
Effect of Different Functional Food Supplements on the Gut Microbiota of Prediabetic Indonesian Individuals during Weight Loss


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Population
2.2. DNA Isolation and Sequencing of the V3–V4 Region of the 16S rRNA Gene
2.3. Statistical Analyses
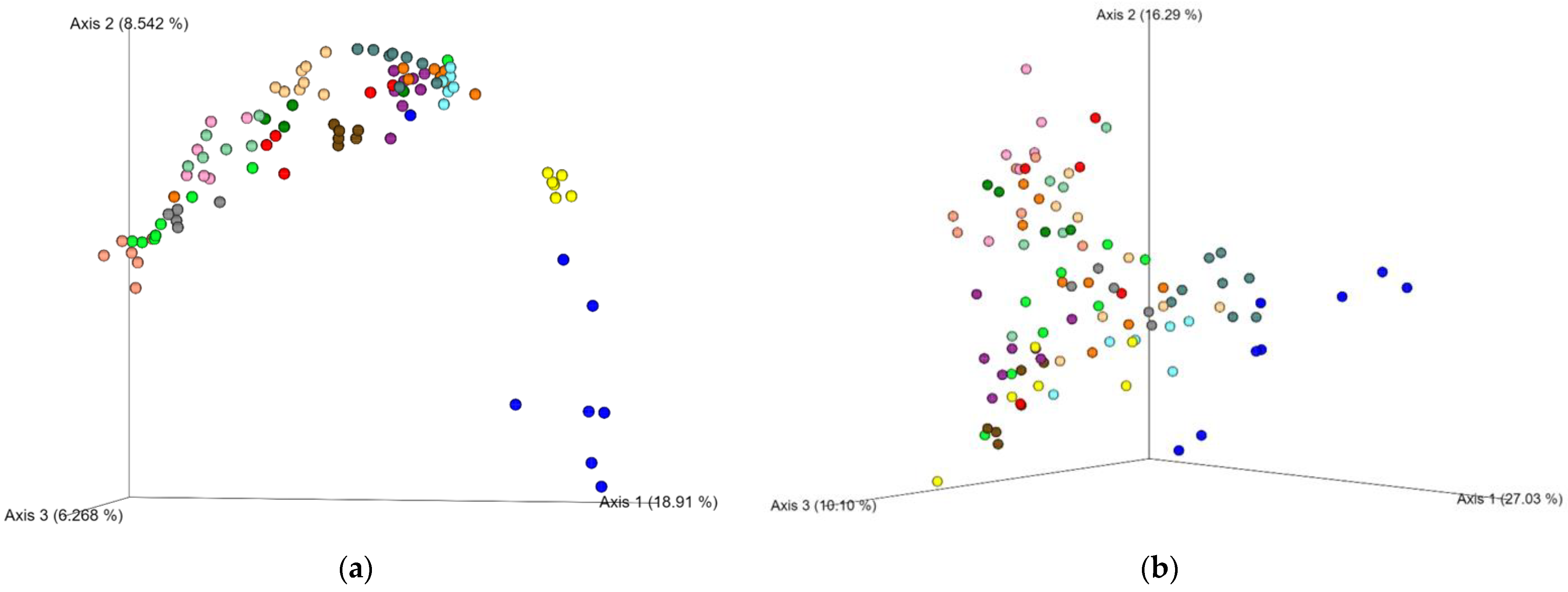
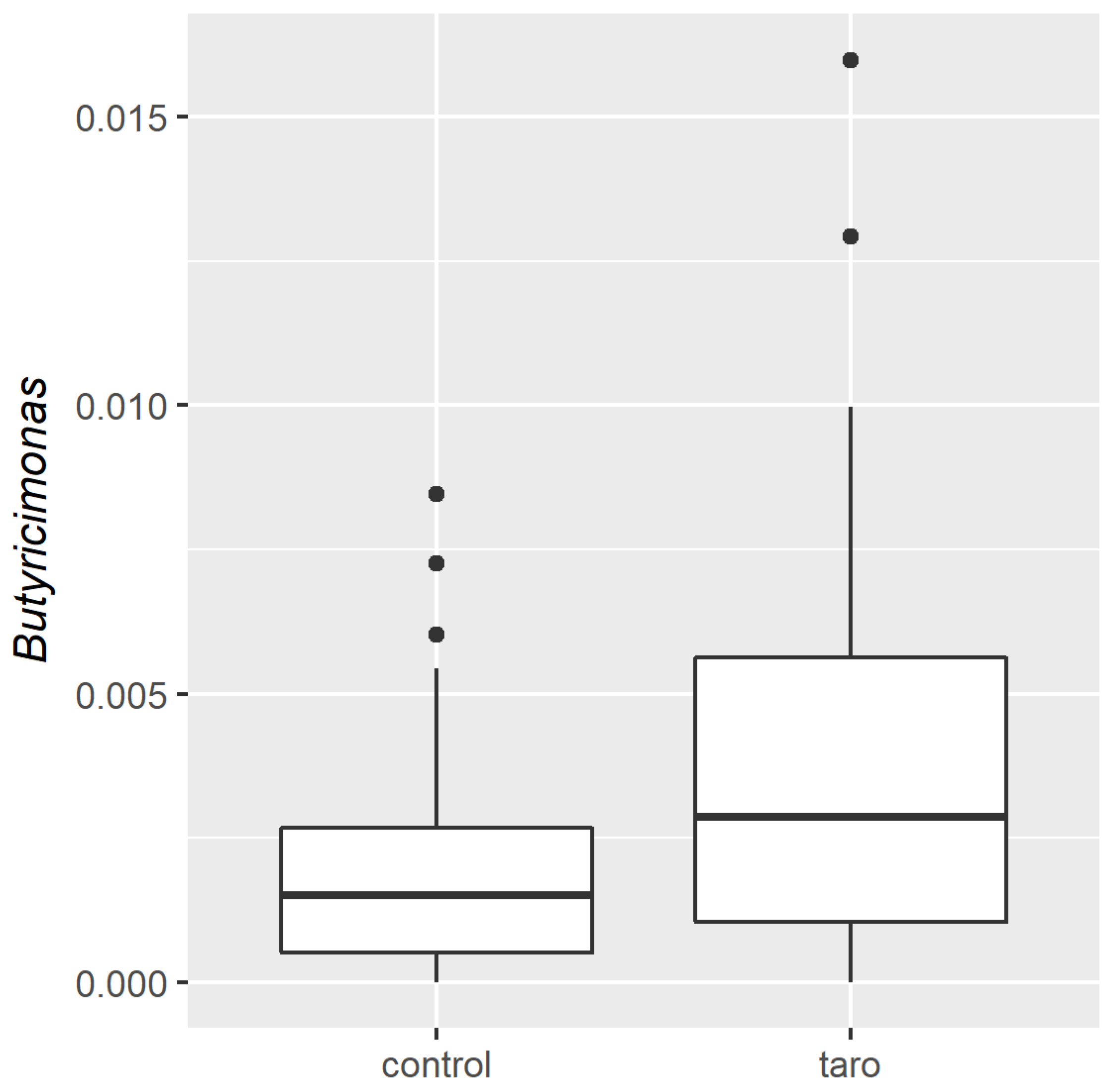
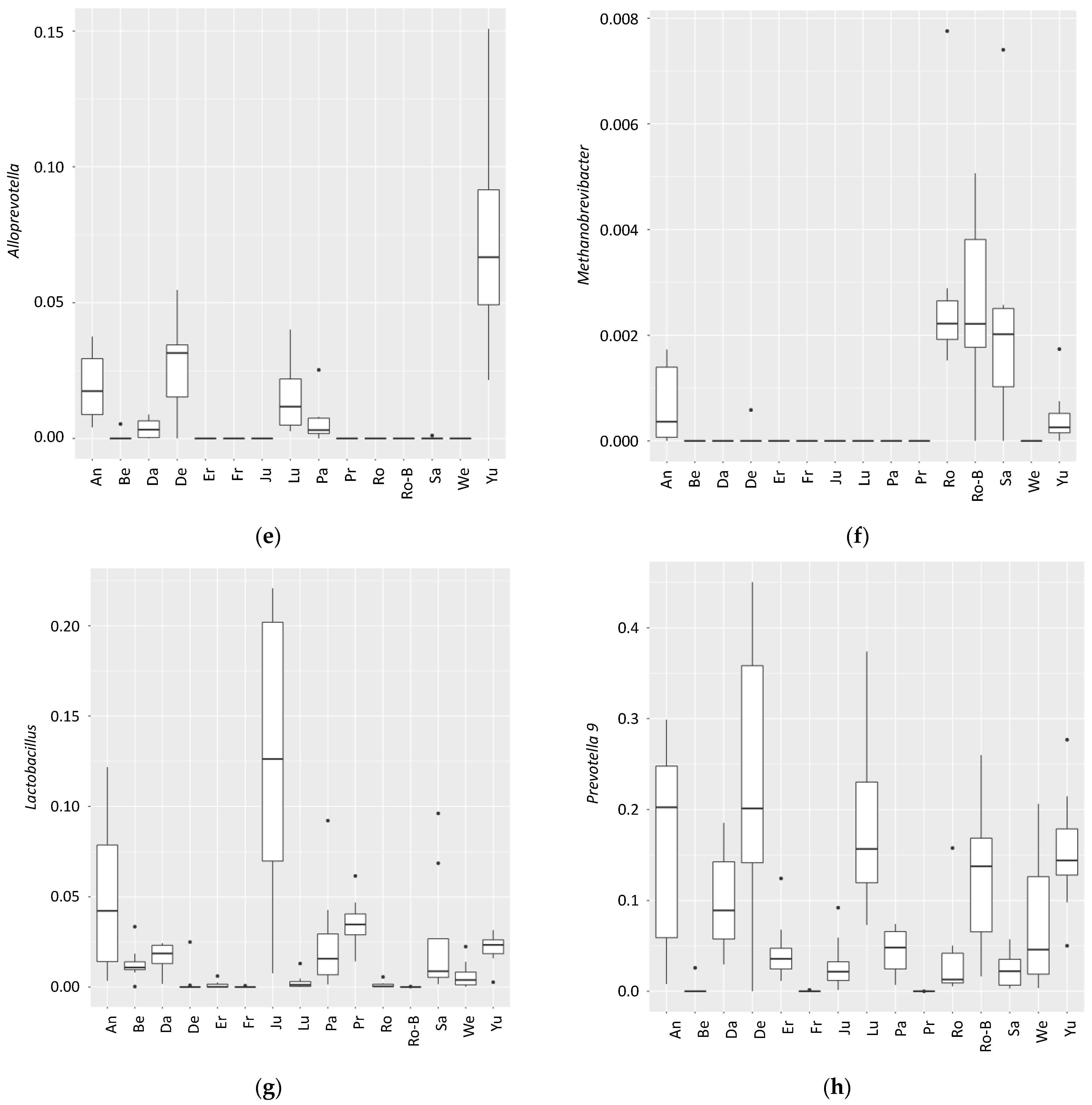
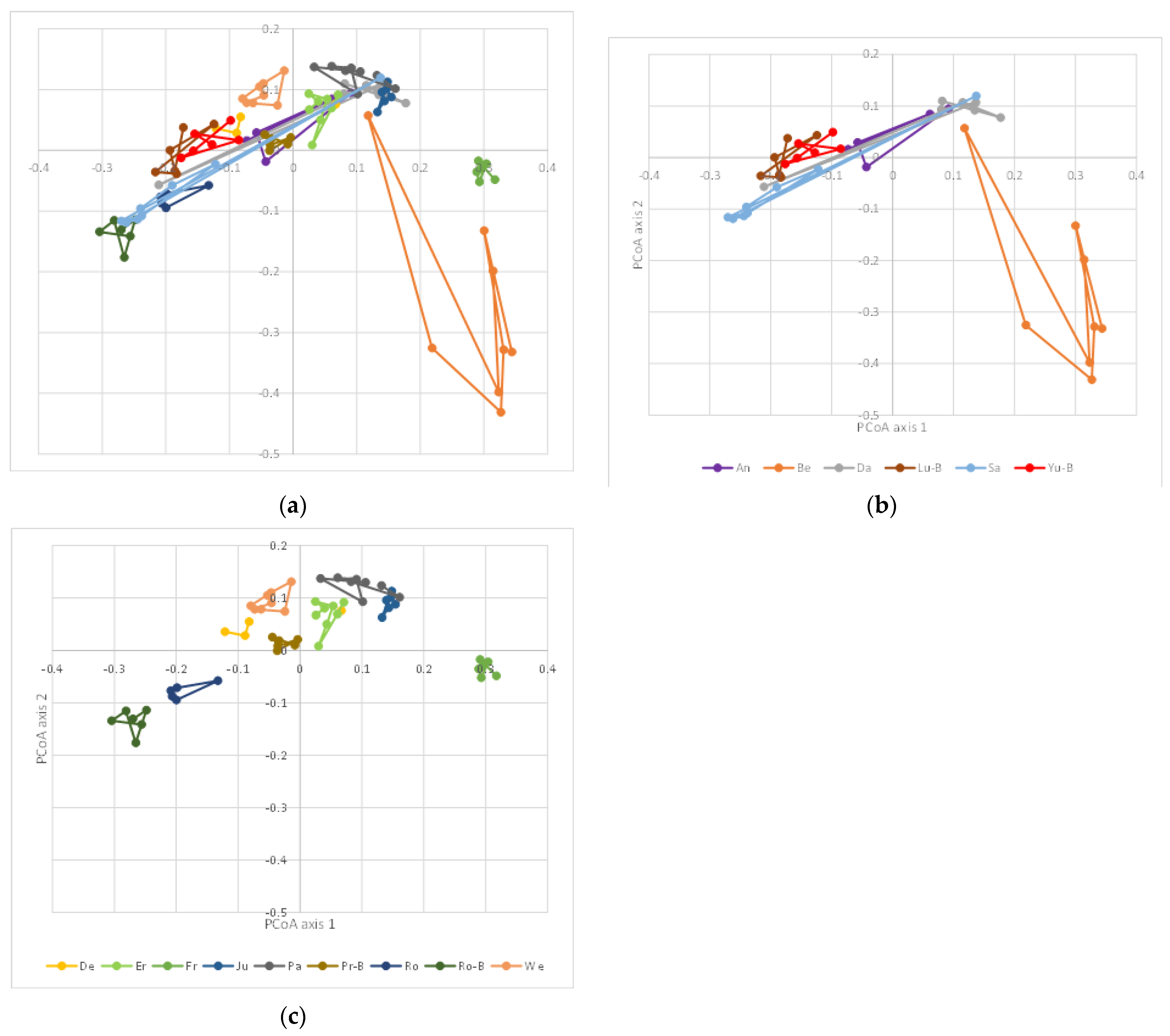
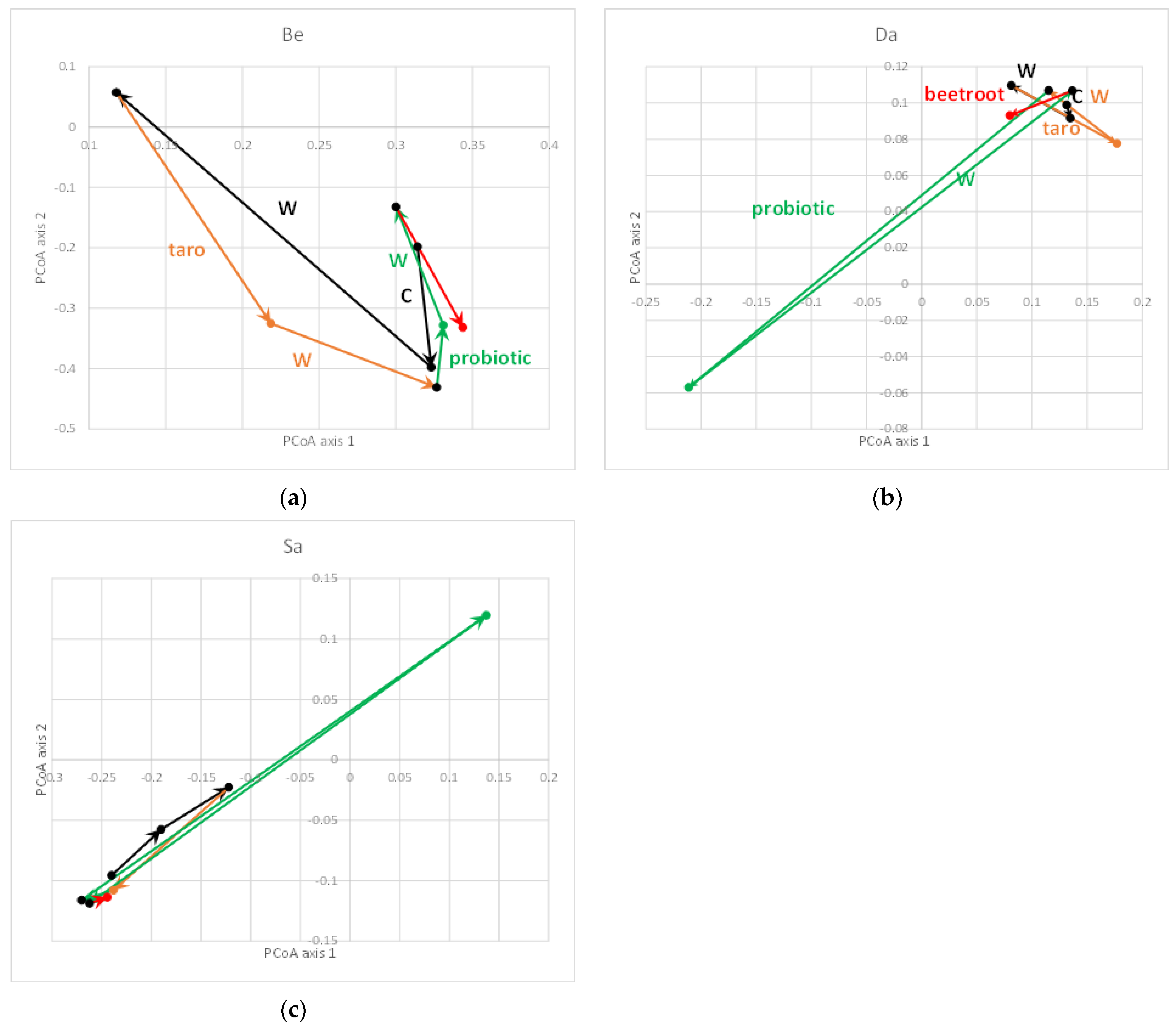
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- UN-News. UN Projects World Population to Reach 8.5 Billion by 2030, Driven by Growth in Developing Countries. Available online: https://news.un.org/en/story/2015/07/505352-un-projects-world-population-reach-85-billion-2030-driven-growth-developing (accessed on 29 March 2020).
- Wild, S.; Roglic, G.; Green, A.; Sicree, R.; King, H. Global prevalence of diabetes: Estimates for the year 2000 and projections for 2030. Diabetes Care 2004, 27, 1047–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapiro, H.; Suez, J.; Elinav, E. Personalized microbiome-based approaches to metabolic syndrome management and prevention. J. Diabetes 2017, 9, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, F.H.; Tremaroli, V.; Nookaew, I.; Bergstrom, G.; Behre, C.J.; Fagerberg, B.; Nielsen, J.; Backhed, F. Gut metagenome in European women with normal, impaired and diabetic glucose control. Nature 2013, 498, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Onwueme, I.C. The Tropical Tuber Crops—Yams, Cassava, Sweet Potato and Cocoyams; John Wiley and Sons: Chichester, UK, 1978. [Google Scholar]
- Asha Devi, A. Genetic diversity analysis in taro using molecular markers—An overview. J. Root Crop. 2012, 38, 15–25. [Google Scholar]
- Solossa, A.H.; Sastrahidayat, I.R.; Hakim, L. Home gardens of the local community surrounding Lake Ayamaru, West Papua province, and its consequences for tourism development and lake conservation. J. Biodivers. Environ. Sci. 2017, 3, 1–11. [Google Scholar]
- Iskandar, J.; Iskandar, B.S. Various Plants of Traditional Rituals: Ethnobotanical Research Among The Baduy Community. Biosaintifika J. Biol. Biol. Educ. 2017, 9, 114–125. [Google Scholar] [CrossRef] [Green Version]
- Kreike, C.M.; Van Eck, H.J.; Lebot, V. Genetic diversity of taro, Colocasia esculenta (L.) Schott, in Southeast Asia and the Pacific. Theor. Appl. Genet. 2004, 109, 761–768. [Google Scholar] [CrossRef]
- Prana, M.S.; Kuswara, T. Taro Cultivation: Diversification to Support National Food Security; Medikom Pustaka Mandiri: Bogor, Indonesia, 2002; 75p. [Google Scholar]
- Surono, I.S.; Wardana, A.A.; Waspodo, P.; Saksono, B.; Verhoeven, J.; Venema, K. Effect of functional food ingredients on gut microbiota in a rodent diabetes model. Nutr. Metab. 2020, 17, 77. [Google Scholar] [CrossRef]
- Surono, I.S.; Venema, K. Modulation of Gut Microbiota Profile and Short-Chain Fatty Acids of Rats Fed with Taro Flour or Taro Starch. Int. J. Microbiol. 2020, 2020, 8893283. [Google Scholar] [CrossRef] [PubMed]
- Surono, I.S.; Verhoeven, J.; Venema, K. Low glycemic load after digestion of native starch from the indigenous tuber Belitung Taro (Xanthosoma sagittifolium) in a dynamic in vitro model of the upper GI tract (TIM-1). Food Nutr. Res. 2020, 64, 4263. [Google Scholar] [CrossRef] [PubMed]
- Venema, K.; Verhoeven, J.; Surono, I.S.; Waspodo, P.; Simatupang, A.; Kusuma, P.D. Differential glucose bioaccessibility from native and modified taro-starches in the absence or presence of beet juice. CyTA-J. Food 2020, 18, 670–674. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO). Joint FAO/WHO Working Group on Drafting Guidelines for the Evaluation of Probiotics in Food: Health and Nutritional Properties of Probiotics in Food including Powder Milk with Live Lactic Acid Bacteria; Food and Agriculture Organization of the United Nations: Rome, Italy, 2001. [Google Scholar]
- Surono, I.; Verhoeven, J.; Verbruggen, S.; Venema, K. Microencapsulation increases survival of the probiotic Lactobacillus plantarum IS-10506, but not Enterococcus faecium IS-27526 in a dynamic, computer-controlled in vitro model of the upper gastrointestinal tract. J. Appl. Microbiol. 2018, 124, 1604–1609. [Google Scholar] [CrossRef] [PubMed]
- Kusuma, R.J.; Azzyati, F.; Purbarani, G.; Sulistyorini, R.; Nofiartika, F.; Huriyati, E. Effect of Traditional Fermented Buffalo Milk (Dadih) On Body Weight, Adipose Tissue Mass and Adiposity Inflammation in High Fat-Induced Obese Rats. EC Nutr. 2015, 1, 106–114. [Google Scholar]
- Prakoeswa, C.R.S.; Herwanto, N.; Prameswari, R.; Astari, L.; Sawitri, S.; Hidayati, A.N.; Indramaya, D.M.; Kusumowidagdo, E.R.; Surono, I.S. Lactobacillus plantarum IS-10506 supplementation reduced SCORAD in children with atopic dermatitis. Benef. Microbes 2017, 8, 833–840. [Google Scholar] [CrossRef]
- Surono, I.S.; Martono, P.D.; Kameo, S.; Suradji, E.W.; Koyama, H. Effect of probiotic L. plantarum IS-10506 and zinc supplementation on humoral immune response and zinc status of Indonesian pre-school children. J. Trace Elem. Med. Biol. 2014, 28, 465–469. [Google Scholar] [CrossRef]
- Dharmawan, J.; Surono, I.S.; Kun, L.Y. Adhesion properties of indigenous dadih lactic acid bacteria on human intestinal mucosal surface. Asian-Australas. J. Anim. Sci. 2006, 19, 751–755. [Google Scholar] [CrossRef]
- Kujala, T.S.; Vienola, M.S.; Klika, K.D.; Loponen, J.M.; Pihlaja, K. Betalain and phenolic compositions of four beetroot (Beta vulgaris) cultivars. Eur. Food Res. Technol. 2002, 214, 505–510. [Google Scholar] [CrossRef]
- Wootton-Beard, P.C.; Ryan, L. Short communication: A beetroot juice shot is a significant and convenient source of bioaccessible antioxidants. J. Funct. Foods 2011, 3, 329–334. [Google Scholar] [CrossRef]
- Henquin, J.C.; Rahier, J. Pancreatic alpha cell mass in European subjects with type 2 diabetes. Diabetologia 2011, 54, 1720–1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsson, B.; Svardsudd, K.; Welin, L.; Wilhelmsen, L.; Bjorntorp, P.; Tibblin, G. Abdominal adipose tissue distribution, obesity, and risk of cardiovascular disease and death: 13 year follow up of participants in the study of men born in 1913. Br. Med. J. (Clin. Res. Ed.) 1984, 288, 1401–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahimi, R.; Nikfar, S.; Larijani, B.; Abdollahi, M. A review on the role of antioxidants in the management of diabetes and its complications. Biomed. Pharmacother. 2005, 59, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, A.; Yamashita, H.; Hiemori, M.; Inagaki, E.; Kimoto, M.; Okamoto, M.; Tsuji, H.; Memon, A.N.; Mohammadio, A.; Natori, Y. Characterization of inhibitors of postprandial hyperglycemia from the leaves of Nerium indicum. J. Nutr. Sci. Vitaminol. 2007, 53, 166–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnbaugh, P.; Ley, R.; Mahowald, M.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Buchner, A.; Lang, A.-G. Statistical power analyses using G*Power 3.1: Tests for correlation and regression analyses. Behav. Res. Methods 2009, 41, 1149–1160. [Google Scholar] [CrossRef] [Green Version]
- Gayoso-Diz, P.; Otero-González, A.; Rodriguez-Alvarez, M.X.; Gude, F.; García, F.; De Francisco, A.; Quintela, A.G. Insulin resistance (HOMA-IR) cut-off values and the metabolic syndrome in a general adult population: Effect of gender and age: EPIRCE cross-sectional study. BMC Endocr. Disord. 2013, 13, 47. [Google Scholar] [CrossRef] [Green Version]
- Vujkovic-Cvijin, I.; Sklar, J.; Jiang, L.; Natarajan, L.; Knight, R.; Belkaid, Y. Host variables confound gut microbiota studies of human disease. Nature 2020, 587, 448–454. [Google Scholar] [CrossRef]
- Chung, W.S.F.; Walker, A.W.; Bosscher, D.; Garcia-Campayo, V.; Wagner, J.; Parkhill, J.; Duncan, S.H.; Flint, H.J. Relative abundance of the Prevotella genus within the human gut microbiota of elderly volunteers determines the inter-individual responses to dietary supplementation with wheat bran arabinoxylan-oligosaccharides. BMC Microbiol. 2020, 20, 283. [Google Scholar] [CrossRef]
- Fassarella, M.; Blaak, E.E.; Penders, J.; Nauta, A.; Smidt, H.; Zoetendal, E.G. Gut microbiome stability and resilience: Elucidating the response to perturbations in order to modulate gut health. Gut 2021, 70, 595–605. [Google Scholar] [CrossRef]
- Costea, P.I.; Hildebrand, F.; Arumugam, M.; Backhed, F.; Blaser, M.J.; Bushman, F.D.; de Vos, W.M.; Ehrlich, S.D.; Fraser, C.M.; Hattori, M.; et al. Enterotypes in the landscape of gut microbial community composition. Nat. Microbiol. 2018, 3, 8–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajilic-Stojanovic, M.; de Vos, W.M. The first 1000 cultured species of the human gastrointestinal microbiota. FEMS Microbiol. Rev. 2014, 38, 996–1047. [Google Scholar] [CrossRef] [PubMed]
- Claesson, M.J.; Cusack, S.; O’Sullivan, O.; Greene-Diniz, R.; de Weerd, H.; Flannery, E.; Marchesi, J.R.; Falush, D.; Dinan, T.; Fitzgerald, G.; et al. Composition, variability, and temporal stability of the intestinal microbiota of the elderly. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4586–4591. [Google Scholar] [CrossRef] [Green Version]
- Tap, J.; Mondot, S.; Levenez, F.; Pelletier, E.; Caron, C.; Furet, J.P.; Ugarte, E.; Munoz-Tamayo, R.; Paslier, D.L.; Nalin, R.; et al. Towards the human intestinal microbiota phylogenetic core. Environ. Microbiol. 2009, 11, 2574–2584. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef] [Green Version]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Aguirre, M.; Eck, A.; Koenen, M.E.; Savelkoul, P.H.; Budding, A.E.; Venema, K. Diet drives quick changes in the metabolic activity and composition of human gut microbiota in a validated in vitro gut model. Res. Microbiol. 2016, 167, 114–125. [Google Scholar] [CrossRef]
- Venema, K.; van Nuenen, H.M.C.; van den Heuvel, E.G.; Pool, W.; van der Vossen, J.M.B.M. The Effect of Lactulose on the Composition of the Intestinal Microbiota and Short-chain Fatty Acid Production in Human Volunteers and a Computer-controlled Model of the Proximal Large Intestine. Microb. Ecol. Health Dis. 2003, 15, 94–105. [Google Scholar] [CrossRef] [Green Version]
- Surono, I.S.; Wardana, A.A.; Waspodo, P.; Saksono, B.; Venema, K. Effect of Taro Starch, Beet Juice, Probiotic, and/or Psicose on Gut Microbiota in a Type 2 Diabetic Rat Model: A Pilot Study. J. Nutr. Metab. 2021, 2021, 1825209. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, M.; Takagaki, A.; Matsumoto, K.; Kato, Y.; Goto, K.; Benno, Y. Butyricimonas synergistica gen. nov., sp. nov. and Butyricimonas virosa sp. nov., butyric acid-producing bacteria in the family ‘Porphyromonadaceae’ isolated from rat faeces. Int. J. Syst. Evol. Microbiol. 2009, 59, 1748–1753. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, M.; Tanaka, Y.; Benno, Y.; Ohkuma, M. Butyricimonas faecihominis sp. nov. and Butyricimonas paravirosa sp. nov., isolated from human faeces, and emended description of the genus Butyricimonas. Int. J. Syst. Evol. Microbiol. 2014, 64, 2992–2997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovatcheva-Datchary, P.; Egert, M.; Maathuis, A.; Rajilic-Stojanovic, M.; de Graaf, A.A.; Smidt, H.; de Vos, W.M.; Venema, K. Linking phylogenetic identities of bacteria to starch fermentation in an in vitro model of the large intestine by RNA-based stable isotope probing. Environ. Microbiol. 2009, 11, 914–926. [Google Scholar] [CrossRef]
- Rose, D.J.; Venema, K.; Keshavarzian, A.; Hamaker, B.R. Starch-entrapped microspheres show a beneficial fermentation profile and decrease in potentially harmful bacteria during in vitro fermentation in faecal microbiota obtained from patients with inflammatory bowel disease. Br. J. Nutr. 2010, 103, 1514–1524. [Google Scholar] [CrossRef] [Green Version]
- Sanders, M.E. Probiotics and microbiota composition. BMC Med. 2016, 14, 82. [Google Scholar] [CrossRef] [Green Version]
- McNulty, N.P.; Yatsunenko, T.; Hsiao, A.; Faith, J.J.; Muegge, B.D.; Goodman, A.L.; Henrissat, B.; Oozeer, R.; Cools-Portier, S.; Gobert, G.; et al. The impact of a consortium of fermented milk strains on the gut microbiome of gnotobiotic mice and monozygotic twins. Sci. Transl. Med. 2011, 3, 106ra106. [Google Scholar] [CrossRef] [Green Version]
- Eloe-Fadrosh, E.A.; Brady, A.; Crabtree, J.; Drabek, E.F.; Ma, B.; Mahurkar, A.; Ravel, J.; Haverkamp, M.; Fiorino, A.M.; Botelho, C.; et al. Functional dynamics of the gut microbiome in elderly people during probiotic consumption. mBio 2015, 6, e00231-15. [Google Scholar] [CrossRef] [Green Version]
- Tom Dieck, H.; Schön, C.; Wagner, T.; Pankoke, H.C.; Fluegel, M.; Speckmann, B. A synbiotic formulation comprising Bacillus subtilis DSM 32315 and L-alanyl-L-glutamine improves intestinal butyrate levels and lipid metabolism in healthy humans. Nutrients 2021, 14, 143. [Google Scholar] [CrossRef]
- Zoetendal, E.G.; Akkermans, A.D.L.; Akkermans-van Vliet, W.A.; de Visser, J.A.G.M.; de Vos, W.M. The Host Genotype Affects the Bacterial Community in the Human Gastronintestinal Tract. Microb. Ecol. Health Dis. 2001, 13, 129–134. [Google Scholar] [CrossRef] [Green Version]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Rachmi, C.N.; Li, M.; Alison Baur, L. Overweight and obesity in Indonesia: Prevalence and risk factors—A literature review. Public Health 2017, 147, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Herningtyas, E.H.; Ng, T.S. Prevalence and distribution of metabolic syndrome and its components among provinces and ethnic groups in Indonesia. BMC Public Health 2019, 19, 377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.; Zhang, M.; Xue, J.; Huang, J.; Zhuang, R.; Zhou, X.; Zhang, H.; Fu, Q.; Hao, Y. Body Mass Index Differences in the Gut Microbiota Are Gender Specific. Front. Microbiol. 2018, 9, 1250. [Google Scholar] [CrossRef]
- Biagi, E.; Candela, M.; Franceschi, C.; Brigidi, P. The aging gut microbiota: New perspectives. Ageing Res. Rev. 2011, 10, 428–429. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Sex | Age | Weight Control Period | Weight Difference Control | Taro Period | Weight Difference Taro | Taro + Probiotic Period | Weight Difference Taro + Probiotic | Taro + Beetroot Period | Weight Difference Taro + Beetroot | Total Weight Loss | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Da | F | 60 | 66 | 63.5 | −2.5 | 65 | 63 | −2 | 64 | 61 | −3 | 64 | 62 | −2 | −4 |
| Be | F | 47 | 80 | 75 | −5 | 76 | 72 | −4 | 72.5 | 69 | −3.5 | 71 | 69 | −2 | −11 |
| Er | F | 53 | 77 | 72 | −5 | 73 | 67 | −6 | 69 | 68 | −1 | 69.5 | 66 | −3.5 | −11 |
| Pa | M | 41 | 86 | 82 | −4 | 85 | 83 | −2 | 82.5 | 82 | −0.5 | 83 | 82 | −1 | −4 |
| We | F | 48 | 61 | 57 | −4 | 59 | 57 | −2 | 58 | 57 | −1 | 59 | 57.5 | −1.5 | −3.5 |
| Sa | F | 45 | 80 | 76 | −4 | 76 | 73 | −3 | 76 | 72 | −4 | 76 | 75 | −1 | −5 |
| Ju | M | 44 | 101 | 94 | −7 | 101 | 100 | −1 | 103.5 | 97.5 | −6 | 101 | 98 | −3 | −3 |
| Ro | F | 43 | 60 | 55 | −5 | 56.5 | 55.5 | −1 | 55.5 | 55 | −0.5 | 55 | 55.5 | 0.5 | −4.5 |
| An | F | 44 | 65 | 63 | −2 | 65 | 62 | −3 | 63.5 | 63 | −0.5 | 63 | 60 | −3 | −5 |
| De | F | 48 | 58 | 53 | −5 | 55 | 53 | −2 | 54 | 52 | −2 | 54 | 51 | −3 | −7 |
| Fr | F | 36 | 78 | 76 | −2 | 78 | 76 | −2 | 79 | 77 | −2 | 79 | 74 | −5 | −4 |
| Pr | M | 62 | 67 | 67 | 0 | 67 | 66.5 | −0.5 | 65 | 64 | −1 | 65 | 64 | −1 | −3 |
| Yu | M | 41 | 72.5 | 72.5 | 0 | 72.5 | 71.5 | −1 | 73 | 70.5 | −2.5 | 72 | 72 | 0 | −0.5 |
| Ro | M | 33 | 68 | 66 | −2 | 67 | 65.5 | −1.5 | 66 | 65 | −1 | 66 | 64 | −2 | −4 |
| Lu | M | 61 | 76.5 | 76.5 | 0 | 76.5 | 75.5 | −1 | 76.5 | 76 | −0.5 | 76 | 75.5 | −0.5 | −1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Surono, I.S.; Simatupang, A.; Kusumo, P.D.; Waspodo, P.; Verbruggen, S.; Verhoeven, J.; Venema, K. Effect of Different Functional Food Supplements on the Gut Microbiota of Prediabetic Indonesian Individuals during Weight Loss. Nutrients 2022, 14, 781. https://doi.org/10.3390/nu14040781
Surono IS, Simatupang A, Kusumo PD, Waspodo P, Verbruggen S, Verhoeven J, Venema K. Effect of Different Functional Food Supplements on the Gut Microbiota of Prediabetic Indonesian Individuals during Weight Loss. Nutrients. 2022; 14(4):781. https://doi.org/10.3390/nu14040781
Chicago/Turabian StyleSurono, Ingrid S., Abraham Simatupang, Pratiwi D. Kusumo, Priyo Waspodo, Sanne Verbruggen, Jessica Verhoeven, and Koen Venema. 2022. "Effect of Different Functional Food Supplements on the Gut Microbiota of Prediabetic Indonesian Individuals during Weight Loss" Nutrients 14, no. 4: 781. https://doi.org/10.3390/nu14040781
APA StyleSurono, I. S., Simatupang, A., Kusumo, P. D., Waspodo, P., Verbruggen, S., Verhoeven, J., & Venema, K. (2022). Effect of Different Functional Food Supplements on the Gut Microbiota of Prediabetic Indonesian Individuals during Weight Loss. Nutrients, 14(4), 781. https://doi.org/10.3390/nu14040781