
Rapid Nontranscriptional Effects of Calcifediol and Calcitriol

 , and
, and 
Abstract
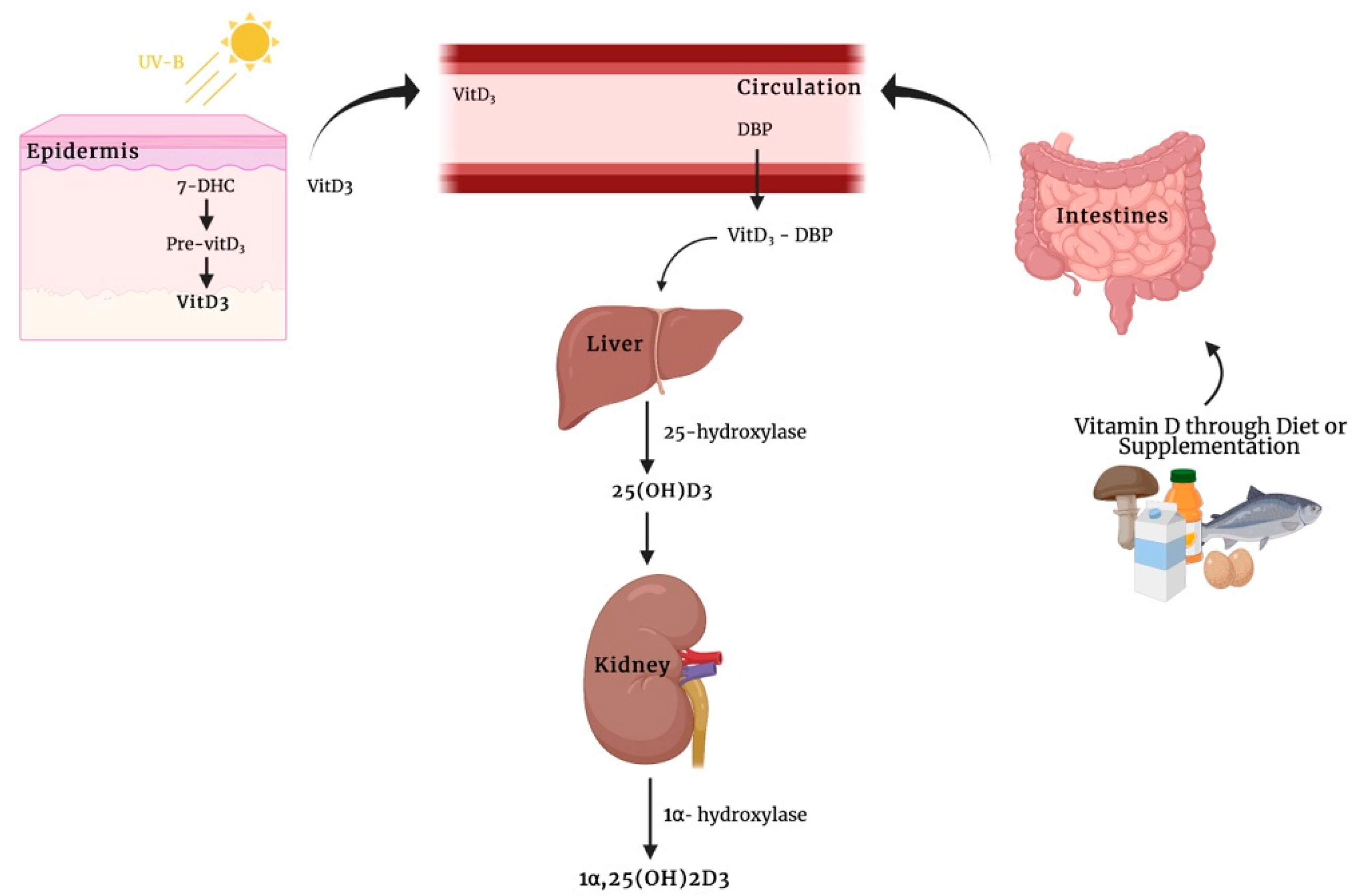
:1. Introduction
2. Rapid, Non-Genomic Steroid Actions
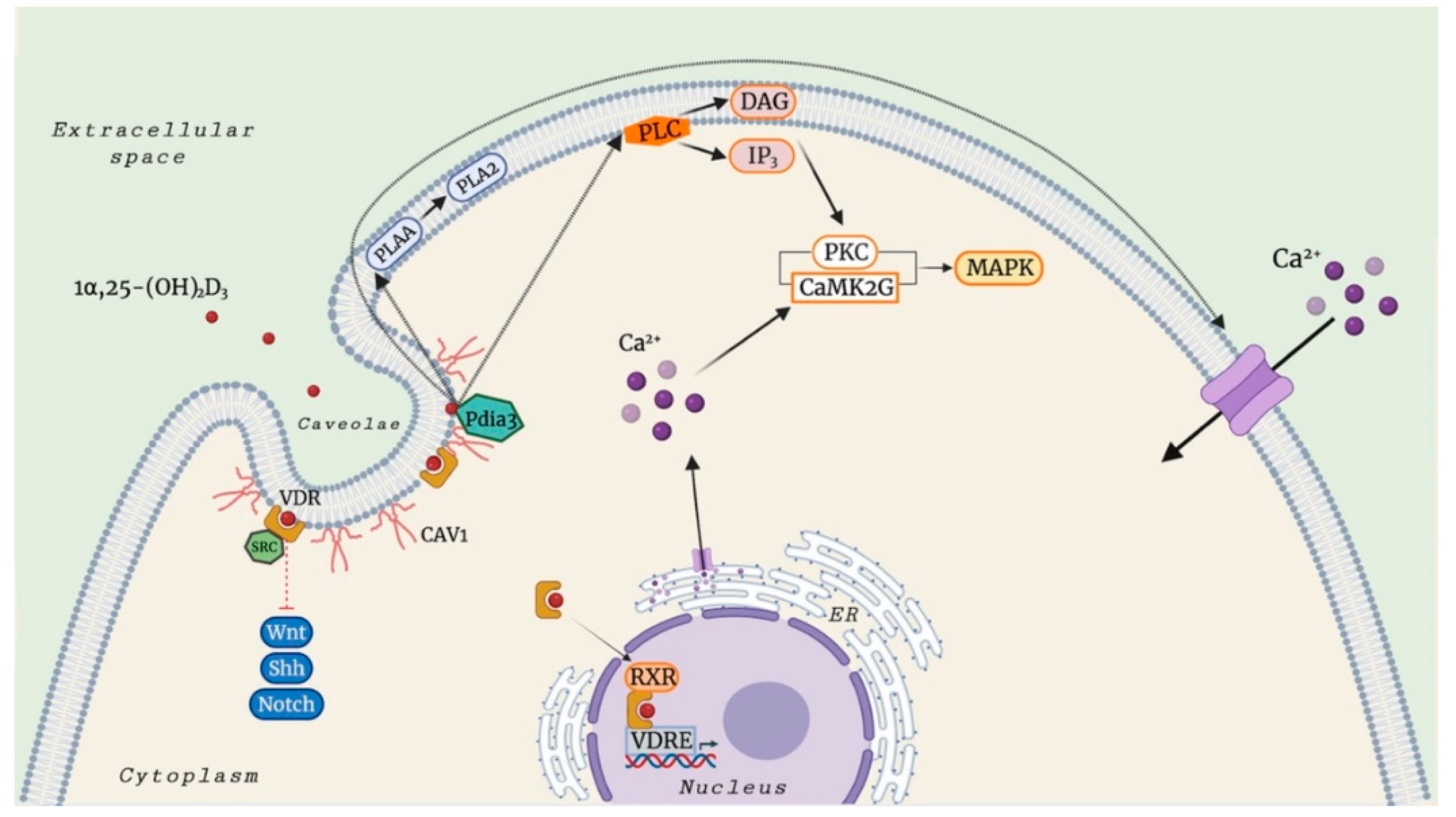
3. Mechanisms of Membrane-Associated Proteins for 1α,25(OH)2D3-Mediated Rapid, Non-Genomic Actions
4. 25(OH)D3-Mediated Rapid, Non-Genomic Actions
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gil, Á.; Plaza-Diaz, J.; Mesa, M.D. Vitamin D: Classic and Novel Actions. Ann. Nutr. Metab. 2018, 72, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Clinckspoor, I.; Verlinden, L.; Mathieu, C.; Bouillon, R.; Verstuyf, A.; Decallonne, B. Vitamin D in thyroid tumorigenesis and development. Prog. Histochem. Cytochem. 2013, 48, 65–98. [Google Scholar] [CrossRef] [PubMed]
- Naveh-Many, T.; Marx, R.; Keshet, E.; Pike, J.W.; Silver, J. Regulation of 1,25-dihydroxyvitamin D3 receptor gene expression by 1,25-dihydroxyvitamin D3 in the parathyroid in vivo. J. Clin. Investig. 1990, 86, 1968–1975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valero Zanuy, M.; Hawkins Carranza, F. Metabolismo, fuentes endógenas y exógenas de vitamina D. Rev. Esp. Enferm. Metab. Oseas 2007, 16, 63–70. [Google Scholar] [CrossRef]
- DeLuca, H.F. The Metabolism and Functions of Vitamin D. In Steroid Hormone Resistance: Mechanisms and Clinical Aspects; Chrousos, G.P., Loriaux, D.L., Lipsett, M.B., Eds.; Advances in Experimental Medicine and Biology; Springer: Boston, MA, USA, 1986; pp. 361–375. ISBN 978-1-4684-5101-6. [Google Scholar]
- Martin, C.E.; Veysey, M.; Yates, Z.R.; Lucock, M.D. Vitamin D: Genetics, Environment & Health. J. Food Nutr. Disord. 2014, 3, 1–19. [Google Scholar]
- Holick, M.F. Vitamin D: Importance in the prevention of cancers, type 1 diabetes, heart disease, and osteoporosis. Am. J. Clin. Nutr. 2004, 79, 362–371. [Google Scholar] [CrossRef] [Green Version]
- Zehnder, D.; Bland, R.; Williams, M.C.; McNinch, R.W.; Howie, A.J.; Stewart, P.M.; Hewison, M. Extrarenal expression of 25-hydroxyvitamin d(3)-1 alpha-hydroxylase. J. Clin. Endocrinol. Metab. 2001, 86, 888–894. [Google Scholar]
- DeLuca, H.F. Overview of general physiologic features and functions of vitamin D. Am. J. Clin. Nutr. 2004, 80, 1689S–1696S. [Google Scholar] [CrossRef] [Green Version]
- Feldman, D.; Malloy, P.J.; Gross, C. Chapter 9—Vitamin D: Biology, Action, and Clinical Implications. In Osteoporosis, 2nd ed.; Marcus, R., Feldman, D., Kelsey, J., Eds.; Academic Press: San Diego, LA, USA, 2001; pp. 257–303. ISBN 978-0-12-470862-4. [Google Scholar]
- Bouillon, R.; Carmeliet, G.; Verlinden, L.; van Etten, E.; Verstuyf, A.; Luderer, H.F.; Lieben, L.; Mathieu, C.; Demay, M. Vitamin D and human health: Lessons from vitamin D receptor null mice. Endocr. Rev. 2008, 29, 726–776. [Google Scholar] [CrossRef]
- DeLuca, H.F. Evolution of our understanding of vitamin D. Nutr. Rev. 2008, 66, S73–S87. [Google Scholar] [CrossRef]
- Schmidt, B.M.; Gerdes, D.; Feuring, M.; Falkenstein, E.; Christ, M.; Wehling, M. Rapid, nongenomic steroid actions: A new age? Front. Neuroendocrinol. 2000, 21, 57–94. [Google Scholar] [CrossRef] [PubMed]
- DeLuca, H.F.; Holick, M.F.; Schnoes, H.K.; Suda, T.; Cousins, R.J. Isolation and identification of 1,25-dihydroxycholecalciferol. A metabolite of vitamin D active in intestine. Biochemistry 1971, 10, 2799–2804. [Google Scholar] [CrossRef] [PubMed]
- Hii, C.S.; Ferrante, A. The Non-Genomic Actions of Vitamin D. Nutrients 2016, 8, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donati, S.; Palmini, G.; Romagnoli, C.; Aurilia, C.; Miglietta, F.; Falsetti, I.; Marini, F.; Zonefrati, R.; Galli, G.; Marcucci, G.; et al. In Vitro Non-Genomic Effects of Calcifediol on Human Preosteoblastic Cells. Nutrients 2021, 13, 4227. [Google Scholar] [CrossRef] [PubMed]
- Selye, H. Correlations between the chemical structure and the pharmacological actions of the steroids. Endocrinology 1942, 30, 437–453. [Google Scholar] [CrossRef]
- Spach, C.; Streeten, D.H.P. Retardation of Sodium Exchange in Dog Erythrocytes by Physiological Concentrations of Aldosterone, In Vitro. J. Clin. Investig. 1964, 43, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Lou, Y.-R.; Molnár, F.; Peräkylä, M.; Qiao, S.; Kalueff, A.V.; St-Arnaud, R.; Carlberg, C.; Tuohimaa, P. 25-Hydroxyvitamin D(3) is an agonistic vitamin D receptor ligand. J. Steroid Biochem. Mol. Biol. 2010, 118, 162–170. [Google Scholar] [CrossRef]
- Gerdes, D.; Christ, M.; K. Haseroth, K.; Notzon, A.; Falkenstein, E.; Wehling, M. Nongenomic Actions of Steroids-From the Laboratory to Clinical Implications. J. Pediat. Endocrinol. Metab. 2000, 13, 853–878. [Google Scholar] [CrossRef]
- Nemere, I.; Yoshimoto, Y.; Norman, A.W. Calcium transport in perfused duodena from normal chicks: Enhancement within fourteen minutes of exposure to 1,25-dihydroxyvitamin D3. Endocrinology 1984, 115, 1476–1483. [Google Scholar] [CrossRef]
- Fleet, J.C. Rapid, Membrane-Initiated Actions of 1,25 Dihydroxyvitamin D: What Are They and What Do They Mean? J. Nutr. 2004, 134, 3215–3218. [Google Scholar] [CrossRef]
- Doroudi, M.; Schwartz, Z.; Boyan, B.D. Membrane-mediated actions of 1,25-dihydroxy vitamin D3: A review of the roles of phospholipase A2 activating protein and Ca (2+)/calmodulin-dependent protein kinase II. J. Steroid Biochem. Mol. Biol. 2015, 147, 81–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dwivedi, P.P.; Hii, C.S.T.; Ferrante, A.; Tan, J.; Der, C.J.; Omdahl, J.L.; Morris, H.A.; May, B.K. Role of MAP kinases in the 1,25-dihydroxyvitamin D3-induced transactivation of the rat cytochrome P450C24 (CYP24) promoter. Specific functions for ERK1/ERK2 and ERK5. J. Biol. Chem. 2002, 277, 29643–29653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nutchey, B.K.; Kaplan, J.S.; Dwivedi, P.P.; Omdahl, J.L.; Ferrante, A.; May, B.K.; Hii, C.S.T. Molecular action of 1,25-dihydroxyvitamin D3 and phorbol ester on the activation of the rat cytochrome P450C24 (CYP24) promoter: Role of MAP kinase activities and identification of an important transcription factor binding site. Biochem. J. 2005, 389, 753–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dwivedi, P.; Gao, X.; Tan, J.; Evdokiou, A.; Ferrante, A.; Morris, H.; May, B.; Hii, C. A role for the phosphatidylinositol 3-kinase--protein kinase C zeta-Sp1 pathway in the 1,25-dihydroxyvitamin D3 induction of the 25-hydroxyvitamin D3 24-hydroxylase gene in human kidney cells. Cell. Signal. 2010, 22, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Norman, A.W. Vitamin D Receptor: New Assignments for an Already Busy Receptor. Endocrinology 2006, 147, 5542–5548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dormanen, M.C.; Bishop, J.E.; Hammond, M.W.; Okamura, W.H.; Nemere, I.; Norman, A.W. Nonnuclear effects of the steroid hormone 1 alpha,25(OH)2-vitamin D3: Analogs are able to functionally differentiate between nuclear and membrane receptors. Biochem. Biophys. Res. Commun. 1994, 201, 394–401. [Google Scholar] [CrossRef]
- Bikle, D.D.; Jiang, Y.; Nguyen, T.; Oda, Y.; Tu, C. Disruption of Vitamin D and Calcium Signaling in Keratinocytes Predisposes to Skin Cancer. Front. Physiol. 2016, 7, 296. [Google Scholar] [CrossRef] [Green Version]
- Bandera Merchan, B.; Morcillo, S.; Martin-Nuñez, G.; Tinahones, F.J.; Macías-González, M. The role of vitamin D and VDR in carcinogenesis: Through epidemiology and basic sciences. J. Steroid Biochem. Mol. Biol. 2017, 167, 203–218. [Google Scholar] [CrossRef]
- Hadden, M.K. Hedgehog and Vitamin D Signaling Pathways in Development and Disease. Vitam. Horm. 2016, 100, 231–253. [Google Scholar]
- Lisse, T.S.; Saini, V.; Zhao, H.; Luderer, H.F.; Gori, F.; Demay, M.B. The Vitamin D Receptor Is Required for Activation of cWnt and Hedgehog Signaling in Keratinocytes. Mol. Endocrinol. 2014, 28, 1698–1706. [Google Scholar] [CrossRef] [Green Version]
- Teichert, A.E.; Elalieh, H.; Elias, P.M.; Welsh, J.; Bikle, D.D. Overexpression of hedgehog signaling is associated with epidermal tumor formation in vitamin D receptor-null mice. J. Investig. Dermatol. 2011, 131, 2289–2297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teichert, A.; Elalieh, H.; Bikle, D. Disruption of the hedgehog signaling pathway contributes to the hair follicle cycling deficiency in Vdr knockout mice. J. Cell. Physiol. 2010, 225, 482–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapia, C.; Suares, A.; De Genaro, P.; González-Pardo, V. In vitro studies revealed a downregulation of Wnt/β-catenin cascade by active vitamin D and TX 527 analog in a Kaposi’s sarcoma cellular model. Toxicol. In Vitr. 2020, 63, 104748. [Google Scholar] [CrossRef] [PubMed]
- Muralidhar, S.; Filia, A.; Nsengimana, J.; Poźniak, J.; O’Shea, S.J.; Diaz, J.M.; Harland, M.; Randerson-Moor, J.A.; Reichrath, J.; Laye, J.P.; et al. Vitamin D–VDR Signaling Inhibits Wnt/β-Catenin–Mediated Melanoma Progression and Promotes Antitumor Immunity. Cancer Res. 2019, 79, 5986–5998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, L.; Fang, W.; Lin, J.; Li, J.; Wu, W.; Xu, J. Vitamin D protects human melanocytes against oxidative damage by activation of Wnt/β-catenin signaling. Lab. Investig. 2018, 98, 1527–1537. [Google Scholar] [CrossRef] [PubMed]
- Larriba, M.J.; González-Sancho, J.M.; Bonilla, F.; Muñoz, A. Interaction of vitamin D with membrane-based signaling pathways. Front. Physiol. 2014, 5, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Wang, X.; Xu, L.; Zhang, J.; Cao, H. A molecular sub-cluster of colon cancer cells with low VDR expression is sensitive to chemotherapy, BRAF inhibitors and PI3K-mTOR inhibitors treatment. Aging 2019, 11, 8587–8603. [Google Scholar] [CrossRef] [PubMed]
- Olsson, K.; Saini, A.; Strömberg, A.; Alam, S.; Lilja, M.; Rullman, E.; Gustafsson, T. Evidence for Vitamin D Receptor Expression and Direct Effects of 1α,25(OH)2D3 in Human Skeletal Muscle Precursor Cells. Endocrinology 2016, 157, 98–111. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, E.; Raghavan, S. Getting under the skin of epidermal morphogenesis. Nat. Rev. Genet. 2002, 3, 199–209. [Google Scholar] [CrossRef]
- Civitelli, R.; Kim, Y.S.; Gunsten, S.L.; Fujimori, A.; Huskey, M.; Avioli, L.V.; Hruska, K.A. Nongenomic Activation of the Calcium Message System by Vitamin D Metabolites in Osteoblast-like Cells. Endocrinology 1990, 127, 2253–2262. [Google Scholar] [CrossRef]
- Selles, J.; Boland, R. Evidence on the participation of the 3′,5′-cyclic AMP pathway in the non-genomic action of 1,25-dihydroxy-vitamin D3 in cardiac muscle. Mol. Cell. Endocrinol. 1991, 82, 229–235. [Google Scholar] [CrossRef]
- Baran, D.T.; Ray, R.; Sorensen, A.M.; Honeyman, T.; Holick, M.F. Binding characteristics of a membrane receptor that recognizes 1 alpha,25-dihydroxyvitamin D3 and its epimer, 1 beta,25-dihydroxyvitamin D3. J. Cell. Biochem. 1994, 56, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Boyan, B.D.; Sylvia, V.L.; McKinney, N.; Schwartz, Z. Membrane actions of vitamin D metabolites 1alpha,25(OH)2D3 and 24R,25(OH)2D3 are retained in growth plate cartilage cells from vitamin D receptor knockout mice. J. Cell. Biochem. 2003, 90, 1207–1223. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Ren, S.; Chen, H.; Chun, R.F.; Gacad, M.A.; Adams, J.S. Intracellular vitamin D binding proteins: Novel facilitators of vitamin D-directed transactivation. Mol. Endocrinol. 2000, 14, 1387–1397. [Google Scholar] [CrossRef] [PubMed]
- Chapron, B.D.; Chapron, A.; Phillips, B.; Okoli, M.C.; Shen, D.D.; Kelly, E.J.; Himmelfarb, J.; Thummel, K.E. Reevaluating the role of megalin in renal vitamin D homeostasis using a human cell-derived microphysiological system. ALTEX 2018, 35, 504–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nykjaer, A.; Fyfe, J.C.; Kozyraki, R.; Leheste, J.R.; Jacobsen, C.; Nielsen, M.S.; Verroust, P.J.; Aminoff, M.; de la Chapelle, A.; Moestrup, S.K.; et al. Cubilin dysfunction causes abnormal metabolism of the steroid hormone 25(OH) vitamin D (3). Proc. Natl. Acad. Sci. USA 2001, 98, 13895–13900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zmijewski, M.A.; Carlberg, C. Vitamin D receptor(s): In the nucleus but also at membranes? Exp. Dermatol. 2020, 29, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Nemere, I.; Dormanen, M.C.; Hammond, M.W.; Okamura, W.H.; Norman, A.W. Identification of a specific binding protein for 1 alpha,25-dihydroxyvitamin D3 in basal-lateral membranes of chick intestinal epithelium and relationship to transcaltachia. J. Biol. Chem. 1994, 269, 23750–23756. [Google Scholar] [CrossRef]
- Nemere, I.; Farach-Carson, M.C.; Rohe, B.; Sterling, T.M.; Norman, A.W.; Boyan, B.D.; Safford, S.E. Ribozyme knockdown functionally links a 1,25(OH)2D3 membrane binding protein (1,25D3-MARRS) and phosphate uptake in intestinal cells. Proc. Natl. Acad. Sci. USA 2004, 101, 7392–7397. [Google Scholar] [CrossRef] [Green Version]
- Nemere, I.; Safford, S.E.; Rohe, B.; DeSouza, M.M.; Farach-Carson, M.C. Identification and characterization of 1,25D3-membrane-associated rapid response, steroid (1,25D3-MARRS) binding protein. J. Steroid Biochem. Mol. Biol. 2004, 89–90, 281–285. [Google Scholar] [CrossRef]
- Hettinghouse, A.; Liu, R.; Liu, C.-J. Multifunctional molecule ERp57: From cancer to neurodegenerative diseases. Pharmacol. Ther. 2018, 181, 34–48. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, F.; Xu, R.; Awan, M.U.N.; Song, Y.; Han, Q.; Xia, X.; Zhang, J. PDIA3: Structure, functions and its potential role in viral infections. Biomed. Pharmacother. 2021, 143, 112110. [Google Scholar] [CrossRef] [PubMed]
- Doroudi, M.; Chen, J.; Boyan, B.D.; Schwartz, Z. New insights on membrane mediated effects of 1α,25-dihydroxy vitamin D3 signaling in the musculoskeletal system. Steroids 2014, 81, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Doroudi, M.; Schwartz, Z.; Boyan, B.D. Phospholipase A2 activating protein is required for 1α,25-dihydroxyvitamin D3 dependent rapid activation of protein kinase C via Pdia3. J. Steroid Biochem. Mol. Biol. 2012, 132, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Sequeira, V.B.; Rybchyn, M.S.; Tongkao-On, W.; Gordon-Thomson, C.; Malloy, P.J.; Nemere, I.; Norman, A.W.; Reeve, V.E.; Halliday, G.M.; Feldman, D.; et al. The role of the vitamin D receptor and ERp57 in photoprotection by 1α,25-dihydroxyvitamin D3. Mol. Endocrinol. 2012, 26, 574–582. [Google Scholar] [CrossRef] [Green Version]
- Khanal, R.; Nemere, I. Membrane receptors for vitamin D metabolites. Crit. Rev. Eukaryot. Gene Expr. 2007, 17, 31–47. [Google Scholar] [CrossRef]
- Yang, W.S.; Yu, H.; Kim, J.J.; Lee, M.J.; Park, S.-K. Vitamin D-induced ectodomain shedding of TNF receptor 1 as a nongenomic action: D3 vs D2 derivatives. J. Steroid Biochem. Mol. Biol. 2016, 155, 18–25. [Google Scholar] [CrossRef]
- Doroudi, M.; Olivares-Navarrete, R.; Boyan, B.D.; Schwartz, Z. A review of 1α,25(OH)2D3 dependent Pdia3 receptor complex components in Wnt5a non-canonical pathway signaling. J. Steroid Biochem. Mol. Biol. 2015, 152, 84–88. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, J.; Lee, C.S.D.; Nizkorodov, A.; Riemenschneider, K.; Martin, D.; Hyzy, S.; Schwartz, Z.; Boyan, B.D. Disruption of Pdia3 gene results in bone abnormality and affects 1alpha,25-dihydroxy-vitamin D3-induced rapid activation of PKC. J. Steroid Biochem. Mol. Biol. 2010, 121, 257–260. [Google Scholar] [CrossRef]
- Garbi, N.; Tanaka, S.; Momburg, F.; Hämmerling, G.J. Impaired assembly of the major histocompatibility complex class I peptide-loading complex in mice deficient in the oxidoreductase ERp57. Nat. Immunol. 2006, 7, 93–102. [Google Scholar] [CrossRef]
- Nemere, I.; Garbi, N.; Hämmerling, G.J.; Khanal, R.C. Intestinal cell calcium uptake and the targeted knockout of the 1,25D3-MARRS (membrane-associated, rapid response steroid-binding) receptor/PDIA3/Erp57. J. Biol. Chem. 2010, 285, 31859–31866. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Lobachev, K.S.; Grindel, B.J.; Farach-Carson, M.C.; Hyzy, S.L.; El-Baradie, K.B.; Olivares-Navarrete, R.; Doroudi, M.; Boyan, B.D.; Schwartz, Z. Chaperone properties of pdia3 participate in rapid membrane actions of 1α,25-dihydroxyvitamin d3. Mol. Endocrinol. 2013, 27, 1065–1077. [Google Scholar] [CrossRef] [Green Version]
- Gaucci, E.; Raimondo, D.; Grillo, C.; Cervoni, L.; Altieri, F.; Nittari, G.; Eufemi, M.; Chichiarelli, S. Analysis of the interaction of calcitriol with the disulfide isomerase ERp57. Sci. Rep. 2016, 6, 37957. [Google Scholar] [CrossRef] [Green Version]
- Blomberg Jensen, M.; Dissing, S. Non-genomic effects of vitamin D in human spermatozoa. Steroids 2012, 77, 903–909. [Google Scholar] [CrossRef]
- Blomberg Jensen, M.; Bjerrum, P.J.; Jessen, T.E.; Nielsen, J.E.; Joensen, U.N.; Olesen, I.A.; Petersen, J.H.; Juul, A.; Dissing, S.; Jørgensen, N. Vitamin D Is Positively Associated with Sperm Motility and Increases Intracellular Calcium in Human Spermatozoa. Hum. Reprod. 2011, 26, 1307–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asano, L.; Watanabe, M.; Ryoden, Y.; Usuda, K.; Yamaguchi, T.; Khambu, B.; Takashima, M.; Sato, S.-I.; Sakai, J.; Nagasawa, K.; et al. Vitamin D Metabolite, 25-Hydroxyvitamin D, Regulates Lipid Metabolism by Inducing Degradation of SREBP/SCAP. Cell Chem. Biol. 2017, 24, 207–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lösel, R.; Wehling, M. Nongenomic actions of steroid hormones. Nat. Rev. Mol. Cell Biol. 2003, 4, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, G.; de Boland, A.R.; Boland, R. Stimulation of Ca2+ release-activated Ca2+ channels as a potential mechanism involved in non-genomic 1,25(OH)2-vitamin D3-induced Ca2+ entry in skeletal muscle cells. Biochem. Biophys. Res. Commun. 1997, 239, 562–565. [Google Scholar] [CrossRef]
- Haussler, M.R.; Jurutka, P.W.; Mizwicki, M.; Norman, A.W. Vitamin D receptor (VDR)-mediated actions of 1α,25(OH)₂vitamin D₃: Genomic and non-genomic mechanisms. Best Pract. Res. Clin. Endocrinol. Metab. 2011, 25, 543–559. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Study | 1α,25(OH)2D3-Mediated Rapid, Non-Genomic Effects | Putative Membrane-Associated Protein Responsible for 1α,25(OH)2D3-Related Rapid, Non-Genomic Effects |
|---|---|---|
| Nemere et al. [21] | Transcaltachia | / 1 |
| Dormanen et al. [27,28] | Transcaltachia | VDR |
| Lisse et al. [32] and Teichert et al. [33,34] | Regulation of the Hedeghog signalling pathway | VDR |
| Tapia et al. [35], Muralidhar et al. [36], and Tang et al. [37] | Regulation of the Wnt signalling pathway | VDR |
| Wang et al. [39] and Olsson et al. [40] | Regulation of the Notch signalling pathway | VDR |
| Civitelli et al. [42] | Increase in intracellular Ca2+ levels | / 1 |
| Selles et al. [43] | Involvement in cAMP signalling pathway | / 1 |
| Nemere et al. [50] | Transcaltachia | Pdia3 |
| Doroudi et al. [56] | Increase of intracellular Ca2+ levels | Pdia3 |
| Sequeira et al. [57] | Protection of UV-induced thymine dimer formation | Pdia3 |
| Khanal et. al. [58] | Regulation of PKC signalling pathway | Pdia3 |
| Yang et al. [59] | Regulation of TNF signalling pathway | Pdia3 |
| Zmijewski et al. [49] | Regulation of PLA2 activation | Pdia3 |
| Doroudi et al. [55] | Regulation of MAPK1 and MAPK2 activation | Pdia3 |
| Doroudi et al. [60] | Regulation of Wnt5A non-canonical signalling pathway | Pdia3 |
| Nemere et al. [63] | Regulation of PKA signalling pathway | Pdia3 |
| Wang et al. [61] and Boyan et al. [45] | Regulation of PKC activity | Pdia3 |
| Study | 25(OH)D3-Mediated Rapid, Non-Genomic Effects | Putative Membrane-Associated Protein Responsible for 25(OH)D3-Related Rapid, Non-Genomic Effects |
|---|---|---|
| Donati et al. [16] | Increase of intracellular Ca2+ levels | / 1 |
| Jensen et al. [67] | Increase of intracellular Ca2+ levels | VDR |
| Asano et al. [68] | Regulation of lipogenesis | SCAP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donati, S.; Palmini, G.; Aurilia, C.; Falsetti, I.; Miglietta, F.; Iantomasi, T.; Brandi, M.L. Rapid Nontranscriptional Effects of Calcifediol and Calcitriol. Nutrients 2022, 14, 1291. https://doi.org/10.3390/nu14061291
Donati S, Palmini G, Aurilia C, Falsetti I, Miglietta F, Iantomasi T, Brandi ML. Rapid Nontranscriptional Effects of Calcifediol and Calcitriol. Nutrients. 2022; 14(6):1291. https://doi.org/10.3390/nu14061291
Chicago/Turabian StyleDonati, Simone, Gaia Palmini, Cinzia Aurilia, Irene Falsetti, Francesca Miglietta, Teresa Iantomasi, and Maria Luisa Brandi. 2022. "Rapid Nontranscriptional Effects of Calcifediol and Calcitriol" Nutrients 14, no. 6: 1291. https://doi.org/10.3390/nu14061291
APA StyleDonati, S., Palmini, G., Aurilia, C., Falsetti, I., Miglietta, F., Iantomasi, T., & Brandi, M. L. (2022). Rapid Nontranscriptional Effects of Calcifediol and Calcitriol. Nutrients, 14(6), 1291. https://doi.org/10.3390/nu14061291