
Effects of Whey Peptide Supplementation on Sarcopenic Obesity in High-Fat Diet-Fed Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Whey Peptide
2.2. Animals and Sarcopenic Obesity Induction
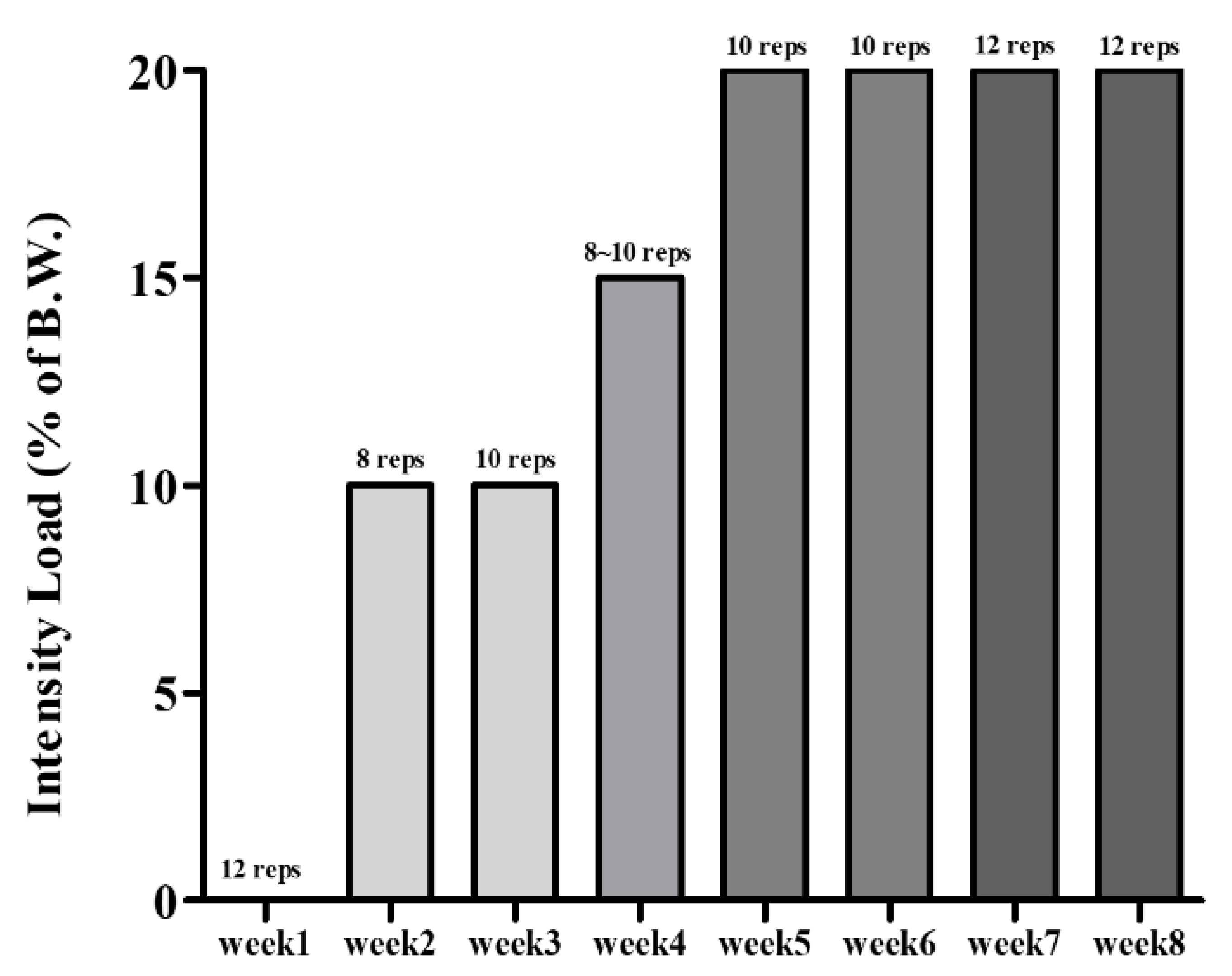
2.3. Experimental Design
2.4. Body Composition Analysis
2.5. Muscle Strength Test
2.6. Histological Assays
2.7. Protein Extraction and Western Blot Analysis
2.8. RNA Extraction and Real-Time PCR Analysis
2.9. Statistical Analysis
3. Results
3.1. Effects of WP Supplementation and RE on Diet Intake, Body, Fat, and Muscle Weights, Body Composition, and Muscle Strength
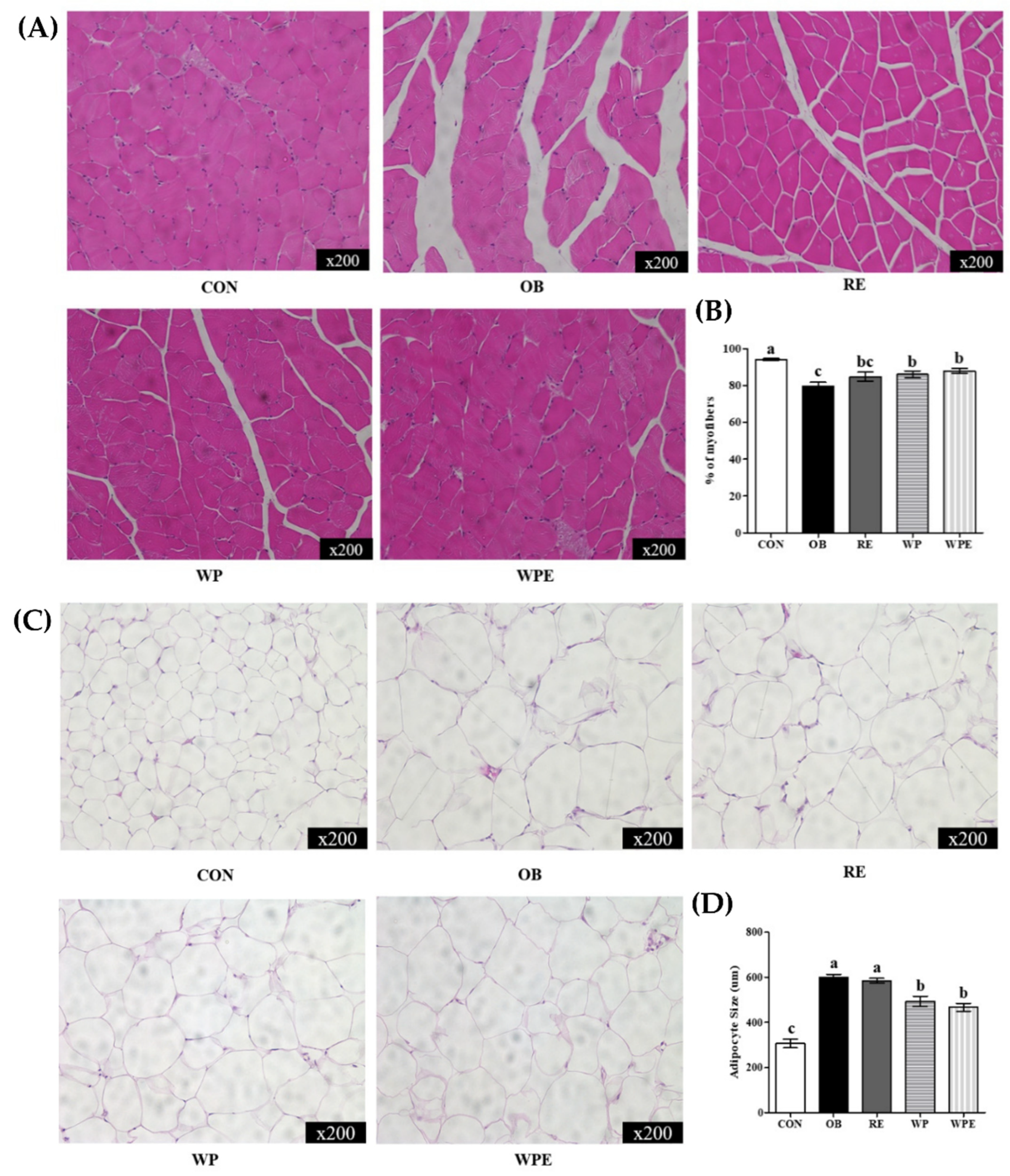
3.2. Effects of WP Supplementation and RE on Morphological Changes in Adipose Tissue and Skeletal Muscle
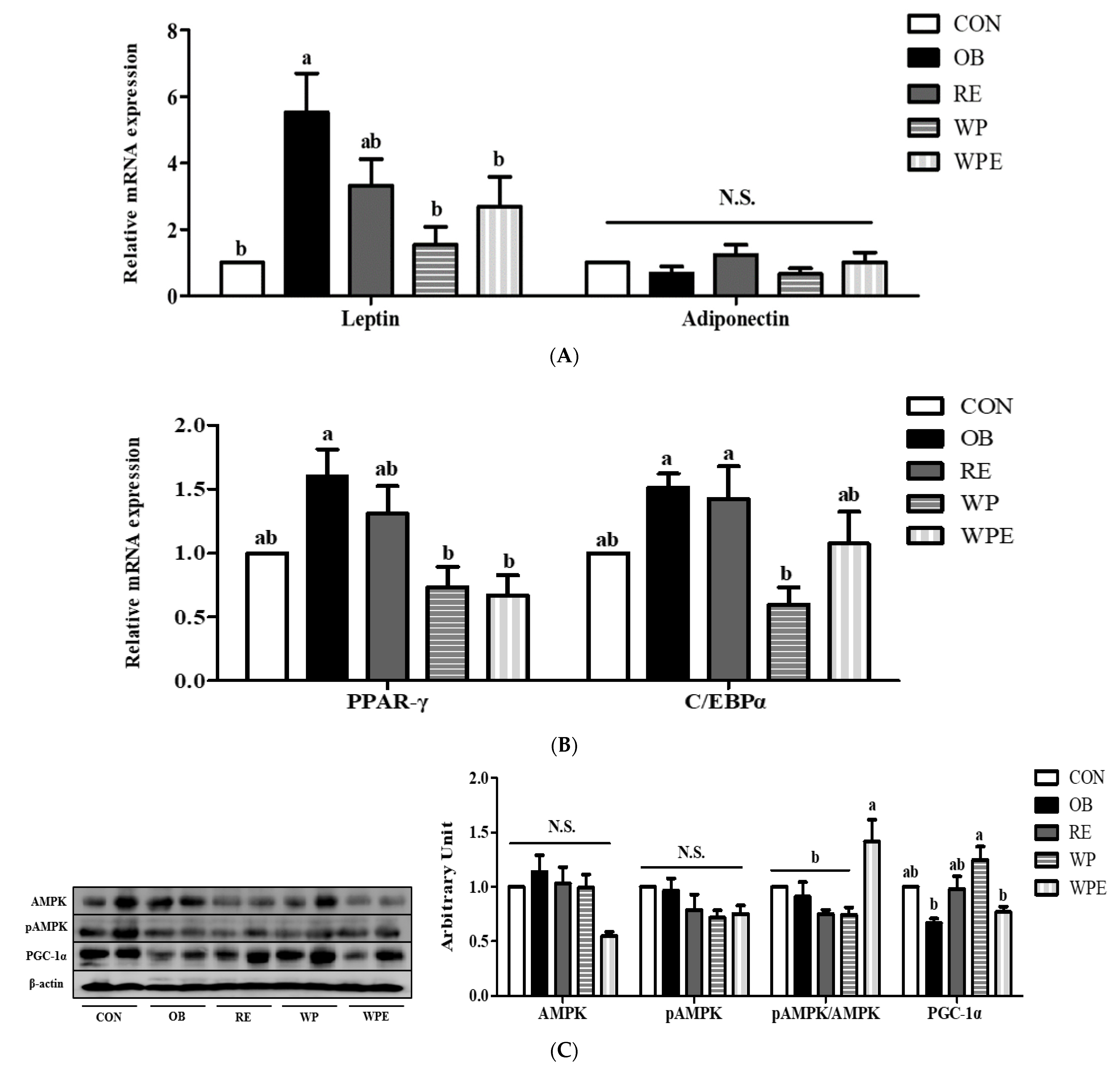
3.3. Effects of WP Supplementation and RE on Adipokines and Lipid and Energy Metabolism in Adipose Tissue
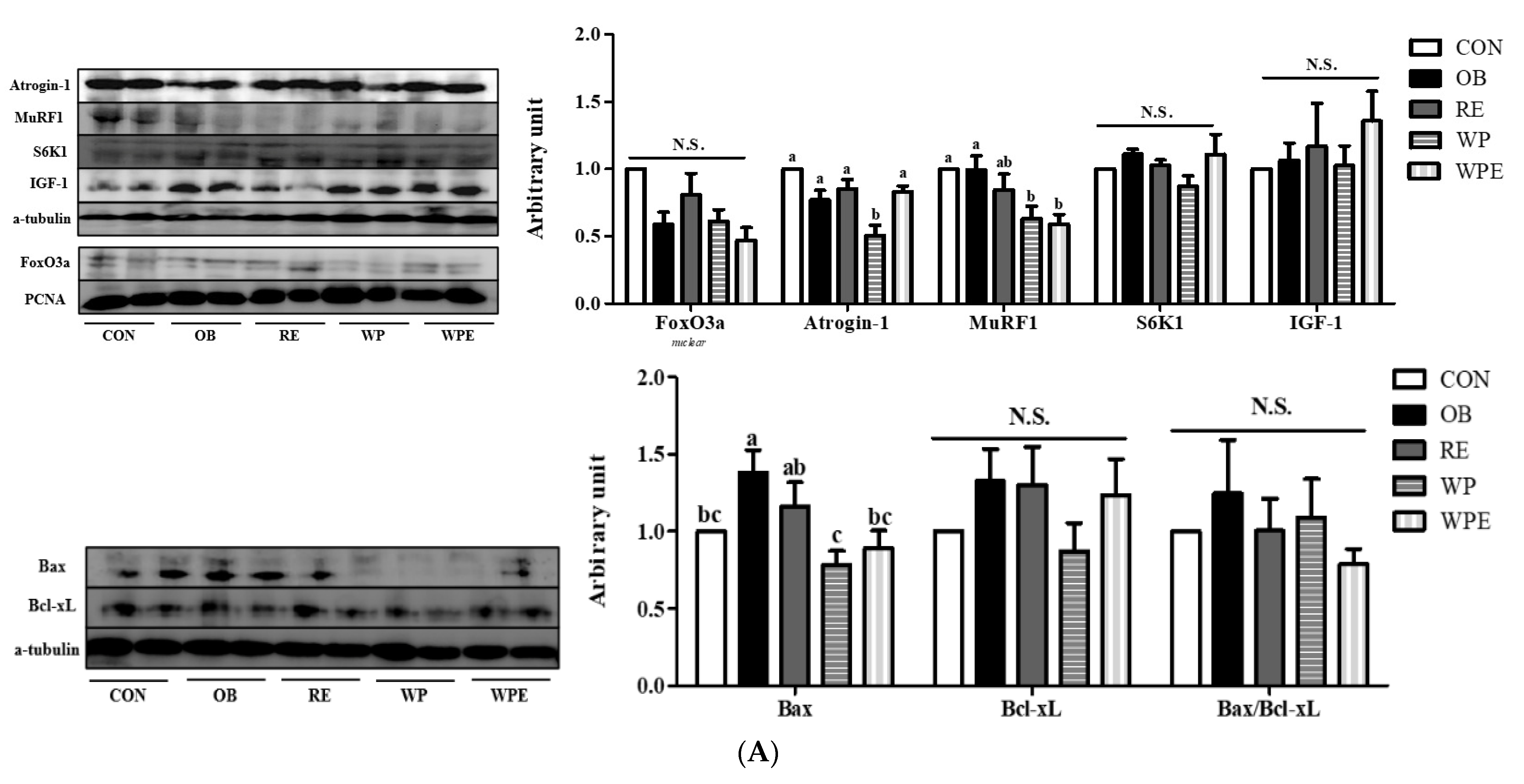
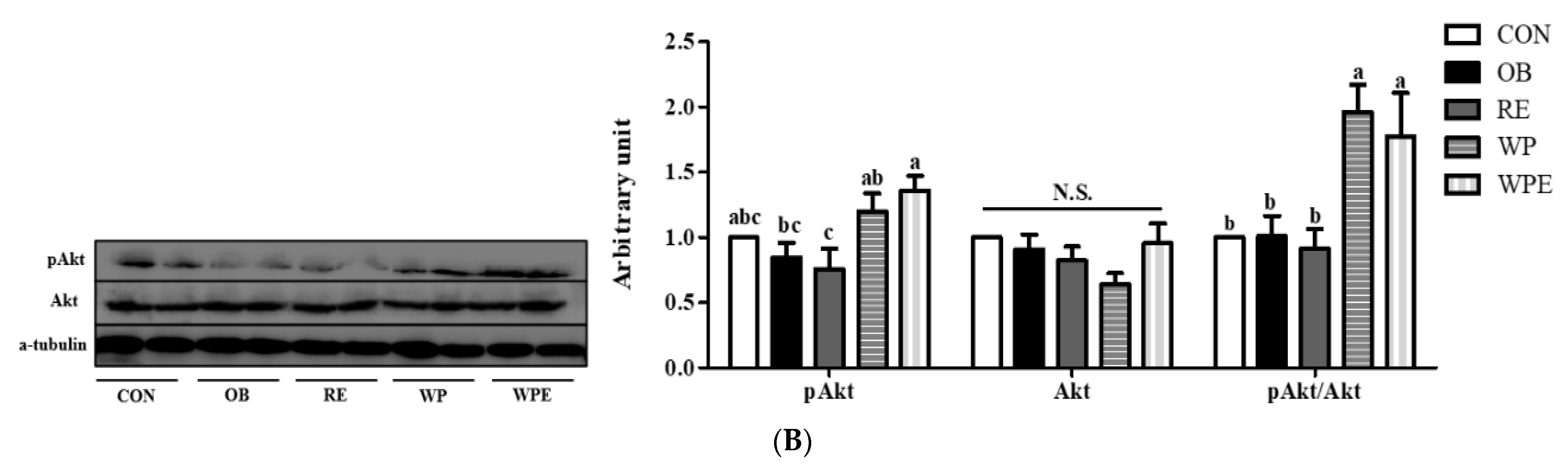
3.4. Effects of WP Supplementation and RE on Protein and Energy Metabolism and Apoptosis in Skeletal Muscle
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, X.; Xie, X.; Dou, Q.; Liu, C.; Zhang, W.; Yang, Y.; Deng, R.; Cheng, A.S.K. Association of Sarcopenic Obesity with the Risk of All-Cause Mortality among Adults over a Broad Range of Different Settings: A Updated Meta-Analysis. BMC Geriatr. 2019, 19, 183. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Xiao, G.; Fan, Y.; He, M.; Lv, S.; Li, Y. Sarcopenic Obesity: Research Advances in Pathogenesis and Diagnostic Criteria. Aging Clin. Exp. Res. 2021, 33, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Batsis, J.A.; Villareal, D.T. Sarcopenic Obesity in Older Adults: Aetiology, Epidemiology and Treatment Strategies. Nat. Rev. Endocrinol. 2018, 14, 513–537. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.L.; Sinha, S. Obesity and Aging: Molecular Mechanisms and Therapeutic Approaches. Ageing Res. Rev. 2021, 67, 101268. [Google Scholar] [CrossRef] [PubMed]
- CDC. Obesity is a Common, Serious, and Costly Disease; CDC: Atlanta, GA, USA, 2020.
- Gao, W.; Liu, J.-L.; Lu, X.; Yang, Q. Epigenetic Regulation of Energy Metabolism in Obesity. J. Mol. Cell Biol. 2021, 13, 480–499. [Google Scholar] [CrossRef]
- Roh, E.; Choi, K.M. Health Consequences of Sarcopenic Obesity: A Narrative Review. Front. Endocrinol. 2020, 11, 332. [Google Scholar] [CrossRef]
- Jin, H.; Oh, H.-J.; Nah, S.-Y.; Lee, B.-Y. Gintonin-Enriched Fraction Protects against Sarcopenic Obesity by Promoting Energy Expenditure and Attenuating Skeletal Muscle Atrophy in High-Fat Diet-Fed Mice. J. Ginseng. Res. 2022, 46, 454–463. [Google Scholar] [CrossRef]
- Wang, Z.-X.; Wang, M.-Y.; Yang, R.-X.; Zhao, Z.-H.; Xin, F.-Z.; Li, Y.; Ren, T.-Y.; Fan, J.-G. Ammonia Scavenger Restores Liver and Muscle Injury in a Mouse Model of Non-Alcoholic Steatohepatitis with Sarcopenic Obesity. Front. Nutr. 2022, 9, 808497. [Google Scholar] [CrossRef]
- Brown, L.A.; Perry, R.A.; Haynie, W.S.; Lee, D.E.; Rosa-Caldwell, M.E.; Brown, J.L.; Greene, N.P.; Wolchok, J.C.; Washington, T.A. Moderators of Skeletal Muscle Maintenance Are Compromised in Sarcopenic Obese Mice. Mech. Ageing Dev. 2021, 194, 111404. [Google Scholar] [CrossRef]
- Liao, C.-D.; Tsauo, J.-Y.; Huang, S.-W.; Ku, J.-W.; Hsiao, D.-J.; Liou, T.-H. Effects of Elastic Band Exercise on Lean Mass and Physical Capacity in Older Women with Sarcopenic Obesity: A Randomized Controlled Trial. Sci. Rep. 2018, 8, 2317. [Google Scholar] [CrossRef]
- Chen, H.-T.; Chung, Y.-C.; Chen, Y.-J.; Ho, S.-Y.; Wu, H.-J. Effects of Different Types of Exercise on Body Composition, Muscle Strength, and IGF-1 in the Elderly with Sarcopenic Obesity. J. Am. Geriatr. Soc. 2017, 65, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Dieli-Conwright, C.M.; Courneya, K.S.; Demark-Wahnefried, W.; Sami, N.; Lee, K.; Buchanan, T.A.; Spicer, D.v.; Tripathy, D.; Bernstein, L.; Mortimer, J.E. Effects of Aerobic and Resistance Exercise on Metabolic Syndrome, Sarcopenic Obesity, and Circulating Biomarkers in Overweight or Obese Survivors of Breast Cancer: A Randomized Controlled Trial. J. Clin. Oncol. 2018, 36, 875–883. [Google Scholar] [CrossRef] [PubMed]
- Peterson, M.D.; Rhea, M.R.; Sen, A.; Gordon, P.M. Resistance Exercise for Muscular Strength in Older Adults: A Meta-Analysis. Ageing Res. Rev. 2010, 9, 226–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, M.D.; Sen, A.; Gordon, P.M. Influence of Resistance Exercise on Lean Body Mass in Aging Adults. Med. Sci. Sports Exerc. 2011, 43, 249–258. [Google Scholar] [CrossRef] [Green Version]
- Giné-Garriga, M.; Roqué-Fíguls, M.; Coll-Planas, L.; Sitjà-Rabert, M.; Salvà, A. Physical Exercise Interventions for Improving Performance-Based Measures of Physical Function in Community-Dwelling, Frail Older Adults: A Systematic Review and Meta-Analysis. Arch. Phys. Med. Rehabil. 2014, 95, 753–769. [Google Scholar] [CrossRef]
- Oh, C.; Jeon, B.H.; Reid Storm, S.N.; Jho, S.; No, J.-K. The Most Effective Factors to Offset Sarcopenia and Obesity in the Older Korean: Physical Activity, Vitamin D, and Protein Intake. Nutrition 2017, 33, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Petroni, M.L.; Caletti, M.T.; Dalle Grave, R.; Bazzocchi, A.; Aparisi Gómez, M.P.; Marchesini, G. Prevention and Treatment of Sarcopenic Obesity in Women. Nutrients 2019, 11, 1302. [Google Scholar] [CrossRef] [Green Version]
- le Couteur, D.G.; Solon-Biet, S.M.; Cogger, V.C.; Ribeiro, R.; de Cabo, R.; Raubenheimer, D.; Cooney, G.J.; Simpson, S.J. Branched Chain Amino Acids, Aging and Age-Related Health. Ageing Res. Rev. 2020, 64, 101198. [Google Scholar] [CrossRef]
- Muscariello, E.; Nasti, G.; Siervo, M.; di Maro, M.; Lapi, D.; D’Addio, G.; Colantuoni, A. Dietary Protein Intake in Sarcopenic Obese Older Women. Clin. Interv. Aging 2016, 11, 133. [Google Scholar] [CrossRef] [Green Version]
- Castro, L.H.A.; de Araújo, F.H.S.; Olimpio, M.Y.M.; Primo, R.B.d.B.; Pereira, T.T.; Lopes, L.A.F.; Trindade, E.B.S.d.M.; Fernandes, R.; Oesterreich, S.A. Comparative Meta-Analysis of the Effect of Concentrated, Hydrolyzed, and Isolated Whey Protein Supplementation on Body Composition of Physical Activity Practitioners. Nutrients 2019, 11, 2047. [Google Scholar] [CrossRef]
- Giglio, B.M.; Schincaglia, R.M.; da Silva, A.S.; Fazani, I.C.S.; Monteiro, P.A.; Mota, J.F.; Cunha, J.P.; Pichard, C.; Pimentel, G.D. Whey Protein Supplementation Compared to Collagen Increases Blood Nesfatin Concentrations and Decreases Android Fat in Overweight Women: A Randomized Double-Blind Study. Nutrients 2019, 11, 2051. [Google Scholar] [CrossRef] [Green Version]
- Bumrungpert, A.; Pavadhgul, P.; Nunthanawanich, P.; Sirikanchanarod, A.; Adulbhan, A. Whey Protein Supplementation Improves Nutritional Status, Glutathione Levels, and Immune Function in Cancer Patients: A Randomized, Double-Blind Controlled Trial. J. Med. Food 2018, 21, 612–616. [Google Scholar] [CrossRef]
- Minj, S.; Anand, S. Whey Proteins and Its Derivatives: Bioactivity, Functionality, and Current Applications. Dairy 2020, 1, 233–258. [Google Scholar] [CrossRef]
- Royle, P.J.; McIntosh, G.H.; Clifton, P.M. Whey Protein Isolate and Glycomacropeptide Decrease Weight Gain and Alter Body Composition in Male Wistar Rats. Br. J. Nutr. 2008, 100, 88–93. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.-C.; Li, Z.; Liu, X.-R.; Hu, J.-N.; Liu, R.; Zhu, N.; Li, Y. The Antioxidant Effects of Whey Protein Peptide on Learning and Memory Improvement in Aging Mice Models. Nutrients 2021, 13, 2100. [Google Scholar] [CrossRef]
- Brown, M.A.; Stevenson, E.J.; Howatson, G. Whey Protein Hydrolysate Supplementation Accelerates Recovery from Exercise-Induced Muscle Damage in Females. Appl. Physiol. Nutr. Metab. 2018, 43, 324–330. [Google Scholar] [CrossRef]
- Chelliah, R.; Wei, S.; Daliri, E.B.-M.; Elahi, F.; Yeon, S.-J.; Tyagi, A.; Liu, S.; Madar, I.H.; Sultan, G.; Oh, D.-H. The Role of Bioactive Peptides in Diabetes and Obesity. Foods 2021, 10, 2220. [Google Scholar] [CrossRef]
- Kumar, M.S. Peptides and Peptidomimetics as Potential Antiobesity Agents: Overview of Current Status. Front. Nutr. 2019, 6, 11. [Google Scholar] [CrossRef]
- Tieland, M.; Dirks, M.L.; van der Zwaluw, N.; Verdijk, L.B.; van de Rest, O.; de Groot, L.C.P.G.M.; van Loon, L.J.C. Protein Supplementation Increases Muscle Mass Gain During Prolonged Resistance-Type Exercise Training in Frail Elderly People: A Randomized, Double-Blind, Placebo-Controlled Trial. J. Am. Med. Dir. Assoc. 2012, 13, 713–719. [Google Scholar] [CrossRef]
- Nabuco, H.C.G.; Tomeleri, C.M.; Fernandes, R.R.; Sugihara Junior, P.; Cavalcante, E.F.; Cunha, P.M.; Antunes, M.; Nunes, J.P.; Venturini, D.; Barbosa, D.S.; et al. Effect of Whey Protein Supplementation Combined with Resistance Training on Body Composition, Muscular Strength, Functional Capacity, and Plasma-Metabolism Biomarkers in Older Women with Sarcopenic Obesity: A Randomized, Double-Blind, Placebo-Controlled Trial. Clin. Nutr. ESPEN 2019, 32, 88–95. [Google Scholar]
- Hsu, K.J.; Liao, C.D.; Tsai, M.W.; Chen, C.N. Effects of Exercise and Nutritional Intervention on Body Composition, Metabolic Health, and Physical Performance in Adults with Sarcopenic Obesity: A Meta-Analysis. Nutrients 2019, 11, 2163. [Google Scholar] [CrossRef]
- Kim, H.; Song, W. Resistance Training Increases Fibroblast Growth Factor-21 and Irisin Levels in the Skeletal Muscle of Zucker Diabetic Fatty Rats. J. Exerc. Nutr. Biochem. 2017, 21, 50–54. [Google Scholar] [CrossRef]
- Effting, P.S.; Brescianini, S.M.S.; Sorato, H.R.; Fernandes, B.B.; Fidelis, G.d.S.P.; da Silva, P.R.L.; Silveira, P.C.L.; Nesi, R.T.; Ceddia, R.B.; Pinho, R.A. Resistance Exercise Modulates Oxidative Stress Parameters and TNF-a Content in the Heart of Mice with Diet-Induced Obesity. Arq. Bras. Cardiol. 2019, 112, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.; Liu, Y.; Zhao, W.; Chen, Q.; Guo, T.; Wei, W.; Luo, Z.; Huang, Y.; Ma, C.; Huang, F.; et al. Aerobic and Resistance Training Enhances Endothelial Progenitor Cell Function via Upregulation of Caveolin-1 in Mice with Type 2 Diabetes. Stem Cell Res. Ther. 2020, 11, 10. [Google Scholar] [CrossRef] [Green Version]
- Eo, H.; Park, J.E.; Jeon, Y.; Lim, Y. Ameliorative Effect of Ecklonia Cava Polyphenol Extract on Renal Inflammation Associated with Aberrant Energy Metabolism and Oxidative Stress in High Fat Diet-Induced Obese Mice. J. Agric. Food Chem. 2017, 65, 3811–3818. [Google Scholar] [CrossRef]
- Lee, H.; Lim, Y. Tocotrienol-Rich Fraction Supplementation Reduces Hyperglycemia-Induced Skeletal Muscle Damage through Regulation of Insulin Signaling and Oxidative Stress in Type 2 Diabetic Mice. J. Nutr. Biochem. 2018, 57, 77–85. [Google Scholar] [CrossRef]
- Lee, D.; Shook, R.P.; Drenowatz, C.; Blair, S.N. Physical Activity and Sarcopenic Obesity: Definition, Assessment, Prevalence and Mechanism. Future Sci. OA 2016, 2, FSO127. [Google Scholar] [CrossRef] [Green Version]
- Lyu, Q.; Wen, Y.; He, B.; Zhang, X.; Chen, J.; Sun, Y.; Zhao, Y.; Xu, L.; Xiao, Q.; Deng, H. The Ameliorating Effects of Metformin on Disarrangement Ongoing in Gastrocnemius Muscle of Sarcopenic and Obese Sarcopenic Mice. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2022, 1868, 166508. [Google Scholar] [CrossRef]
- Sepandi, M.; Samadi, M.; Shirvani, H.; Alimohamadi, Y.; Taghdir, M.; Goudarzi, F.; Akbarzadeh, I. Effect of Whey Protein Supplementation on Weight and Body Composition Indicators: A Meta-Analysis of Randomized Clinical Trials. Clin. Nutr. ESPEN 2022, 50, 74–83. [Google Scholar] [CrossRef]
- Zemel, B.S. Body Composition during Growth and Development. In Human Growth and Development; Elsevier: Amsterdam, The Netherlands, 2022; pp. 517–545. [Google Scholar]
- Welch, A.A.; Hayhoe, R.P.G.; Cameron, D. The Relationships between Sarcopenic Skeletal Muscle Loss during Ageing and Macronutrient Metabolism, Obesity and Onset of Diabetes. Proc. Nutr. Soc. 2020, 79, 158–169. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; He, J.; Wang, H.; Zhu, D.; Bi, Y. Adipose Morphology: A Critical Factor in Regulation of Human Metabolic Diseases and Adipose Tissue Dysfunction. Obes. Surg. 2020, 30, 5086–5100. [Google Scholar] [CrossRef]
- Griffen, C.; Duncan, M.; Hattersley, J.; Weickert, M.O.; Dallaway, A.; Renshaw, D. Effects of Resistance Exercise and Whey Protein Supplementation on Skeletal Muscle Strength, Mass, Physical Function, and Hormonal and Inflammatory Biomarkers in Healthy Active Older Men: A Randomised, Double-Blind, Placebo-Controlled Trial. Exp. Gerontol. 2022, 158, 111651. [Google Scholar] [CrossRef]
- Li, F.; Li, Y.; Duan, Y.; Hu, C.-A.A.; Tang, Y.; Yin, Y. Myokines and Adipokines: Involvement in the Crosstalk between Skeletal Muscle and Adipose Tissue. Cytokine Growth Factor Rev. 2017, 33, 73–82. [Google Scholar] [CrossRef]
- Zhou, X.; Wu, W.; Chen, J.; Wang, X.; Wang, Y. AMP-Activated Protein Kinase Is Required for the Anti-Adipogenic Effects of Alpha-Linolenic Acid. Nutr. Metab. 2015, 12, 10. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Jin, H.; Chei, S.; Oh, H.-J.; Lee, J.-Y.; Lee, B.-Y. Effect of Dietary Silk Peptide on Obesity, Hyperglycemia, and Skeletal Muscle Regeneration in High-Fat Diet-Fed Mice. Cells 2020, 9, 377. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Fernández, L.; Fernández-Galilea, M.; Felix-Soriano, E.; Escoté, X.; González-Muniesa, P.; Moreno-Aliaga, M.J. Inflammation and Oxidative Stress in Adipose Tissue. In Obesity; Elsevier: Amsterdam, The Netherlands, 2018; pp. 63–92. [Google Scholar]
- Lenhard, J.M. Lipogenic Enzymes as Therapeutic Targets for Obesity and Diabetes. Curr. Pharm. Des. 2011, 17, 325–331. [Google Scholar] [CrossRef]
- Kobayashi, M.; Deguchi, Y.; Nozaki, Y.; Higami, Y. Contribution of PGC-1α to Obesity- and Caloric Restriction-Related Physiological Changes in White Adipose Tissue. Int. J. Mol. Sci. 2021, 22, 6025. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, L.; Li, B.; Jiang, H.; Duan, Y.; Xie, Z.; Shuai, L.; Li, J.; Li, J. AMP-Activated Protein Kinase (AMPK) Regulates Energy Metabolism through Modulating Thermogenesis in Adipose Tissue. Front. Physiol. 2018, 9, 122. [Google Scholar] [CrossRef] [Green Version]
- Stern, J.H.; Rutkowski, J.M.; Scherer, P.E. Adiponectin, Leptin, and Fatty Acids in the Maintenance of Metabolic Homeostasis through Adipose Tissue Crosstalk. Cell Metab. 2016, 23, 770–784. [Google Scholar] [CrossRef] [Green Version]
- Khodamoradi, K.; Khosravizadeh, Z.; Seetharam, D.; Mallepalli, S.; Farber, N.; Arora, H. The Role of Leptin and Low Testosterone in Obesity. Int. J. Impot. Res. 2022; ahead of print. [Google Scholar] [CrossRef]
- Lago, F.; Gómez, R.; Gómez-Reino, J.J.; Dieguez, C.; Gualillo, O. Adipokines as Novel Modulators of Lipid Metabolism. Trends Biochem. Sci. 2009, 34, 500–510. [Google Scholar] [CrossRef]
- Kim, J.-E.; Kim, J.-S.; Jo, M.-J.; Cho, E.; Ahn, S.-Y.; Kwon, Y.-J.; Ko, G.-J. The Roles and Associated Mechanisms of Adipokines in Development of Metabolic Syndrome. Molecules 2022, 27, 334. [Google Scholar] [CrossRef]
- Wilhelmsen, A.; Tsintzas, K.; Jones, S.W. Recent Advances and Future Avenues in Understanding the Role of Adipose Tissue Cross Talk in Mediating Skeletal Muscle Mass and Function with Ageing. Geroscience 2021, 43, 85–110. [Google Scholar] [CrossRef]
- Kohara, K.; Ochi, M.; Tabara, Y.; Nagai, T.; Igase, M.; Miki, T. Leptin in Sarcopenic Visceral Obesity: Possible Link between Adipocytes and Myocytes. PLoS ONE 2011, 6, e24633. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Zhu, Y.; Schultz, R.D.; Li, N.; He, Z.; Zhang, Z.; Caron, A.; Zhu, Q.; Sun, K.; Xiong, W.; et al. Partial Leptin Reduction as an Insulin Sensitization and Weight Loss Strategy. Cell Metab. 2019, 30, 706–719. e6. [Google Scholar] [CrossRef]
- Jaiswal, N.; Gavin, M.G.; Quinn, W.J.; Luongo, T.S.; Gelfer, R.G.; Baur, J.A.; Titchenell, P.M. The Role of Skeletal Muscle Akt in the Regulation of Muscle Mass and Glucose Homeostasis. Mol. Metab. 2019, 28, 1–13. [Google Scholar] [CrossRef]
- Westerblad, H.; Bruton, J.D.; Katz, A. Skeletal Muscle: Energy Metabolism, Fiber Types, Fatigue and Adaptability. Exp. Cell Res. 2010, 316, 3093–3099. [Google Scholar] [CrossRef]
- Han, M.J.; Choung, S.-Y. Codonopsis Lanceolata Ameliorates Sarcopenic Obesity via Recovering PI3K/Akt Pathway and Lipid Metabolism in Skeletal Muscle. Phytomedicine 2022, 96, 153877. [Google Scholar] [CrossRef]
- Lee, Y.-S.; Park, E.-J.; Kim, S.-M.; Kim, J.-Y.; Lee, H.-J. Anti-Sarcopenic Obesity Effects of Lonicera Caerulea Extract in High-Fat Diet-Fed Mice. Antioxidants 2021, 10, 1633. [Google Scholar] [CrossRef]
- Lee, K.; Jin, H.; Chei, S.; Lee, J.-Y.; Oh, H.-J.; Lee, B.-Y. Dietary Silk Peptide Prevents High-Fat Diet-Induced Obesity and Promotes Adipose Browning by Activating AMP-Activated Protein Kinase in Mice. Nutrients 2020, 12, 201. [Google Scholar] [CrossRef] [Green Version]
- Wu, G. Amino Acids: Metabolism, Functions, and Nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef]
- Caldow, M.K.; Ham, D.J.; Godeassi, D.P.; Chee, A.; Lynch, G.S.; Koopman, R. Glycine Supplementation during Calorie Restriction Accelerates Fat Loss and Protects against Further Muscle Loss in Obese Mice. Clin. Nutr. 2016, 35, 1118–1126. [Google Scholar] [CrossRef]
- Mantuano, P.; Boccanegra, B.; Bianchini, G.; Conte, E.; de Bellis, M.; Sanarica, F.; Camerino, G.M.; Pierno, S.; Cappellari, O.; Allegretti, M.; et al. BCAAs and Di-Alanine Supplementation in the Prevention of Skeletal Muscle Atrophy: Preclinical Evaluation in a Murine Model of Hind Limb Unloading. Pharmacol. Res. 2021, 171, 105798. [Google Scholar] [CrossRef]
- Luciano, T.F.; Marques, S.O.; Pieri, B.L.; de Souza, D.R.; Araújo, L.v.; Nesi, R.T.; Scheffer, D.L.; Comin, V.H.; Pinho, R.A.; Muller, A.P.; et al. Responses of Skeletal Muscle Hypertrophy in Wistar Rats to Different Resistance Exercise Models. Physiol. Res. 2017, 317–323. [Google Scholar] [CrossRef]
- Gacitua, T.; Karachon, L.; Romero, E.; Parra, P.; Poblete, C.; Russell, J.; Rodrigo, R. Effects of Resistance Training on Oxidative Stress-Related Biomarkers in Metabolic Diseases: A Review. Sport Sci. Health 2018, 14, 1–7. [Google Scholar] [CrossRef]
- Fry, C.S.; Drummond, M.J.; Glynn, E.L.; Dickinson, J.M.; Gundermann, D.M.; Timmerman, K.L.; Walker, D.K.; Dhanani, S.; Volpi, E.; Rasmussen, B.B. Aging Impairs Contraction-Induced Human Skeletal Muscle MTORC1 Signaling and Protein Synthesis. Skelet. Muscle 2011, 1, 11. [Google Scholar] [CrossRef] [Green Version]
- Leal, L.G.; Lopes, M.A.; Batista, M.L. Physical Exercise-Induced Myokines and Muscle-Adipose Tissue Crosstalk: A Review of Current Knowledge and the Implications for Health and Metabolic Diseases. Front. Physiol. 2018, 9, 1307. [Google Scholar] [CrossRef]
- Nair, A.; Jacob, S. A Simple Practice Guide for Dose Conversion between Animals and Human. J. Basic Clin. Pharm. 2016, 7, 27. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Direction | Sequence (5′–3′) |
|---|---|---|
| GAPDH | Forward | AGG TTG TCT CCT GCG ACT |
| Reverse | TGC TGT AGC CGT ATT CAT TGT CA | |
| Leptin | Forward | GAG ACC CCT GTG TCG GTT C |
| Reverse | CTG CGT GTG TGA AAT GTC ATT G | |
| Adiponectin | Forward | TGA CGA CAC CAA AAG GGC TC |
| Reverse | ACC TGC ACA AGT TCC CTT GG | |
| C/EBPα | Forward | CGC AAG AGC CGA GAT AAA GC |
| Reverse | CAC GGC TCA GCT GTT CCA | |
| PPAR-γ | Forward | CGC TGA TGC ACT GCC TAT GA |
| Reverse | AGA GGT CCA CAG AGC TGA TTC C |
| Group | CON | OB | RE | WP. | WPE | |
|---|---|---|---|---|---|---|
| Body Composition (% of Total Mass) | ||||||
| Before 8 weeks of RE and WP (right after sarcopenic obesity induction) | Fat Mass | 22.87 ± 0.90 b | 36.82 ± 1.36 a | 37.13 ± 1.49 a | 36.59 ± 1.56 a | 36.82 ± 1.28 a |
| Lean Mass | 74.58 ± 0.87 a | 61.08 ± 1.34 b | 60.82 ± 1.47 a | 61.38 ± 1.49 a | 61.06 ± 1.22 a | |
| After 8 weeks of RE and WP | Fat Mass | 25.22 ± 1.07 b | 40.51 ± 1.02 a | 39.89 ± 1.45 a | 37.61 ± 1.75 a | 36.70 ± 1.64 a |
| Lean Mass | 72.11 ± 1.03 a | 57.41 ± 0.99 b | 57.91 ± 1.43 b | 60.27 ± 1.66 b | 61.02 ± 1.56 b | |
| Difference made during 8 weeks of RE and WP | Fat Mass | 2.36 ± 0.83 abc | 3.69 ± 0.73 a | 2.76 ± 0.49 ab | 0.73 ± 0.66 bc | 0.01 ± 1.20 c |
| Lean Mass | −2.47 ± 0.81 bc | −3.67 ± 0.70 c | −2.90 ± 0.49 bc | −0.81 ± 0.63 ab | −0.20 ± 1.12 a | |
| Body Weight (g) | ||||||
| Before 8 weeks of RE and WP (sarcopenic obesity induced) | 31.56 ± 0.75 b | 40.46 ± 0.07 a | 40.25 ± 1.24 a | 41.16 ± 2.05 a | 40.01 ± 1.49 a | |
| After 8 weeks of RE and WP | 31.88 ± 0.85 c | 45.09 ± 0.92 a | 42.16 ± 1.49 ab | 42.63 ± 2.66 ab | 39.62 ± 2.36 b | |
| Difference made during 8 weeks of RE and WP | 0.33 ± 0.35 b | 4.63 ± 0.58 a | 1.90 ± 0.56 b | 1.47 ± 1.08 b | −1.39 ± 1.02 b | |
| Fat Weight (% B.W.) (After 8 weeks of RE and WP) | ||||||
| Total Fat | 4.96 ± 1.43 c | 16.03 ± 2.51 a | 14.98 ± 3.04 ab | 13.06 ± 3.54 b | 12.53 ± 3.65 b | |
| Subcutaneous Fat | 2.68 ± 0.96 b | 10.00 ± 2.53 a | 9.41 ± 2.16 a | 8.40 ± 3.33 a | 7.54 ± 2.83 a | |
| Epididymis Fat | 1.93 ± 0.51 d | 5.72 ± 0.67 a | 5.21 ± 1.07 ab | 4.34 ± 0.68 c | 4.72 ±1.07 bc | |
| Brown Adipose Tissue | 0.34 ± 0.09 | 0.32 ± 0.08 | 0.35 ± 0.06 | 0.32 ± 0.14 | 0.28 ± 0.06 | |
| Muscle Weight (% B.W.) (After 8 weeks of RE and WP) | ||||||
| Total Muscle | 2.20 ± 0.14 a | 1.60 ± 0.19 b | 1.67 ± 0.22 b | 1.70 ± 0.32 b | 1.80 ± 0.22 b | |
| Gastrocnemius | 1.02 ± 0.07 a | 0.72 ± 0.07 c | 0.79 ± 0.09 bc | 0.77 ± 0.14 bc | 0.83 ± 0.08 b | |
| Quadricep | 1.13 ± 0.09 a | 0.84 ± 0.12 b | 0.84 ± 0.14 b | 0.89 ± 0.18 b | 0.92 ± 0.16 b | |
| Soleus | 0.05 ± 0.01 a | 0.04 ± 0.00 b | 0.04 ± 0.01 b | 0.04 ± 0.00 b | 0.05 ± 0.01 b | |
| Muscle Strength (strength(N)/B.W(g)) (After 8 weeks of RE and WP) | 0.034 ± 0.002 a | 0.019 ± 0.002 c | 0.025 ± 0.001 b | 0.018 ± 0.003 c | 0.027 ± 0.002 b | |
| Daily Diet Intake (g) | 3.25 ± 0.11 a | 2.88 ± 0.08 b | 2.62 ± 0.08 bc | 2.70 ± 0.13 bc | 2.51 ± 0.08 c | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, G.; Lim, Y. Effects of Whey Peptide Supplementation on Sarcopenic Obesity in High-Fat Diet-Fed Mice. Nutrients 2022, 14, 4402. https://doi.org/10.3390/nu14204402
Lim G, Lim Y. Effects of Whey Peptide Supplementation on Sarcopenic Obesity in High-Fat Diet-Fed Mice. Nutrients. 2022; 14(20):4402. https://doi.org/10.3390/nu14204402
Chicago/Turabian StyleLim, Gahyun, and Yunsook Lim. 2022. "Effects of Whey Peptide Supplementation on Sarcopenic Obesity in High-Fat Diet-Fed Mice" Nutrients 14, no. 20: 4402. https://doi.org/10.3390/nu14204402
APA StyleLim, G., & Lim, Y. (2022). Effects of Whey Peptide Supplementation on Sarcopenic Obesity in High-Fat Diet-Fed Mice. Nutrients, 14(20), 4402. https://doi.org/10.3390/nu14204402