
High-Fat Diet and Female Fertility across Lifespan: A Comparative Lesson from Mammal Models

 ,
,  ,
, 
 ,
,  ,
,  ,
,  , , and
, , and 
Abstract
:1. Introduction
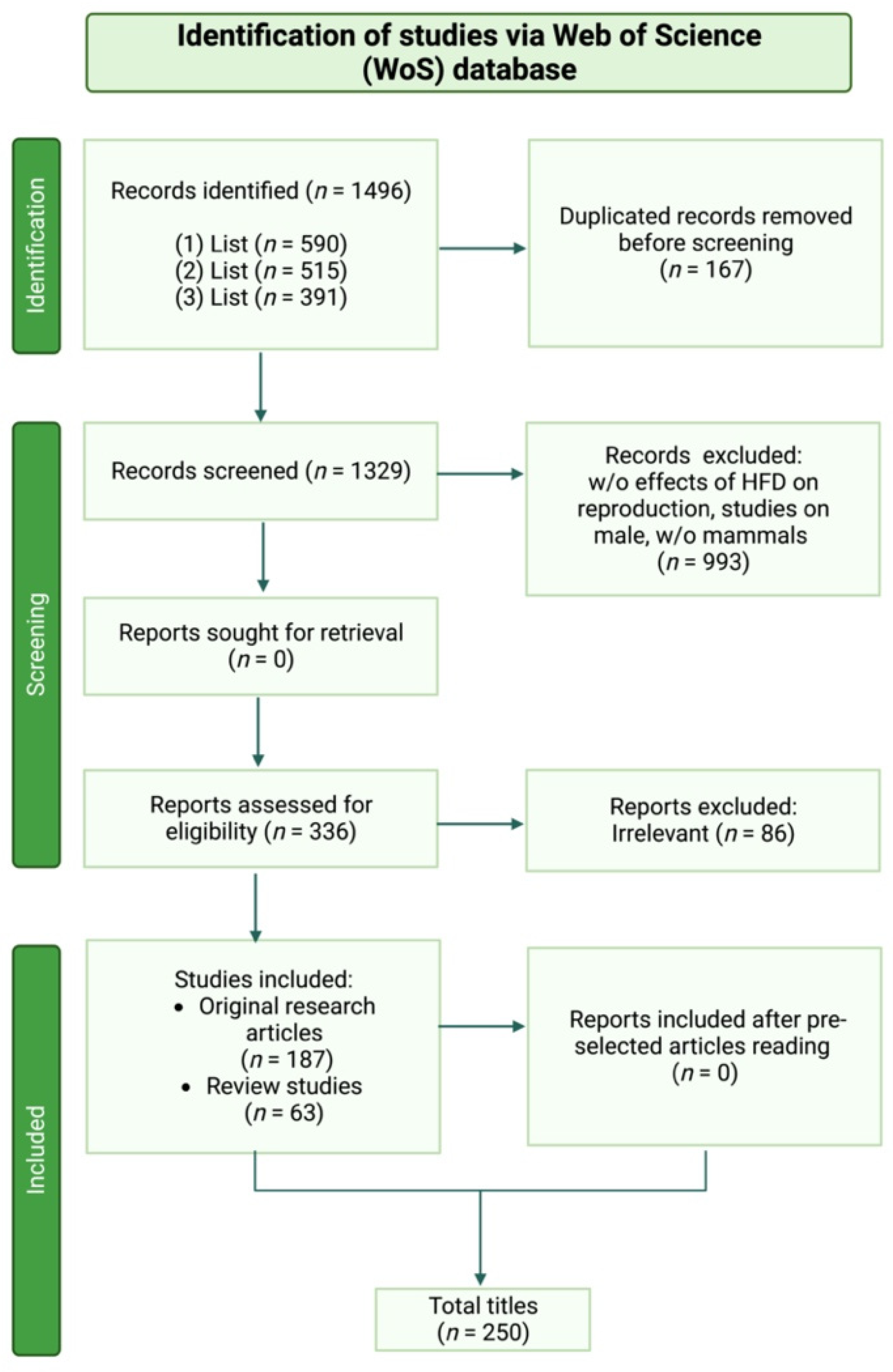
2. Materials and Methods
Bibliographic Search Methods
- (1)
- List: “High fat diet influence on female reproductive endocrine control”
- (2)
- List: “High fat diet affecting ovarian folliculogenesis”
- (3)
- List: “High fat diet impact on oocyte developmental competence”
3. High-Fat Diet Influence on Female Reproductive Endocrine Control
3.1. Transition toward Puberty
3.2. Reproduction in Adulthood
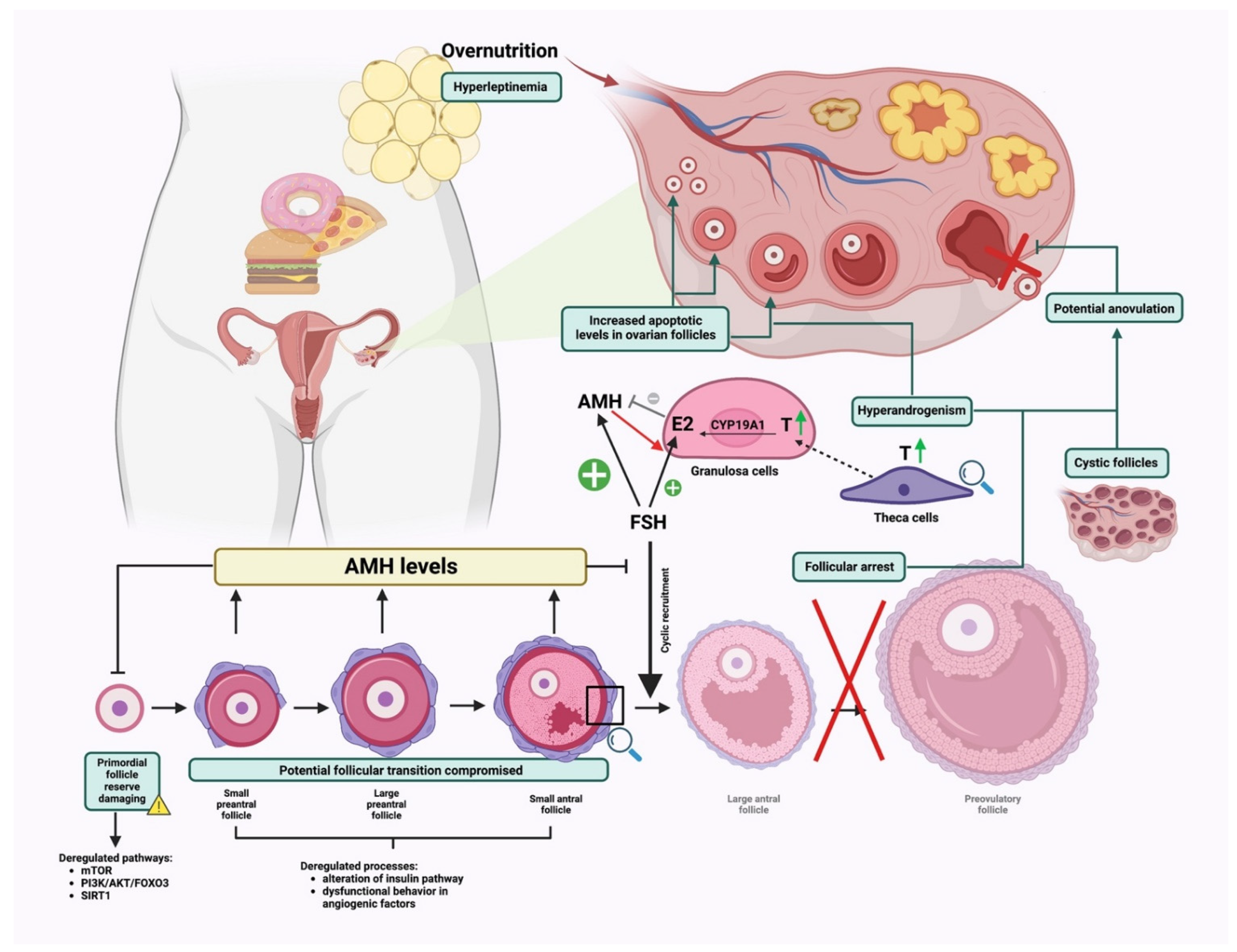
4. High-Fat Diet Affecting Ovarian Folliculogenesis
4.1. Effect on Early Ovarian Folliculogenesis
4.2. Effects on Late Follicle Development under Physiological and Hormonal-Induced Conditions
4.3. Effects on Ovulation Rate
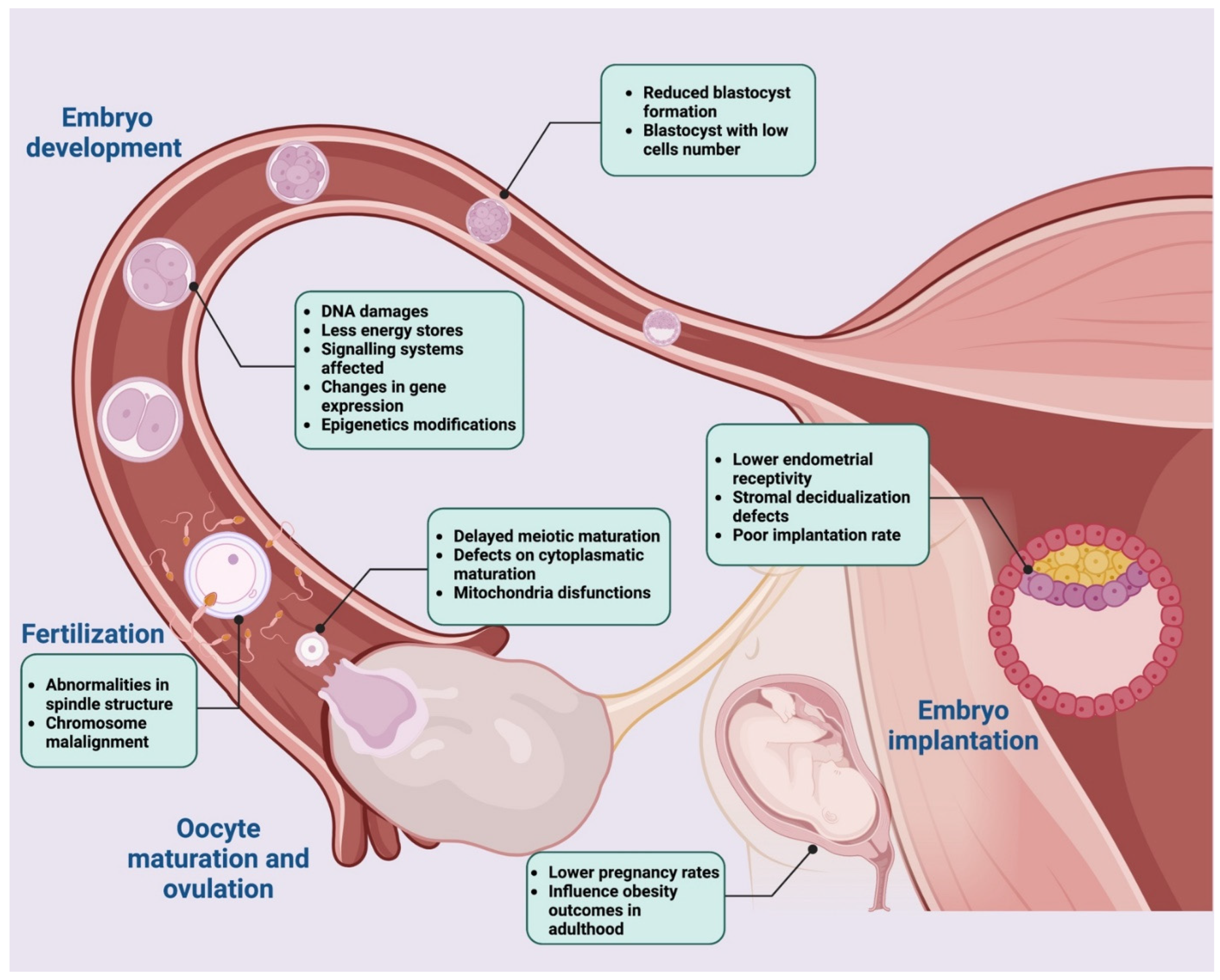
5. High-Fat Diet Impact on Oocyte Developmental Competence
5.1. High-Fat Diet and Oocyte Quality
5.2. High-Fat Diet Affects Oocyte In Vitro Maturation Performance
5.3. Relation between High-Fat Diet and Fertilization Rate
5.4. Effects of High-Fat Diet Composition on Embryonic Implantation and Pregnancy Rate
5.5. High-Fat Diet Impact on Early Embryogenesis
6. Discussion and Conclusions
Overall Sum Up, Key Points and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coss, D. Regulation of reproduction via tight control of gonadotropin hormone levels. Mol. Cell. Endocrinol. 2018, 463, 116–130. [Google Scholar] [CrossRef] [PubMed]
- Gonnella, F.; Konstantinidou, F.; Berardino, C.D.; Capacchietti, G.; Peserico, A.; Russo, V.; Barboni, B.; Stuppia, L.; Gatta, V. A Systematic Review of the Effects of High-Fat Diet Exposure on Oocyte and Follicular Quality: A Molecular Point of View. Int. J. Mol. Sci. 2022, 23, 8890. [Google Scholar] [CrossRef] [PubMed]
- Silvestris, E.; de Pergola, G.; Rosania, R.; Loverro, G. Obesity as disruptor of the female fertility. Reprod. Biol. Endocrinol. 2018, 16, 22. [Google Scholar] [CrossRef] [PubMed]
- Pflanz, N.C.; Daszkowski, A.W.; James, K.A.; Mihic, S.J. Ketone body modulation of ligand-gated ion channels. Neuropharmacology 2019, 148, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Kermack, A.J.; Wellstead, S.J.; Fisk, H.L.; Cheong, Y.; Houghton, F.D.; Macklon, N.S.; Calder, P.C. The Fatty Acid Composition of Human Follicular Fluid Is Altered by a 6-Week Dietary Intervention That Includes Marine Omega-3 Fatty Acids. Lipids 2021, 56, 201–209. [Google Scholar] [CrossRef]
- Hohos, N.M.; Skaznik-Wikiel, M.E. High-Fat Diet and Female Fertility. Endocrinology 2017, 158, 2407–2419. [Google Scholar] [CrossRef] [Green Version]
- Gaskins, A.J.; Chavarro, J.E. Diet and fertility: A review. Am. J. Obstet. Gynecol. 2018, 218, 379–389. [Google Scholar] [CrossRef]
- Richards, J.A.S. The Ovarian Cycle. Vitam. Horm. 2018, 107, 1–25. [Google Scholar] [CrossRef]
- Haisenleder, D.J.; Dalkin, A.C.; Ortolano, G.A.; Marshall, J.C.; Shupnik, M.A. A pulsatile gonadotropin-releasing hormone stimulus is required to increase transcription of the gonadotropin subunit genes: Evidence for differential regulation of transcription by pulse frequency in vivo. Endocrinology 1991, 128, 509–517. [Google Scholar] [CrossRef]
- Marshall, J.C.; Griffin, M.L. The role of changing pulse frequency in the regulation of ovulation. Hum. Reprod. 1993, 8 (Suppl. 2), 57–61. [Google Scholar] [CrossRef]
- Goldsammler, M.; Merhi, Z.; Buyuk, E. Role of hormonal and inflammatory alterations in obesity-related reproductive dysfunction at the level of the hypothalamic-pituitary-ovarian axis. Reprod. Biol. Endocrinol. 2018, 16, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adashi, E.Y. Endocrinology of the ovary. Hum. Reprod. 1994, 9, 815–827. [Google Scholar] [CrossRef]
- Fontana, R.; Della Torre, S. The Deep Correlation between Energy Metabolism and Reproduction: A View on the Effects of Nutrition for Women Fertility. Nutrients 2016, 8, 87. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, S.R.; Shaver, P.R.; Shewchuk, B.M. High Fat Diet Dysregulates Hypothalamic-Pituitary Axis Gene Expression Levels which are Differentially Rescued by EPA and DHA Ethyl Esters. Mol. Nutr. Food Res. 2018, 62, 1800219. [Google Scholar] [CrossRef] [PubMed]
- Rimon-Dahari, N.; Yerushalmi-Heinemann, L.; Alyagor, L.; Dekel, N. Ovarian Folliculogenesis. Results Probl. Cell Differ. 2016, 58, 167–190. [Google Scholar] [CrossRef]
- Monniaux, D. Factors influencing establishment of the ovarian reserve and their effects on fertility. Anim. Reprod. 2018, 15, 635–647. [Google Scholar] [CrossRef]
- Mitchell, M.; Armstrong, D.T.; Robker, R.L.; Norman, R.J. Adipokines: Implications for female fertility and obesity. Reproduction 2005, 130, 583–597. [Google Scholar] [CrossRef] [Green Version]
- Cordier, A.G.; Léveillé, P.; Dupont, C.; Tarrade, A.; Picone, O.; Larcher, T.; Dahirel, M.; Poumerol, E.; Mandon-Pepin, B.; Lévy, R.; et al. Dietary lipid and cholesterol induce ovarian dysfunction and abnormal LH response to stimulation in rabbits. PLoS ONE 2013, 8, e63101. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Chen, H.; Yang, S.; Li, Y.; Wang, B.; Chen, Y.; Wu, X. High-fat diet decreases the expression of Kiss1 mRNA and kisspeptin in the ovary, and increases ovulatory dysfunction in postpubertal female rats. Reprod. Biol. Endocrinol. 2014, 12, 127. [Google Scholar] [CrossRef] [Green Version]
- Bach, À. Effects of nutrition and genetics on fertility in dairy cows. Reprod. Fertil. Dev. 2018, 31, 40–54. [Google Scholar] [CrossRef]
- Cohen, P.; Zhao, C.; Cai, X.; Montez, J.M.; Rohani, S.C.; Feinstein, P.; Mombaerts, P.; Friedman, J.M. Selective deletion of leptin receptor in neurons leads to obesity. J. Clin. Investig. 2001, 108, 1113–1121. [Google Scholar] [CrossRef] [PubMed]
- Nieuwenhuis, D.; Pujol-Gualdo, N.; Arnoldussen, I.A.C.; Kiliaan, A.J. Adipokines: A gear shift in puberty. Obes. Rev. 2020, 21, e13005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broughton, D.E.; Moley, K.H. Obesity and female infertility: Potential mediators of obesity’s impact. Fertil. Steril. 2017, 107, 840–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Lin, Q.; Xu, X.; Illahi, G.S.; Dong, C.; Wu, X. Maternal high-fat diet impairs follicular development of offspring through intraovarian kisspeptin/GPR54 system. Reprod. Biol. Endocrinol. 2019, 17, 13. [Google Scholar] [CrossRef] [Green Version]
- Patel, R.; Shah, G. High-fat diet exposure from pre-pubertal age induces polycystic ovary syndrome (PCOS) in rats. Reproduction 2018, 155, 141–151. [Google Scholar] [CrossRef]
- Roberts, J.S.; Perets, R.A.; Sarfert, K.S.; Bowman, J.J.; Ozark, P.A.; Whitworth, G.B.; Blythe, S.N.; Toporikova, N. High-fat high-sugar diet induces polycystic ovary syndrome in a rodent model. Biol. Reprod. 2017, 96, 551–562. [Google Scholar] [CrossRef] [Green Version]
- Barella, L.F.; De Oliveira, J.C.; Branco, R.C.S.; Camargo, R.L.; Gomes, R.M.; Mendes, F.C.V.; Miranda, R.A.; Gravena, C.; Torrezan, R.; Grassiolli, S.; et al. Early exposure to a high-fat diet has more drastic consequences on metabolism compared with exposure during adulthood in rats. Horm. Metab. Res. 2012, 44, 458–464. [Google Scholar] [CrossRef]
- Newell-Fugate, A.E.; Taibl, J.N.; Alloosh, M.; Sturek, M.; Bahr, J.M.; Nowak, R.A.; Krisher, R.L. Effects of Obesity and Metabolic Syndrome on Steroidogenesis and Folliculogenesis in the Female Ossabaw Mini-Pig. PLoS ONE 2015, 10, e0128749. [Google Scholar] [CrossRef] [Green Version]
- Newell-Fugate, A.E.; Taibl, J.N.; Clark, S.G.; Alloosh, M.; Sturek, M.; Krisher, R.L. Effects of diet-induced obesity on metabolic parameters and reproductive function in female Ossabaw minipigs. Comp. Med. 2014, 64, 44–49. [Google Scholar]
- McGee, W.K.; Bishop, C.V.; Pohl, C.R.; Chang, R.J.; Marshall, J.C.; Pau, F.K.; Stouffer, R.L.; Cameron, J.L. Effects of hyperandrogenemia and increased adiposity on reproductive and metabolic parameters in young adult female monkeys. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E1292–E1304. [Google Scholar] [CrossRef] [Green Version]
- Lalanza, J.F.; Snoeren, E.M.S. The cafeteria diet: A standardized protocol and its effects on behavior. Neurosci. Biobehav. Rev. 2021, 122, 92–119. [Google Scholar] [CrossRef] [PubMed]
- Bazzano, M.V.; Torelli, C.; Pustovrh, M.C.; Paz, D.A.; Elia, E.M. Obesity induced by cafeteria diet disrupts fertility in the rat by affecting multiple ovarian targets. Reprod. Biomed. Online 2015, 31, 655–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bermejo-Alvarez, P.; Rosenfeld, C.S.; Roberts, R.M. Effect of maternal obesity on estrous cyclicity, embryo development and blastocyst gene expression in a mouse model. Hum. Reprod. 2012, 27, 3513–3522. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.L.; Xu, J.J.; Ni, Y.H.; Chen, X.C.; Zhang, H.X.; Zhang, X.M.; Liu, W.J.; Luo, L.L.; Fu, Y.C. SIRT1 activator (SRT1720) improves the follicle reserve and prolongs the ovarian lifespan of diet-induced obesity in female mice via activating SIRT1 and suppressing mTOR signaling. J. Ovarian Res. 2014, 7, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pohlmeier, W.E.; Xie, F.; Kurz, S.G.; Lu, N.; Wood, J.R. Progressive obesity alters the steroidogenic response to ovulatory stimulation and increases the abundance of mRNAs stored in the ovulated oocyte. Mol. Reprod. Dev. 2014, 81, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Hohos, N.M.; Cho, K.J.; Swindle, D.C.; Skaznik-Wikiel, M.E. High-fat diet exposure, regardless of induction of obesity, is associated with altered expression of genes critical to normal ovulatory function. Mol. Cell. Endocrinol. 2018, 470, 199–207. [Google Scholar] [CrossRef]
- Sirotkin, A.V.; Fabian, D.; Babeľová (Kubandová), J.; Vlčková, R.; Alwasel, S.; Harrath, A.H. Body fat affects mouse reproduction, ovarian hormone release, and response to follicular stimulating hormone. Reprod. Biol. 2018, 18, 5–11. [Google Scholar] [CrossRef]
- Clark, K.L.; Talton, O.O.; Ganesan, S.; Schulz, L.C.; Keating, A.F. Developmental origins of ovarian disorder: Impact of maternal lean gestational diabetes on the offspring ovarian proteome in mice. Biol. Reprod. 2019, 101, 771–781. [Google Scholar] [CrossRef]
- Leong, I. Link between maternal obesity and offspring is STELLA. Nat. Rev. Endocrinol. 2018, 14, 189. [Google Scholar] [CrossRef]
- Ge, Z.J.; Luo, S.M.; Lin, F.; Liang, Q.X.; Huang, L.; Wei, Y.C.; Hou, Y.; Han, Z.M.; Schatten, H.; Sun, Q.Y. DNA methylation in oocytes and liver of female mice and their offspring: Effects of high-fat-diet-induced obesity. Environ. Health Perspect. 2014, 122, 159–164. [Google Scholar] [CrossRef] [Green Version]
- Cheong, Y.; Sadek, K.H.; Bruce, K.D.; MacKlon, N.; Cagampang, F.R. Diet-induced maternal obesity alters ovarian morphology and gene expression in the adult mouse offspring. Fertil. Steril. 2014, 102, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Tsoulis, M.W.; Chang, P.E.; Moore, C.J.; Chan, K.A.; Gohir, W.; Petrik, J.J.; Vickers, M.H.; Connor, K.L.; Sloboda, D.M. Maternal High-Fat Diet-Induced Loss of Fetal Oocytes Is Associated with Compromised Follicle Growth in Adult Rat Offspring. Biol. Reprod. 2016, 94, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreas, E.; Reid, M.; Zhang, W.; Moley, K.H. The effect of maternal high-fat/high-sugar diet on offspring oocytes and early embryo development. Mol. Hum. Reprod. 2019, 25, 717–728. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, F.; Xie, J.; Huang, D.; Xie, M. Fetal and neonatal genistein exposure aggravates to interfere with ovarian follicle development of obese female mice induced by high-fat diet. Food Chem. Toxicol. 2020, 135, 110982. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Che, L.; Yang, Z.; Zhang, P.; Shi, J.; Li, J.; Lin, Y.; Fang, Z.; Che, L.; Feng, B.; et al. Effect of High Fat Dietary Intake during Maternal Gestation on Offspring Ovarian Health in a Pig Model. Nutrients 2016, 8, 498. [Google Scholar] [CrossRef] [Green Version]
- Sohrabi, M.; Mohammadi Roushandeh, A.; Alizadeh, Z.; Vahidinia, A.; Vahabian, M.; Hosseini, M. Effect of a high fat diet on ovary morphology, in vitro development, in vitro fertilisation rate and oocyte quality in mice. Singap. Med. J. 2015, 56, 573–579. [Google Scholar] [CrossRef] [Green Version]
- Aiken, C.E.; Tarry-Adkins, J.L.; Penfold, N.C.; Dearden, L.; Ozanne, S.E. Decreased ovarian reserve, dysregulation of mitochondrial biogenesis, and increased lipid peroxidation in female mouse offspring exposed to an obesogenic maternal diet. FASEB J. 2016, 30, 1548–1556. [Google Scholar] [CrossRef] [Green Version]
- Mirihagalle, S.; You, T.; Suh, L.; Patel, C.; Gao, L.; Rattan, S.; Qiao, H. Prenatal exposure to di-(2-ethylhexyl) phthalate and high-fat diet synergistically disrupts mouse fetal oogenesis and affects folliculogenesis. Biol. Reprod. 2019, 100, 1561–1570. [Google Scholar] [CrossRef]
- Howell, K.R.; Powell, T.L. Effects of maternal obesity on placental function and fetal development. Reproduction 2017, 153, R97–R108. [Google Scholar] [CrossRef] [Green Version]
- Merhi, Z.; Du, X.Q.; Charron, M.J. Perinatal exposure to high dietary advanced glycation end products affects the reproductive system in female offspring in mice. Mol. Hum. Reprod. 2020, 26, 615–623. [Google Scholar] [CrossRef]
- King, S.E.; Skinner, M.K. Epigenetic Transgenerational Inheritance of Obesity Susceptibility. Trends Endocrinol. Metab. 2020, 31, 478–494. [Google Scholar] [CrossRef] [PubMed]
- Kaspar, D.; Hastreiter, S.; Irmler, M.; Hrabé de Angelis, M.; Beckers, J. Nutrition and its role in epigenetic inheritance of obesity and diabetes across generations. Mamm. Genome 2020, 31, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Andreas, E.; Winstanley, Y.E.; Robker, R.L. Effect of obesity on the ovarian follicular environment and developmental competence of the oocyte. Curr. Opin. Endocr. Metab. Res. 2021, 18, 152–158. [Google Scholar] [CrossRef]
- Wołodko, K.; Castillo-fernandez, J.; Kelsey, G.; Galvão, A. Revisiting the Impact of Local Leptin Signaling in Folliculogenesis and Oocyte Maturation in Obese Mothers. Int. J. Mol. Sci. 2021, 22, 4270. [Google Scholar] [CrossRef] [PubMed]
- Marei, W.F.A.; Smits, A.; Mohey-Elsaeed, O.; Pintelon, I.; Ginneberge, D.; Bols, P.E.J.; Moerloose, K.; Leroy, J.L.M.R. Differential effects of high fat diet-induced obesity on oocyte mitochondrial functions in inbred and outbred mice. Sci. Rep. 2020, 10, 9806. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, Z.; Liao, X.; Wang, Z. High fat diet triggers cell cycle arrest and excessive apoptosis of granulosa cells during the follicular development. Biochem. Biophys. Res. Commun. 2015, 466, 599–605. [Google Scholar] [CrossRef]
- Ravisankar, S.; Ting, A.Y.; Murphy, M.J.; Redmayne, N.; Wang, D.; McArthur, C.A.; Takahashi, D.L.; Kievit, P.; Chavez, S.L.; Hennebold, J.D. Short-term Western-style diet negatively impacts reproductive outcomes in primates. JCI Insight 2021, 6, e138312. [Google Scholar] [CrossRef]
- Hsueh, A.J.W.; Kawamura, K.; Cheng, Y.; Fauser, B.C.J.M. Intraovarian control of early folliculogenesis. Endocr. Rev. 2015, 36, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Gershon, E.; Dekel, N. Newly Identified Regulators of Ovarian Folliculogenesis and Ovulation. Int. J. Mol. Sci. 2020, 21, 4565. [Google Scholar] [CrossRef]
- Liu, K.; Zhang, H.; Risal, S.; Gorre, N.; Busayavalasa, K.; Li, X.; Shen, Y.; Bosbach, B.; Brännström, M. Somatic cells initiate primordial follicle activation and govern the development of dormant oocytes in mice. Curr. Biol. 2014, 24, 2501–2508. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Lin, F.; Zhao, J.; Wang, Z. Expression Regulation and Physiological Role of Transcription Factor FOXO3a During Ovarian Follicular Development. Front. Physiol. 2020, 11, 1413. [Google Scholar] [CrossRef] [PubMed]
- Long, G.Y.; Yang, J.Y.; Xu, J.J.; Ni, Y.H.; Zhou, X.L.; Ma, J.Y.; Fu, Y.C.; Luo, L.L. SIRT1 knock-in mice preserve ovarian reserve resembling caloric restriction. Gene 2019, 686, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Bordone, L.; Cohen, D.; Robinson, A.; Motta, M.C.; Van Veen, E.; Czopik, A.; Steele, A.D.; Crowe, H.; Marmor, S.; Luo, J.; et al. SIRT1 transgenic mice show phenotypes resembling calorie restriction. Aging Cell 2007, 6, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Du, X.; Zhao, L.; He, M.; Lin, L.; Guo, C.; Zhang, X.; Han, J.; Yan, H.; Huang, K.; et al. SIRT1 facilitates primordial follicle recruitment independent of deacetylase activity through directly modulating Akt1 and mTOR transcription. FASEB J. 2019, 33, 14703–14716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Rajareddy, S.; Reddy, P.; Du, C.; Jagarlamudi, K.; Shen, Y.; Gunnarsson, D.; Selstam, G.; Boman, K.; Liu, K. Infertility caused by retardation of follicular development in mice with oocyte-specific expression of Foxo3a. Development 2007, 134, 199–209. [Google Scholar] [CrossRef] [Green Version]
- John, G.B.; Shirley, L.J.; Gallardo, T.D.; Castrillon, D.G. Specificity of the requirement for Foxo3 in primordial follicle activation. Reproduction 2007, 133, 855–863. [Google Scholar] [CrossRef]
- Castrillon, D.H.; Miao, L.; Kollipara, R.; Horner, J.W.; DePinho, R.A. Suppression of ovarian follicle activation in mice by the transcription factor Foxo3a. Science 2003, 301, 5630. [Google Scholar] [CrossRef]
- Zheng, W.; Nagaraju, G.; Liu, Z.; Liu, K. Functional roles of the phosphatidylinositol 3-kinases (PI3Ks) signaling in the mammalian ovary. Mol. Cell. Endocrinol. 2012, 356, 24–30. [Google Scholar] [CrossRef]
- Brown, C.; LaRocca, J.; Pietruska, J.; Ota, M.; Anderson, L.; Smith, S.D.; Weston, P.; Rasoulpour, T.; Hixon, M.L. Subfertility caused by altered follicular development and oocyte growth in female mice lacking PKB alpha/Akt1. Biol. Reprod. 2010, 82, 246–256. [Google Scholar] [CrossRef] [Green Version]
- Hilal, G.; Fatma, T.; Ferruh, Y.; Sabire, G.; Yüksel, A. Effect of high-fat diet on the various morphological parameters of the ovary. Anat. Cell Biol. 2020, 53, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Luo, L.L.; Xu, J.J.; Xu, M.Y.; Zhang, X.M.; Zhou, X.L.; Liu, W.J.; Fu, Y.C. Obesity accelerates ovarian follicle development and follicle loss in rats. Metabolism 2014, 63, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.L.; Chen, X.C.; Fu, Y.C.; Xu, J.J.; Li, L.; Lin, X.H.; Xiang, Y.F.; Zhang, X.M. The effects of caloric restriction and a high-fat diet on ovarian lifespan and the expression of SIRT1 and SIRT6 proteins in rats. Aging Clin. Exp. Res. 2012, 24, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Nteeba, J.; Ross, J.W.; Perfield, J.W.; Keating, A.F. High fat diet induced obesity alters ovarian phosphatidylinositol-3 kinase signaling gene expression. Reprod. Toxicol. 2013, 42, 68–77. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Zhang, Z.; Liao, X.; Qi, L.; Liu, Y.; Wang, Z. Effect of high-fat diet-induced obesity on the Akt/FoxO/Smad signaling pathway and the follicular development of the mouse ovary. Mol. Med. Rep. 2016, 14, 3894–3900. [Google Scholar] [CrossRef] [Green Version]
- Fullston, T.; Shehadeh, H.; Sandeman, L.Y.; Kang, W.X.; Wu, L.L.; Robker, R.L.; McPherson, N.O.; Lane, M. Female offspring sired by diet induced obese male mice display impaired blastocyst development with molecular alterations to their ovaries, oocytes and cumulus cells. J. Assist. Reprod. Genet. 2015, 32, 725–735. [Google Scholar] [CrossRef] [Green Version]
- Asemota, O.; Thornton, K.; Merhi, Z.; Berger, D.S.; Williams, L.; Seki, Y.; Du, X.Q.; Jindal, S.; Charron, M.J.; Buyuk, E. Monocyte chemotactic protein-1 plays a role in ovarian dysfunction related to high-fat diet-induced obesity. Syst. Biol. Reprod. Med. 2020, 66, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Merhi, Z.; Thornton, K.; Bonney, E.; Cipolla, M.J.; Charron, M.J.; Buyuk, E. Ovarian kisspeptin expression is related to age and to monocyte chemoattractant protein-1. J. Assist. Reprod. Genet. 2016, 33, 535–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senosy, W.; Mahmoud, G.B.; Abdel-Raheem, S.M. Influence of short-term energy supplementation on estrus, ovarian activity, and blood biochemistry in Ossimi ewes synchronized with fluorogestone acetate in the subtropics. Theriogenology 2017, 88, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Raviv, S.; Hantisteanu, S.; Sharon, S.M.; Atzmon, Y.; Michaeli, M.; Shalom-Paz, E. Lipid droplets in granulosa cells are correlated with reduced pregnancy rates. J. Ovarian Res. 2020, 13, 4. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, S.; Wang, B.; Wei, H.; Liu, X.; Hao, J.; Duan, Y.; Hua, J.; Zheng, X.; Feng, X.; et al. High-fat-diet impaired mitochondrial function of cumulus cells but improved the efficiency of parthenogenetic embryonic quality in mice. Anim. Cells Syst. (Seoul) 2018, 22, 243–252. [Google Scholar] [CrossRef]
- Xia, H.; Zhang, R.; Guan, H.; Zhang, W. Follicle loss and PTEN/PI3K/mTOR signaling pathway activated in LepR-mutated mice. Gynecol. Endocrinol. 2019, 35, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Panwar, S.; Herrid, M.; Kauter, K.G.; McFarlane, J.R. Effect of passive immunization against leptin on ovarian follicular development in prepubertal mice. J. Reprod. Immunol. 2012, 96, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Wołodko, K.; Walewska, E.; Adamowski, M.; Castillo-Fernandez, J.; Kelsey, G.; Galvão, A. Leptin Resistance in the Ovary of Obese Mice is Associated with Profound Changes in the Transcriptome of Cumulus Cells. Cell. Physiol. Biochem. 2020, 54, 417–437. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.A.; Carpenter, R.M.; Blythe, S.N.; Toporikova, N. FSH/AMH Ratio and Adipocyte Size are Linked to Ovarian Dysfunction. Endocr. Res. 2020, 45, 174–189. [Google Scholar] [CrossRef]
- Shibahara, H.; Ishiguro, A.; Inoue, Y.; Koumei, S.; Kuwayama, T.; Iwata, H. Mechanism of palmitic acid-induced deterioration of in vitro development of porcine oocytes and granulosa cells. Theriogenology 2020, 141, 54–61. [Google Scholar] [CrossRef]
- Bishop, C.V.; Reiter, T.E.; Erikson, D.W.; Hanna, C.B.; Daughtry, B.L.; Chavez, S.L.; Hennebold, J.D.; Stouffer, R.L. Chronically elevated androgen and/or consumption of a Western-style diet impairs oocyte quality and granulosa cell function in the nonhuman primate periovulatory follicle. J. Assist. Reprod. Genet. 2019, 36, 1497–1511. [Google Scholar] [CrossRef]
- Bishop, C.V.; Xu, F.; Xu, J.; Ting, A.Y.; Galbreath, E.; McGee, W.K.; Zelinski, M.B.; Hennebold, J.D.; Cameron, J.L.; Stouffer, R.L. Western-style diet, with and without chronic androgen treatment, alters the number, structure, and function of small antral follicles in ovaries of young adult monkeys. Fertil. Steril. 2016, 105, 1023–1034. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.S.K.; Shikanov, A. Follicle development as an orchestrated signaling network in a 3D organoid. J. Biol. Eng. 2019, 13, 2. [Google Scholar] [CrossRef]
- Murabayashi, N.; Sugiyama, T.; Zhang, L.; Kamimoto, Y.; Umekawa, T.; Ma, N.; Sagawa, N. Maternal high-fat diets cause insulin resistance through inflammatory changes in fetal adipose tissue. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 169, 39–44. [Google Scholar] [CrossRef]
- Dupont, J.; Scaramuzzi, R.J. Insulin signalling and glucose transport in the ovary and ovarian function during the ovarian cycle. Biochem. J. 2016, 473, 1483–1501. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, C.M.; Emanuelli, B.; Kahn, C.R. Critical nodes in signalling pathways: Insights into insulin action. Nat. Rev. Mol. Cell Biol. 2006, 7, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Samoto, T.; Maruo, T.; Cecilia, A.L.L.; Matsuo, H.; Deguchi, J.; Barnea, E.R. Insulin receptor expression in follicular and stromal compartments of the human ovary over the course of follicular growth, regression and atresia. Endocr. J. 1993, 40, 715–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Roeiy, A.; Chen, X.; Robertss, V.J.; Shimasaki, S.; Ling, N.; Leroith, D.; Roberts, C.T.; Yen, S.S.C. Expression of the genes encoding the insulin-like growth factors (IGF-I and II), the IGF and insulin receptors, and IGF-binding proteins-1-6 and the localization of their gene products in normal and polycystic ovary syndrome ovaries. J. Clin. Endocrinol. Metab. 1994, 78, 1488–1496. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.K.; Chou, C.H.; Huang, C.C.; Wen, W.F.; Chen, H.F.; Shun, C.T.; Ho, H.N.; Chen, M.J. Obesity alters ovarian folliculogenesis through disrupted angiogenesis from increased IL-10 production. Mol. Metab. 2021, 49, 101189. [Google Scholar] [CrossRef]
- Hohos, N.M.; Elliott, E.M.; Cho, K.J.; Lin, I.S.; Rudolph, M.C.; Skaznik-Wikiel, M.E. High-fat diet-induced dysregulation of ovarian gene expression is restored with chronic omega-3 fatty acid supplementation. Mol. Cell. Endocrinol. 2020, 499, 110615. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Anderson, C.L.; Timme, K.R.; Kurz, S.G.; Fernando, S.C.; Wood, J.R. Obesity-dependent increases in oocyte mRNAs are associated with increases in proinflammatory signaling and gut microbial abundance of lachnospiraceae in female mice. Endocrinology 2016, 157, 1630–1643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, M.O.; Sharma, S.; Kim, S.; Rickert, E.; Hsueh, K.; Hwang, V.; Olefsky, J.M.; Webster, N.J.G. Obese Neuronal PPARγ Knockout Mice Are Leptin Sensitive but Show Impaired Glucose Tolerance and Fertility. Endocrinology 2017, 158, 121–133. [Google Scholar] [CrossRef]
- Ma, T.; Cui, P.; Tong, X.; Hu, W.; Shao, L.R.; Zhang, F.; Li, X.; Feng, Y. Endogenous Ovarian Angiogenesis in Polycystic Ovary Syndrome-Like Rats Induced by Low-Frequency Electro-Acupuncture: The CLARITY Three-Dimensional Approach. Int. J. Mol. Sci. 2018, 19, 3500. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Hayes, E.; Prizant, H.; Srivastava, R.K.; Hammes, S.R.; Sen, A. Leptin-Induced CART (Cocaine- and Amphetamine-Regulated Transcript) Is a Novel Intraovarian Mediator of Obesity-Related Infertility in Females. Endocrinology 2016, 157, 1248–1257. [Google Scholar] [CrossRef]
- Jia, Z.; Feng, Z.; Wang, L.; Li, H.; Wang, H.; Xu, D.; Zhao, X.; Feng, D.; Feng, X. Resveratrol reverses the adverse effects of a diet-induced obese murine model on oocyte quality and zona pellucida softening. Food Funct. 2018, 9, 2623–2633. [Google Scholar] [CrossRef]
- Yang, L.; Chen, L.; Lu, X.; Tan, A.; Chen, Y.; Li, Y.; Peng, X.; Yuan, S.; Cai, D.; Yu, Y. Peri-ovarian adipose tissue contributes to intraovarian control during folliculogenesis in mice. Reproduction 2018, 156, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N.; Frantz, G.; LeCouter, J.; Dillard-Telm, L.; Pham, T.; Draksharapu, A.; Giordano, T.; Peale, F. Differential expression of the angiogenic factor genes vascular endothelial growth factor (VEGF) and endocrine gland-derived VEGF in normal and polycystic human ovaries. Am. J. Pathol. 2003, 162, 1881–1893. [Google Scholar] [CrossRef] [Green Version]
- Kamat, B.R.; Brown, L.F.; Manseau, E.J.; Senger, D.R.; Dvorak, H.F. Expression of vascular permeability factor/vascular endothelial growth factor by human granulosa and theca lutein cells. Role in corpus luteum development. Am. J. Pathol. 1995, 146, 157–165. [Google Scholar]
- Artini, P.G.; Ruggiero, M.; Parisen Toldin, M.R.; Monteleone, P.; Monti, M.; Cela, V.; Genazzani, A.R. Vascular endothelial growth factor and its soluble receptor in patients with polycystic ovary syndrome undergoing IVF. Hum. Fertil. (Camb.) 2009, 12, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Scotti, L.; Parborell, F.; Irusta, G.; De Zuñiga, I.; Bisioli, C.; Pettorossi, H.; Tesone, M.; Abramovich, D. Platelet-derived growth factor BB and DD and angiopoietin1 are altered in follicular fluid from polycystic ovary syndrome patients. Mol. Reprod. Dev. 2014, 81, 748–756. [Google Scholar] [CrossRef]
- Tal, R.; Seifer, D.B.; Shohat-Tal, A.; Grazi, R.V.; Malter, H.E. Transforming growth factor-β1 and its receptor soluble endoglin are altered in polycystic ovary syndrome during controlled ovarian stimulation. Fertil. Steril. 2013, 100, 538–543. [Google Scholar] [CrossRef]
- Raja-Khan, N.; Kunselman, A.R.; Demers, L.M.; Ewens, K.G.; Spielman, R.S.; Legro, R.S. A variant in the fibrillin-3 gene is associated with TGF-β and inhibin B levels in women with polycystic ovary syndrome. Fertil. Steril. 2010, 94, 2916–2919. [Google Scholar] [CrossRef] [Green Version]
- Artini, P.G.; Monti, M.; Matteucci, C.; Valentino, V.; Cristello, F.; Genazzani, A.R. Vascular endothelial growth factor and basic fibroblast growth factor in polycystic ovary syndrome during controlled ovarian hyperstimulation. Gynecol. Endocrinol. 2006, 22, 465–470. [Google Scholar] [CrossRef]
- Tal, R.; Seifer, D.B.; Grazi, R.V.; Malter, H.E. Angiopoietin-1 and angiopoietin-2 are altered in polycystic ovarian syndrome (PCOS) during controlled ovarian stimulation. Vasc. Cell 2013, 5, 18. [Google Scholar] [CrossRef] [Green Version]
- Tal, R.; Seifer, D.B.; Grazi, R.V.; Malter, H.E. Follicular fluid placental growth factor is increased in polycystic ovarian syndrome: Correlation with ovarian stimulation. Reprod. Biol. Endocrinol. 2014, 12, 82. [Google Scholar] [CrossRef] [Green Version]
- Tal, R.; Seifer, D.B.; Arici, A. The emerging role of angiogenic factor dysregulation in the pathogenesis of polycystic ovarian syndrome. Semin. Reprod. Med. 2015, 33, 195–207. [Google Scholar] [CrossRef] [PubMed]
- McFee, R.M.; Romereim, S.M.; Snider, A.P.; Summers, A.F.; Pohlmeier, W.E.; Kurz, S.G.; Cushman, R.A.; Davis, J.S.; Wood, J.R.; Cupp, A.S. A high-androgen microenvironment inhibits granulosa cell proliferation and alters cell identity. Mol. Cell. Endocrinol. 2021, 531, 111288. [Google Scholar] [CrossRef] [PubMed]
- Bradford, A.P.; Jones, K.; Kechris, K.; Chosich, J.; Montague, M.; Warren, W.C.; May, M.C.; Al-Safi, Z.; Kuokkanen, S.; Appt, S.E.; et al. Joint MiRNA/mRNA expression profiling reveals changes consistent with development of dysfunctional corpus luteum after weight gain. PLoS ONE 2015, 10, e0135163. [Google Scholar] [CrossRef] [Green Version]
- Fabian, D.; Babeľová, J.; Čikoš, Š.; Šefčíková, Z. Overweight negatively affects outcome of superovulation treatment in female mice. Zygote 2017, 25, 751–759. [Google Scholar] [CrossRef]
- Smits, A.; Marei, W.F.A.; De Neubourg, D.; Leroy, J.L.M.R. Diet normalization or caloric restriction as a preconception care strategy to improve metabolic health and oocyte quality in obese outbred mice. Reprod. Biol. Endocrinol. 2021, 19, 166. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Li, Y.; Liao, X.; Wang, Z.; Li, R.; Zou, S.; Jiang, T.; Zheng, B.; Duan, P.; Xiao, J. Diabetes Induces Abnormal Ovarian Function via Triggering Apoptosis of Granulosa Cells and Suppressing Ovarian Angiogenesis. Int. J. Biol. Sci. 2017, 13, 1297–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, Y.; Wei, Z.; Hunter, N. Maternal obesity enhances oocyte chromosome abnormalities associated with aging. Chromosoma 2019, 128, 413–421. [Google Scholar] [CrossRef]
- Wallace, J.; Milne, J.S.; Adam, C.L.; Aitken, R.P. Impact of donor and recipient adiposity on placental and fetal growth in adolescent sheep. Reproduction 2017, 153, 381–394. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.J.; Yang, W.S.; Chen, C.L.; Wu, M.Y.; Yang, Y.S.; Ho, H.N. The relationship between anti-Mullerian hormone, androgen and insulin resistance on the number of antral follicles in women with polycystic ovary syndrome. Hum. Reprod. 2008, 23, 952–957. [Google Scholar] [CrossRef] [Green Version]
- Dewailly, D.; Barbotin, A.L.; Dumont, A.; Catteau-Jonard, S.; Robin, G. Role of Anti-Müllerian Hormone in the Pathogenesis of Polycystic Ovary Syndrome. Front. Endocrinol. 2020, 11, 641. [Google Scholar] [CrossRef]
- Adamowski, M.; Wołodko, K.; Oliveira, J.; Castillo-Fernandez, J.; Murta, D.; Kelsey, G.; Galvão, A.M. Leptin Signaling in the Ovary of Diet-Induced Obese Mice Regulates Activation of NOD-Like Receptor Protein 3 Inflammasome. Front. Cell Dev. Biol. 2021, 9, 738731. [Google Scholar] [CrossRef] [PubMed]
- Hohos, N.M.; Elliott, E.M.; Giornazi, A.; Silva, E.; Rice, J.D.; Skaznik-Wikiel, M.E. High-fat diet induces an ovulatory defect associated with dysregulated endothelin-2 in mice. Reproduction 2021, 161, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Gambineri, A.; Laudisio, D.; Marocco, C.; Radellini, S.; Colao, A.; Savastano, S. Female infertility: Which role for obesity? Int. J. Obes. Suppl. 2019, 9, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Hohos, N.M.; Cho, K.; Swindle, D.; Polotsky, A.J.; Skaznik-Wikiel, M.E. Expression of genes governing ovulation and steroidogenesis is affected by high fat diet irrespective of obesity. Fertil. Steril. 2017, 108, e14. [Google Scholar] [CrossRef] [Green Version]
- Li, F.X.; Liu, Y.; Miao, X.P.; Fu, G.Q.; Curry, T.E. Expression and regulation of the differentiation regulators ERBB Receptor Feedback Inhibitor 1 (ERRFI1) and Interferon-related Developmental Regulator 1 (IFRD1) during the periovulatory period in the rat ovary. Mol. Reprod. Dev. 2016, 83, 714–723. [Google Scholar] [CrossRef]
- Brown, H.M.; Dunning, K.R.; Robker, R.L.; Pritchard, M.; Russell, D.L. Requirement for ADAMTS-1 in extracellular matrix remodeling during ovarian folliculogenesis and lymphangiogenesis. Dev. Biol. 2006, 300, 699–709. [Google Scholar] [CrossRef]
- Lim, H.; Paria, B.C.; Das, S.K.; Dinchuk, J.E.; Langenbach, R.; Trzaskos, J.M.; Dey, S.K. Multiple female reproductive failures in cyclooxygenase 2-deficient mice. Cell 1997, 91, 197–208. [Google Scholar] [CrossRef] [Green Version]
- Dahm-Kähler, P.; Ghahremani, M.; Lind, A.K.; Sundfeldt, K.; Brännström, M. Monocyte chemotactic protein-1 (MCP-1), its receptor, and macrophages in the perifollicular stroma during the human ovulatory process. Fertil. Steril. 2009, 91, 231–239. [Google Scholar] [CrossRef]
- Kanda, H.; Tateya, S.; Tamori, Y.; Kotani, K.; Hiasa, K.I.; Kitazawa, R.; Kitazawa, S.; Miyachi, H.; Maeda, S.; Egashira, K.; et al. MCP-1 contributes to macrophage infiltration into adipose tissue, insulin resistance, and hepatic steatosis in obesity. J. Clin. Investig. 2006, 116, 1494–1505. [Google Scholar] [CrossRef]
- Gonzalez, M.B.; Lane, M.; Knight, E.J.; Robker, R.L. Inflammatory markers in human follicular fluid correlate with lipid levels and Body Mass Index. J. Reprod. Immunol. 2018, 130, 25–29. [Google Scholar] [CrossRef]
- Dunning, K.R.; Russell, D.L.; Robker, R.L. Lipids and oocyte developmental competence: The role of fatty acids and β-oxidation. Reproduction 2014, 148, R15–R27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boudoures, A.L.; Chi, M.; Thompson, A.; Zhang, W.; Moley, K.H. The effects of voluntary exercise on oocyte quality in a diet-induced obese murine model. Reproduction 2016, 151, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Dunning, K.R.; Anastasi, M.R.; Zhang, V.J.; Russell, D.L.; Robker, R.L. Regulation of Fatty Acid Oxidation in Mouse Cumulus-Oocyte Complexes during Maturation and Modulation by PPAR Agonists. PLoS ONE 2014, 9, e87327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradley, J.; Swann, K. Mitochondria and lipid metabolism in mammalian oocytes and early embryos. Int. J. Dev. Biol. 2019, 63, 93–103. [Google Scholar] [CrossRef] [Green Version]
- Greaney, J.; Wei, Z.; Homer, H. Regulation of chromosome segregation in oocytes and the cellular basis for female meiotic errors. Hum. Reprod. Update 2018, 24, 135–161. [Google Scholar] [CrossRef]
- Li, R.; Albertini, D.F. The road to maturation: Somatic cell interaction and self-organization of the mammalian oocyte. Nat. Rev. Mol. Cell Biol. 2013, 14, 141–152. [Google Scholar] [CrossRef]
- Homer, H.A. The Role of Oocyte Quality in Explaining “Unexplained” Infertility. Semin. Reprod. Med. 2020, 38, 21–28. [Google Scholar] [CrossRef]
- Cimadomo, D.; Fabozzi, G.; Vaiarelli, A.; Ubaldi, N.; Ubaldi, F.M.; Rienzi, L. Impact of maternal age on oocyte and embryo competence. Front. Endocrinol. 2018, 9, 327. [Google Scholar] [CrossRef] [Green Version]
- He, M.; Zhang, T.; Yang, Y.; Wang, C. Mechanisms of Oocyte Maturation and Related Epigenetic Regulation. Front. Cell Dev. Biol. 2021, 9, 596. [Google Scholar] [CrossRef]
- Fontana, J.; Martínková, S.; Petr, J.; Žalmanová, T.; Trnka, J. Metabolic cooperation in the ovarian follicle. Physiol. Res. 2020, 69, 33–48. [Google Scholar] [CrossRef]
- Monniaux, D. Driving folliculogenesis by the oocyte-somatic cell dialog: Lessons from genetic models. Theriogenology 2016, 86, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Da Broi, M.G.; Giorgi, V.S.I.; Wang, F.; Keefe, D.L.; Albertini, D.; Navarro, P.A. Influence of follicular fluid and cumulus cells on oocyte quality: Clinical implications. J. Assist. Reprod. Genet. 2018, 35, 735–751. [Google Scholar] [CrossRef] [PubMed]
- Fortune, J.E. Ovarian Follicular Growth and Development in Mammals. Biol. Reprod. 1994, 50, 225–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revelli, A.; Piane, L.D.; Casano, S.; Molinari, E.; Massobrio, M.; Rinaudo, P. Follicular fluid content and oocyte quality: From single biochemical markers to metabolomics. Reprod. Biol. Endocrinol. 2009, 7, 40. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, N.; Asada, R.; Saito, A.; Kanemoto, S.; Imaizumi, K. Obesity-induced endoplasmic reticulum stress causes chronic inflammation in adipose tissue. Sci. Rep. 2012, 2, 799. [Google Scholar] [CrossRef] [Green Version]
- Kirillova, A.; Smitz, J.E.J.; Sukhikh, G.T.; Mazunin, I. The Role of Mitochondria in Oocyte Maturation. Cells 2021, 10, 2484. [Google Scholar] [CrossRef]
- Kazemi, A.; Ramezanzadeh, F.; Nasr-Esfahani, M.H.; Saboor Yaraghi, A.A.; Ahmadi, M. Does dietary fat intake influence oocyte competence and embryo quality by inducing oxidative stress in follicular fluid? Iran. J. Reprod. Med. 2013, 11, 1005–1012. [Google Scholar]
- Song, J.; Xiang, S.; Pang, C.; Guo, J.; Sun, Z. Metabolomic alternations of follicular fluid of obese women undergoing in-vitro fertilization treatment. Sci. Rep. 2020, 10, 5968. [Google Scholar] [CrossRef] [Green Version]
- Yan, S.; Wang, F.; Shi, Q. The effect of maternal high-fat-diet mediated oxidative stress on ovarian function in mice offspring. Exp. Ther. Med. 2020, 20, 135. [Google Scholar] [CrossRef]
- Leroy, J.L.M.R.; Vanholder, T.; Mateusen, B.; Christophe, A.; Opsomer, G.; de Kruif, A.; Genicot, G.; Van Soom, A. Non-esterified fatty acids in follicular fluid of dairy cows and their effect on developmental capacity of bovine oocytes in vitro. Reproduction 2005, 130, 485–495. [Google Scholar] [CrossRef] [Green Version]
- Igosheva, N.; Abramov, A.Y.; Poston, L.; Eckert, J.J.; Fleming, T.P.; Duchen, M.R.; McConnell, J. Maternal diet-induced obesity alters mitochondrial activity and redox status in mouse oocytes and zygotes. PLoS ONE 2010, 5, e10074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valckx, S.D.M.; De Pauw, I.; De Neubourg, D.; Inion, I.; Berth, M.; Fransen, E.; Bols, P.E.J.; Leroy, J.L.M.R. BMI-related metabolic composition of the follicular fluid of women undergoing assisted reproductive treatment and the consequences for oocyte and embryo quality. Hum. Reprod. 2012, 27, 3531–3539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valckx, S.D.M.; Van Hoeck, V.; Arias-Alvarez, M.; Maillo, V.; Lopez-Cardona, A.P.; Gutierrez-Adan, A.; Berth, M.; Cortvrindt, R.; Bols, P.E.J.; Leroy, J.L.M.R. Elevated non-esterified fatty acid concentrations during in vitro murine follicle growth alter follicular physiology and reduce oocyte developmental competence. Fertil. Steril. 2014, 102, 1769–1776.e1. [Google Scholar] [CrossRef] [PubMed]
- Wakefield, S.L.; Lane, M.; Schulz, S.J.; Hebart, M.L.; Thompson, J.G.; Mitchell, M. Maternal supply of omega-3 polyunsaturated fatty acids alter mechanisms involved in oocyte and early embryo development in the mouse. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E425–E434. [Google Scholar] [CrossRef]
- Wu, L.L.Y.; Dunning, K.R.; Yang, X.; Russell, D.L.; Lane, M.; Norman, R.J.; Robker, R.L. High-fat diet causes lipotoxicity responses in cumulus-oocyte complexes and decreased fertilization rates. Endocrinology 2010, 151, 5438–5445. [Google Scholar] [CrossRef]
- Chen, Z.; Lei, L.; Wen, D.; Yang, L. Melatonin attenuates palmitic acid-induced mouse granulosa cells apoptosis via endoplasmic reticulum stress. J. Ovarian Res. 2019, 12, 43. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Wu, L.L.; Chura, L.R.; Liang, X.; Lane, M.; Norman, R.J.; Robker, R.L. Exposure to lipid-rich follicular fluid is associated with endoplasmic reticulum stress and impaired oocyte maturation in cumulus-oocyte complexes. Fertil. Steril. 2012, 97, 1438–1443. [Google Scholar] [CrossRef]
- Mu, Y.M.; Yanase, T.; Nishi, Y.; Tanaka, A.; Saito, M.; Jin, C.H.; Mukasa, C.; Okabe, T.; Nomura, M.; Goto, K.; et al. Saturated FFAs, palmitic acid and stearic acid, induce apoptosis in human granulosa cells. Endocrinology 2001, 142, 3590–3597. [Google Scholar] [CrossRef]
- Vanholder, T.; Lmr Leroy, J.; Van Soom, A.; Maes, D.; Coryn, M.; Fiers, T.; De Kruif, A.; Opsomer, G. Effect of non-esterified fatty acids on bovine theca cell steroidogenesis and proliferation in vitro. Anim. Reprod. Sci. 2006, 92, 51–63. [Google Scholar] [CrossRef]
- Wu, L.L.; Russell, D.L.; Norman, R.J.; Robker, R.L. Endoplasmic reticulum (ER) stress in cumulus-oocyte complexes impairs pentraxin-3 secretion, mitochondrial membrane potential (DeltaPsi m), and embryo development. Mol. Endocrinol. 2012, 26, 562–573. [Google Scholar] [CrossRef] [Green Version]
- Aardema, H.; Lolicato, F.; van de Lest, C.H.A.; Brouwers, J.F.; Vaandrager, A.B.; van Tol, H.T.A.; Roelen, B.A.J.; Vos, P.L.A.M.; Helms, J.B.; Gadella, B.M. Bovine cumulus cells protect maturing oocytes from increased fatty acid levels by massive intracellular lipid storage. Biol. Reprod. 2013, 88, 164. [Google Scholar] [CrossRef] [PubMed]
- Sutton-McDowall, M.L.; Wu, L.L.Y.; Purdey, M.; Abell, A.D.; Goldys, E.M.; MacMillan, K.L.; Thompson, J.G.; Robker, R.L. Nonesterified Fatty Acid-Induced Endoplasmic Reticulum Stress in Cattle Cumulus Oocyte Complexes Alters Cell Metabolism and Developmental Competence. Biol. Reprod. 2016, 94, 23. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Baddela, V.S.; Becker, F.; Dannenberger, D.; Viergutz, T.; Vanselow, J. Elevated free fatty acids affect bovine granulosa cell function: A molecular cue for compromised reproduction during negative energy balance. Endocr. Connect. 2019, 8, 493–505. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, C.; Li, J.; Wang, G.; Li, L. Non-Esterified Fatty Acid-Induced Reactive Oxygen Species Mediated Granulosa Cells Apoptosis Is Regulated by Nrf2/p53 Signaling Pathway. Antioxidants 2020, 9, 523. [Google Scholar] [CrossRef] [PubMed]
- Yenuganti, V.R.; Koczan, D.; Vanselow, J. Genome wide effects of oleic acid on cultured bovine granulosa cells: Evidence for the activation of pathways favoring folliculo-luteal transition. BMC Genom. 2021, 22, 486. [Google Scholar] [CrossRef] [PubMed]
- Aardema, H.; Vos, P.L.A.M.; Lolicato, F.; Roelen, B.A.J.; Knijn, H.M.; Vaandrager, A.B.; Helms, B.J.; Gadella, B.M. Oleic acid prevents detrimental effects of saturated fatty acids on bovine oocyte developmental competence. Biol. Reprod. 2011, 85, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Carro, M.; Buschiazzo, J.; Ríos, G.L.; Oresti, G.M.; Alberio, R.H. Linoleic acid stimulates neutral lipid accumulation in lipid droplets of maturing bovine oocytes. Theriogenology 2013, 79, 687–694. [Google Scholar] [CrossRef]
- Zachut, M.; Dekel, I.; Lehrer, H.; Arieli, A.; Arav, A.; Livshitz, L.; Yakoby, S.; Moallem, U. Effects of dietary fats differing in n-6:n-3 ratio fed to high-yielding dairy cows on fatty acid composition of ovarian compartments, follicular status, and oocyte quality. J. Dairy Sci. 2010, 93, 529–545. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, K.; Itami, N.; Ueda, M.; Kansaku, K.; Shirasuna, K.; Kuwayama, T.; Iwata, H. Non-esterified fatty acid-associated ability of follicular fluid to support porcine oocyte maturation and development. Reprod. Med. Biol. 2018, 17, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Pawlak, P.; Malyszka, N.; Szczerbal, I.; Kolodziejski, P. Fatty acid induced lipolysis influences embryo development, gene expression and lipid droplet formation in the porcine cumulus cells. Biol. Reprod. 2020, 103, 36–48. [Google Scholar] [CrossRef] [Green Version]
- Itami, N.; Shirasuna, K.; Kuwayama, T.; Iwata, H. Palmitic acid induces ceramide accumulation, mitochondrial protein hyperacetylation, and mitochondrial dysfunction in porcine oocytes. Biol. Reprod. 2018, 98, 644–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wonnacott, K.E.; Kwong, W.Y.; Hughes, J.; Salter, A.M.; Lea, R.G.; Garnsworthy, P.C.; Sinclair, K.D. Dietary omega-3 and -6 polyunsaturated fatty acids affect the composition and development of sheep granulosa cells, oocytes and embryos. Reproduction 2010, 139, 57–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamayou, S. Oocyte quality in assisted reproduction techniques. Minerva Endocrinol. 2022, 47, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Warzych, E.; Lipinska, P. Energy metabolism of follicular environment during oocyte growth and maturation. J. Reprod. Dev. 2020, 66, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Hoeck, V.; Sturmey, R.G.; Bermejo-Alvarez, P.; Rizos, D.; Gutierrez-Adan, A.; Leese, H.J.; Bols, P.E.J.; Leroy, J.L.M.R. Elevated non-esterified fatty acid concentrations during bovine oocyte maturation compromise early embryo physiology. PLoS ONE 2011, 6, e23183. [Google Scholar] [CrossRef]
- Hou, Y.J.; Zhu, C.C.; Duan, X.; Liu, H.L.; Wang, Q.; Sun, S.C. Both diet and gene mutation induced obesity affect oocyte quality in mice. Sci. Rep. 2016, 6, 18858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, K.A.; Boudoures, A.L.; Chi, M.M.Y.; Wang, Q.; Moley, K.H. Adverse effects of obesity and/or high-fat diet on oocyte quality and metabolism are not reversible with resumption of regular diet in mice. Reprod. Fertil. Dev. 2015, 27, 716–724. [Google Scholar] [CrossRef] [Green Version]
- Iljas, J.D.; Homer, H.A. Sirt3 is dispensable for oocyte quality and female fertility in lean and obese mice. FASEB J. 2020, 34, 6641–6653. [Google Scholar] [CrossRef] [Green Version]
- Wen, X.; Han, Z.; Liu, S.J.; Hao, X.; Zhang, X.J.; Wang, X.Y.; Zhou, C.J.; Ma, Y.Z.; Liang, C.G. Phycocyanin Improves Reproductive Ability in Obese Female Mice by Restoring Ovary and Oocyte Quality. Front. Cell Dev. Biol. 2020, 8, 595373. [Google Scholar] [CrossRef]
- Rhee, J.S.; Saben, J.L.; Mayer, A.L.; Schulte, M.B.; Asghar, Z.; Stephens, C.; Chi, M.M.Y.; Moley, K.H. Diet-induced obesity impairs endometrial stromal cell decidualization: A potential role for impaired autophagy. Hum. Reprod. 2016, 31, 1315–1326. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Lu, T.; Gao, L.; Fu, X.; Zhu, S.; Hou, Y. Enriched endoplasmic reticulum-mitochondria interactions result in mitochondrial dysfunction and apoptosis in oocytes from obese mice. J. Anim. Sci. Biotechnol. 2017, 8, 62. [Google Scholar] [CrossRef] [PubMed]
- Marei, W.F.A.; Alvarez, M.A.; Van Hoeck, V.; Gutierrez-Adan, A.; Bols, P.E.J.; Leroy, J.L.M.R. Effect of nutritionally induced hyperlipidaemia on in vitro bovine embryo quality depends on the type of major fatty acid in the diet. Reprod. Fertil. Dev. 2017, 29, 1856–1867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boots, C.E.; Boudoures, A.; Zhang, W.; Drury, A.; Moley, K.H. Obesity-induced oocyte mitochondrial defects are partially prevented and rescued by supplementation with co-enzyme Q10 in a mouse model. Hum. Reprod. 2016, 31, 2090–2097. [Google Scholar] [CrossRef]
- Saben, J.L.; Boudoures, A.L.; Asghar, Z.; Thompson, A.; Drury, A.; Zhang, W.; Chi, M.; Cusumano, A.; Scheaffer, S.; Moley, K.H. Maternal Metabolic Syndrome Programs Mitochondrial Dysfunction via Germline Changes across Three Generations. Cell Rep. 2016, 16, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Han, L.; Ma, R.; Hou, X.; Yu, Y.; Sun, S.; Xu, Y.; Schedl, T.; Moley, K.H.; Wang, Q. Sirt3 prevents maternal obesity-associated oxidative stress and meiotic defects in mouse oocytes. Cell Cycle 2015, 14, 2959–2968. [Google Scholar] [CrossRef]
- Hostens, M.; Fievez, V.; Leroy, J.L.M.R.; van de Burgwal, E.J.; Van Ranst, B.; Vlaeminck, B.; Opsomer, G. Milk fat saturation and reproductive performance in dairy cattle. Anim. Reprod. Sci. 2013, 141, 116–123. [Google Scholar] [CrossRef]
- Han, L.; Wang, H.; Li, L.; Li, X.; Ge, J.; Reiter, R.J.; Wang, Q. Melatonin protects against maternal obesity-associated oxidative stress and meiotic defects in oocytes via the SIRT3-SOD2-dependent pathway. J. Pineal Res. 2017, 63, e12431. [Google Scholar] [CrossRef]
- Khan, R.; Jiang, X.; Hameed, U.; Shi, Q. Role of Lipid Metabolism and Signaling in Mammalian Oocyte Maturation, Quality, and Acquisition of Competence. Front. Cell Dev. Biol. 2021, 9, 384. [Google Scholar] [CrossRef]
- Snider, A.P.; Wood, J.R. Obesity induces ovarian inflammation and reduces oocyte quality. Reproduction 2019, 158, R79–R90. [Google Scholar] [CrossRef] [Green Version]
- Schultz, R.M.; Kopf, G.S. Molecular basis of mammalian egg activation. Curr. Top. Dev. Biol. 1995, 30, 21–62. [Google Scholar] [CrossRef]
- Stricker, S.A. Comparative biology of calcium signaling during fertilization and egg activation in animals. Dev. Biol. 1999, 211, 157–176. [Google Scholar] [CrossRef] [PubMed]
- Ducibella, T.; Huneau, D.; Angelichio, E.; Xu, Z.; Schultz, R.M.; Kopf, G.S.; Fissore, R.; Madoux, S.; Ozil, J.P. Egg-to-embryo transition is driven by differential responses to Ca2+ oscillation number. Dev. Biol. 2002, 250, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, S.I.; Yuzaki, M.; Nakada, K.; Shirakawa, H.; Nakanishi, S.; Nakade, S.; Mikoshiba, K. Block of Ca2+ wave and Ca2+ oscillation by antibody to the inositol 1,4,5-trisphosphate receptor in fertilized hamster eggs. Science 1992, 257, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Yang, L.; Li, P.; Hofmann, O.; Dicker, L.; Hide, W.; Lin, X.; Watkins, S.M.; Ivanov, A.R.; Hotamisligil, G.S. Aberrant lipid metabolism disrupts calcium homeostasis causing liver endoplasmic reticulum stress in obesity. Nature 2011, 473, 528–531. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, A.; Chaube, S.K. High cytosolic free calcium level signals apoptosis through mitochondria-caspase mediated pathway in rat eggs cultured in vitro. Apoptosis 2012, 17, 439–448. [Google Scholar] [CrossRef]
- Luzzo, K.M.; Wang, Q.; Purcell, S.H.; Chi, M.; Jimenez, P.T.; Grindler, N.; Schedl, T.; Moley, K.H. High fat diet induced developmental defects in the mouse: Oocyte meiotic aneuploidy and fetal growth retardation/brain defects. PLoS ONE 2012, 7, e49217. [Google Scholar] [CrossRef] [Green Version]
- Aitken, R.J.; Findlay, J.K.; Hutt, K.J.; Kerr, J.B. Apoptosis in the germ line. Reproduction 2011, 141, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Eisenbach, M. Sperm chemotaxis. Rev. Reprod. 1999, 4, 56–66. [Google Scholar] [CrossRef]
- Bedford, J.M.; Kim, H.H. Cumulus oophorus as a sperm sequestering device, in vivo. J. Exp. Zool. 1993, 265, 321–328. [Google Scholar] [CrossRef]
- Carrell, D.T.; Middleton, R.G.; Peterson, C.M.; Jones, K.P.; Urry, R.L. Role of the cumulus in the selection of morphologically normal sperm and induction of the acrosome reaction during human in vitro fertilization. Arch. Androl. 1993, 31, 133–137. [Google Scholar] [CrossRef] [Green Version]
- Tanghe, S.; Van Soom, A.; Nauwynck, H.; Coryn, M.; De Kruif, A. Minireview: Functions of the cumulus oophorus during oocyte maturation, ovulation, and fertilization. Mol. Reprod. Dev. 2002, 61, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.J.; Wu, S.N.; Shen, J.P.; Wang, D.H.; Kong, X.W.; Lu, A.; Li, Y.J.; Zhou, H.X.; Zhao, Y.F.; Liang, C.G. The beneficial effects of cumulus cells and oocyte-cumulus cell gap junctions depends on oocyte maturation and fertilization methods in mice. PeerJ 2016, 4, e1761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jungheim, E.S.; MacOnes, G.A.; Odem, R.R.; Patterson, B.W.; Lanzendorf, S.E.; Ratts, V.S.; Moley, K.H. Associations between free fatty acids, cumulus oocyte complex morphology and ovarian function during in vitro fertilization. Fertil. Steril. 2011, 95, 1970–1974. [Google Scholar] [CrossRef]
- Mullen, T.J.; Davis-Roca, A.C.; Wignall, S.M. Spindle assembly and chromosome dynamics during oocyte meiosis. Curr. Opin. Cell Biol. 2019, 60, 53–59. [Google Scholar] [CrossRef]
- Lane, S.; Kauppi, L. Meiotic spindle assembly checkpoint and aneuploidy in males versus females. Cell. Mol. Life Sci. 2019, 76, 1135–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, H.; Hamatani, T.; Kamijo, S.; Iwai, M.; Kobanawa, M.; Ogawa, S.; Miyado, K.; Tanaka, M. Impact of Oxidative Stress on Age-Associated Decline in Oocyte Developmental Competence. Front. Endocrinol. 2019, 10, 811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, D.; Yasui, T.; Kobayashi, C.; Kitazato, T.; Iwasa, T.; Irahara, M. Effect of high fat diet on artificial oocyte activation following superovulation in mice. Zygote 2016, 24, 286–292. [Google Scholar] [CrossRef]
- Scheffler, K.; Uraji, J.; Jentoft, I.; Cavazza, T.; Mönnich, E.; Mogessie, B.; Schuh, M. Two mechanisms drive pronuclear migration in mouse zygotes. Nat. Commun. 2021, 12, 841. [Google Scholar] [CrossRef]
- The Developmental Mechanics of Cell Specification—Developmental Biology—NCBI Bookshelf. Available online: https://www.ncbi.nlm.nih.gov/books/NBK9968/ (accessed on 17 September 2022).
- Cakmak, H.; Taylor, H.S. Implantation failure: Molecular mechanisms and clinical treatment. Hum. Reprod. Update 2011, 17, 242–253. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Dey, S.K. Roadmap to embryo implantation: Clues from mouse models. Nat. Rev. Genet. 2006, 7, 185–199. [Google Scholar] [CrossRef]
- Yousif, M.D.; Calder, M.D.; Du, J.T.; Ruetz, K.N.; Crocker, K.; Urquhart, B.L.; Betts, D.H.; Rafea, B.A.; Watson, A.J. Oleic Acid Counters Impaired Blastocyst Development Induced by Palmitic Acid During Mouse Preimplantation Development: Understanding Obesity-Related Declines in Fertility. Reprod. Sci. 2020, 27, 2038–2051. [Google Scholar] [CrossRef] [PubMed]
- Kubandová, J.; Čikoš, Š.; Burkuš, J.; Czikková, S.; Koppel, J.; Fabian, D. Amount of maternal body fat significantly affected the quality of isolated mouse preimplantation embryos and slowed down their development. Theriogenology 2014, 81, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Reis, A.; Rooke, J.A.; McCallum, G.J.; Staines, M.E.; Ewen, M.; Lomax, M.A.; McEvoy, T.G. Consequences of exposure to serum, with or without vitamin E supplementation, in terms of the fatty acid content and viability of bovine blastocysts produced in vitro. Reprod. Fertil. Dev. 2003, 15, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Sudano, M.J.; Rascado, T.D.S.; Tata, A.; Belaz, K.R.A.; Santos, V.G.; Valente, R.S.; Mesquita, F.S.; Ferreira, C.R.; Araújo, J.P.; Eberlin, M.N.; et al. Lipidome signatures in early bovine embryo development. Theriogenology 2016, 86, 472–484.e1. [Google Scholar] [CrossRef] [Green Version]
- Lessey, B.A.; Young, S.L. What exactly is endometrial receptivity? Fertil. Steril. 2019, 111, 611–617. [Google Scholar] [CrossRef]
- Carson, D.D.; Lagow, E.; Thathiah, A.; Al-Shami, R.; Farach-Carson, M.C.; Vernon, M.; Yuan, L.; Fritz, M.A.; Lessey, B. Changes in gene expression during the early to mid-luteal (receptive phase) transition in human endometrium detected by high-density microarray screening. Mol. Hum. Reprod. 2002, 8, 871–879. [Google Scholar] [CrossRef] [Green Version]
- Altmäe, S.; Reimand, J.R.; Hovatta, O.; Zhang, P.; Kere, J.; Laisk, T.; Saare, M.; Peters, M.; Vilo, J.; Stavreus-Evers, A.; et al. Research resource: Interactome of human embryo implantation: Identification of gene expression pathways, regulation, and integrated regulatory networks. Mol. Endocrinol. 2012, 26, 203–217. [Google Scholar] [CrossRef]
- Ng, S.W.; Norwitz, G.A.; Pavlicev, M.; Tilburgs, T.; Simón, C.; Norwitz, E.R. Endometrial Decidualization: The Primary Driver of Pregnancy Health. Int. J. Mol. Sci. 2020, 21, 4092. [Google Scholar] [CrossRef]
- Norwitz, E.R.; Schust, D.J.; Fisher, S.J. Implantation and the survival of early pregnancy. N. Engl. J. Med. 2001, 345, 1400–1408. [Google Scholar] [CrossRef]
- Adastra, K.L.; Frolova, A.I.; Chi, M.M.; Cusumano, D.; Bade, M.; Carayannopoulos, M.O.; Moley, K.H. Slc2a8 deficiency in mice results in reproductive and growth impairments. Biol. Reprod. 2012, 87, 49. [Google Scholar] [CrossRef]
- Yang, S.; Wang, H.; Li, D.; Li, M. Role of Endometrial Autophagy in Physiological and Pathophysiological Processes. J. Cancer 2019, 10, 3459–3471. [Google Scholar] [CrossRef] [PubMed]
- Salamun, V.; Bokal, E.V.; Maver, A.; Papler, T.B. Transcriptome study of receptive endometrium in overweight and obese women shows important expression differences in immune response and inflammatory pathways in women who do not conceive. PLoS ONE 2021, 16, e0261873. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.J.; Uyehara, C.F.T.; Hashiro, G.M.; Frattarelli, J.L. The utility of serum leptin and follicular fluid leptin, estradiol, and progesterone levels during an in vitro fertilization cycle. J. Assist. Reprod. Genet. 2007, 24, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Metwally, M.; Preece, R.; Thomas, J.; Ledger, W.; Li, T.C. A proteomic analysis of the endometrium in obese and overweight women with recurrent miscarriage: Preliminary evidence for an endometrial defect. Reprod. Biol. Endocrinol. 2014, 12, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellver, J.; Marín, C.; Lathi, R.B.; Murugappan, G.; Labarta, E.; Vidal, C.; Giles, J.; Cabanillas, S.; Marzal, A.; Galliano, D.; et al. Obesity Affects Endometrial Receptivity by Displacing the Window of Implantation. Reprod. Sci. 2021, 28, 3171–3180. [Google Scholar] [CrossRef] [PubMed]
- Erel, C.T.; Senturk, L.M. The impact of body mass index on assisted reproduction. Curr. Opin. Obstet. Gynecol. 2009, 21, 228–235. [Google Scholar] [CrossRef]
- Tanaka, T.; Umesaki, N. Leptin regulates the proliferation and apoptosis of human endometrial epithelial cells. Int. J. Mol. Med. 2008, 22, 683–689. [Google Scholar] [CrossRef] [Green Version]
- Niu, R.; Okamoto, T.; Iwase, K.; Nomura, S.; Mizutani, S. Quantitative analysis of matrix metalloproteinases-2 and -9, and their tissue inhibitors-1 and -2 in human placenta throughout gestation. Life Sci. 2000, 66, 1127–1137. [Google Scholar] [CrossRef]
- Maffeis, C.; Morandi, A. Effect of Maternal Obesity on Foetal Growth and Metabolic Health of the Offspring. Obes. Facts 2017, 10, 112–117. [Google Scholar] [CrossRef]
- Moley, K.H.; Vaughn, W.K.; DeCherney, A.H.; Diamond, M.P. Effect of diabetes mellitus on mouse pre-implantation embryo development. J. Reprod. Fertil. 1991, 93, 325–332. [Google Scholar] [CrossRef]
- Wyman, A.; Pinto, A.B.; Sheridan, R.; Moley, K.H. One-cell zygote transfer from diabetic to nondiabetic mouse results in congenital malformations and growth retardation in offspring. Endocrinology 2008, 149, 466–469. [Google Scholar] [CrossRef] [PubMed]
- Minge, C.E.; Bennett, B.D.; Norman, R.J.; Robker, R.L. Peroxisome proliferator-activated receptor-gamma agonist rosiglitazone reverses the adverse effects of diet-induced obesity on oocyte quality. Endocrinology 2008, 149, 2646–2656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jungheim, E.S.; Schoeller, E.L.; Marquard, K.L.; Louden, E.D.; Schaffer, J.E.; Moley, K.H. Diet-induced obesity model: Abnormal oocytes and persistent growth abnormalities in the offspring. Endocrinology 2010, 151, 4039–4046. [Google Scholar] [CrossRef] [PubMed]
- Bazzano, M.V.; Sarrible, G.B.; de Astrada, M.B.; Elia, E. Obesity modifies the implantation window and disrupts intrauterine embryo positioning in rats. Reproduction 2021, 162, 61–72. [Google Scholar] [CrossRef]
- Kazemi, A.; Ramezanzadeh, F.; Nasr-Esfahani, M.H. Relationship between Dietary Fat Intake, Its Major Food Sources and Assisted Reproduction Parameters. J. Reprod. Infertil. 2014, 15, 214–221. [Google Scholar]
- Valckx, S.D.M.; De Bie, J.; Michiels, E.D.; Goovaerts, I.G.; Punjabi, U.; Ramos-Ibeas, P.; Gutierrez-Adan, A.; Bols, P.E.; Leroy, J.L. The effect of human follicular fluid on bovine oocyte developmental competence and embryo quality. Reprod. Biomed. Online 2015, 30, 203–207. [Google Scholar] [CrossRef] [Green Version]
- Matorras, R.; Exposito, A.; Ferrando, M.; Mendoza, R.; Larreategui, Z.; Laínz, L.; Aranburu, L.; Andrade, F.; Aldámiz-Echevarria, L.; Ruiz-Larrea, M.B.; et al. Oocytes of women who are obese or overweight have lower levels of n-3 polyunsaturated fatty acids compared with oocytes of women with normal weight. Fertil. Steril. 2020, 113, 53–61. [Google Scholar] [CrossRef]
- Leary, C.; Leese, H.J.; Sturmey, R.G. Human embryos from overweight and obese women display phenotypic and metabolic abnormalities. Hum. Reprod. 2015, 30, 122–132. [Google Scholar] [CrossRef] [Green Version]
- Baddela, V.S.; Sharma, A.; Vanselow, J. Non-esterified fatty acids in the ovary: Friends or foes? Reprod. Biol. Endocrinol. 2020, 18, 60. [Google Scholar] [CrossRef]
- Li, L.; Wu, C.S.; Hou, G.M.; Dong, M.Z.; Wang, Z.B.; Hou, Y.; Schatten, H.; Zhang, G.R.; Sun, Q.Y. Type 2 diabetes increases oocyte mtDNA mutations which are eliminated in the offspring by bottleneck effect. Reprod. Biol. Endocrinol. 2018, 16, 110. [Google Scholar] [CrossRef]
- Wang, H.; Cheng, Q.; Li, X.; Hu, F.; Han, L.; Zhang, H.; Li, L.; Ge, J.; Ying, X.; Guo, X.; et al. Loss of TIGAR Induces Oxidative Stress and Meiotic Defects in Oocytes from Obese Mice. Mol. Cell. Proteomics 2018, 17, 1354–1364. [Google Scholar] [CrossRef] [Green Version]
- Lopomo, A.; Burgio, E.; Migliore, L. Epigenetics of Obesity. Prog. Mol. Biol. Transl. Sci. 2016, 140, 151–184. [Google Scholar] [CrossRef] [PubMed]
- Gu, T.P.; Guo, F.; Yang, H.; Wu, H.P.; Xu, G.F.; Liu, W.; Xie, Z.G.; Shi, L.; He, X.; Jin, S.G.; et al. The role of Tet3 DNA dioxygenase in epigenetic reprogramming by oocytes. Nature 2011, 477, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Belchetz, P.E.; Plant, T.M.; Nakai, Y.; Keogh, E.J.; Knobil, E. Hypophysial responses to continuous and intermittent delivery of hypopthalamic gonadotropin-releasing hormone. Science 1978, 202, 631–633. [Google Scholar] [CrossRef] [PubMed]
- Francés-Herrero, E.; Lopez, R.; Hellström, M.; de Miguel-Gómez, L.; Herraiz, S.; Brännström, M.; Pellicer, A.; Cervelló, I. Bioengineering trends in female reproduction: A systematic review. Hum. Reprod. Update 2022, 28, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Eddie, S.L.; Quartuccio, S.M.; Zhu, J.; Shepherd, J.A.; Kothari, R.; Kim, J.J.; Woodruff, T.K.; Burdette, J.E. Three-dimensional modeling of the human fallopian tube fimbriae. Gynecol. Oncol. 2015, 136, 348–354. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Park, E.S.; Xiang, D.; Li, Z. Long-term organoid culture reveals enrichment of organoid-forming epithelial cells in the fimbrial portion of mouse fallopian tube. Stem Cell Res. 2018, 32, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Xu, Y.; Rashedi, A.S.; Pavone, M.E.; Julie Kim, J.; Woodruff, T.K.; Burdette, J.E. Human fallopian tube epithelium co-culture with murine ovarian follicles reveals crosstalk in the reproductive cycle. Mol. Hum. Reprod. 2016, 22, 756–767. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; West, E.; Shea, L.D.; Woodruff, T.K. Identification of a Stage-Specific Permissive In Vitro Culture Environment for Follicle Growth and Oocyte Development1. Biol. Reprod. 2006, 75, 916–923. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rodent Models of Obesity (%) | Diet Macro-Nutrients (~ %) | Morphological Effects on Female Fertility [References] | ||||
|---|---|---|---|---|---|---|
| Endocrine | Ovarian Folliculogenesis | Oocyte Competencies | ||||
| Granulosa Cells | Theca Cells | |||||
| 60% of mammal models | Mouse (51%) | ~44% Fat ~52% Carbohydrate ~20% Protein | 2. Estrous cycle impairments [33,34,36,75,95,99,100,115,116,117,132,179,207] 3. Ovary reserve depletion [36,47,81,82,100,129] 4. Altered follicle gene profiles [33,35,36,50,75,77,100,115] 5. Gonadotropin release defects [14,33,34,36,37,75,95,99,100,115,132,179,207] 6. Steroid release impairment [33,36,50,99,100] 7. Perinatal growth delay [50,89,100,196,241] 8. Pubertal onset defects [21,50] | 1. Impaired cell control [34,41,47,54,56,73,74,95] 2. Estradiol synthesis [34,36,38,41,44,46,47,48,50,56,70,74,75,77,80,83,94,95,96,99,100,179] 3. Development failures [75,76,77,151,155,156,233] 4. Lipid droplets [80,154] | 1. Androgen production dysregulation [34,36,41,44,46,47,48,50,70,74,75,80,94,95,96,99,100,122,179] 2. LH receptors [44,97,221] | 1. Oocyte apoptosis [48,99,151,153,154,155,176,179,180,181] 2. Partial meiotic competence [33,40,46,48,99,100,114,132,176,177,178] 3. MII oocyte [33,35,37,40,43,46,48,55,76,96,99,100,114,115,117,132,149,153,155,157,176,177,178,180,183,185,207,213,233,234] 4. Fertilization rate [33,37,40,43,46,114,121,153,155,183] 5. Oocyte lipid droplets [43,47,55,115,132,145,154,212] 6. Mitochondrial de-regulation [33,43,47,55,80,100,115,133,145,149,151,176,177,178,179,181,183,184,185,212,241,242] 7. Spindle defects [46,48,55,100,117,132,176,177,178,179,183,185,187,196,242] 8. Chromosome’s misalignment [46,100,117,177,178,185,187,196,242] 9. Epigenetic modifications [39,40,44,47,96,117,176,179,184] |
| Rats (9%) | ~40% Fat ~66% Carbohydrate ~20% Protein | 1. Pubertal onset defects [19,25] 2. Estrous cycle impairments [19,25,26,32,42,71,72,98] 3. Ovary reserve depletion [19,42,71] 4. Altered follicle gene profiles [19,25,26,32,42,72,84,98] 5. Gonadotropin release defects [9,19,25,42,84,98] 6. Steroid release impairment [19,25,26,84,98] 7. Perinatal growth delay [27] | 1. Impaired cell control [26,32,42,71,72,235] 2. Estradiol synthesis [2,19,24,25,26,32,42,71,84,98] | 1. Androgen synthesis dysregulation [19,25,26,32,42,71,72,84,98] 2. LH receptors [25] | 1. Oocyte apoptosis [42,195] 6. Mitochondrial de-regulation [42] | |
| Non-Rodent Models of Obesity (% of Mammal Models) | Diet Macro-Nutrients (~ %) | Morphological Effects on Female Fertility [References] | ||||
|---|---|---|---|---|---|---|
| Endocrine | Ovarian Folliculogenesis | Oocyte Competencies | ||||
| POLI GASTRIC (15%) | Ovi- caprine (3%) | n.s* Fat n.s* Carbohydrate ~17% Protein | 2. Estrous cycle impairments [78,118] 9. Ovulation timing changes [78,118,148] | 3. Development failures [78] | 3. Theca dysfunction [78] | 4. Fertilization rate [118,148] |
| Bovine (12%) | 60% Fat; ~57% Carbohydrate ~88% Protein | 1. Pubertal onset defects [20] 2. Estrous cycle impairments [112,182,186] 3. Ovarian reserve depletion [182,186] | 3. Development failures [20,150,159,160,161,162,163,164,165,166,167,168] | 3. Theca dysfunction [20] 4. Late folliculogenesis defects mediated by angiogenesis [112] | 1. Oocyte apoptosis [182] 3. MII oocytes [20,150,159,160,161,162,163,164,165,166,167,168,175,182] 4. Fertilization rate [150,182,186] 5. Oocyte lipid droplets [237] | |
| MONO GASTRIC (5%) | Pig (4%) | ~62% Fat; ~20% Carbohydrate | 5. Gonadotropin release defects [28,29] 6. Steroid release defects [28,29] | 2. Estradiol synthesis [28,29,45,85] 3. Development failures [85,169,170,171] | 3. Theca dysfunction [28,29,45] | 2. Partial meiotic competence [45,85] 3. MII oocytes [45,85] |
| Rabbit (1%) | ~8% Fat n.s* Carbohydrate n.s* Protein | 3. Ovarian reserve depletion [18] | 3. Development failures [18] | 3. Theca dysfunction [18] | - | |
| PRIMATES (20%) | Human (17%) | n.s* | 3. Ovarian reserve depletion [128] 5. Gonadotropin release defects [79] 6. Steroid release defects [79] | 3. Development failures [5,79,92,93,158] 4. Lipid droplets [79] 5. Altered angiogenesis [102,103,104,105,106,107,108,109,110] 6. FF oxidative biomarkers [147,148,152,153,203] | 3. Theca dysfunction [79,119] | 3. MII oocytes [4,79,152,236,237,238,239] 4. Fertilization rate [4,79,225,237,238] |
| Non-human (3%) | ~35% Fat ~49% Carbohydrate ~18% Protein | 2. Estrous cycle impairment [30,57,86,87,245] 4. Altered follicle gene profiles [30,57,86,87] 5. Gonadotropin release defects [30,57,86,87] 6. Steroid release defects [30,57,86,87] 9. Ovulation timing changes [30,57,86,87] | 2. Estradiol synthesis [30,57,86,87] | 3. Theca dysfunction [57,86,87] | 3. MII oocytes [57,86,87] 4. Fertilization rate [57,86,87] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Berardino, C.; Peserico, A.; Capacchietti, G.; Zappacosta, A.; Bernabò, N.; Russo, V.; Mauro, A.; El Khatib, M.; Gonnella, F.; Konstantinidou, F.; et al. High-Fat Diet and Female Fertility across Lifespan: A Comparative Lesson from Mammal Models. Nutrients 2022, 14, 4341. https://doi.org/10.3390/nu14204341
Di Berardino C, Peserico A, Capacchietti G, Zappacosta A, Bernabò N, Russo V, Mauro A, El Khatib M, Gonnella F, Konstantinidou F, et al. High-Fat Diet and Female Fertility across Lifespan: A Comparative Lesson from Mammal Models. Nutrients. 2022; 14(20):4341. https://doi.org/10.3390/nu14204341
Chicago/Turabian StyleDi Berardino, Chiara, Alessia Peserico, Giulia Capacchietti, Alex Zappacosta, Nicola Bernabò, Valentina Russo, Annunziata Mauro, Mohammad El Khatib, Francesca Gonnella, Fani Konstantinidou, and et al. 2022. "High-Fat Diet and Female Fertility across Lifespan: A Comparative Lesson from Mammal Models" Nutrients 14, no. 20: 4341. https://doi.org/10.3390/nu14204341
APA StyleDi Berardino, C., Peserico, A., Capacchietti, G., Zappacosta, A., Bernabò, N., Russo, V., Mauro, A., El Khatib, M., Gonnella, F., Konstantinidou, F., Stuppia, L., Gatta, V., & Barboni, B. (2022). High-Fat Diet and Female Fertility across Lifespan: A Comparative Lesson from Mammal Models. Nutrients, 14(20), 4341. https://doi.org/10.3390/nu14204341