
Mechanisms of Sodium/Iodide Symporter-Mediated Mammary Gland Iodine Compensation during Lactation


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Animals and Treatments
2.2. Urine
2.3. Rats’ Milk
2.4. Measurement of Iodine in the Mammary Gland, Milk, Urine, and Serum
2.5. Thyroid Function Tests
2.6. HE Staining
2.7. Western Blot Analysis
2.8. Immunohistochemistry
2.9. Statistical Analysis
3. Results
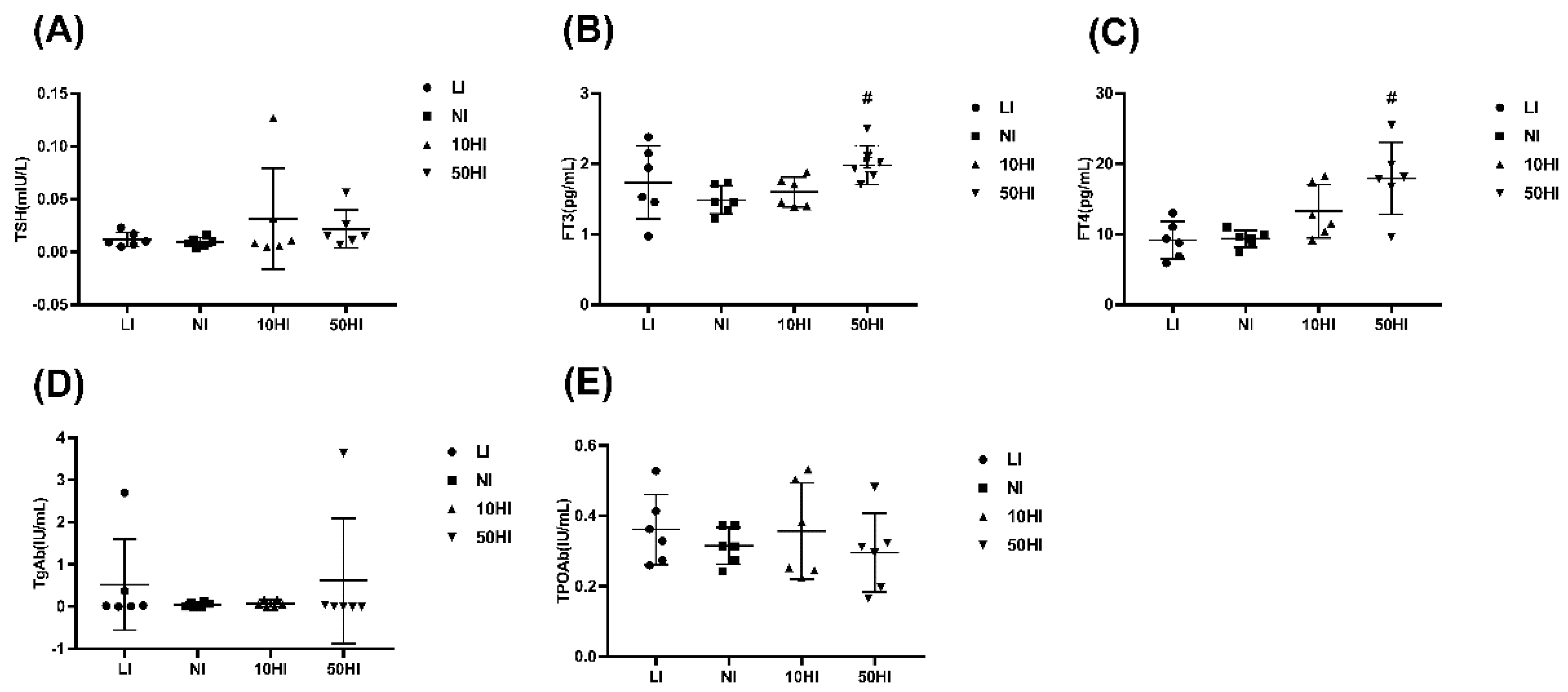
3.1. Iodine Nutrition and Thyroid Hormone Levels in Lactating Rats
3.2. Effects of Different Iodine Intakes on Breast Milk Iodine Concentration and Mammary Gland Iodine Content in Lactating Rats
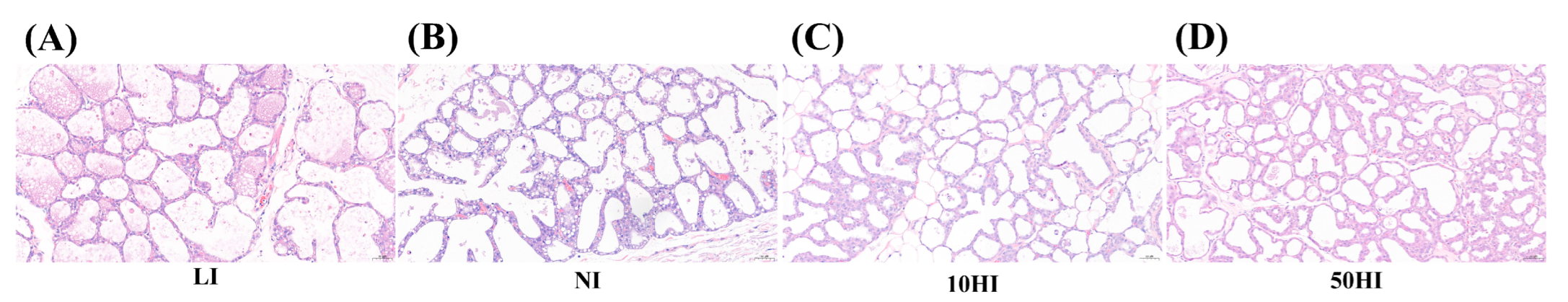
3.3. Effects of Iodine on Mammary Gland Cell Morphology in Lactating Rats
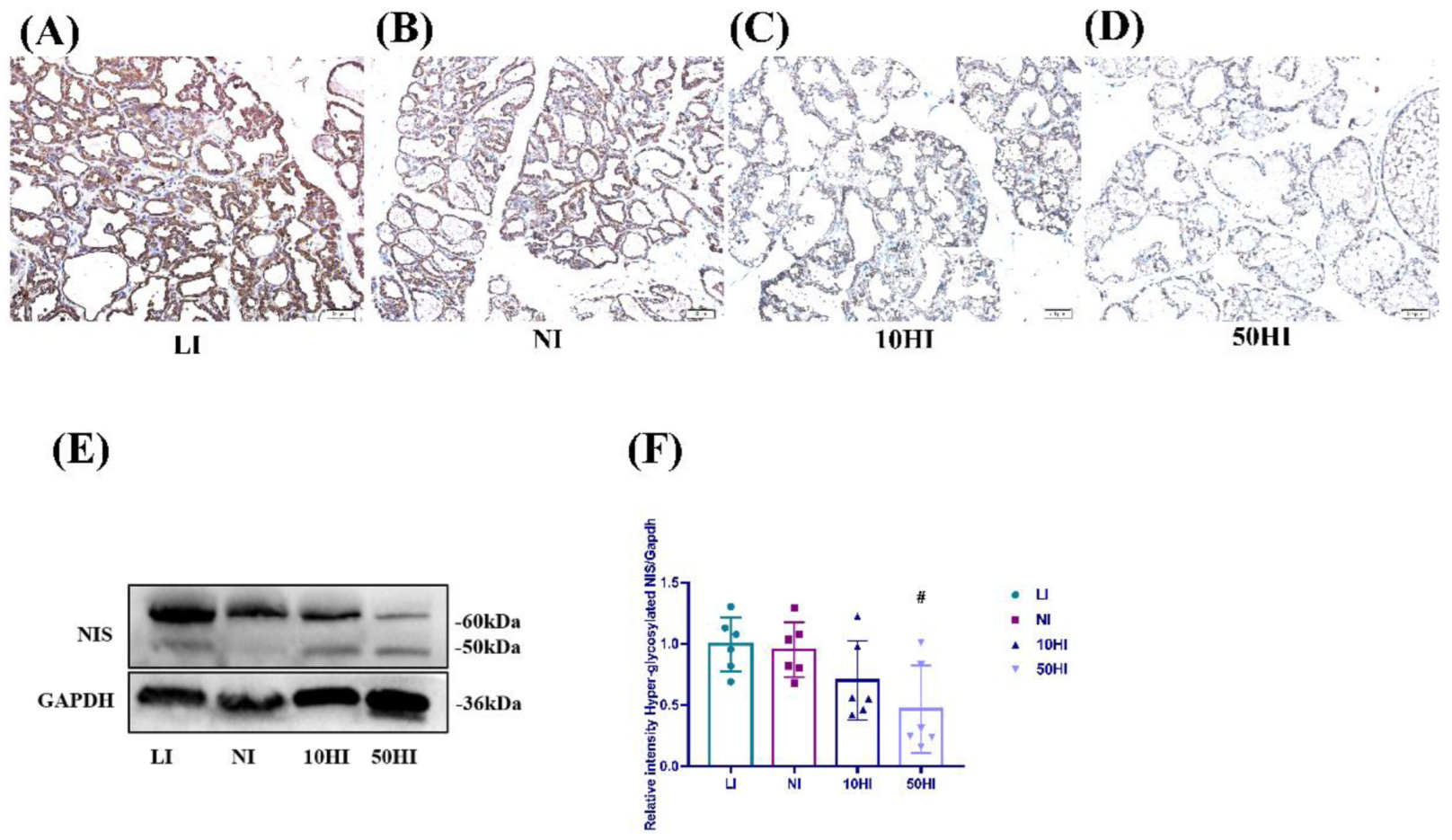
3.4. Effect of Iodine on NIS Expression in the Mammary Gland in Lactating Rats
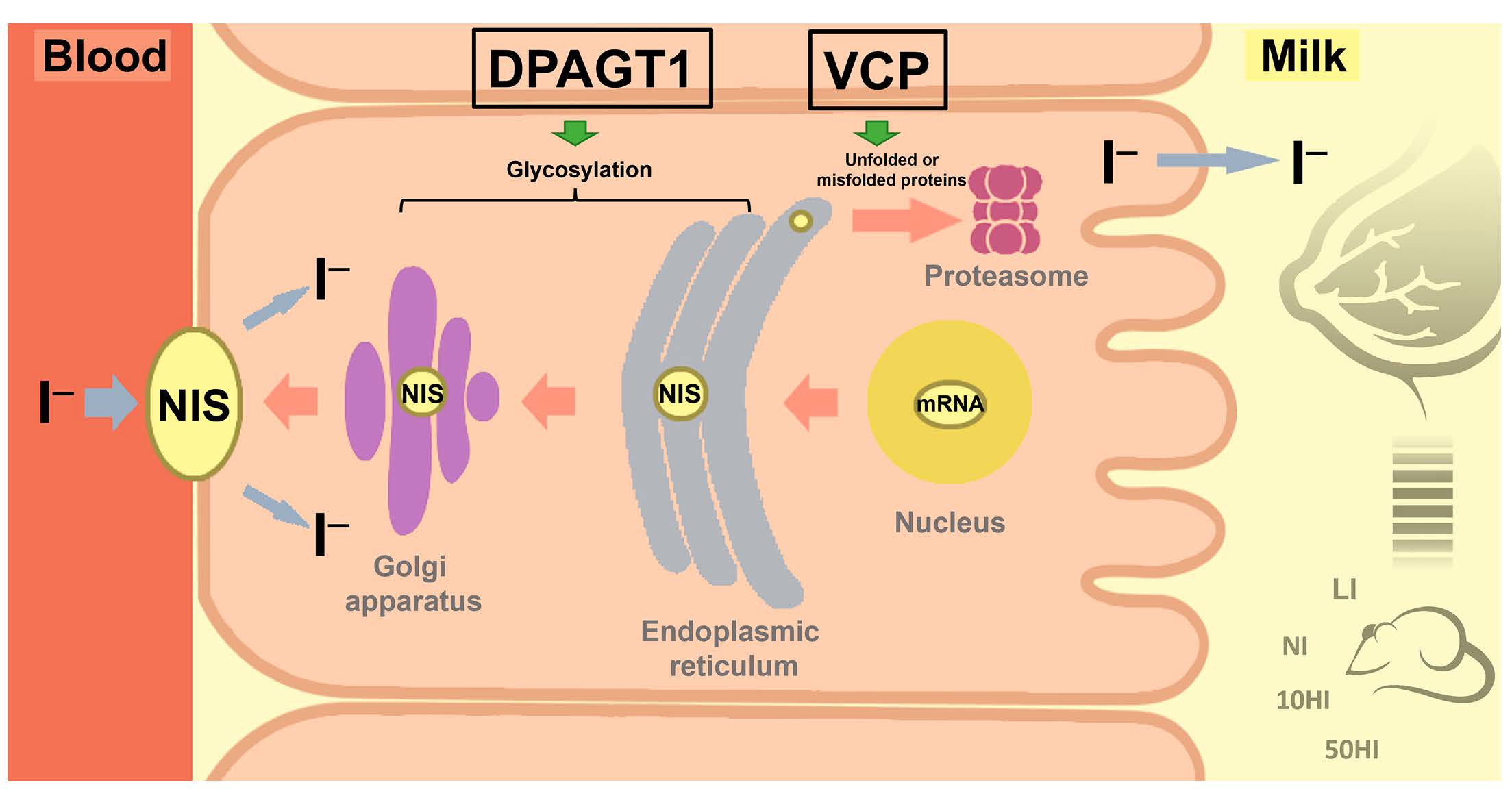
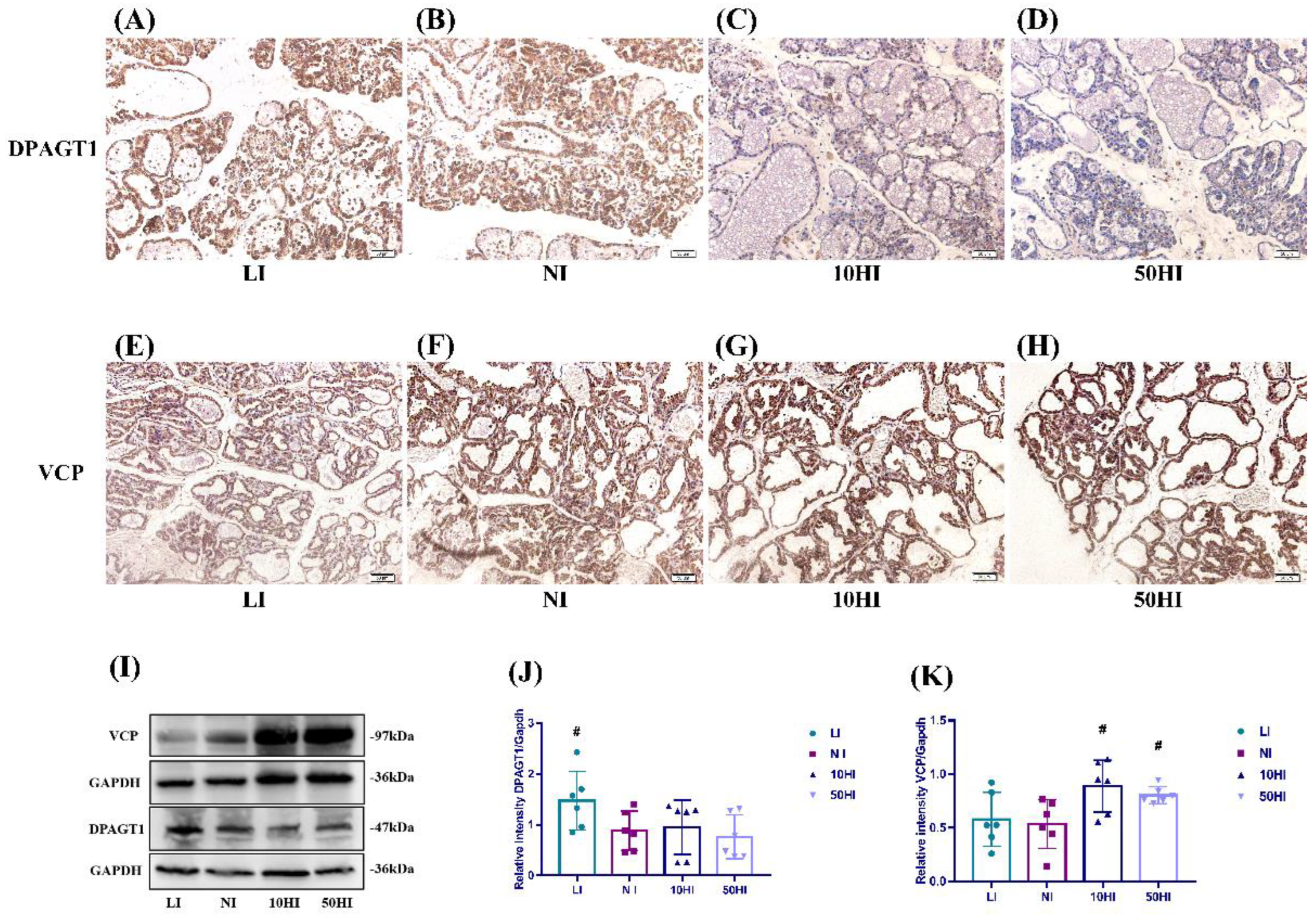
3.5. Effect of Iodine on NIS Glycosylation and Degradation in the Mammary Gland during Lactation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Velasco, I.; Bath, S.C.; Rayman, M.P. Iodine as Essential Nutrient during the First 1000 Days of Life. Nutrients 2018, 10, 290. [Google Scholar] [CrossRef]
- Schwarzenberg, S.J.; Georgieff, M.K. Advocacy for Improving Nutrition in the First 1000 Days to Support Childhood Development and Adult Health. Pediatrics 2018, 141, e20173716. [Google Scholar] [CrossRef]
- Mulrine, H.M.; Skeaff, S.A.; Ferguson, E.L.; Gray, A.R.; Valeix, P. Breast-milk iodine concentration declines over the first 6 mo postpartum in iodine-deficient women. Am. J. Clin. Nutr. 2010, 92, 849–856. [Google Scholar] [CrossRef]
- Chen, Z.-P.; Hetzel, B.S. Cretinism revisited. Best Pract. Res. Clin. Endocrinol. Metab. 2010, 24, 39–50. [Google Scholar] [CrossRef]
- Andersson, M.; Braegger, C.P. The Role of Iodine for Thyroid Function in Lactating Women and Infants. Endocr. Rev. 2021, 43, 469–506. [Google Scholar] [CrossRef]
- Swanson, C.A.; Pearce, E.N. Iodine Insufficiency: A Global Health Problem? Adv. Nutr. Int. Rev. J. 2013, 4, 533–535. [Google Scholar] [CrossRef]
- Liu, L.; Wang, D.; Liu, P.; Meng, F.; Wen, D.; Jia, Q.; Liu, J.; Zhang, X.; Jiang, P.; Shen, H. The relationship between iodine nutrition and thyroid disease in lactating women with different iodine intakes. Br. J. Nutr. 2015, 114, 1487–1495. [Google Scholar] [CrossRef]
- Abel, M.H.; Caspersen, I.H.; Meltzer, H.M.; Haugen, M.; Brandlistuen, R.E.; Aase, H.; Alexander, J.; Torheim, L.E.; Brantsæter, A.-L. Suboptimal Maternal Iodine Intake Is Associated with Impaired Child Neurodevelopment at 3 Years of Age in the Norwegian Mother and Child Cohort Study. J. Nutr. 2017, 147, 1314–1324. [Google Scholar] [CrossRef]
- Hannan, M.A.; Faraji, B.; Tanguma, J.; Longoria, N.; Rodriguez, R.C. Maternal Milk Concentration of Zinc, Iron, Selenium, and Iodine and Its Relationship to Dietary Intakes. Biol. Trace Element Res. 2008, 127, 6–15. [Google Scholar] [CrossRef]
- Dold, S.; Zimmermann, M.B.; Aboussad, A.; Cherkaoui, M.; Jia, Q.; Jukic, T.; Kusic, Z.; Quirino, A.; Sang, Z.; Luis, T.O.S.; et al. Breast Milk Iodine Concentration Is a More Accurate Biomarker of Iodine Status Than Urinary Iodine Concentration in Exclusively Breastfeeding Women. J. Nutr. 2017, 147, 528–537. [Google Scholar] [CrossRef] [Green Version]
- Brown-Grant, K. The iodide concentrating mechanism of the mammary gland. J. Physiol. 1957, 135, 644–654. [Google Scholar] [CrossRef]
- Rillema, J.A.; Hill, M.A. Prolactin regulation of the pendrin-iodide transporter in the mammary gland. Am. J. Physiol. Metab. 2003, 284, E25–E28. [Google Scholar] [CrossRef]
- Poole, V.L.; McCabe, C.J. Iodide transport and breast cancer. J. Endocrinol. 2015, 227, R1–R12. [Google Scholar] [CrossRef]
- Huet, G.; Gouyer, V.; Delacour, D.; Richet, C.; Zanetta, J.; Delannoy, P.; Degand, P. Involvement of glycosylation in the intracellular trafficking of glycoproteins in polarized epithelial cells. Biochimie 2003, 85, 323–330. [Google Scholar] [CrossRef]
- Beyer, S.; Lakshmanan, A.; Liu, Y.-Y.; Zhang, X.; Wapnir, I.; Smolenski, A.; Jhiang, S. KT5823 Differentially Modulates Sodium Iodide Symporter Expression, Activity, and Glycosylation between Thyroid and Breast Cancer Cells. Endocrinology 2011, 152, 782–792. [Google Scholar] [CrossRef]
- Chung, T.; Youn, H.; Yeom, C.J.; Kang, K.W.; Chung, J.-K. Glycosylation of Sodium/Iodide Symporter (NIS) Regulates Its Membrane Translocation and Radioiodine Uptake. PLoS ONE 2015, 10, e0142984. [Google Scholar] [CrossRef]
- Feng, F.; Yehia, L.; Ni, Y.; Chang, Y.S.; Jhiang, S.M.; Eng, C. A Nonpump Function of Sodium Iodide Symporter in Thyroid Cancer via Cross-talk with PTEN Signaling. Cancer Res. 2018, 78, 6121–6133. [Google Scholar] [CrossRef]
- Fletcher, A.; Read, M.L.; Thornton, C.E.M.; Larner, D.P.; Poole, V.L.; Brookes, K.; Nieto, H.R.; Alshahrani, M.; Thompson, R.J.; Lavery, G.G.; et al. Targeting Novel Sodium Iodide Symporter Interactors ADP-Ribosylation Factor 4 and Valosin-Containing Protein Enhances Radioiodine Uptake. Cancer Res. 2020, 80, 102–115. [Google Scholar] [CrossRef]
- Bastola, P.; Neums, L.; Schoenen, F.J.; Chien, J. VCP inhibitors induce endoplasmic reticulum stress, cause cell cycle arrest, trigger caspase-mediated cell death and synergistically kill ovarian cancer cells in combination with Salubrinal. Mol. Oncol. 2016, 10, 1559–1574. [Google Scholar] [CrossRef]
- Song, C.; Wang, Q.; Rogers, T.J. Valosin-containing protein (VCP/p97) is capable of unfolding polyubiquitinated proteins through its ATPase domains. Biochem. Biophys. Res. Commun. 2015, 463, 453–457. [Google Scholar] [CrossRef] [Green Version]
- WHO/UNICEF/ICCIDD. Assessment of Iodine Deficiency Disorders and Monitoring their Elimination: A Guide for Programs Managers, 3rd ed.; World Health Organization: Geneva, Switzerland, 2007.
- Semba, R.D.; Delange, F. Iodine in Human Milk: Perspectives for Infant Health. Nutr. Rev. 2001, 59, 269–278. [Google Scholar] [CrossRef]
- Nazeri, P.; Mirmiran, P.; Shiva, N.; Mehrabi, Y.; Mojarrad, M.; Azizi, F. Iodine Nutrition Status in Lactating Mothers Residing in Countries with Mandatory and Voluntary Iodine Fortification Programs: An Updated Systematic Review. Thyroid 2015, 25, 611–620. [Google Scholar] [CrossRef]
- Azizi, F.; Smyth, P. Breastfeeding and maternal and infant iodine nutrition. Clin. Endocrinol. 2009, 70, 803–809. [Google Scholar] [CrossRef]
- Micali, S.; Bulotta, S.; Puppin, C.; Territo, A.; Navarra, M.; Bianchi, G.; Damante, G.; Filetti, S.; Russo, D. Sodium iodide symporter (NIS) in extrathyroidal malignancies: Focus on breast and urological cancer. BMC Cancer 2014, 14, 303. [Google Scholar] [CrossRef]
- Tazebay, U.H.; Wapnir, I.L.; Levy, O.; Dohan, O.; Zuckier, L.; Zhao, Q.H.; Deng, H.F.; Amenta, P.S.; Fineberg, S.; Pestell, R.G.; et al. The mammary gland iodide transporter is expressed during lactation and in breast cancer. Nat. Med. 2000, 6, 871–878. [Google Scholar] [CrossRef]
- Rillema, J.A.; Yu, T.X.; Jhiang, S.M. Effect of prolactin on sodium iodide symporter expression in mouse mammary gland explants. Am. J. Physiol. Metab. 2000, 279, E769–E772. [Google Scholar] [CrossRef]
- Yu, X.; Shen, H.; Liu, L.; Lin, L.; Gao, M.; Wang, S. Changes of Sodium Iodide Symporter Regulated by IGF-I and TGF-β1 in Mammary Gland Cells from Lactating Mice at Different Iodine Levels. Biol. Trace Element Res. 2011, 146, 73–78. [Google Scholar] [CrossRef]
- Serrano-Nascimento, C.; Salgueiro, R.B.; Vitzel, K.F.; Pantaleão, T.; Corrêa da Costa, V.M.; Nunes, M.T. Iodine excess exposure during pregnancy and lactation impairs maternal thyroid function in rats. Endocr. Connect. 2017, 6, 510–521. [Google Scholar] [CrossRef]
- Sun, Y.; Han, Y.; Qian, M.; Li, Y.; Ye, Y.; Lin, L.; Liu, Y. Defending Effects of Iodide Transfer in Placental Barrier Against Maternal Iodine Deficiency. Thyroid 2021, 31, 509–518. [Google Scholar] [CrossRef]
- Belaya, K.; Finlayson, S.; Cossins, J.; Liu, W.W.; Maxwell, S.; Palace, J.; Beeson, D. Identification of DPAGT1 as a new gene in which mutations cause a congenital myasthenic syndrome. Ann. N. Y. Acad. Sci. 2012, 1275, 29–35. [Google Scholar] [CrossRef]
- Jiang, Z.; Kuo, Y.-H.; Zhong, M.; Zhang, J.; Zhou, X.X.; Xing, L.; Wells, J.A.; Wang, Y.; Arkin, M.R. Adaptor-Specific Antibody Fragment Inhibitors for the Intracellular Modulation of p97 (VCP) Protein–Protein Interactions. J. Am. Chem. Soc. 2022, 144, 13218–13225. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | N | Serum | Urine | |
|---|---|---|---|---|
| STIC (μg/L) | SNBIC (μg/L) | UIC (μg/L) a | ||
| LI | 6 | 29.12 (25.08, 33.27) | 19.79 (18.61, 21.66) | 87.45 (78.71, 95.64) |
| NI | 6 | 55.77 (42.10, 71.76) | 54.33 (41.95, 63.48) | 369.96 (312.10, 433.28) |
| 10HI | 6 | 338.18 (218.43, 368.25) | 257.19 (220.31, 293.38) | 3569.18 (3287.96, 3793.56) |
| 50HI | 6 | 1090.08 (981.04, 1199.83) # | 841.83 (758.49, 903.45) # | 12,286.04 (11,470.80,13,362.59) # |
| Group | N | BMIC (μg/L) a | MIC (μg) |
|---|---|---|---|
| LI | 6 | 26.46 (22.91, 44.91) | 0.38 (0.29, 0.50) |
| NI | 6 | 220.46 (160.61, 224.36) | 0.81 (0.74, 1.37) |
| 10HI | 6 | 1188.75 (936.76, 1473.97) | 6.30 (2.37, 8.10) |
| 50HI | 6 | 7413.31 (4226.06, 9175.44) # | 72.19 (56.71, 99.63) # |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, M.; Gao, Y.; Guo, W.; Meng, Q.; Jin, Q.; Yang, R.; Yang, Y.; Zhang, Y.; Zhang, W. Mechanisms of Sodium/Iodide Symporter-Mediated Mammary Gland Iodine Compensation during Lactation. Nutrients 2022, 14, 3592. https://doi.org/10.3390/nu14173592
Fu M, Gao Y, Guo W, Meng Q, Jin Q, Yang R, Yang Y, Zhang Y, Zhang W. Mechanisms of Sodium/Iodide Symporter-Mediated Mammary Gland Iodine Compensation during Lactation. Nutrients. 2022; 14(17):3592. https://doi.org/10.3390/nu14173592
Chicago/Turabian StyleFu, Min, Yuanpeng Gao, Wenxing Guo, Qi Meng, Qi Jin, Rui Yang, Ying Yang, Yaqi Zhang, and Wanqi Zhang. 2022. "Mechanisms of Sodium/Iodide Symporter-Mediated Mammary Gland Iodine Compensation during Lactation" Nutrients 14, no. 17: 3592. https://doi.org/10.3390/nu14173592
APA StyleFu, M., Gao, Y., Guo, W., Meng, Q., Jin, Q., Yang, R., Yang, Y., Zhang, Y., & Zhang, W. (2022). Mechanisms of Sodium/Iodide Symporter-Mediated Mammary Gland Iodine Compensation during Lactation. Nutrients, 14(17), 3592. https://doi.org/10.3390/nu14173592