
Hypothesis: Low Vitamin A and D Levels Worsen Clinical Outcomes When Children with Sickle Cell Disease Encounter Parvovirus B19

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participant Selection and Clinical Data Collection
2.2. Cytokine/Chemokine Assays
2.3. Virus-Specific Enzyme-Linked Immunosorbent Assay (ELISA)
2.4. Vitamin Measurements
2.5. Statistical Analyses
3. Results
3.1. Patient Characteristics
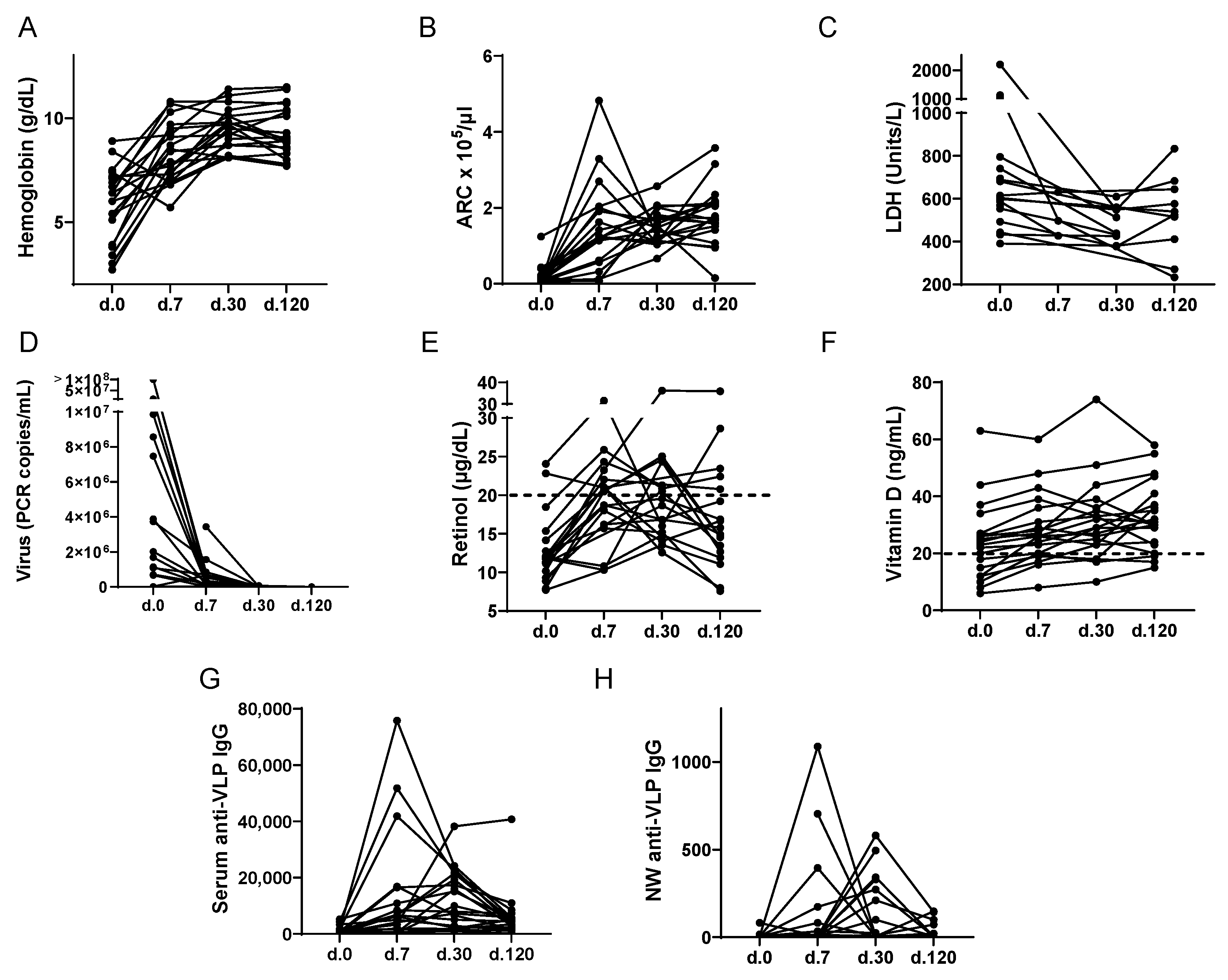
3.2. Laboratory Kinetics during Acute Parvovirus B19 Infection
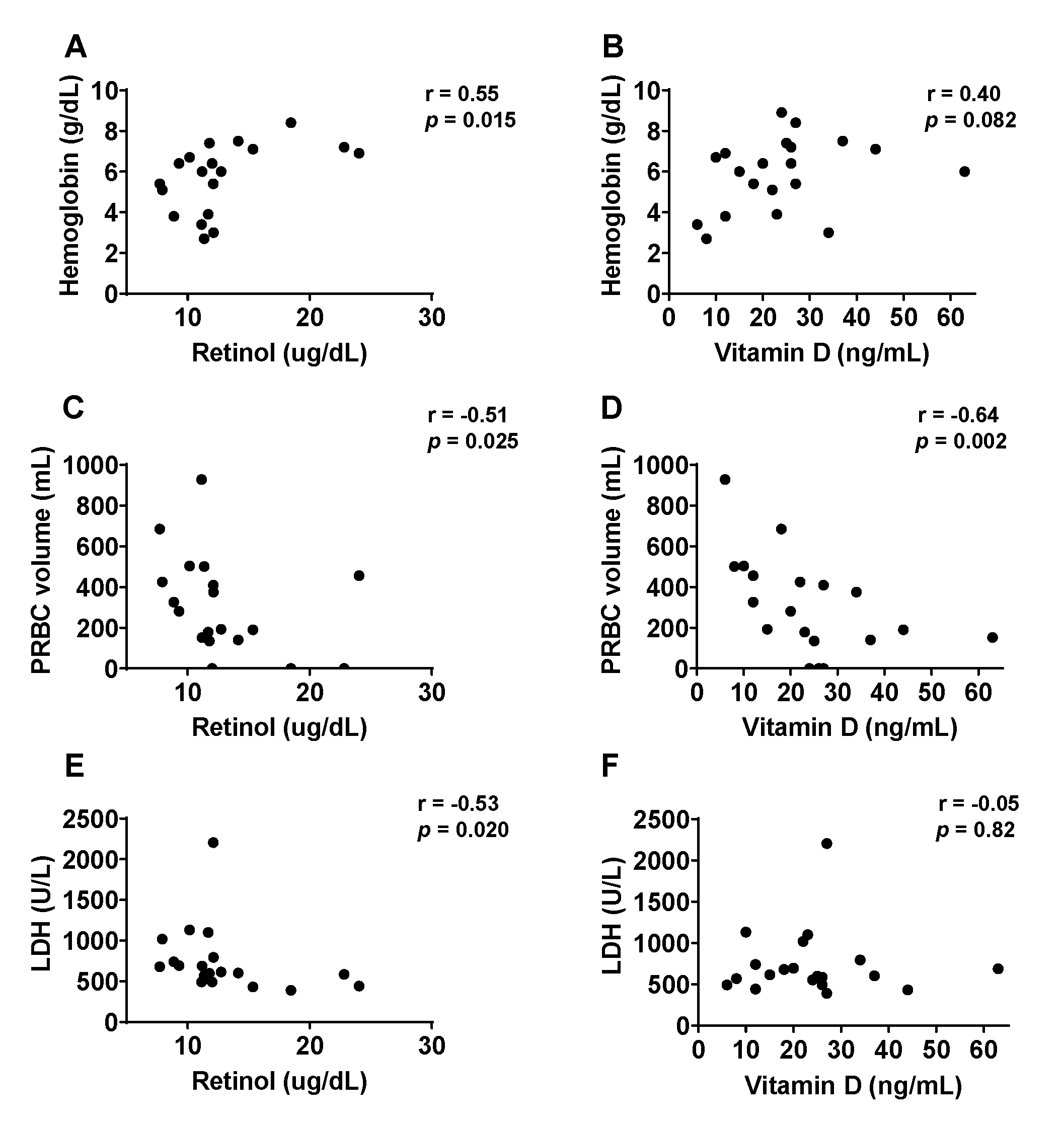
3.3. Correlation between Vitamin Levels and Disease Symptoms upon Hospitalization
3.4. Vitamins, Virus, ARC, and the Virus-Specific Antibody Response
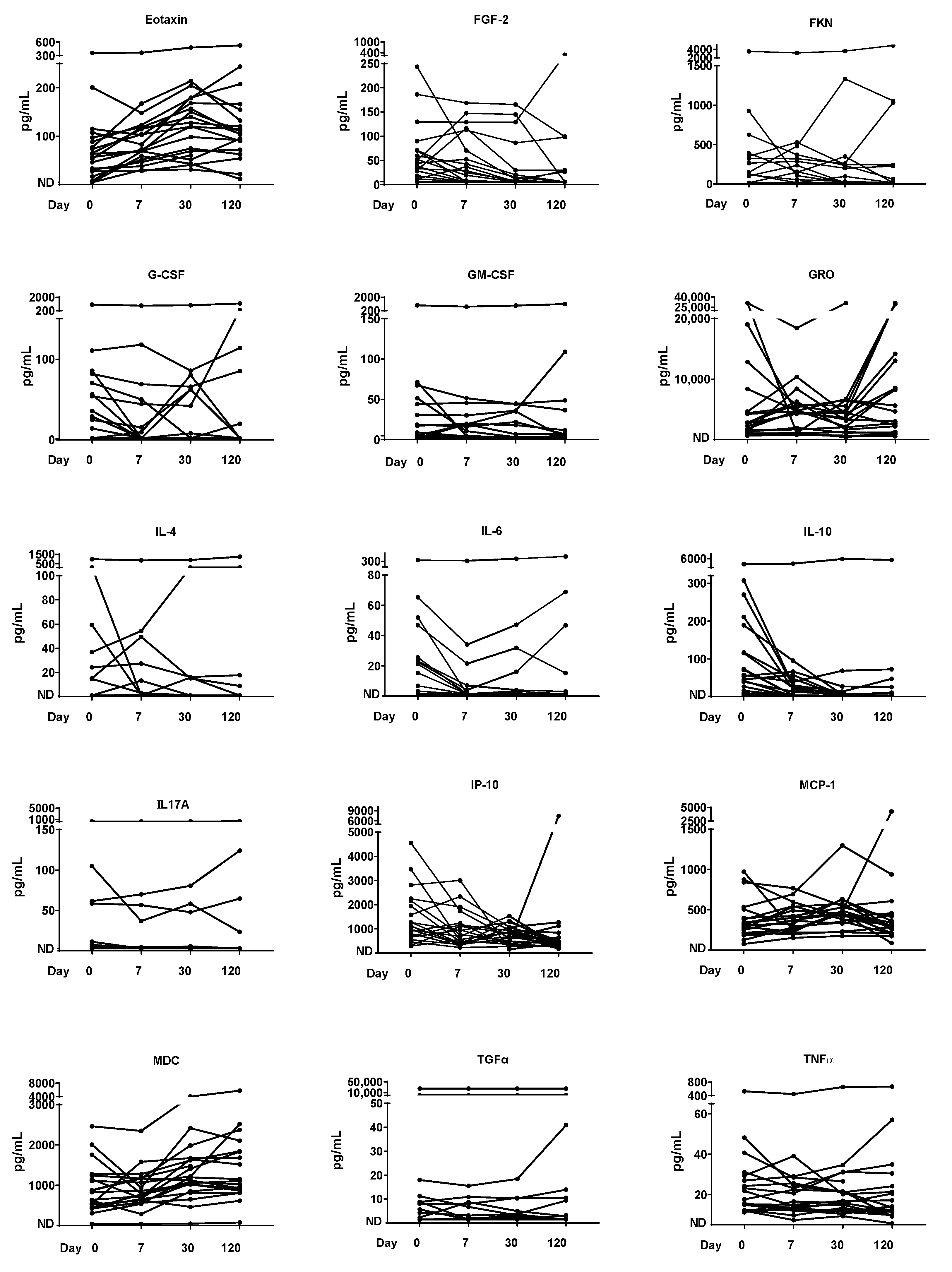
3.5. Cytokine/Chemokine Time Course
3.6. Vitamin Correlations with Cytokines/Chemokines
3.7. Virus, ARC and Virus-Specific Antibody Correlations with Cytokines/Chemokines
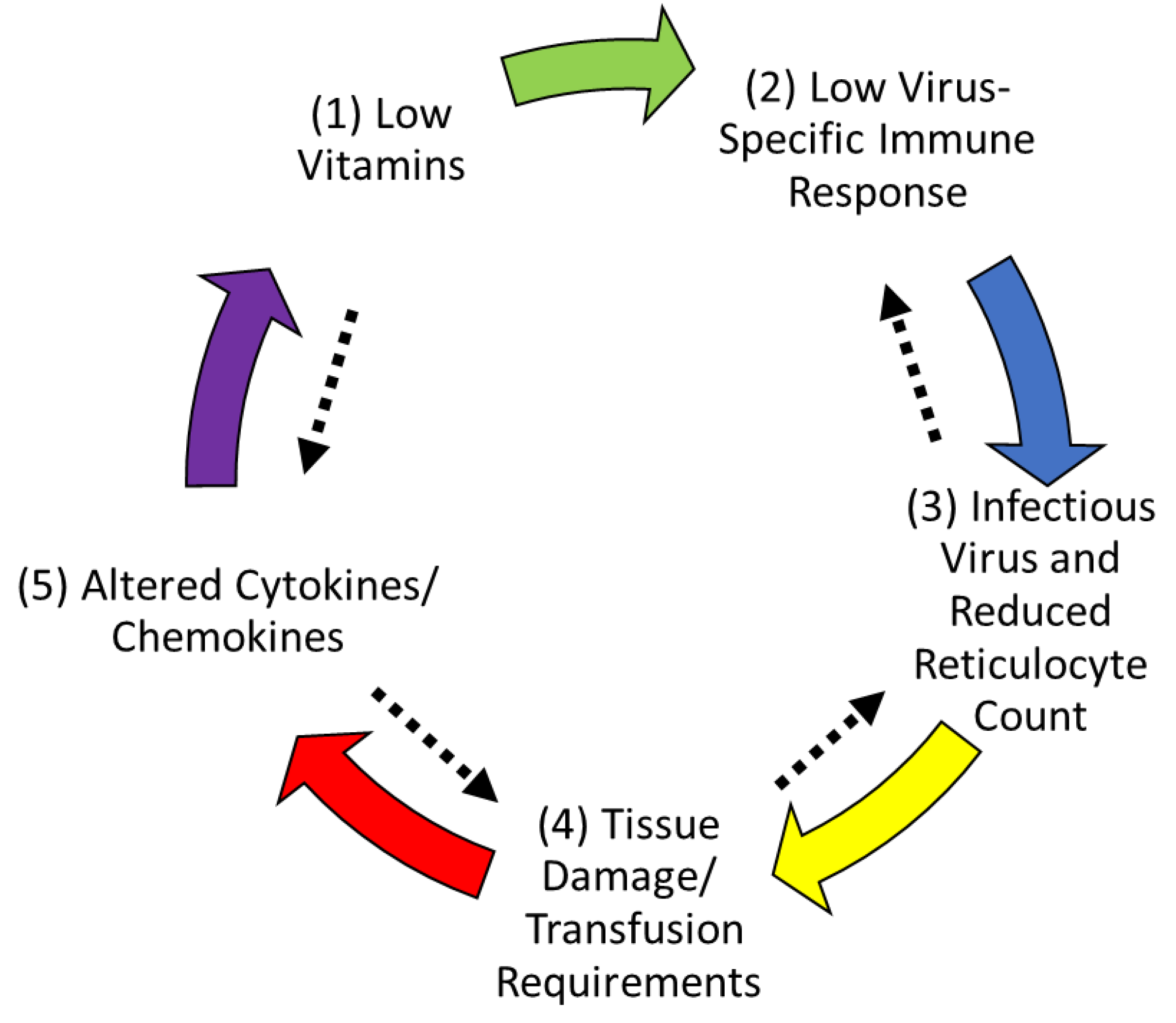
3.8. Hypothesis
4. Discussion
4.1. Low Vitamins Inhibit Virus-Specific Immune Responses and Thereby Increase Virus Amplification and Disease
4.2. Disease Further Reduces Vitamin Levels
4.2.1. RBP Is an Acute Phase Protein
4.2.2. Renal Mechanisms of Hypovitaminosis
4.3. Pediatric Under-Nutrition and Vitamin Supplementation
4.3.1. Under-Nutrition among Children with SCD
4.3.2. Benefits/Risks of Vitamin Supplementation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Young, N.S.; Brown, K.E. Parvovirus B19. N. Engl. J. Med. 2004, 350, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, J.F., Jr.; Luban, N.L.; Cohen, B.J.; Mortimer, P.P. Human serum parvovirus as the cause of aplastic crisis in sickle cell disease. Am. J. Dis. Child. 1984, 138, 401–403. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.P.; Miller, S.T.; Cohen, B.J. Transient aplastic crisis in patients with sickle cell disease. B19 parvovirus studies during a 7-year period. Am. J. Dis. Child. 1992, 146, 1328–1330. [Google Scholar] [CrossRef] [PubMed]
- Yazdanbakhsh, K.; Ware, R.E.; Noizat-Pirenne, F. Red blood cell alloimmunization in sickle cell disease: Pathophysiology, risk factors, and transfusion management. Blood 2012, 120, 528–537. [Google Scholar] [CrossRef]
- Tanumihardjo, S.A.; Russell, R.M.; Stephensen, C.B.; Gannon, B.M.; Craft, N.E.; Haskell, M.J.; Lietz, G.; Schulze, K.; Raiten, D.J. Biomarkers of Nutrition for Development (BOND)—Vitamin A Review. J. Nutr. 2016, 146, 1816S–1848S. [Google Scholar] [CrossRef]
- Chesney, R.W. Vitamin D and the Magic Mountain: The Anti-Infectious Role of the Vitamin. J. Pediatr. 2010, 156, 698–703. [Google Scholar] [CrossRef]
- Penkert, R.R.; Cortez, V.; Karlsson, E.A.; Livingston, B.; Surman, S.L.; Li, Y.; Ross, A.C.; Schultz-Cherry, S.; Hurwitz, J.L. Vitamin A Corrects Tissue Deficits in Diet-Induced Obese Mice and Reduces Influenza Infection After Vaccination and Challenge. Obesity 2020, 28, 1631–1636. [Google Scholar] [CrossRef]
- Penkert, R.R.; Rowe, H.M.; Surman, S.L.; Sealy, R.E.; Rosch, J.; Hurwitz, J.L. Influences of Vitamin A on Vaccine Immunogenicity and Efficacy. Front. Immunol. 2019, 10, 1576. [Google Scholar] [CrossRef]
- Rudraraju, R.; Jones, B.G.; Surman, S.L.; Sealy, R.E.; Thomas, P.G.; Hurwitz, J.L. Respiratory Tract Epithelial Cells Express Retinaldehyde Dehydrogenase ALDH1A and Enhance IgA Production by Stimulated B Cells in the Presence of Vitamin A. PLoS ONE 2014, 9, e86554. [Google Scholar] [CrossRef]
- Rudraraju, R.; Surman, S.L.; Jones, B.G.; Sealy, R.; Woodland, D.L.; Hurwitz, J.L. Reduced Frequencies and Heightened CD103 Expression among Virus-Induced CD8+T Cells in the Respiratory Tract Airways of Vitamin A-Deficient Mice. Clin. Vaccine Immunol. 2012, 19, 757–765. [Google Scholar] [CrossRef]
- Surman, S.; Jones, B.; Sealy, R.; Rudraraju, R.; Hurwitz, J. Oral retinyl palmitate or retinoic acid corrects mucosal IgA responses toward an intranasal influenza virus vaccine in vitamin A deficient mice. Vaccine 2014, 32, 2521–2524. [Google Scholar] [CrossRef] [PubMed]
- Surman, S.L.; Penkert, R.R.; Jones, B.G.; Sealy, R.E.; Hurwitz, J.L. Vitamin Supplementation at the Time of Immunization with a Cold-Adapted Influenza Virus Vaccine Corrects Poor Mucosal Antibody Responses in Mice Deficient for Vitamins A and D. Clin. Vaccine Immunol. 2016, 23, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Surman, S.L.; Penkert, R.R.; Sealy, R.E.; Jones, B.G.; Marion, T.N.; Vogel, P.; Hurwitz, J.L. Consequences of Vitamin A Deficiency: Immunoglobulin Dysregulation, Squamous Cell Metaplasia, Infectious Disease, and Death. Int. J. Mol. Sci. 2020, 21, 5570. [Google Scholar] [CrossRef] [PubMed]
- Mora, J.R.; Iwata, M.; von Andrian, U.H. Vitamin effects on the immune system: Vitamins A and D take centre stage. Nat. Rev. Immunol. 2008, 8, 685–698. [Google Scholar] [CrossRef]
- Tejon, G.P.; Manríquez, V.; De Calisto, J.; Flores-Santibañez, F.; Hidalgo, Y.; Crisóstomo, N.; Fernandez, D.; Sauma, D.; Mora, J.R.; Bono, M.R.; et al. Vitamin A Impairs the Reprogramming of Tregs into IL-17-Producing Cells during Intestinal Inflammation. BioMed Res. Int. 2015, 2015, 137893. [Google Scholar] [CrossRef]
- Gembillo, G.; Siligato, R.; Amatruda, M.; Conti, G.; Santoro, D. Vitamin D and Glomerulonephritis. Medicina 2021, 57, 186. [Google Scholar] [CrossRef]
- D’Souza, R.M. Vitamin A for the Treatment of Children with Measles—A Systematic Review. J. Trop. Pediatr. 2002, 48, 323–327. [Google Scholar] [CrossRef]
- Sabetta, J.R.; DePetrillo, P.; Cipriani, R.J.; Smardin, J.; Burns, L.A.; Landry, M.L. Serum 25-Hydroxyvitamin D and the Incidence of Acute Viral Respiratory Tract Infections in Healthy Adults. PLoS ONE 2010, 5, e11088. [Google Scholar] [CrossRef]
- Patel, N.; Penkert, R.R.; Jones, B.G.; Sealy, R.E.; Surman, S.L.; Sun, Y.; Tang, L.; DeBeauchamp, J.; Webb, A.; Richardson, J.; et al. Baseline Serum Vitamin A and D Levels Determine Benefit of Oral Vitamin A&D Supplements to Humoral Immune Responses Following Pediatric Influenza Vaccination. Viruses 2019, 11, 907. [Google Scholar] [CrossRef]
- Hurwitz, J.L.; Jones, B.G.; Penkert, R.R.; Gansebom, S.; Sun, Y.; Tang, L.; Bramley, A.M.; Jain, S.; McCullers, J.A.; Arnold, S.R. Low Retinol-Binding Protein and Vitamin D Levels Are Associated with Severe Outcomes in Children Hospitalized with Lower Respiratory Tract Infection and Respiratory Syncytial Virus or Human Metapneumovirus Detection. J. Pediatr. 2017, 187, 323–327. [Google Scholar] [CrossRef]
- Adesina, O.O.; Gurney, J.G.; Kang, G.; Villavicencio, M.; Hodges, J.R.; Chemaitilly, W.; Kaste, S.C.; Zemel, B.S.; Hankins, J.S. Height-corrected low bone density associates with severe outcomes in sickle cell disease: SCCRIP cohort study results. Blood Adv. 2019, 3, 1476–1488. [Google Scholar] [CrossRef] [PubMed]
- Brown, B.; Long, K.; Agdere, L.; Kulpa, J.; Zarzoso-Fernandez, S.; Choudhary, D.; Sundarum, R. The association between vitamin D deficiency and hospitalization outcomes in pediatric patients with sickle cell disease. Blood Cells Mol. Dis. 2020, 82, 102415. [Google Scholar] [CrossRef] [PubMed]
- Nolan, V.G.; Nottage, K.A.; Cole, E.W.; Hankins, J.S.; Gurney, J.G. Prevalence of Vitamin D Deficiency in Sickle Cell Disease: A Systematic Review. PLoS ONE 2015, 10, e0119908. [Google Scholar] [CrossRef] [PubMed]
- Arlet, J.-B.; Courbebaisse, M.; Chatellier, G.; Eladari, D.; Souberbielle, J.-C.; Friedlander, G.; de Montalembert, M.; Prié, D.; Pouchot, J.; Ribeil, J.-A. Relationship between vitamin D deficiency and bone fragility in sickle cell disease: A cohort study of 56 adults. Bone 2013, 52, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.T.; Licursi, M.; McMahon, D.J. Vitamin D deficiency and acute vaso-occlusive complications in children with sickle cell disease. Pediatr. Blood Cancer 2015, 62, 643–647. [Google Scholar] [CrossRef] [PubMed]
- Schall, J.I.; Zemel, B.S.; Kawchak, D.A.; Ohene-Frempong, K.; Stallings, V.A. Vitamin A status, hospitalizations, and other outcomes in young children with sickle cell disease. J. Pediatr. 2004, 145, 99–106. [Google Scholar] [CrossRef]
- Penkert, R.; Chandramouli, S.; Dormitzer, P.; Settembre, E.; Sealy, R.; Wong, S.; Young, N.; Sun, Y.; Tang, L.; Cotton, A.; et al. Novel Surrogate Neutralizing Assay Supports Parvovirus B19 Vaccine Development for Children with Sickle Cell Disease. Vaccines 2021, 9, 860. [Google Scholar] [CrossRef]
- Almekinder, J.; Manda, W.; Soko, D.; Lan, Y.; Hoover, D.R.; Semba, R.D. Evaluation of plasma retinol-binding protein as a surrogate measure for plasma retinol concentrations. Scand. J. Clin. Lab. Investig. 2000, 60, 199–204. [Google Scholar] [CrossRef]
- Rosen, C.J.; Abrams, S.; Aloia, J.F.; Brannon, P.M.; Clinton, S.K.; Durazo-Arvizu, R.A.; Gallagher, J.C.; Gallo, R.; Jones, G.; Kovacs, C.; et al. IOM Committee Members Respond to Endocrine Society Vitamin D Guideline. J. Clin. Endocrinol. Metab. 2012, 97, 1146–1152. [Google Scholar] [CrossRef]
- Lewis, C.J.; McDowell, M.A.; Sempos, C.T.; Lewis, K.C.; Yetley, E.A. Relationship between age and serum vitamin A in children aged 4–11 y. Am. J. Clin. Nutr. 1990, 52, 353–360. [Google Scholar] [CrossRef]
- Potter, C.G.; Potter, A.C.; Hatton, C.S.; Chapel, H.M.; Anderson, M.J.; Pattison, J.R.; Tyrrell, D.A.; Higgins, P.G.; Willman, J.S.; Parry, H.F. Variation of erythroid and myeloid precursors in the marrow and peripheral blood of volunteer subjects infected with human parvovirus (B19). J. Clin. Investig. 1987, 79, 1486–1492. [Google Scholar] [CrossRef] [PubMed]
- Penkert, R.R.; Hurwitz, J.L.; Thomas, P.; Rosch, J.; Dowdy, J.; Sun, Y.; Tang, L.; Hankins, J.S. Inflammatory molecule reduction with hydroxyurea therapy in children with sickle cell anemia. Haematologica 2018, 103, e50–e54. [Google Scholar] [CrossRef] [PubMed]
- Garcia, N.P.; Júnior, A.L.S.; Soares, G.A.S.; Costa, T.C.C.; dos Santos, A.P.C.; Costa, A.G.; Tarragô, A.M.; Martins, R.N.; Pontes, F.D.C.L.; de Almeida, E.G.; et al. Sickle Cell Anemia Patients Display an Intricate Cellular and Serum Biomarker Network Highlighted by TCD4+CD69+ Lymphocytes, IL-17/MIP-1β, IL-12/VEGF, and IL-10/IP-10 Axis. J. Immunol. Res. 2020, 2020, 4585704. [Google Scholar] [CrossRef]
- Sennikov, S.V.; Injelevskaya, T.V.; Krysov, S.V.; Silkov, A.N.; Kovinev, I.B.; Dyachkova, N.J.; Zenkov, A.N.; Loseva, M.I.; Kozlov, V.A. Production of hemo- and immunoregulatory cytokines by erythroblast antigen+ and glycophorin A+ cells from human bone marrow. BMC Cell Biol. 2004, 5, 39. [Google Scholar] [CrossRef]
- Oppenheim, J.J.; Zachariae, C.O.C.; Mukaida, N.; Matsushima, K. Properties of the Novel Proinflammatory Supergene “Intercrine” Cytokine Family. Annu. Rev. Immunol. 1991, 9, 617–648. [Google Scholar] [CrossRef]
- Gandrillon, O.; Schmidt, U.; Beug, H.; Samarut, J. TGF-beta cooperates with TGF-alpha to induce the self-renewal of normal erythrocytic progenitors: Evidence for an autocrine mechanism. EMBO J. 1999, 18, 2764–2781. [Google Scholar] [CrossRef]
- Karsten, E.; Breen, E.; Herbert, B. Red blood cells are dynamic reservoirs of cytokines. Sci. Rep. 2018, 8, 3101. [Google Scholar] [CrossRef] [PubMed]
- Honarvar, N.M.; Harirchian, M.H.; Koohdani, F.; Siassi, F.; Abdolahi, M.; Bitarafan, S.; Salehi, E.; Sahraian, M.A.; Eshraghian, M.R.; Saboor-Yarghi, A.A. The Effect of Vitamin A Supplementation on Retinoic Acid-Related Orphan Receptor γt (RORγt) and Interleukin-17 (IL-17) Gene Expression in Avonex-Treated Multiple Sclerotic Patients. J. Mol. Neurosci. 2013, 51, 749–753. [Google Scholar] [CrossRef]
- Williams, A.M.; Ladva, C.; Leon, J.S.; Lopman, B.A.; Tangpricha, V.; Whitehead, R.D.; Armitage, A.E.; Wray, K.; Morovat, A.; Pasricha, S.-R.; et al. Changes in micronutrient and inflammation serum biomarker concentrations after a norovirus human challenge. Am. J. Clin. Nutr. 2019, 110, 1456–1464. [Google Scholar] [CrossRef]
- Penkert, R.R.; Surman, S.L.; Jones, B.G.; Sealy, R.E.; Vogel, P.; Neale, G.; Hurwitz, J.L. Vitamin A deficient mice exhibit increased viral antigens and enhanced cytokine/chemokine production in nasal tissues following respiratory virus infection despite the presence of FoxP3+ T cells. Int. Immunol. 2016, 28, 139–152. [Google Scholar] [CrossRef]
- Penkert, R.R.; Iverson, A.; Rosch, J.W.; Hurwitz, J.L. Prevnar-13 vaccine failure in a mouse model for vitamin A deficiency. Vaccine 2017, 35, 6264–6268. [Google Scholar] [CrossRef] [PubMed]
- Sealy, R.E.; Surman, S.L.; Vogel, P.; Hurwitz, J.L. Might routine vitamin A monitoring in cystic fibrosis patients reduce virus-mediated lung pathology? Front. Immunol. 2021, 12, 704391. [Google Scholar] [CrossRef] [PubMed]
- Penkert, R.R.; Smith, A.P.; Hrincius, E.R.; McCullers, J.A.; Vogel, P.; Smith, A.M.; Hurwitz, J.L. Effect of Vitamin A Deficiency in Dysregulating Immune Responses to Influenza Virus and Increasing Mortality Rates After Bacterial Coinfections. J. Infect. Dis. 2021, 223, 1806–1816. [Google Scholar] [CrossRef] [PubMed]
- Surman, S.L.; Jones, B.G.; Rudraraju, R.; Sealy, R.E.; Hurwitz, J.L. Intranasal Administration of Retinyl Palmitate with a Respiratory Virus Vaccine Corrects Impaired Mucosal IgA Response in the Vitamin A-Deficient Host. Clin. Vaccine Immunol. 2014, 21, 598–601. [Google Scholar] [CrossRef]
- Raiten, D.J.; Ashour, F.A.S.; Ross, A.C.; Meydani, S.N.; Dawson, H.D.; Stephensen, C.B.; Brabin, B.; Suchdev, P.; Van Ommen, B.; Group, I.C. Inflammation and Nutritional Science for Programs/Policies and Interpretation of Research Evidence (INSPIRE). J. Nutr. 2015, 145, 1039S–1108S. [Google Scholar] [CrossRef]
- Rosales, F.J.; Ritter, S.J.; Zolfaghari, R.; Smith, J.E.; Ross, A.C. Effects of acute inflammation on plasma retinol, retinol-binding protein, and its mRNA in the liver and kidneys of vitamin A-sufficient rats. J. Lipid Res. 1996, 37, 962–971. [Google Scholar] [CrossRef]
- Gieng, J.; Green, M.H.; Green, J.B.; Rosales, F.J. Model-based compartmental analysis indicates a reduced mobilization of hepatic vitamin A during inflammation in rats. J. Lipid Res. 2007, 48, 904–913. [Google Scholar] [CrossRef]
- Rosales, F.J.; Ross, A.C. A Low Molar Ratio of Retinol Binding Protein to Transthyretin Indicates Vitamin A Deficiency during Inflammation: Studies in Rats and A Posteriori Analysis of Vitamin A-Supplemented Children with Measles. J. Nutr. 1998, 128, 1681–1687. [Google Scholar] [CrossRef]
- Rosales, F.J.; Ross, A.C. Inflammation in Human Immunodeficiency Virus Type 1 Infection as a Cause of Decreased Plasma Retinol. J. Infect. Dis. 1996, 173, 507–508. [Google Scholar] [CrossRef]
- Rosales, F.J.; Topping, J.D.; Smith, J.E.; Shankar, A.H.; Ross, A.C. Relation of serum retinol to acute phase proteins and malarial morbidity in Papua New Guinea children. Am. J. Clin. Nutr. 2000, 71, 1582–1588. [Google Scholar] [CrossRef]
- Ataga, K.I.; Saraf, S.L.; Derebail, V.K. The nephropathy of sickle cell trait and sickle cell disease. Nat. Rev. Nephrol. 2022, 18, 361–377. [Google Scholar] [CrossRef] [PubMed]
- Ataga, K.I.; Derebail, V.K.; Archer, D.R. The glomerulopathy of sickle cell disease. Am. J. Hematol. 2014, 89, 907–914. [Google Scholar] [CrossRef] [PubMed]
- Gliozzi, M.L.; Rbaibi, Y.; Long, K.R.; Vitturi, D.A.; Weisz, O.A. Hemoglobin alters vitamin carrier uptake and vitamin D metabolism in proximal tubule cells: Implications for sickle cell disease. Am. J. Physiol. Cell Physiol. 2019, 317, C993–C1000. [Google Scholar] [CrossRef] [PubMed]
- Raila, J.; Willnow, T.; Schweigert, F. Megalin-Mediated Reuptake of Retinol in the Kidneys of Mice Is Essential for Vitamin A Homeostasis. J. Nutr. 2005, 135, 2512–2516. [Google Scholar] [CrossRef] [PubMed]
- Christensen, E.I.; Moskaug, J.; Vorum, H.; Jacobsen, C.; Gundersen, T.E.; Nykjaer, A.; Blomhoff, R.; Willnow, T.; Moestrup, S.K. Evidence for an Essential Role of Megalin in Transepithelial Transport of Retinol. J. Am. Soc. Nephrol. 1999, 10, 685–695. [Google Scholar] [CrossRef]
- Sousa, M.; Norden, A.G.W.; Jacobsen, C.; Willnow, T.; Christensen, E.I.; Thakker, R.; Verroust, P.J.; Moestrup, S.K.; Saraiva, M.J. Evidence for the Role of Megalin in Renal Uptake of Transthyretin. J. Biol. Chem. 2000, 275, 38176–38181. [Google Scholar] [CrossRef]
- Negri, A.L. Proximal tubule endocytic apparatus as the specific renal uptake mechanism for vitamin D-binding protein/25-(OH)D3 complex. Nephrology 2006, 11, 510–515. [Google Scholar] [CrossRef]
- Gembillo, G.; Cernaro, V.; Siligato, R.; Curreri, F.; Catalano, A.; Santoro, D. Protective Role of Vitamin D in Renal Tubulopathies. Metabolites 2020, 10, 115. [Google Scholar] [CrossRef]
- Samarasinghe, A.E.; Penkert, R.R.; Hurwitz, J.L.; Sealy, R.E.; LeMessurier, K.S.; Hammond, C.; Dubin, P.J.; Lew, D.B. Questioning Cause and Effect: Children with Severe Asthma Exhibit High Levels of Inflammatory Biomarkers Including Beta-Hexosaminidase, but Low Levels of Vitamin A and Immunoglobulins. Biomedicines 2020, 8, 393. [Google Scholar] [CrossRef]
- Jones, B.G.; Oshansky, C.M.; Bajracharya, R.; Tang, L.; Sun, Y.; Wong, S.S.; Webby, R.; Thomas, P.G.; Hurwitz, J.L. Retinol binding protein and vitamin D associations with serum antibody isotypes, serum influenza virus-specific neutralizing activities and airway cytokine profiles. Clin. Exp. Immunol. 2016, 183, 239–247. [Google Scholar] [CrossRef]
- Reid, M. Nutrition and sickle cell disease. C. R. Biol. 2013, 336, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Ghafuri, D.L.; Rodeghier, M.; DeBaun, M.R. Rate of Food Insecurity Among Households with Children with Sickle Cell Disease is Above the National Average. South. Med. J. 2020, 113, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.; George, E.O.; Wang, W. Nutrition assessment in children with sickle cell disease. J. Assoc. Acad. Minor. Physicians 1997, 8, 44–48. [Google Scholar]
- Badaloo, A.; Jackson, A.A.; Jahoor, F. Whole Body Protein Turnover and Resting Metabolic Rate in Homozygous Sickle Cell Disease. Clin. Sci. 1989, 77, 93–97. [Google Scholar] [CrossRef]
- Barden, E.M.; Zemel, B.S.; Kawchak, D.A.; Goran, M.I.; Ohene-Frempong, K.; Stallings, V.A. Total and resting energy expenditure in children with sickle cell disease. J. Pediatr. 2000, 136, 73–79. [Google Scholar] [CrossRef]
- Singhal, A.; Parker, S.; Linsell, L.; Serjeant, G. Energy intake and resting metabolic rate in preschool Jamaican children with homozygous sickle cell disease. Am. J. Clin. Nutr. 2002, 75, 1093–1097. [Google Scholar] [CrossRef]
- Hibbert, J.M.; Creary, M.S.; Gee, B.; Buchanan, I.D.; Quarshie, A.; Hsu, L. Erythropoiesis and Myocardial Energy Requirements Contribute to the Hypermetabolism of Childhood Sickle Cell Anemia. J. Pediatr. Gastroenterol. Nutr. 2006, 43, 680–687. [Google Scholar] [CrossRef]
- Singhal, A.; Davies, P.; Sahota, A.; Thomas, P.W.; Serjeant, G.R. Resting metabolic rate in homozygous sickle cell disease. Am. J. Clin. Nutr. 1993, 57, 32–34. [Google Scholar] [CrossRef]
- Gray, N.T.; Bartlett, J.M.; Kolasa, K.M.; Marcuard, S.P.; Holbrook, C.T.; Horner, R.D. Nutritional Status and Dietary Intake of Children with Sickle Cell Anemia. J. Pediatr. Hematol. 1992, 14, 57–61. [Google Scholar] [CrossRef]
- Okany, C.C.; Akinyanju, O.O. The influence of socio-economic status on the severity of sickle cell disease. Afr. J. Med. Med. Sci. 1993, 22, 57–60. [Google Scholar]
- Jesus, A.C.D.S.D.; Konstantyner, T.; Lôbo, I.K.V.; Braga, J.A.P. Socioeconomic and Nutritional Characteristics of Children and Adolescents with Sickle Cell Anemia: A Systematic Review. Rev. Paul. Pediatr. 2018, 36, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Newton, D.A.; Baatz, J.E.; Kindy, M.S.; Gattoni-Celli, S.; Shary, J.R.; Hollis, B.W.; Wagner, C.L. Vitamin D binding protein polymorphisms significantly impact vitamin D status in children. Pediatr. Res. 2019, 86, 662–669. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.T.; Kattan, M.; Fennoy, I.; Arpadi, S.M.; Miller, R.L.; Cremers, S.; McMahon, N.J.; Nieves, J.; Brittenham, G.M. Randomized phase 2 trial of monthly vitamin D to prevent respiratory complications in children with sickle cell disease. Blood Adv. 2018, 2, 969–978. [Google Scholar] [CrossRef] [PubMed]
- Hood, A.M.; Quinn, C.T.; King, C.D.; Shook, L.M.; Peugh, J.L.; Crosby, L.E. Vitamin D supplementation and pain-related emergency department visits in children with sickle cell disease. Complement. Ther. Med. 2020, 49, 102342. [Google Scholar] [CrossRef]
- Imdad, A.; Mayo-Wilson, E.; Haykal, M.R.; Regan, A.; Sidhu, J.; Smith, A.; Bhutta, Z.A. Vitamin A supplementation for preventing morbidity and mortality in children from six months to five years of age. Cochrane Database Syst. Rev. 2022, 3, CD008524. [Google Scholar] [CrossRef]
- Dougherty, K.A.; Schall, J.I.; Bertolaso, C.; Smith-Whitley, K.; Stallings, V.A. Vitamin D Supplementation Improves Health-Related Quality of Life and Physical Performance in Children with Sickle Cell Disease and in Healthy Children. J. Pediatr. Health Care 2020, 34, 424–434. [Google Scholar] [CrossRef]
- Brownell, J.N.; Schall, J.I.; Mcanlis, C.R.; Smith-Whitley, K.; Norris, C.F.; Stallings, V.A. Effect of High-dose Vitamin A Supplementation in Children With Sickle Cell Disease: A Randomized, Double-blind, Dose-finding Pilot Study. J. Pediatr. Hematol. 2020, 42, 83–91. [Google Scholar] [CrossRef]
- Grégoire-Pelchat, P.; Pastore, Y.; Robitaille, N.; Lemay, S.; Khamessan, A.; Kleiber, N.; Nyalendo, C.; Gagné, N.; Alos, N.; Mailhot, G. Comparison of two vitamin D supplementation strategies in children with sickle cell disease: A randomized controlled trial. Br. J. Haematol. 2021, 192, 385–394. [Google Scholar] [CrossRef]
- Adekunle, M.O.; Dada, A.O.; Njokanma, F.O.; Solarin, A.U.; Animasahun, B.A.; Lamina, M.O. Comparative Effectiveness of a Six-Week Treatment Course of Vitamin D2 and D3 in Children With Sickle Cell Anemia in Steady State With Hypovitaminosis D: A Randomized Clinical Trial. J. Hematol. 2021, 10, 114–122. [Google Scholar] [CrossRef]
- Soe, H.H.K.; Abas, A.B.; Than, N.N.; Ni, H.; Singh, J.; Said, A.R.B.M.; Osunkwo, I. Vitamin D supplementation for sickle cell disease. Cochrane Database Syst. Rev. 2017, 1, CD010858. [Google Scholar] [CrossRef]
- Urashima, M.; Segawa, T.; Okazaki, M.; Kurihara, M.; Wada, Y.; Ida, H. Randomized trial of vitamin D supplementation to prevent seasonal influenza A in schoolchildren. Am. J. Clin. Nutr. 2010, 91, 1255–1260. [Google Scholar] [CrossRef] [PubMed]
- Bresee, J.S.; Fischer, M.; Dowell, S.F.; Johnston, B.D.; Biggs, V.M.; Levine, R.S.; Lingappa, J.R.; Keyserling, H.L.; Petersen, K.M.; Bak, J.R.; et al. Vitamin A therapy for children with respiratory syncytial virus infection: A multicenter trial in the United States. Pediatr. Infect. Dis. J. 1996, 15, 777–782. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Cameron, N.A.; Linder, J.A. Multivitamins and Supplements—Benign Prevention or Potentially Harmful Distraction? JAMA 2022, 327, 2294–2295. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, E.A.; Evans, C.V.; Ivlev, I.; Rushkin, M.C.; Thomas, R.G.; Martin, A.; Lin, J.S. Vitamin and Mineral Supplements for the Primary Prevention of Cardiovascular Disease and Cancer: Updated Evidence Report and Systematic Review for the US Preventive Services Task Force. JAMA 2022, 327, 2334–2347. [Google Scholar] [CrossRef] [PubMed]
- Force, U.P.S.T.; Mangione, C.M.; Barry, M.J.; Nicholson, W.K.; Cabana, M.; Chelmow, D.; Coker, T.R.; Davis, E.M.; Donahue, K.E.; Doubeni, C.A.; et al. Vitamin, Mineral, and Multivitamin Supplementation to Prevent Cardiovascular Disease and Cancer: US Preventive Services Task Force Recommendation Statement. JAMA 2022, 327, 2326–2333. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Sex | Age of Patient upon Enrollment (Years) | Sickle Genotype | Retinol (μg/dL) | Vitamin D (ng/mL) |
|---|---|---|---|---|---|
| SC001 | Male | 6.8 | SS | 7.9 | 22 |
| SC002 | Male | 5.5 | SS | 8.9 | 12 |
| SC004 | Female | 7.2 | SS | 7.7 | 18 |
| SC005 | Male | 5.8 | SS | 12.7 | 15 |
| SC006 | Female | 8.5 | SC | 9.3 | 20 |
| SC007 | Female | 6.7 | SS | 11.7 | 23 |
| SC008 | Male | 6.1 | SS | 11.8 | 25 |
| SC011 | Male | 14.3 | SS | 11.1 | 6 |
| SC012 | Female | 9.8 | SS | 12.0 | 26 |
| SC013 | Male | 4.5 | SS | 14.1 | 37 |
| SC014 | Female | 7.2 | SS | 12.1 | 34 |
| SC016 | Male | 10.6 | SS | ND | 24 |
| SC017 | Female | 6.2 | SS | 15.4 | 44 |
| SC018 | Female | 11.4 | S Beta + Thal | 22.8 | 26 |
| SC019 | Male | 6.0 | SD | 11.2 | 63 |
| SC020 | Female | 7.0 | SS | 11.4 | 8 |
| SC021 | Male | 11.8 | SS | 12.1 | 27 |
| SC022 | Female | 8.8 | SC | 10.2 | 10 |
| SC023 | Female | 5.3 | SS | 18.5 | 27 |
| SC024 | Male | 12.8 | SC | 24.1 | 12 |
| Factor | Day 0 Retinol | Day 0 Vit D | Day 0 Virus | Day 0 ARC | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Virus-specific Antibodies | Corr | p | r | Corr | p | R | Corr | p | R | Corr | p | r |
| Serum Day 0 IgM Diagnostic assay | ----- | 0.23 | 0.29 | ----- | 0.58 | −0.13 | ----- | >0.99 | −0.14 | ----- | 0.48 | 0.17 |
| Serum Day 0 IgG Diagnostic assay | ----- | 0.92 | 0.03 | ----- | 0.45 | 0.18 | Negative | 0.049 | −0.49 | Positive | 0.002 | 0.65 |
| Serum Peak IgG VLP ELISA | ----- | 0.17 | 0.33 | ----- | 0.25 | 0.27 | ----- | 0.96 | −0.01 | ----- | 0.31 | 0.24 |
| NW Day 0 IgG VLP ELISA | ----- | 0.38 | 0.24 | ----- | 0.72 | 0.10 | ----- | 0.93 | 0.06 | Positive | 0.032 | 0.51 |
| NW Peak Ig GVLP ELISA | Positive | 0.020 | 0.53 | Positive | 0.047 | 0.46 | ----- | 0.94 | 0.02 | ----- | 0.55 | 0.15 |
| Vitamin | Retinol | Vitamin D | ||||
|---|---|---|---|---|---|---|
| Factor | Correlation | p | r | Correlation | P | r |
| FKN | Negative | 0.008 | −0.59 | ----- | 0.089 | −0.39 |
| Flt-3L | Negative | 0.044 | −0.47 | ----- | 0.34 | −0.22 |
| G-CSF | Negative | 0.011 | −0.57 | ----- | 0.22 | −0.29 |
| GM-CSF | Negative | 0.023 | −0.52 | ----- | 0.40 | −0.20 |
| IFNα2 | Negative | 0.010 | −0.58 | ----- | 0.071 | −0.41 |
| IL−1RA | Negative | 0.016 | −0.55 | ----- | 0.19 | −0.31 |
| IL-3 | Negative | 0.049 | −0.46 | ----- | 0.12 | −0.36 |
| IL-4 | Negative | 0.008 | −0.59 | ----- | 0.053 | −0.44 |
| IL-5 | Negative | 0.013 | −0.56 | ----- | 0.14 | −0.34 |
| IL-6 | Negative | 0.009 | −0.58 | ----- | 0.086 | −0.39 |
| IL-7 | Negative | 0.021 | −0.53 | ----- | 0.10 | −0.38 |
| IL-9 | Negative | 0.019 | −0.53 | ----- | 0.10 | −0.38 |
| IL-12p40 | Negative | 0.003 | −0.65 | Negative | 0.044 | −0.45 |
| IL-12p70 | Negative | 0.006 | −0.61 | ----- | 0.071 | −0.41 |
| IL-13 | Negative | 0.025 | −0.51 | ----- | 0.22 | −0.29 |
| IL-15 | Negative | 0.003 | −0.65 | Negative | 0.045 | −0.45 |
| IL-17A | Negative | 0.002 | −0.66 | Negative | 0.005 | −0.60 |
| MIP-1b | Negative | 0.013 | −0.56 | ----- | 0.43 | −0.19 |
| TNFα | Negative | 0.049 | −0.46 | ----- | 0.39 | −0.21 |
| TNFβ | Negative | 0.015 | −0.55 | ----- | 0.12 | −0.36 |
| Virus | ARC | Serum Virus-Specific IgG (Day 0, Diagnostic Assay) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Factor | Corr | p value | R value | Corr | p value | R value | Corr | p value | R value |
| G-CSF | ----- | 0.41 | −0.21 | Positive | 0.041 | 0.46 | ----- | 0.74 | 0.08 |
| GRO | Negative | 0.025 | −0.56 | ----- | 0.092 | 0.41 | ----- | 0.12 | 0.38 |
| IFNγ | ----- | 0.88 | 0.04 | ----- | 0.14 | −0.34 | Negative | 0.006 | −0.59 |
| IL-10 | Positive | 0.006 | 0.64 | Negative | 0.011 | −0.55 | Negative | 0.003 | −0.62 |
| IP-10 | ----- | 0.099 | 0.41 | ----- | 0.27 | −0.26 | Negative | 0.001 | −0.69 |
| TGFα | ----- | 0.42 | −0.22 | Positive | 0.008 | 0.59 | 0.77 | 0.07 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Penkert, R.R.; Azul, M.; Sealy, R.E.; Jones, B.G.; Dowdy, J.; Hayden, R.T.; Tang, L.; Ross, A.C.; Hankins, J.S.; Hurwitz, J.L. Hypothesis: Low Vitamin A and D Levels Worsen Clinical Outcomes When Children with Sickle Cell Disease Encounter Parvovirus B19. Nutrients 2022, 14, 3415. https://doi.org/10.3390/nu14163415
Penkert RR, Azul M, Sealy RE, Jones BG, Dowdy J, Hayden RT, Tang L, Ross AC, Hankins JS, Hurwitz JL. Hypothesis: Low Vitamin A and D Levels Worsen Clinical Outcomes When Children with Sickle Cell Disease Encounter Parvovirus B19. Nutrients. 2022; 14(16):3415. https://doi.org/10.3390/nu14163415
Chicago/Turabian StylePenkert, Rhiannon R., Melissa Azul, Robert E. Sealy, Bart G. Jones, Jola Dowdy, Randall T. Hayden, Li Tang, A. Catharine Ross, Jane S. Hankins, and Julia L. Hurwitz. 2022. "Hypothesis: Low Vitamin A and D Levels Worsen Clinical Outcomes When Children with Sickle Cell Disease Encounter Parvovirus B19" Nutrients 14, no. 16: 3415. https://doi.org/10.3390/nu14163415
APA StylePenkert, R. R., Azul, M., Sealy, R. E., Jones, B. G., Dowdy, J., Hayden, R. T., Tang, L., Ross, A. C., Hankins, J. S., & Hurwitz, J. L. (2022). Hypothesis: Low Vitamin A and D Levels Worsen Clinical Outcomes When Children with Sickle Cell Disease Encounter Parvovirus B19. Nutrients, 14(16), 3415. https://doi.org/10.3390/nu14163415