
In Vitro Gut Fermentation of Whey Protein Hydrolysate: An Evaluation of Its Potential Modulation on Infant Gut Microbiome

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Determination of Amino Acids and Peptides in WPH
2.3. In Vitro Anaerobic Culture
2.4. Simulated Gastrointestinal Digestion of WPH
2.5. Determination of Molecular Weight Distribution
2.6. Infant Fecal Inoculum
2.7. Determination of pH Value and SCFAs Content during In Vitro Fermentation
2.8. Microbiome Analysis
2.9. Statistical Analysis
3. Results
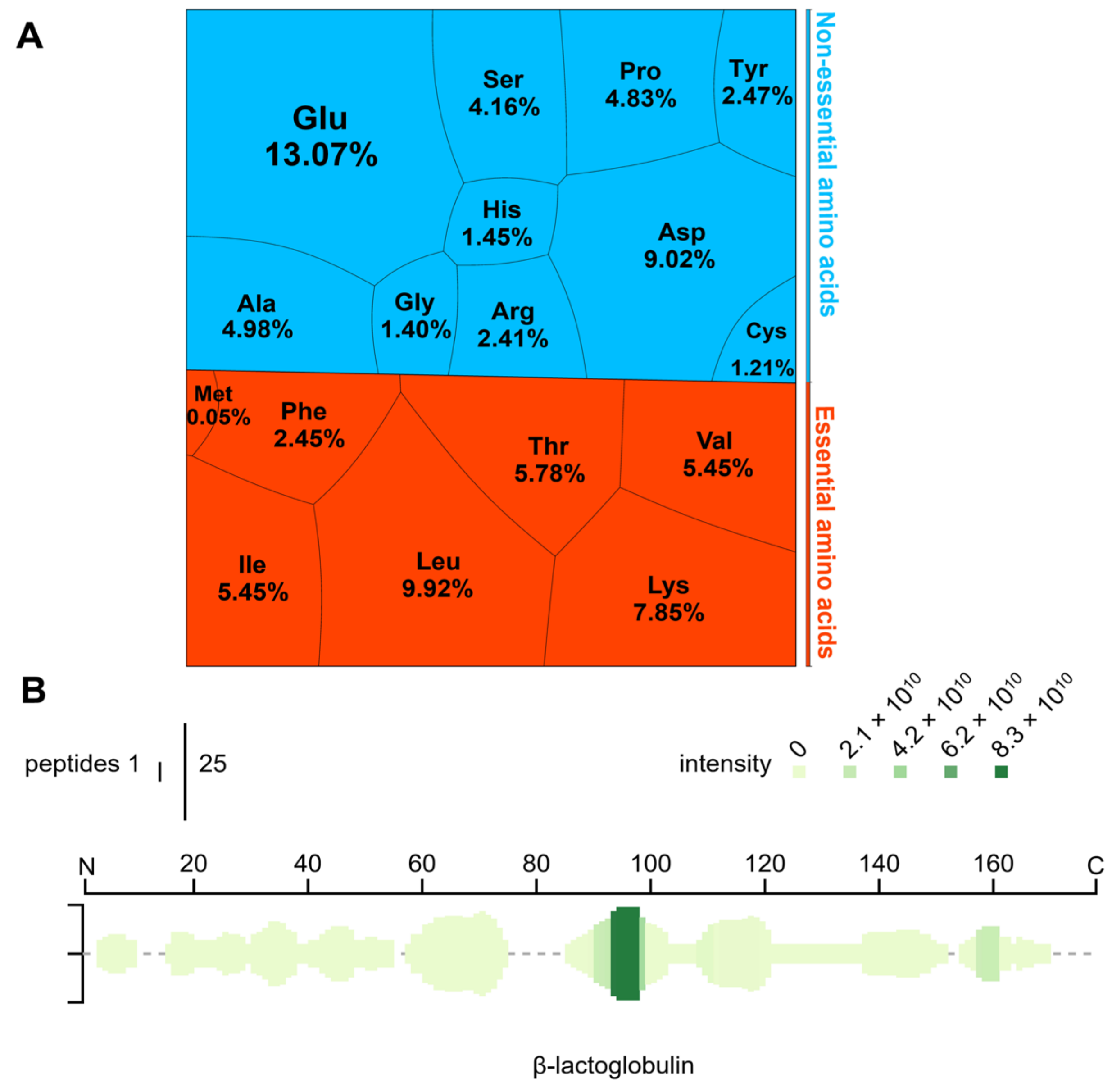
3.1. Amino Acid Composition and Peptide Profiling
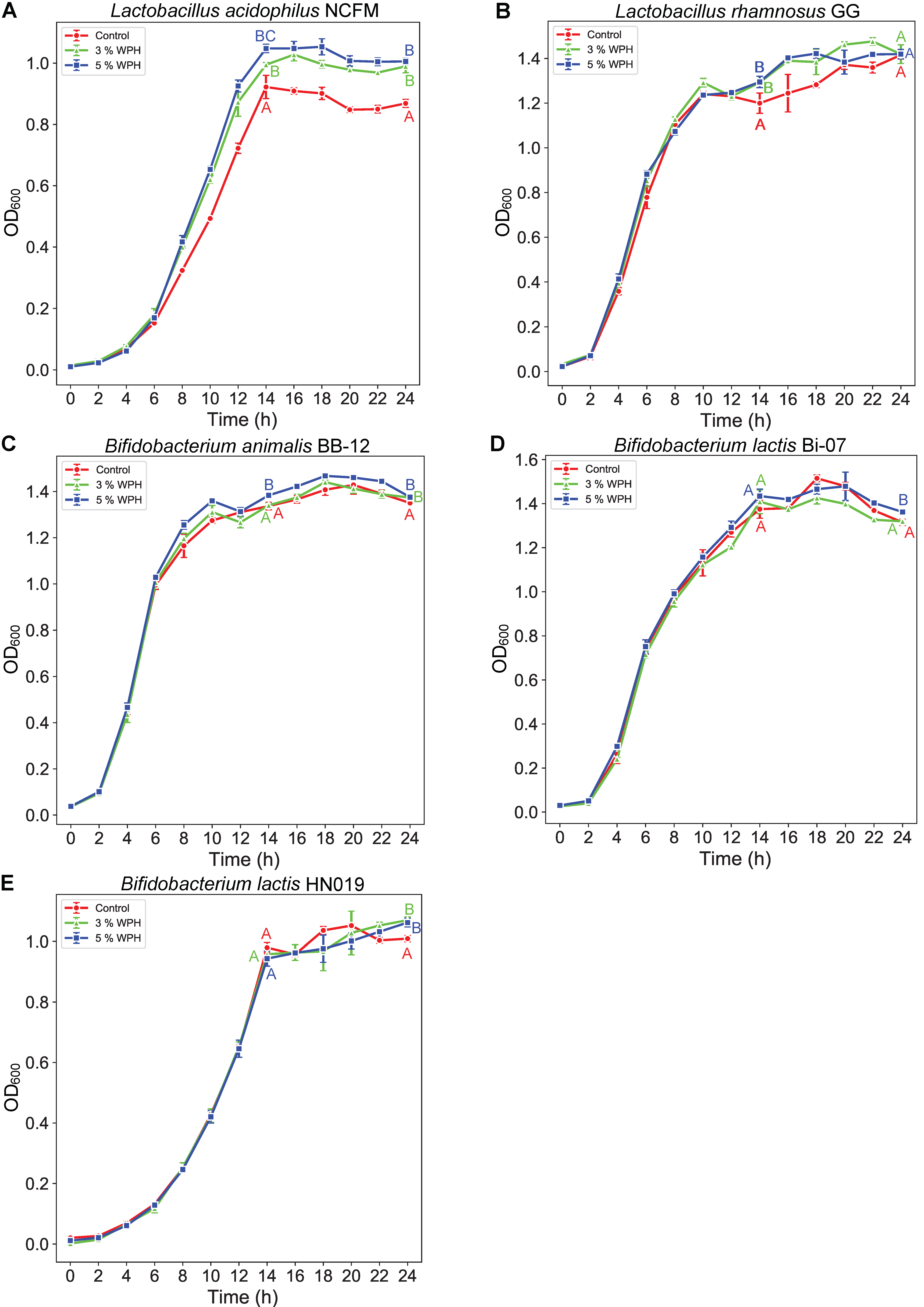
3.2. Growth of Lactobacilli and Bifidobacteria with WPH as Selectable Nitrogen Source
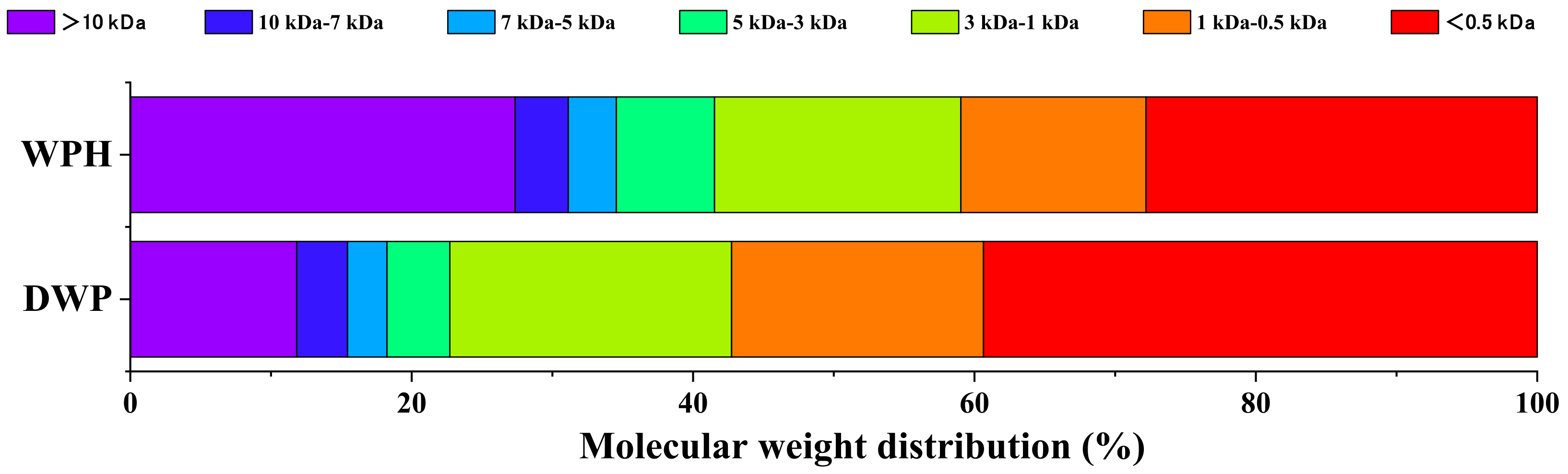
3.3. Changes in Molecular Weight Distribution after Digestion
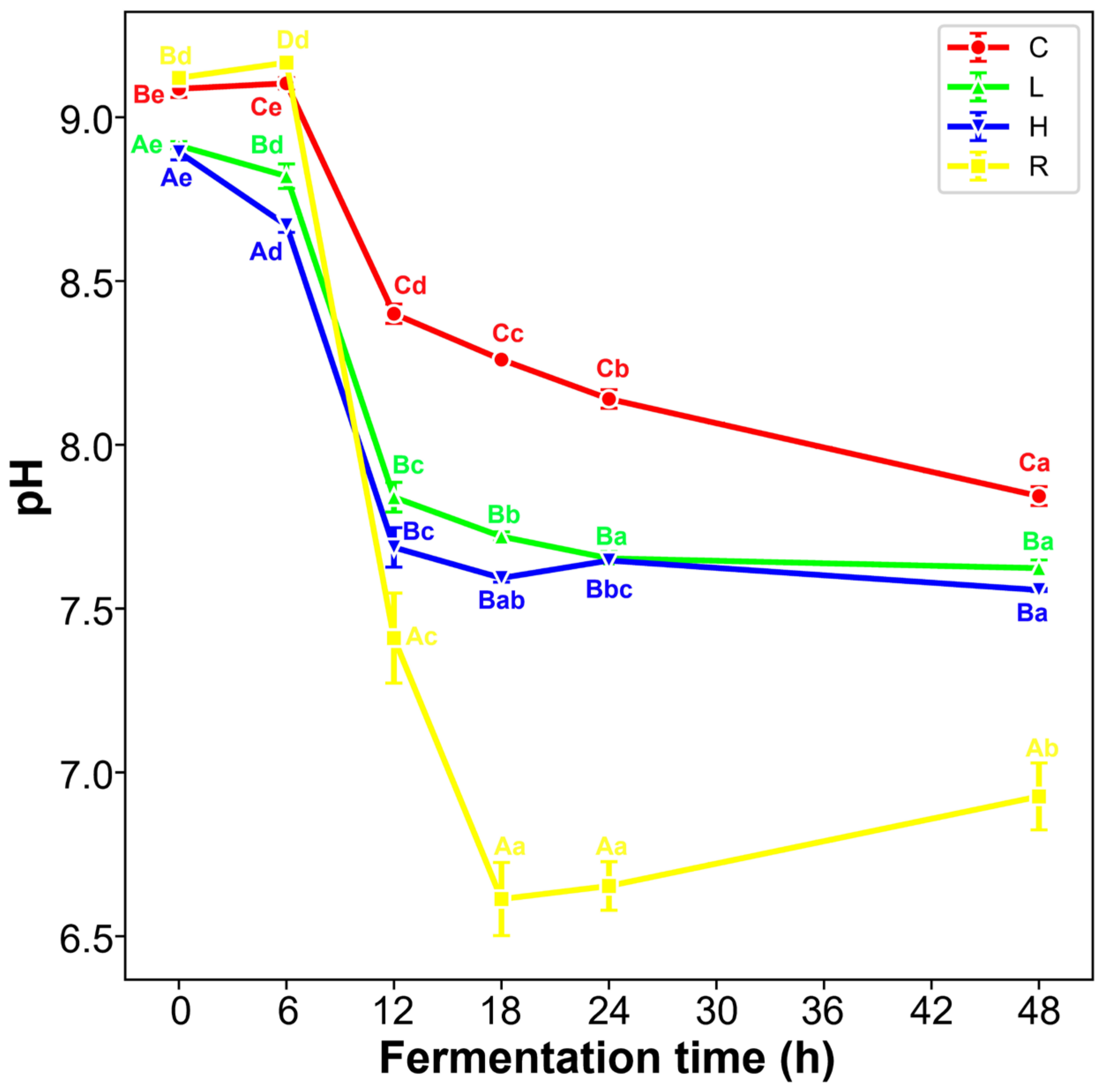
3.4. The pH Curve of Fermented Broth
3.5. Changes in SCFAs Formation during Fermentation
3.6. Effects of DWP on Fecal Microbiota
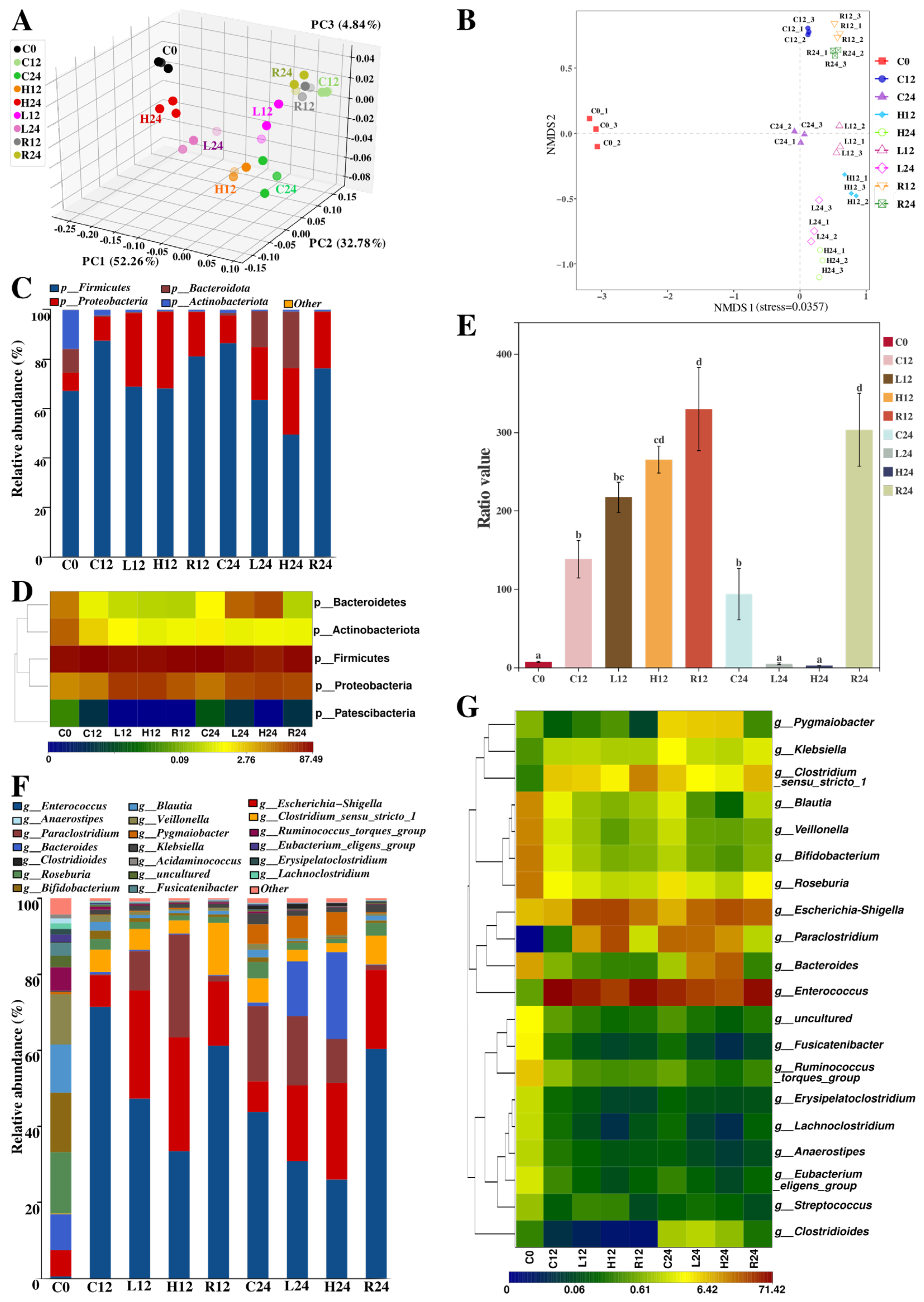
3.6.1. Alpha and Beta Diversity Analysis
3.6.2. Taxonomic Analysis
3.7. Correlation Analysis of SCFAs with Fecal Microbiota at the Genus Level
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Patel, S. Functional food relevance of whey protein: A review of recent findings and scopes ahead. J. Funct. Food 2015, 19, 308–319. [Google Scholar] [CrossRef]
- Fenelon, M.A.; Hickey, R.M.; Buggy, A.; McCarthy, N.; Murphy, E.G. Chapter 12—Whey Proteins in Infant Formula. In Whey Proteins; Deeth, H.C., Bansal, N., Eds.; Academic Press: London, UK, 2019; pp. 439–494. [Google Scholar]
- Dullius, A.; Goettert, M.I.; de Souza, C.F.V. Whey Protein Hydrolysates as a Source of Bioactive Peptides for Functional Foods—Biotechnological Facilitation of Industrial Scale-Up. J. Funct. Foods 2018, 42, 58–74. [Google Scholar] [CrossRef]
- Chatterton, D.E.W.; Smithers, G.; Roupas, P.; Brodkorb, A. Bioactivity of β-lactoglobulin and α-lactalbumin—Technological implications for processing. Int. Dairy J. 2006, 16, 1229–1240. [Google Scholar] [CrossRef]
- Wu, S.-L.; Ding, D.; Fang, A.-P.; Chen, P.-Y.; Chen, S.; Jing, L.-P.; Chen, Y.-M.; Zhu, H.-L. Growth, Gastrointestinal Tolerance and Stool Characteristics of Healthy Term Infants Fed an Infant Formula Containing Hydrolyzed Whey Protein (63%) and Intact Casein (37%): A Randomized Clinical Trial. Nutrients 2017, 9, 1254. [Google Scholar] [CrossRef] [Green Version]
- Picaud, J.-C.; Pajek, B.; Arciszewska, M.; Tarczón, I.; Escribano, J.; Porcel, R.; Adelt, T.; Hassink, E.; Rijnierse, A.; Abrahamse-Berkeveld, M.; et al. An Infant Formula with Partially Hydrolyzed Whey Protein Supports Adequate Growth and Is Safe and Well-Tolerated in Healthy, Term Infants: A Randomized, Double-Blind, Equivalence Trial. Nutrients 2020, 12, 2072. [Google Scholar] [CrossRef]
- Yu, Y.-J.; Amorim, M.; Marques, C.; Calhau, C.; Pintado, M. Effects of Whey Peptide Extract on the Growth of Probiotics and Gut Microbiota. J. Funct. Foods 2016, 21, 507–516. [Google Scholar] [CrossRef]
- Boscaini, S.; Cabrera-Rubio, R.; Nychyk, O.; Speakman, J.R.; Cryan, J.F.; Cotter, P.D.; Nilaweera, K.N. Age- and duration-dependent effects of whey protein on high-fat diet-induced changes in body weight, lipid metabolism, and gut microbiota in mice. Physiol. Rep. 2020, 8, e14523. [Google Scholar] [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids from Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [Green Version]
- Mueller, N.T.; Bakacs, E.; Combellick, J.; Grigoryan, Z.; Dominguez-Bello, M.G. The infant microbiome development: Mom matters. Trends Mol. Med. 2015, 21, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Markowiak-Kopeć, P.; Śliżewska, K. The Effect of Probiotics on the Production of Short-Chain Fatty Acids by Human Intestinal Microbiome. Nutrients 2020, 12, 1107. [Google Scholar] [CrossRef]
- Peled, S.; Livney, Y.D. The role of dietary proteins and carbohydrates in gut microbiome composition and activity: A review. Food Hydrocoll. 2021, 120, 106911. [Google Scholar] [CrossRef]
- Liu, X.-R.; Zhu, N.; Hao, Y.-T.; Yu, X.-C.; Li, Z.; Mao, R.-X.; Liu, R.; Kang, J.-W.; Hu, J.-N.; Li, Y. Radioprotective Effect of Whey Hydrolysate Peptides against γ-Radiation-Induced Oxidative Stress in BALB/c Mice. Nutrients 2021, 13, 816. [Google Scholar] [CrossRef]
- Liang, C.; Wang, L.; Zhang, Y.; Zhang, J.; Zhang, X.; Ye, N. The effects of elevated CO2 concentrations on changes in fatty acids and amino acids of three species of microalgae. Phycologia 2020, 59, 208–217. [Google Scholar] [CrossRef]
- Long, S.; Yang, Y.; Shen, C.; Wang, Y.; Deng, A.; Qin, Q.; Qiao, L. Metaproteomics characterizes human gut microbiome function in colorectal cancer. NPJ Biofilm. Microbiomes 2020, 6, 14. [Google Scholar] [CrossRef]
- Manguy, J.; Jehl, P.; Dillon, E.T.; Davey, N.E.; Shields, D.C.; Holton, T.A. Peptigram: A Web-Based Application for Peptidomics Data Visualization. J. Proteome Res. 2017, 16, 712–719. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Huang, R.; Shah, N.P.; Tao, X.; Xiong, Y.; Wei, H. Antioxidant and antibacterial activities of exopolysaccharides from Bifidobacterium bifidum WBIN03 and Lactobacillus plantarum R315. J. Dairy Sci. 2014, 97, 7334–7343. [Google Scholar] [CrossRef] [Green Version]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Wang, K.; Luo, Q.; Hong, H.; Liu, H.; Luo, Y. Novel antioxidant and ACE inhibitory peptide identified from Arthrospira platensis protein and stability against thermal/pH treatments and simulated gastrointestinal digestion. Food Res. Int. 2021, 139, 109908. [Google Scholar] [CrossRef]
- Chen, Y.; Chang, S.K.C.; Zhang, Y.; Hsu, C.-Y.; Nannapaneni, R. Gut microbiota and short chain fatty acid composition as affected by legume type and processing methods as assessed by simulated in vitro digestion assays. Food Chem. 2020, 312, 126040. [Google Scholar] [CrossRef]
- Yun, C.; Liu, E.; Rippa, M.; Mormile, P.; Sun, D.; Yan, C.; Liu, Q. Effects of Chemical and Solar Soil-Disinfection Methods on Soil Bacterial Communities. Sustainability 2020, 12, 9833. [Google Scholar] [CrossRef]
- Sousa, R.; Portmann, R.; Dubois, S.; Recio, I.; Egger, L. Protein digestion of different protein sources using the INFOGEST static digestion model. Food Res. Int. 2020, 130, 108996. [Google Scholar] [CrossRef]
- Hoppe, C.; Andersen, G.S.; Jacobsen, S.; Mølgaard, C.; Friis, H.; Sangild, P.T.; Michaelsen, K.F. The Use of Whey or Skimmed Milk Powder in Fortified Blended Foods for Vulnerable Groups. J. Nutr. 2008, 138, 145S–161S. [Google Scholar] [CrossRef] [Green Version]
- McGregor, R.A.; Poppitt, S.D. Milk protein for improved metabolic health: A review of the evidence. Nutr. Metab. 2013, 10, 46. [Google Scholar] [CrossRef] [Green Version]
- Mølgaard, C.; Larnkjær, A.; Arnberg, K.; Michaelsen, K.F. Milk and Growth in Children: Effects of Whey and Casein. In Nestlé Nutrition Institute Workshop Series: Pediatric Program; Clemens, R.A., Hernell, O., Michaelsen, K.F., Eds.; Nestec Ltd.: Vevey, Switzerland, 2011; Volume 67, pp. 67–78. [Google Scholar]
- Mei, F.; Duan, Z.; Chen, M.; Lu, J.; Zhao, M.; Li, L.; Shen, X.; Xia, G.; Chen, S. Effect of a high-collagen peptide diet on the gut microbiota and short-chain fatty acid metabolism. J. Funct. Food 2020, 75, 104278. [Google Scholar] [CrossRef]
- Diether, N.E.; Willing, B.P. Microbial Fermentation of Dietary Protein: An Important Factor in Diet–Microbe–Host Interaction. Microorganisms 2019, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Lactobacillus Acidophilus NCFM®|Database. Available online: https://www.optibacprobiotics.com/professionals/probiotics-database/lactobacillus/lactobacillus-acidophilus/lactobacillus-acidophilus-ncfm (accessed on 28 November 2021).
- Coscia, A.; Bardanzellu, F.; Caboni, E.; Fanos, V.; Peroni, D.G. When a Neonate Is Born, So Is a Microbiota. Life 2021, 11, 148. [Google Scholar] [CrossRef]
- Stewart, C.J. Breastfeeding Promotes Bifidobacterial Immunomodulatory Metabolites. Nat. Microbiol. 2021, 6, 1335–1336. [Google Scholar] [CrossRef]
- Skolnick, J.; Chou, C.; Miklavcic, J. Insights into Novel Infant Milk Formula Bioactives. Nutr. Diet. Suppl. 2020, 12, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Moya, T.; López-Nicolás, R.; Planes, D.; González-Bermúdez, C.A.; Ros-Berruezo, G.; Frontela-Saseta, C. In vitro modulation of gut microbiota by whey protein to preserve intestinal health. Food Funct. 2017, 8, 3053–3063. [Google Scholar] [CrossRef]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- Davila, A.-M.; Blachier, F.; Gotteland, M.; Andriamihaja, M.; Benetti, P.-H.; Sanz, Y.; Tomé, D. Re-Print of “Intestinal luminal nitrogen metabolism: Role of the gut microbiota and consequences for the host”. Pharmacol. Res. 2013, 69, 114–126. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Neis, E.P.J.G.; Dejong, C.H.C.; Rensen, S.S. The Role of Microbial Amino Acid Metabolism in Host Metabolism. Nutrients 2015, 7, 2930–2946. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.; Zhai, Q.; Chen, W. Investigations of Bacteroides spp. towards next-generation probiotics. Food Res. Int. 2019, 116, 637–644. [Google Scholar] [CrossRef]
- Xiao, L.; Wang, J.; Zheng, J.; Li, X.; Zhao, F. Deterministic transition of enterotypes shapes the infant gut microbiome at an early age. Genome Biol. 2021, 22, 243. [Google Scholar] [CrossRef]
- Liu, L.; He, Y.; Wang, K.; Miao, J.; Zheng, Z. Metagenomics approach to the intestinal microbiome structure and function in high fat diet-induced obesity in mice fed with conjugated linoleic acid (CLA). Food Funct. 2020, 11, 9729–9739. [Google Scholar] [CrossRef]
- Li, T.; Gao, J.; Du, M.; Mao, X. Bovine α-lactalbumin hydrolysates ameliorate obesity-associated endotoxemia and inflammation in high-fat diet-fed mice through modulation of gut microbiota. Food Funct. 2019, 10, 3368–3378. [Google Scholar] [CrossRef]
- Moreno-Pérez, D.; Bressa, C.; Bailén, M.; Hamed-Bousdar, S.; Naclerio, F.; Carmona, M.; Pérez, M.; González-Soltero, R.; Montalvo-Lominchar, M.; Carabaña, C.; et al. Effect of a Protein Supplement on the Gut Microbiota of Endurance Athletes: A Randomized, Controlled, Double-Blind Pilot Study. Nutrients 2018, 10, 337. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SCFA | Sample | 0 h | 6 h | 12 h | 18 h | 24 h | 48 h |
|---|---|---|---|---|---|---|---|
| Acetic acid | C | 96.04 ± 5.83 A,a | 110.35 ± 3.34 A,a | 354.65 ± 70.06 A,b | 480.95 ± 9.26 A,c | 675.90 ± 68.17 A,d | 704.89 ± 56.67 A,d |
| L | 101.52 ± 9.92 A,a | 129.76 ± 5.15 B,a | 467.93 ± 6.30 B,b | 679.48 ± 19.82 B,c | 966.73 ± 72.05 B,d | 1068.40 ± 91.21 C,d | |
| H | 98.25 ± 9.74 A,a | 154.25 ± 7.88 C,a | 476.35 ± 1.94 B,b | 690.45 ± 17.61 B,c | 1029.74 ± 44.60 B,d | 978.99 ± 161.96 BC,d | |
| R | 96.05 ± 6.09 A,a | 111.97 ± 11.91 AB,a | 450.27 ± 58.52 AB,b | 599.20 ± 99.70 AB,c | 592.45 ± 66.52 A,c | 810.98 ± 51.61 AB,d | |
| Propionic acid | C | 7.97 ± 0.66 B,a | 5.62 ± 0.50 A,a | 17.06 ± 2.96 AB,a | 48.09 ± 4.70 B,b | 91.01 ± 19.42 B,c | 126.48 ± 4.92 B,d |
| L | 6.91 ± 1.16 AB,a | 7.61 ± 1.07 B,a | 35.89 ± 2.72 BC,b | 89.10 ± 2.10 C,c | 144.74 ± 7.18 C,d | 189.79 ± 3.30 C,e | |
| H | 5.91 ± 0.82 AB,a | 8.03 ± 0.15 B,a | 63.96 ± 24.10 C,b | 100.27 ± 2.73 D,c | 162.49 ± 0.99 C,d | 171.81 ± 29.20 C,d | |
| R | 4.63 ± 1.53 A,a | 6.25 ± 1.07 AB,a | 6.40 ± 2.16 A,a | 10.56 ± 3.22 A,a | 12.74 ± 3.69 A,a | 43.61 ± 2.84 A,a | |
| Butyric acid | C | 4.20 ± 0.99 B,a | 2.77 ± 0.10 A,a | 3.96 ± 1.10 A,a | 6.94 ± 0.31 A,a | 39.36 ± 3.21 B,b | 180.29 ± 4.62 B,c |
| L | 3.15 ± 0.52 AB,a | 4.51 ± 1.14 B,a | 5.47 ± 0.15 A,a | 41.08 ± 4.21 B,b | 101.31 ± 3.81 C,c | 201.53 ± 10.30 BC,d | |
| H | 2.73 ± 0.35 AB,a | 3.66 ± 0.11 AB,a | 8.83 ± 4.31 A,a | 66.53 ± 4.22 C,b | 123.37 ± 9.59 D,c | 208.20 ± 19.41 C,d | |
| R | 2.17 ± 0.52 A,a | 2.76 ± 0.53 A,a | 7.59 ± 1.94 A,b | 11.41 ± 2.26 A,c | 19.36 ± 0.92 A,d | 144.03 ± 0.25 A,e | |
| Iso-butyric acid | C | ND | ND | ND | 18.83 ± 5.99 A,a | 51.38 ± 4.08 A,b | 91.35 ± 4.30 B,c |
| L | ND | ND | ND | 33.31 ± 2.60 B,a | 60.29 ± 3.27 B,b | 120.16 ± 0.44 C,c | |
| H | ND | ND | 6.48 ± 0.61 A,a | 34.27 ± 0.42 B,b | 57.07 ± 1.02 AB,c | 98.58 ± 14.28 B,d | |
| R | ND | ND | ND | ND | ND | 22.92 ± 3.65 A,a | |
| Valeric acid | C | ND | ND | ND | ND | ND | 4.50 ± 0.50 B,a |
| L | ND | ND | ND | ND | 1.13 ± 0.19 A,a | 8.69 ± 0.34 C,b | |
| H | ND | ND | ND | ND | 1.26 ± 0.09 A,a | 8.30 ± 0.89 C,b | |
| R | ND | ND | ND | ND | ND | 2.30 ± 1.33 A,a | |
| Iso-valeric acid | C | ND | ND | ND | ND | ND | ND |
| L | ND | ND | ND | ND | ND | ND | |
| H | ND | ND | ND | ND | ND | ND | |
| R | ND | ND | ND | ND | ND | ND |
| Sample ID | Observed Species | Chao1 | Shannon | Simpson | Coverage |
|---|---|---|---|---|---|
| C0 | 82.00 ± 0.82 A | 88.18 ± 3.48 A | 4.27 ± 0.05 A | 0.08 ± 0.00 E | 1.00 ± 0.00 |
| C12 | 67.67 ± 0.94 BC | 82.27 ± 9.38 AB | 2.29 ± 0.07 D | 0.41 ± 0.01 A | 1.00 ± 0.00 |
| C24 | 70.67 ± 2.05 B | 82.54 ± 1.15 AB | 3.17 ± 0.19 B | 0.20 ± 0.02 C | 1.00 ± 0.00 |
| L12 | 66.33 ± 2.49 C | 86.98 ± 9.17 A | 2.63 ± 0.04 C | 0.24 ± 0.01 B | 1.00 ± 0.00 |
| L24 | 67.00 ± 0.82 C | 78.26 ± 7.54 ABC | 3.31 ± 0.07 B | 0.15 ± 0.01 D | 1.00 ± 0.00 |
| H12 | 57.33 ± 1.70 E | 64.56 ± 4.28 C | 2.51 ± 0.08 C | 0.24 ± 0.01 B | 1.00 ± 0.00 |
| H24 | 61.67 ± 0.47 D | 67.29 ± 5.45 BC | 3.25 ± 0.00 B | 0.15 ± 0.00 D | 1.00 ± 0.00 |
| R12 | 60.33 ± 2.05 DE | 71.33 ± 3.92 ABC | 1.99 ± 0.07 E | 0.40 ± 0.02 A | 1.00 ± 0.00 |
| R24 | 62.67 ± 0.47 D | 75.83 ± 12.22 ABC | 2.13 ± 0.15 DE | 0.39 ± 0.03 A | 1.00 ± 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, C.; Tian, L.; Hong, H.; Wang, Q.; Zhan, X.; Luo, Y.; Tan, Y. In Vitro Gut Fermentation of Whey Protein Hydrolysate: An Evaluation of Its Potential Modulation on Infant Gut Microbiome. Nutrients 2022, 14, 1374. https://doi.org/10.3390/nu14071374
Feng C, Tian L, Hong H, Wang Q, Zhan X, Luo Y, Tan Y. In Vitro Gut Fermentation of Whey Protein Hydrolysate: An Evaluation of Its Potential Modulation on Infant Gut Microbiome. Nutrients. 2022; 14(7):1374. https://doi.org/10.3390/nu14071374
Chicago/Turabian StyleFeng, Chunsong, Li Tian, Hui Hong, Quanyu Wang, Xin Zhan, Yongkang Luo, and Yuqing Tan. 2022. "In Vitro Gut Fermentation of Whey Protein Hydrolysate: An Evaluation of Its Potential Modulation on Infant Gut Microbiome" Nutrients 14, no. 7: 1374. https://doi.org/10.3390/nu14071374
APA StyleFeng, C., Tian, L., Hong, H., Wang, Q., Zhan, X., Luo, Y., & Tan, Y. (2022). In Vitro Gut Fermentation of Whey Protein Hydrolysate: An Evaluation of Its Potential Modulation on Infant Gut Microbiome. Nutrients, 14(7), 1374. https://doi.org/10.3390/nu14071374