, P.; Martini, D.; Del Bo’, C.; Marino, M.; Meroño, T.; Nikoloudaki, O.; Calabrese, F.M.; Turroni, S.; Taverniti, V.; et al. Effects of Dietary Fibers on Short-Chain Fatty Acids and Gut Microbiota Composition in Healthy Adults: A Systematic Review. Nutrients 2022, 14, 2559. https://doi.org/10.3390/nu14132559
Vinelli V, Biscotti P, Martini D, Del Bo’ C, Marino M, Meroño T, Nikoloudaki O, Calabrese FM, Turroni S, Taverniti V, et al. Effects of Dietary Fibers on Short-Chain Fatty Acids and Gut Microbiota Composition in Healthy Adults: A Systematic Review. Nutrients. 2022; 14(13):2559. https://doi.org/10.3390/nu14132559
Chicago/Turabian StyleVinelli, Valentina, Paola Biscotti, Daniela Martini, Cristian Del Bo’, Mirko Marino, Tomás Meroño, Olga Nikoloudaki, Francesco Maria Calabrese, Silvia Turroni, Valentina Taverniti, and et al. 2022. "Effects of Dietary Fibers on Short-Chain Fatty Acids and Gut Microbiota Composition in Healthy Adults: A Systematic Review" Nutrients 14, no. 13: 2559. https://doi.org/10.3390/nu14132559
APA StyleVinelli, V., Biscotti, P., Martini, D., Del Bo’, C., Marino, M., Meroño, T., Nikoloudaki, O., Calabrese, F. M., Turroni, S., Taverniti, V., Unión Caballero, A., Andrés-Lacueva, C., Porrini, M., Gobbetti, M., De Angelis, M., Brigidi, P., Pinart, M., Nimptsch, K., Guglielmetti, S., & Riso, P. (2022). Effects of Dietary Fibers on Short-Chain Fatty Acids and Gut Microbiota Composition in Healthy Adults: A Systematic Review. Nutrients, 14(13), 2559. https://doi.org/10.3390/nu14132559