
Comparative Effects of Allulose, Fructose, and Glucose on the Small Intestine

 ,
,  ,
,
Abstract
:1. Introduction
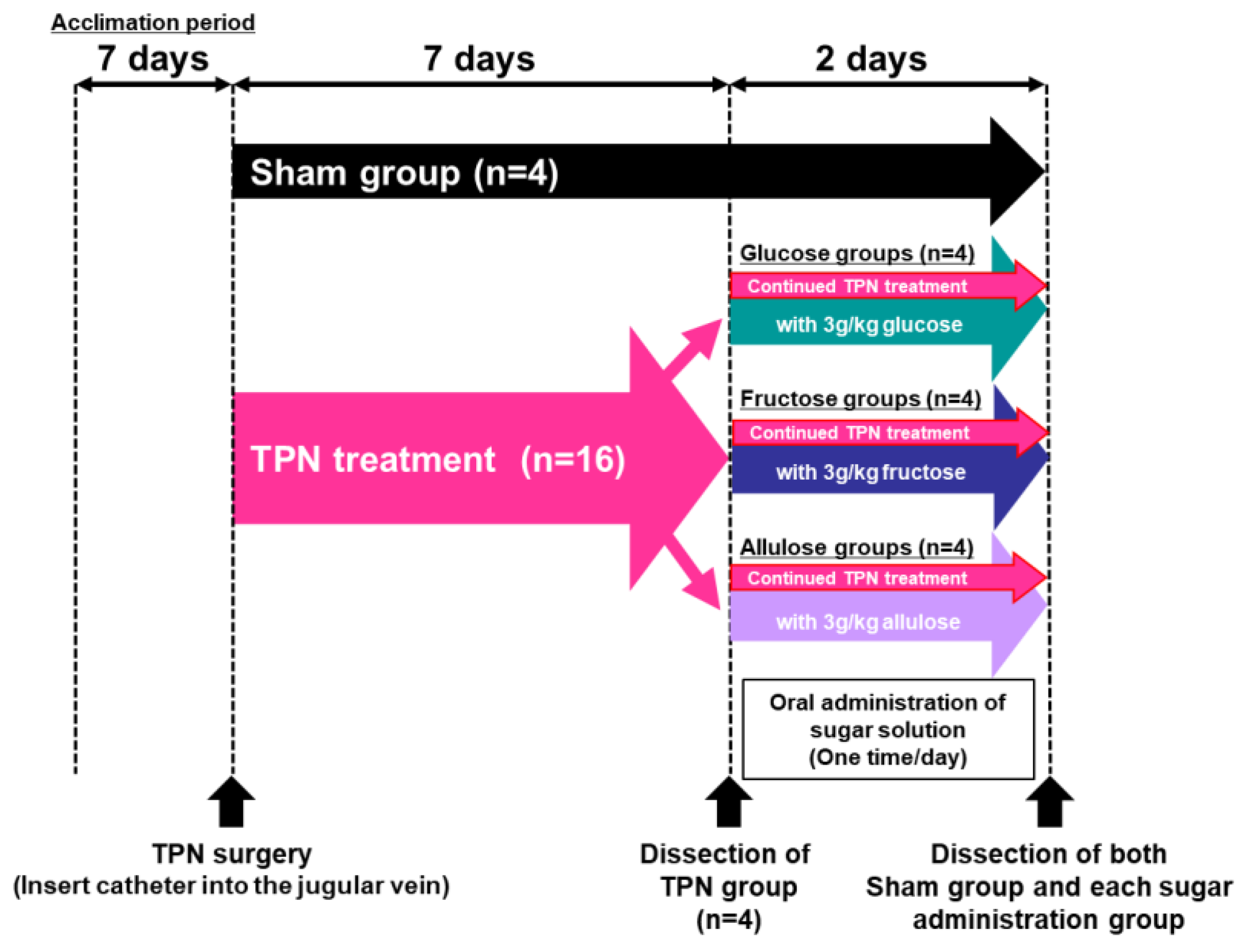
2. Materials and Methods
3. Results
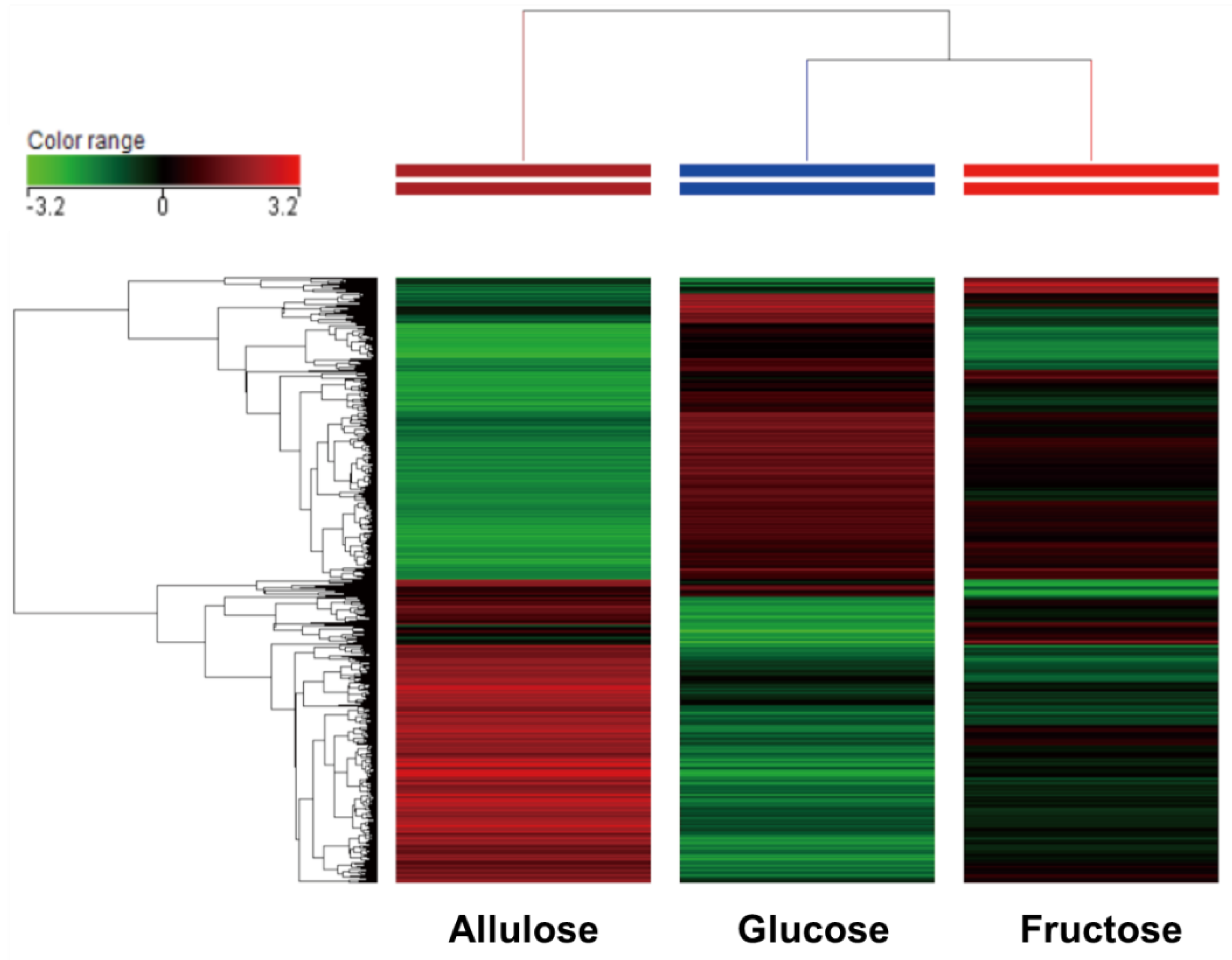
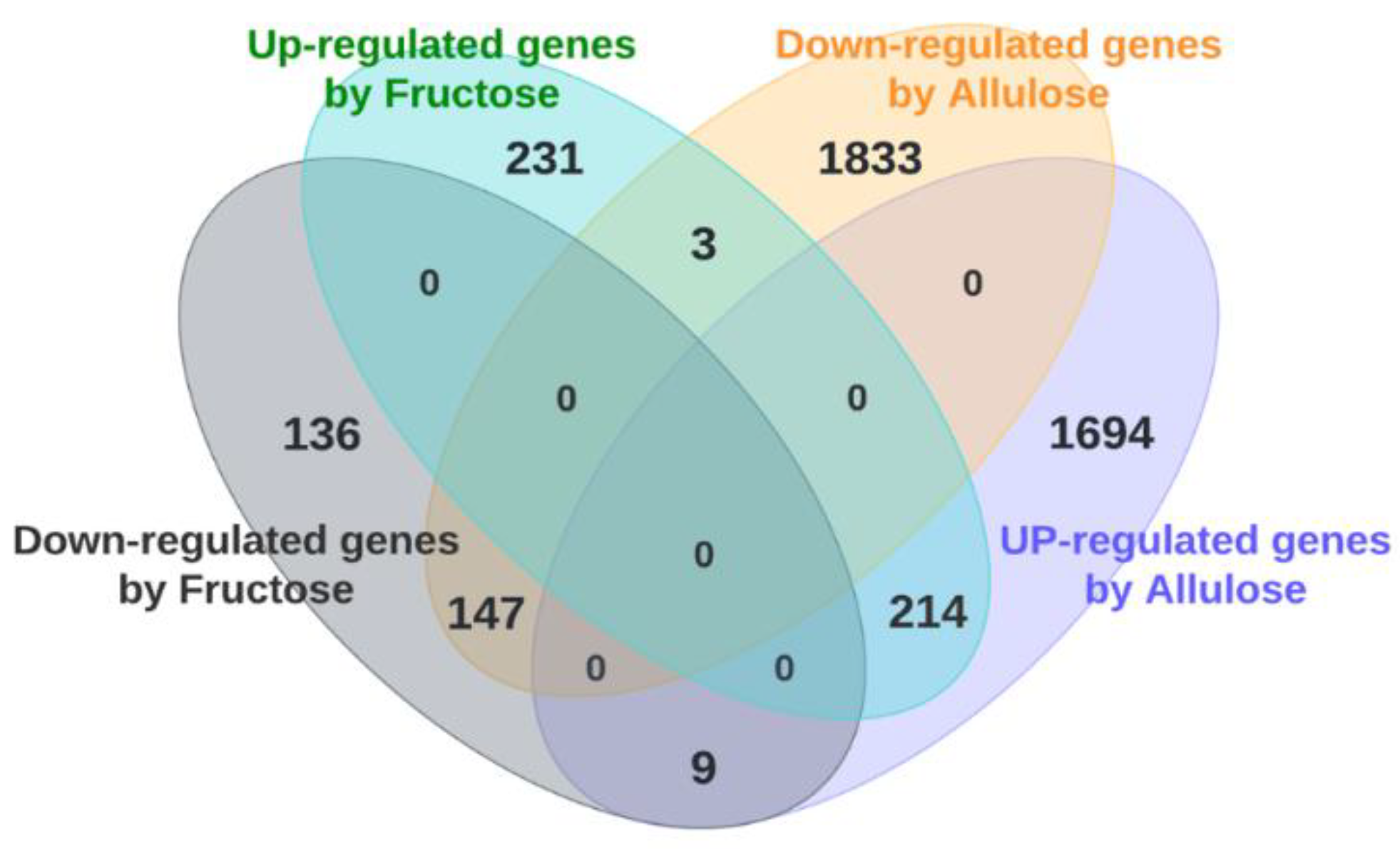
3.1. DNA Microarray Analysis (Clustering Analysis, Venn Diagram Analysis and Transcriptome Analysis)
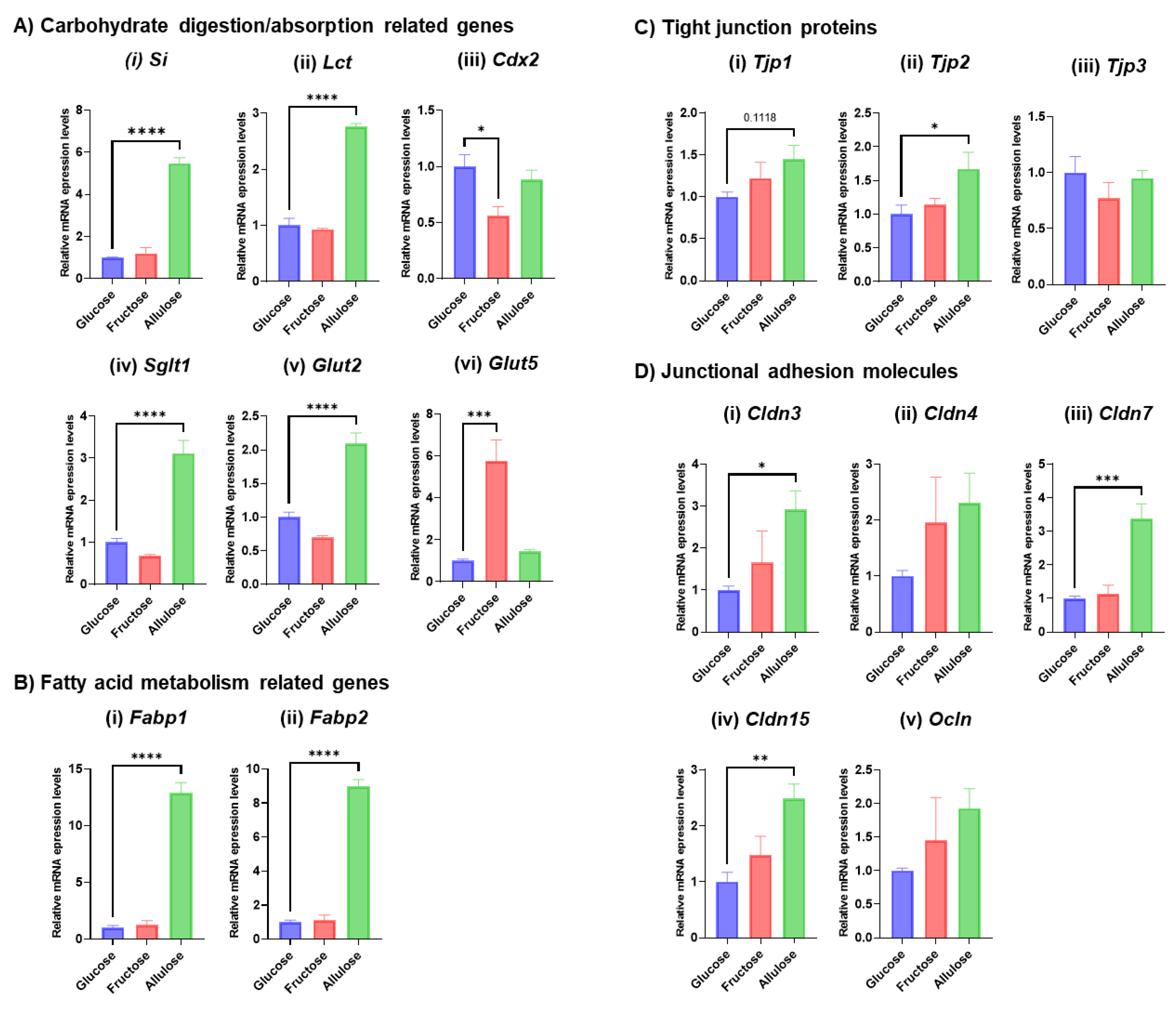
3.2. Expression of Representative Genes Related to Digestive System Development and Function
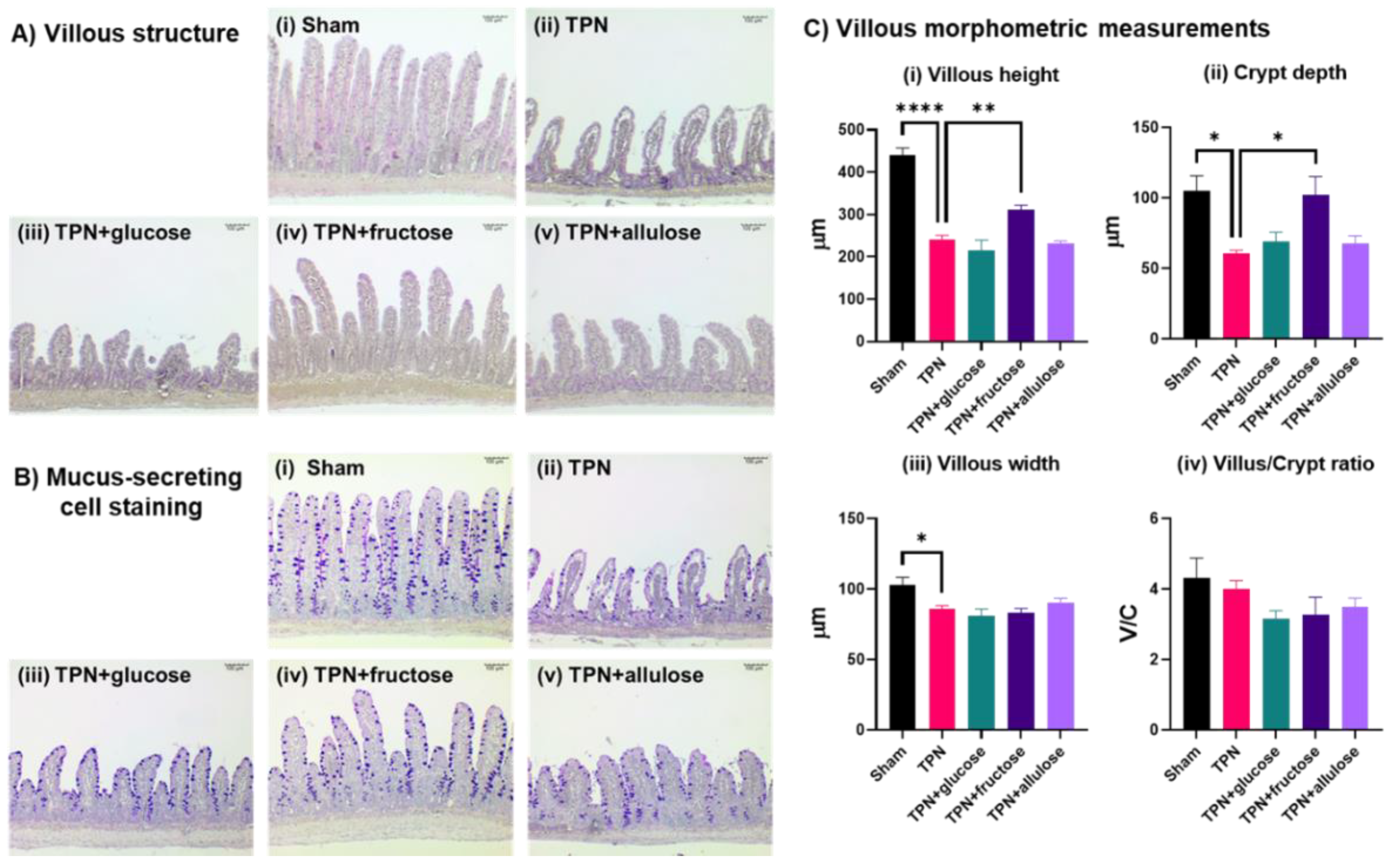
3.3. Evaluating the Potential of Allulose in Rescuing TPN-Induced Mucosal Atrophy
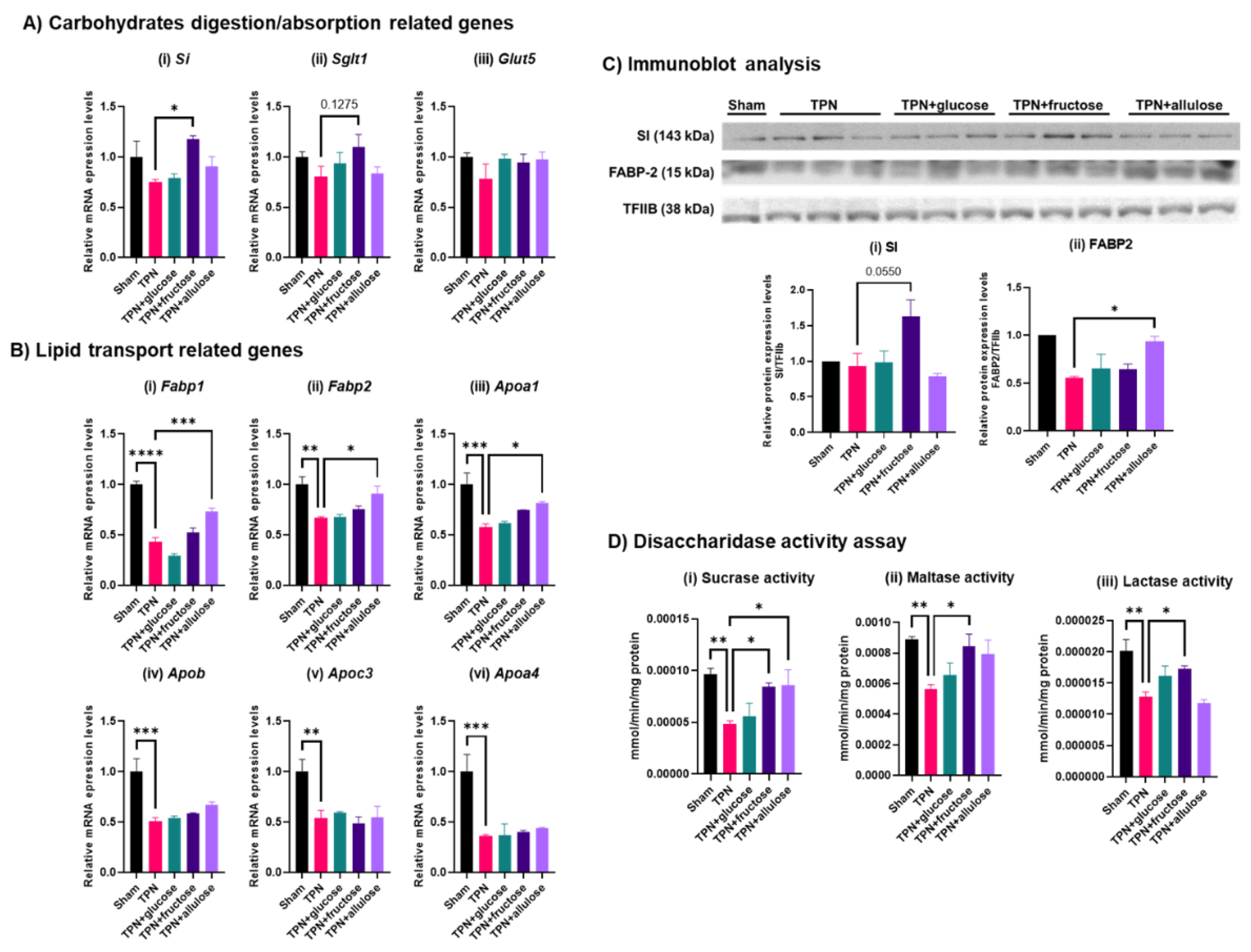
3.4. Effect of Monosaccharide Supplementation on Nutrient Metabolism and Transport Genes in TPN
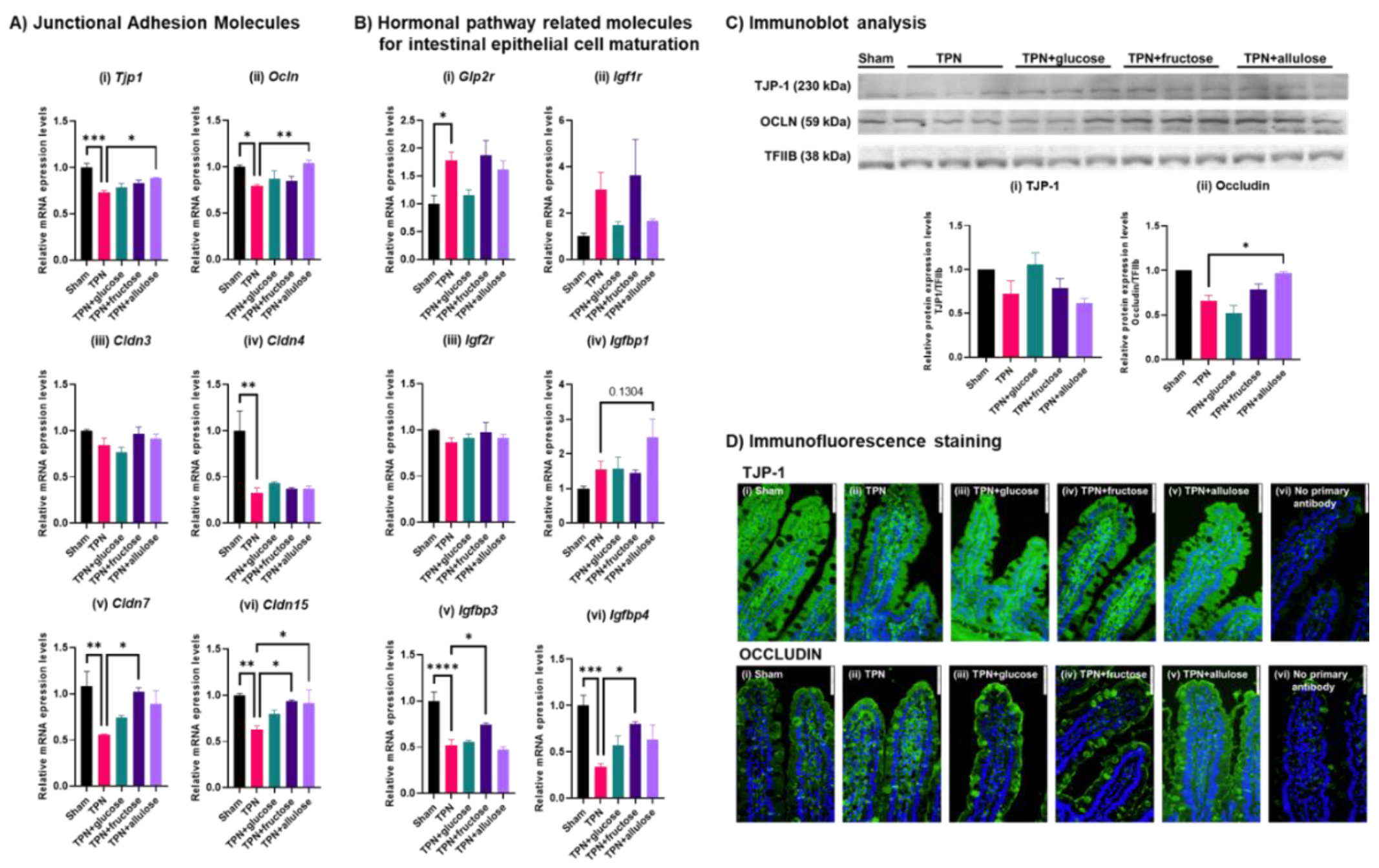
3.5. Evaluating the Potential of Allulose in Rescuing TPN-Induced Alterations in the Gut Barrier
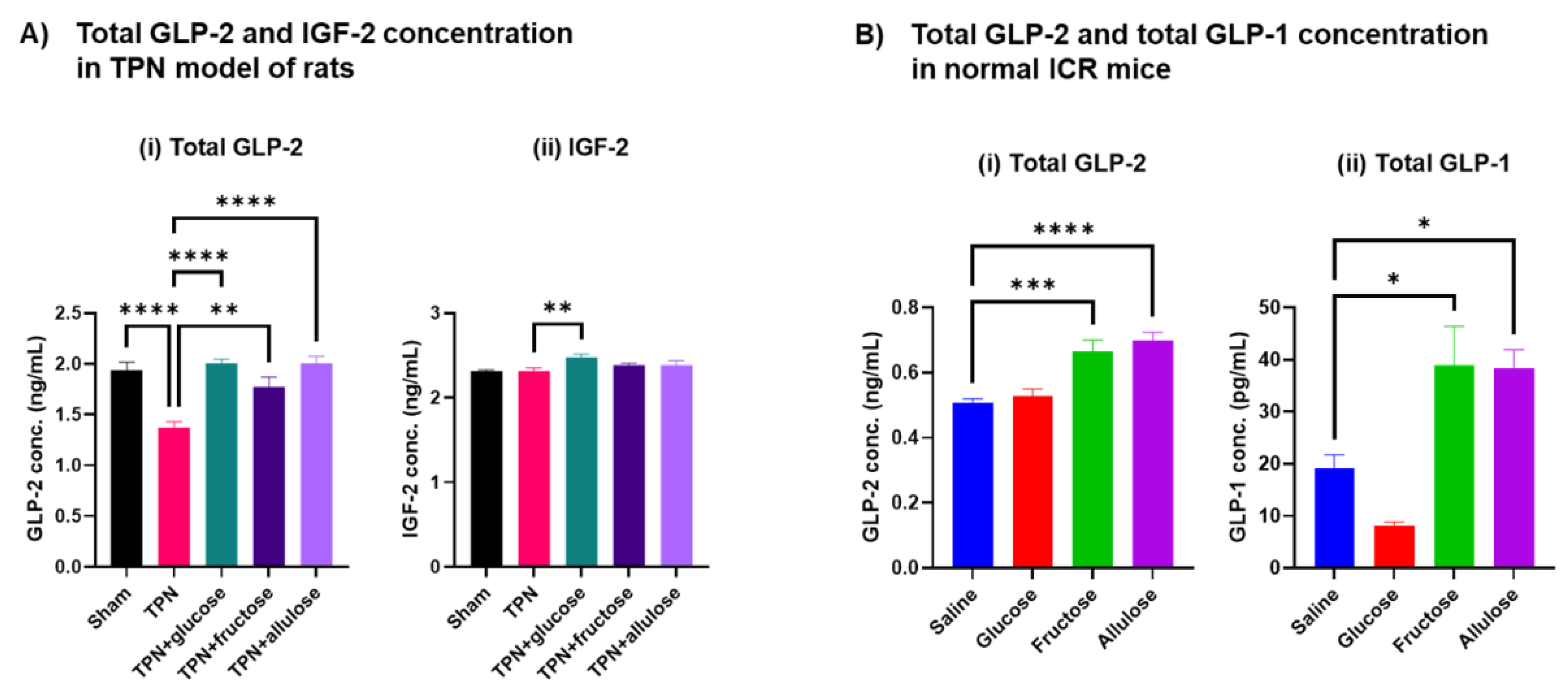
3.6. Effect of Allulose on Hormones Regulating Gut Differentiation and Barrier
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Oshima:, H.; Kimura, I.; Izumori, K. Psicose contents in various food products and its origin. Food Sci. Technol. Res. 2006, 12, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.A.; Kitagaki, S.; Nakano, D.; Nishiyama, A.; Funamoto, Y.; Matsunaga, T.; Tsukamoto, I.; Yamaguchi, F.; Kamitori, K.; Dong, Y.; et al. Rare sugar D-psicose improves insulin sensitivity and glucose tolerance in type 2 diabetes Otsuka Long-Evans Tokushima Fatty (OLETF) rats. Biochem. Biophys. Res. Commun. 2011, 405, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.; Yamaguchi, F.; Matsunaga, T.; Hirata, Y.; Kamitori, K.; Dong, Y.; Sui, L.; Tsukamoto, I.; Ueno, M.; Tokuda, M. Rare sugar D-psicose protects pancreas beta-islets and thus improves insulin resistance in OLETF rats. Biochem. Biophys. Res. Commun. 2012, 425, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.; Yamaguchi, F.; Hirose, K.; Matsunaga, T.; Sui, L.; Hirata, Y.; Noguchi, C.; Katagi, A.; Kamitori, K.; Dong, Y.; et al. Rare sugar D-psicose prevents progression and development of diabetes in T2DM model Otsuka Long-Evans Tokushima Fatty rats. Drug Des. Devel. Ther. 2015, 9, 525–535. [Google Scholar] [CrossRef] [Green Version]
- Hayakawa, M.; Hira, T.; Nakamura, M.; Iida, T.; Kishimoto, Y.; Hara, H. Secretion of GLP-1 but not GIP is potently stimulated by luminal d-Allulose (d-Psicose) in rats. Biochem. Biophys. Res. Commun. 2018, 496, 898–903. [Google Scholar] [CrossRef]
- Drucker, D.J. The biology of incretin hormones. Cell Metab. 2006, 3, 153–165. [Google Scholar] [CrossRef] [Green Version]
- Iwasaki, Y.; Sendo, M.; Dezaki, K.; Hira, T.; Sato, T.; Nakata, M.; Goswami, C.; Aoki, R.; Arai, T.; Kumari, P.; et al. GLP-1 release and vagal afferent activation mediate the beneficial metabolic and chronotherapeutic effects of D-allulose. Nat. Commun. 2018, 9, 113. [Google Scholar] [CrossRef]
- Itoh, K.; Mizuno, S.; Hama, S.; Oshima, W.; Kawamata, M.; Hossain, A.; Ishihara, Y.; Tokuda, M. Beneficial Effects of Supplementation of the Rare Sugar “D-allulose” Against Hepatic Steatosis and Severe Obesity in Lep(ob)/Lep(ob) Mice. J. Food Sci. 2015, 80, H1619–H1626. [Google Scholar] [CrossRef]
- Nagata, Y.; Kanasaki, A.; Tamaru, S.; Tanaka, K. D-psicose, an epimer of D-fructose, favorably alters lipid metabolism in Sprague-Dawley rats. J. Agric. Food Chem. 2015, 63, 3168–3176. [Google Scholar] [CrossRef]
- Choi, B.R.; Kwon, E.Y.; Kim, H.J.; Choi, M.S. Role of Synbiotics Containing d-Allulose in the Alteration of Body Fat and Hepatic Lipids in Diet-Induced Obese Mice. Nutrients 2018, 10, 1797. [Google Scholar] [CrossRef] [Green Version]
- Yagi, K.; Matsuo, T. The study on long-term toxicity of d-psicose in rats. J. Clin. Biochem. Nutr. 2009, 45, 271–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iida, T.; Hayashi, N.; Yamada, T.; Yoshikawa, Y.; Miyazato, S.; Kishimoto, Y.; Okuma, K.; Tokuda, M.; Izumori, K. Failure of d-psicose absorbed in the small intestine to metabolize into energy and its low large intestinal fermentability in humans. Metabolism 2010, 59, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Hishiike, T.; Ogawa, M.; Hayakawa, S.; Nakajima, D.; O’Charoen, S.; Ooshima, H.; Sun, Y. Transepithelial transports of rare sugar D-psicose in human intestine. J. Agric. Food Chem. 2013, 61, 7381–7386. [Google Scholar] [CrossRef] [PubMed]
- Kishida, K.; Martinez, G.; Iida, T.; Yamada, T.; Ferraris, R.P.; Toyoda, Y. d-Allulose is a substrate of glucose transporter type 5 (GLUT5) in the small intestine. Food Chem. 2019, 277, 604–608. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Yoon, J.; Choi, M.S. Tracing the Anti-Inflammatory Mechanism/Triggers of d-Allulose: A Profile Study of Microbiome Composition and mRNA Expression in Diet-Induced Obese Mice. Mol. Nutr. Food Res. 2020, 64, e1900982. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Han, H.J.; Kim, A.H.; Choi, J.Y.; Cho, S.J.; Park, Y.B.; Jung, U.J.; Choi, M.S. d-Allulose supplementation normalized the body weight and fat-pad mass in diet-induced obese mice via the regulation of lipid metabolism under isocaloric fed condition. Mol. Nutr. Food Res. 2016, 60, 1695–1706. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.; Yamaguchi, F.; Matsuo, T.; Tsukamoto, I.; Toyoda, Y.; Ogawa, M.; Nagata, Y.; Tokuda, M. Rare sugar D-allulose: Potential role and therapeutic monitoring in maintaining obesity and type 2 diabetes mellitus. Pharmacol. Ther. 2015, 155, 49–59. [Google Scholar] [CrossRef]
- Goldstein, R.M.; Hebiguchi, T.; Luk, G.D.; Taqi, F.; Guilarte, T.R.; Franklin, F.A., Jr.; Niemiec, P.W.; Dudgeon, D.L. The effects of total parenteral nutrition on gastrointestinal growth and development. J. Pediatr. Surg. 1985, 20, 785–791. [Google Scholar] [CrossRef]
- Suzuki, T.; Mayamagi, Y.; Keta, A.; Kasahara, A.; Sato, A.; Takahashi, T. Oral administration of fructose improves jejunal villous morphology and nutrient digestion and absorption capabilities in a rat model of total parenteral nutrition. Biomed. Res. Clin. Pract. 2019, 4, 1–8. [Google Scholar] [CrossRef]
- Jiang, L.; Ferraris, R.P. Developmental reprogramming of rat GLUT-5 requires de novo mRNA and protein synthesis. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 280, G113–G120. [Google Scholar] [CrossRef] [Green Version]
- Dahlqvist, A. Assay of intestinal disaccharidases. Scand. J. Clin. Lab. Investig. 1984, 44, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Aoki, K.; Shimokobe, K.; Omiya, S.; Funayama, C.; Takahashi, T.; Kato, M. Age-related morphological and functional changes in the small intestine of senescence-accelerated mouse. Exp. Gerontol. 2022, 163, 111795. [Google Scholar] [CrossRef] [PubMed]
- Lorentz, O.; Duluc, I.; Arcangelis, A.D.; Simon-Assmann, P.; Kedinger, M.; Freund, J.N. Key role of the Cdx2 homeobox gene in extracellular matrix-mediated intestinal cell differentiation. J. Cell Biol. 1997, 139, 1553–1565. [Google Scholar] [CrossRef] [Green Version]
- Douard, V.; Cui, X.L.; Soteropoulos, P.; Ferraris, R.P. Dexamethasone sensitizes the neonatal intestine to fructose induction of intestinal fructose transporter (Slc2A5) function. Endocrinology 2008, 149, 409–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T.; Douard, V.; Mochizuki, K.; Goda, T.; Ferraris, R.P. Diet-induced epigenetic regulation in vivo of the intestinal fructose transporter Glut5 during development of rat small intestine. Biochem. J. 2011, 435, 43–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudley, M.A.; Wykes, L.J.; Dudley, A.W., Jr.; Burrin, D.G.; Nichols, B.L.; Rosenberger, J.; Jahoor, F.; Heird, W.C.; Reeds, P.J. Parenteral nutrition selectively decreases protein synthesis in the small intestine. Am. J. Physiol. 1998, 274, G131–G137. [Google Scholar] [CrossRef] [PubMed]
- Herman, M.A.; Birnbaum, M.J. Molecular aspects of fructose metabolism and metabolic disease. Cell Metab. 2021, 33, 2329–2354. [Google Scholar] [CrossRef]
- Tsukamoto, I.; Hossain, A.; Yamaguchi, F.; Hirata, Y.; Dong, Y.; Kamitori, K.; Sui, L.; Nonaka, M.; Ueno, M.; Nishimoto, K.; et al. Intestinal absorption, organ distribution, and urinary excretion of the rare sugar D-psicose. Drug Des. Devel. Ther. 2014, 8, 1955–1964. [Google Scholar] [CrossRef] [Green Version]
- Shu, R.; David, E.S.; Ferraris, R.P. Dietary fructose enhances intestinal fructose transport and GLUT5 expression in weaning rats. Am. J. Physiol. 1997, 272, G446–G453. [Google Scholar] [CrossRef]
- Kishi, K.; Tanaka, T.; Igawa, M.; Takase, S.; Goda, T. Sucrase-isomaltase and hexose transporter gene expressions are coordinately enhanced by dietary fructose in rat jejunum. J. Nutr. 1999, 129, 953–956. [Google Scholar] [CrossRef]
- Cui, X.L.; Soteropoulos, P.; Tolias, P.; Ferraris, R.P. Fructose-responsive genes in the small intestine of neonatal rats. Physiol. Genom. 2004, 18, 206–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, S.R.; Ramsamooj, S.; Liang, R.J.; Katti, A.; Pozovskiy, R.; Vasan, N.; Hwang, S.K.; Nahiyaan, N.; Francoeur, N.J.; Schatoff, E.M.; et al. Dietary fructose improves intestinal cell survival and nutrient absorption. Nature 2021, 597, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Ding, Y.; Xu, C.; Hao, M.; Li, H.; Ding, L. Cldn-7 deficiency promotes experimental colitis and associated carcinogenesis by regulating intestinal epithelial integrity. Oncoimmunology 2021, 10, 1923910. [Google Scholar] [CrossRef] [PubMed]
- Brubaker, P.L.; Crivici, A.; Izzo, A.; Ehrlich, P.; Tsai, C.H.; Drucker, D.J. Circulating and tissue forms of the intestinal growth factor, glucagon-like peptide-2. Endocrinology 1997, 138, 4837–4843. [Google Scholar] [CrossRef]
- Xiao, Q.; Boushey, R.P.; Drucker, D.J.; Brubaker, P.L. Secretion of the intestinotropic hormone glucagon-like peptide 2 is differentially regulated by nutrients in humans. Gastroenterology 1999, 117, 99–105. [Google Scholar] [CrossRef]
- Martin, G.R.; Wallace, L.E.; Hartmann, B.; Holst, J.J.; Demchyshyn, L.; Toney, K.; Sigalet, D.L. Nutrient-stimulated GLP-2 release and crypt cell proliferation in experimental short bowel syndrome. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, G431–G438. [Google Scholar] [CrossRef] [Green Version]
- Guan, X.; Karpen, H.E.; Stephens, J.; Bukowski, J.T.; Niu, S.; Zhang, G.; Stoll, B.; Finegold, M.J.; Holst, J.J.; Hadsell, D.; et al. GLP-2 receptor localizes to enteric neurons and endocrine cells expressing vasoactive peptides and mediates increased blood flow. Gastroenterology 2006, 130, 150–164. [Google Scholar] [CrossRef]
- Leen, J.L.; Izzo, A.; Upadhyay, C.; Rowland, K.J.; Dube, P.E.; Gu, S.; Heximer, S.P.; Rhodes, C.J.; Storm, D.R.; Lund, P.K.; et al. Mechanism of action of glucagon-like peptide-2 to increase IGF-I mRNA in intestinal subepithelial fibroblasts. Endocrinology 2011, 152, 436–446. [Google Scholar] [CrossRef] [Green Version]
- Cheeseman, C.I. Upregulation of SGLT-1 transport activity in rat jejunum induced by GLP-2 infusion in vivo. Am. J. Physiol. 1997, 273, R1965–R1971. [Google Scholar] [CrossRef] [Green Version]
- Kato, Y.; Yu, D.; Schwartz, M.Z. Glucagonlike peptide-2 enhances small intestinal absorptive function and mucosal mass in vivo. J. Pediatr. Surg. 1999, 34, 18–20. [Google Scholar] [CrossRef]
- Damholt, A.B.; Buchan, A.M.; Holst, J.J.; Kofod, H. Proglucagon processing profile in canine L cells expressing endogenous prohormone convertase 1/3 and prohormone convertase 2. Endocrinology 1999, 140, 4800–4808. [Google Scholar] [CrossRef] [PubMed]
- Choi, P.; Guo, J.; Erwin, C.R.; Warner, B.W. IGF-2 mediates intestinal mucosal hyperplasia in retinoblastoma protein (Rb)-deficient mice. J. Pediatr. Surg. 2013, 48, 1340–1347. [Google Scholar] [CrossRef] [Green Version]
- Dube, P.E.; Forse, C.L.; Bahrami, J.; Brubaker, P.L. The essential role of insulin-like growth factor-1 in the intestinal tropic effects of glucagon-like peptide-2 in mice. Gastroenterology 2006, 131, 589–605. [Google Scholar] [CrossRef]
- Markovic, M.A.; Brubaker, P.L. The roles of glucagon-like peptide-2 and the intestinal epithelial insulin-like growth factor-1 receptor in regulating microvillus length. Sci. Rep. 2019, 9, 13010. [Google Scholar] [CrossRef] [PubMed]
- Fesler, Z.; Mitova, E.; Brubaker, P.L. GLP-2, EGF, and the Intestinal Epithelial IGF-1 Receptor Interactions in the Regulation of Crypt Cell Proliferation. Endocrinology 2020, 161, bqaa040. [Google Scholar] [CrossRef] [PubMed]
- Murali, S.G.; Brinkman, A.S.; Solverson, P.; Pun, W.; Pintar, J.E.; Ney, D.M. Exogenous GLP-2 and IGF-I induce a differential intestinal response in IGF binding protein-3 and -5 double knockout mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G794–G804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austin, K.; Imam, N.A.; Pintar, J.E.; Brubaker, P.L. IGF binding protein-4 is required for the growth effects of glucagon-like peptide-2 in murine intestine. Endocrinology 2015, 156, 429–436. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, J.; Longuet, C.; Maida, A.; Bahrami, J.; Xu, E.; Baker, C.L.; Brubaker, P.L.; Drucker, D.J.; Adeli, K. Glucagon-like peptide-2 increases intestinal lipid absorption and chylomicron production via CD36. Gastroenterology 2009, 137, 997–1005.e4. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | |||||
| Categories | Diseases or Functions Annotation | Activation z-Score | #Molecules | ||
| F/G | A/G | A/F | |||
| Cancer, Organismal Injury and Abnormalities | Advanced extracranial solid tumor | 2.13 | 100 | ||
| Advanced malignant tumor | 2.13 | 174 | |||
| Secondary tumor | 2.13 | 163 | |||
| Advanced lung cancer | 2.13 | 51 | |||
| Carbohydrate Metabolism | Uptake of carbohydrate | 2.92 | 2.42 | 56 | |
| Uptake of monosaccharide | 2.43 | 52 | |||
| Carbohydrate Metabolism, Lipid Metabolism, Small Molecule Biochemistry | Metabolism of phosphatidic acid | 2.41 | 41 | ||
| Synthesis of phosphatidic acid | 2.00 | 37 | |||
| Carbohydrate Metabolism, Molecular Transport, Small Molecule Biochemistry | Uptake of D-glucose | 2.47 | 2.47 | 47 | |
| Uptake of D-hexose | 2.59 | 2.32 | 48 | ||
| Cell Death and Survival, Organismal Injury and Abnormalities | Cell death of cervical cancer cell lines | 2.40 | 2.21 | 71 | |
| Cell death of epithelial cell lines | 2.15 | 55 | |||
| Cell Morphology, Cellular Assembly and Organization, Cellular Function and Maintenance | Formation of cellular protrusions | 2.55 | 2.82 | 142 | |
| Cell-To-Cell Signaling and Interaction | Adhesion of tumor cell lines | 2.29 | 2.63 | 55 | |
| Binding of tumor cell lines | 3.08 | 3.39 | 72 | ||
| Interaction of tumor cell lines | 2.70 | 3.26 | 76 | ||
| Cellular Assembly and Organization, Cellular Function, Maintenance and Tissue Development | Development of cytoplasm | 2.48 | 2.70 | 96 | |
| Formation of cytoskeleton | 2.25 | 2.49 | 67 | ||
| Formation of membrane ruffles | 2.40 | 17 | |||
| Microtubule dynamics | 2.10 | 182 | |||
| Formation of actin filaments | 2.28 | 2.53 | 57 | ||
| Formation of actin stress fibers | 2.01 | 47 | |||
| Fibrogenesis | 2.42 | 2.20 | 79 | ||
| Formation of filaments | 2.35 | 2.41 | 72 | ||
| Cellular Compromise, Inflammatory Response | Degranulation of cells | 2.03 | 2.42 | 108 | |
| Degranulation of phagocytes | 2.27 | 2.27 | 96 | ||
| Cellular Function and Maintenance | Endocytosis | 3.36 | 3.36 | 94 | |
| Engulfment of cells | 3.41 | 3.41 | 83 | ||
| Internalization by tumor cell lines | 2.41 | 2.74 | 27 | ||
| Cellular Function and Maintenance, Inflammatory Response | Phagocytosis | 3.63 | 3.63 | 58 | |
| Infectious Diseases | HIV infection | 2.07 | 120 | ||
| Infection by HIV-1 | 2.03 | 104 | |||
| Infection by Retroviridae | 2.22 | 2.37 | 124 | ||
| Infection by RNA virus | 2.43 | 2.57 | 155 | ||
| Infection of cells | 2.26 | 2.40 | 140 | ||
| Infection of cervical cancer cell lines | 2.50 | 2.69 | 78 | ||
| Infection of tumor cell lines | 2.13 | 2.84 | 3.02 | 92 | |
| Viral Infection | 2.56 | 2.63 | 294 | ||
| Lipid Metabolism, Small Molecule Biochemistry | Fatty acid metabolism | 2.05 | 112 | ||
| Metabolism of membrane lipid derivative | 2.64 | 3.17 | 99 | ||
| Molecular Transport | Secretion of molecule | 2.94 | 2.74 | 103 | |
| Transport of molecule | 4.48 | 4.17 | 305 | ||
| Organismal Development | Size of body | 2.90 | 2.90 | 131 | |
| Post-Translational Modification | Ubiquitination | 2.34 | 70 | ||
| Ubiquitination of protein | 2.01 | 2.01 | 69 | ||
| Protein Trafficking | Interaction of protein | 2.24 | 2.24 | 48 | |
| B | |||||
| Categories | Diseases or Functions Annotation | Activation z-Score | Molecules | ||
| F/G | A/G | A/F | |||
| Cardiovascular Disease, Hematological Disease, Organismal Injury and Abnormalities | Anemia | −2.49 | −2.49 | 74 | |
| Cell Cycle, Cellular Movement | Cytokinesis of tumor cell lines | −2.05 | −2.05 | 17 | |
| Cell Death and Survival | Apoptosis of breast cancer cell lines | −2.03 | 61 | ||
| Cellular Compromise | Stress response of cells | −2.02 | 28 | ||
| Stress response of tumor cell lines | −2.58 | −2.05 | −2.05 | 15 | |
| Cellular Development, Cellular Growth and Proliferation, Connective Tissue Development and Function, Tissue Development | Cell proliferation of fibroblasts | −3.09 | 72 | ||
| Connective Tissue Disorders, Inflammatory Disease, Inflammatory Response, Organismal Injury and Abnormalities, Skeletal and Muscular Disorders | Inflammation of joint | −2.39 | −2.39 | 153 | |
| Connective Tissue Disorders, Inflammatory Disease, Organismal Injury and Abnormalities, Skeletal and Muscular Disorders | Rheumatic Disease | −2.21 | −2.21 | 199 | |
| Free Radical Scavenging | Generation of reactive oxygen species | −2.05 | 42 | ||
| Inflammatory Response | Inflammation of absolute anatomical region | −2.23 | −2.23 | 181 | |
| Inflammation of body cavity | −3.05 | −3.05 | 152 | ||
| Inflammatory Response, Organismal Injury and Abnormalities | Inflammation of organ | −2.43 | −2.43 | 219 | |
| Lipid Metabolism, Molecular Transport, Small Molecule Biochemistry | Concentration of fatty acid | −2.5 | −2.12 | 52 | |
| Neurological Disease | Motor dysfunction or movement disorder | −2.13 | −2.53 | 185 | |
| Movement Disorders | −2.04 | −2.45 | 182 | ||
| Organismal Injury and Abnormalities, Renal and Urological Disease | Urination disorder | −3.24 | −3.55 | 54 | |
| Organismal Survival | Morbidity or mortality | −3.12 | −3.12 | 402 | |
| Organismal death | −3.19 | −3.19 | 396 | ||
| Protein Synthesis | Expression of protein | −2.57 | −2.57 | 80 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, T.; Sato, Y.; Kadoya, S.; Takahashi, T.; Otomo, M.; Kobayashi, H.; Aoki, K.; Kantake, M.; Sugiyama, M.; Ferraris, R.P. Comparative Effects of Allulose, Fructose, and Glucose on the Small Intestine. Nutrients 2022, 14, 3230. https://doi.org/10.3390/nu14153230
Suzuki T, Sato Y, Kadoya S, Takahashi T, Otomo M, Kobayashi H, Aoki K, Kantake M, Sugiyama M, Ferraris RP. Comparative Effects of Allulose, Fructose, and Glucose on the Small Intestine. Nutrients. 2022; 14(15):3230. https://doi.org/10.3390/nu14153230
Chicago/Turabian StyleSuzuki, Takuji, Yuki Sato, Sumire Kadoya, Takumi Takahashi, Moeko Otomo, Hanna Kobayashi, Kai Aoki, Mai Kantake, Maika Sugiyama, and Ronaldo P. Ferraris. 2022. "Comparative Effects of Allulose, Fructose, and Glucose on the Small Intestine" Nutrients 14, no. 15: 3230. https://doi.org/10.3390/nu14153230
APA StyleSuzuki, T., Sato, Y., Kadoya, S., Takahashi, T., Otomo, M., Kobayashi, H., Aoki, K., Kantake, M., Sugiyama, M., & Ferraris, R. P. (2022). Comparative Effects of Allulose, Fructose, and Glucose on the Small Intestine. Nutrients, 14(15), 3230. https://doi.org/10.3390/nu14153230