
The Controversial Role of HCY and Vitamin B Deficiency in Cardiovascular Diseases


{kind=link}
{kind=link}
Abstract
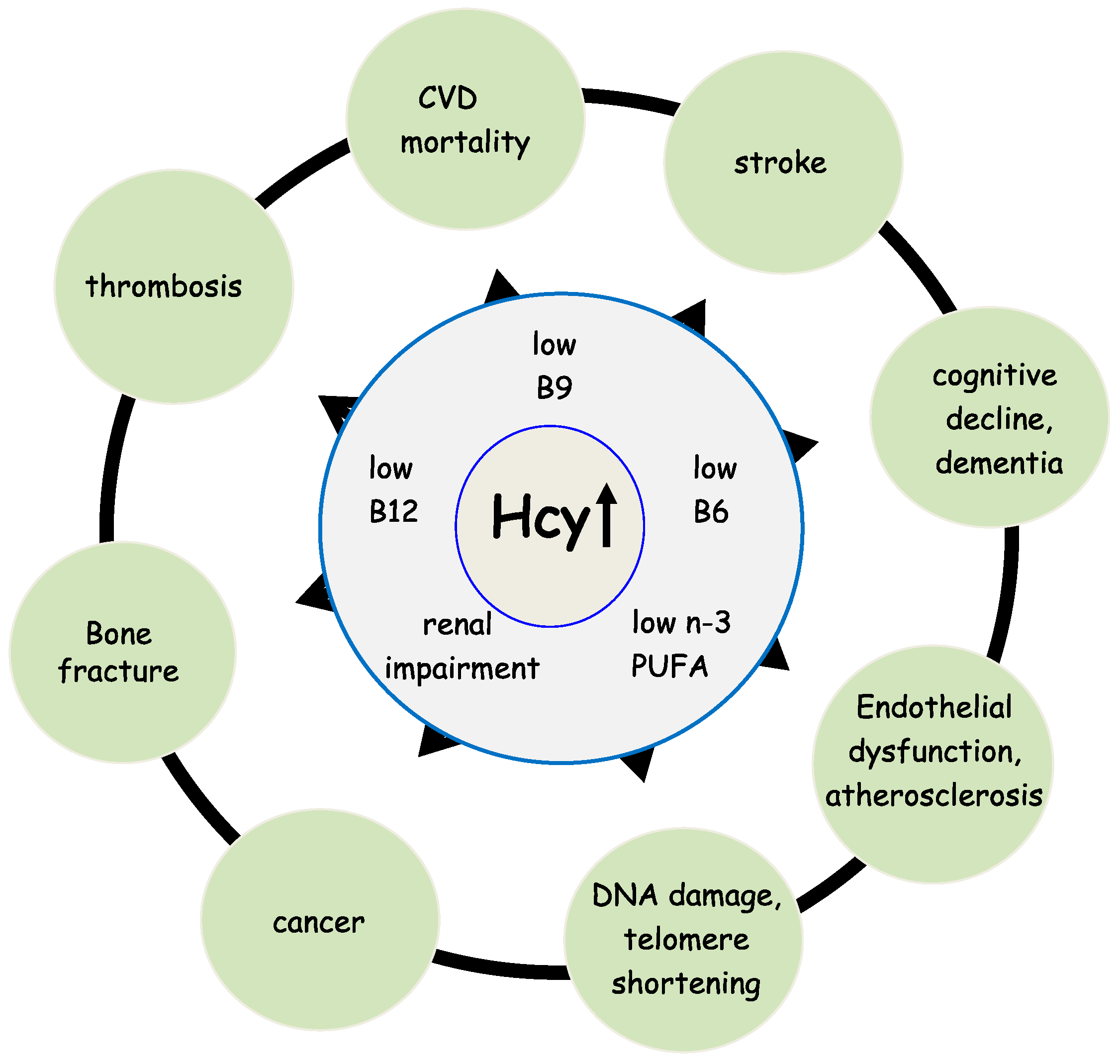
:1. Introduction
2. Homocysteine, B-Vitamin Deficiency, and Mortality
3. HCY, B-Vitamins, and CVD Risk
4. Hyperhomocysteinemia and Vascular Disease—Mechanistic Aspects
5. HHcy, B-Vitamin Deficiency and Genomic Effects, and Telomere Length
6. Controversial Aspects of Hcy and B Vitamins in CVD
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pusceddu, I.; Herrmann, W.; Kleber, M.E.; Scharnagl, H.; Hoffmann, M.M.; Winklhofer-Roob, B.M.; März, W.; Herrmann, M. Subclinical inflammation, telomere shortening, homocysteine, vitamin B6, and mortality: The Ludwigshafen Risk and Cardiovascular Health Study. Eur. J. Nutr. 2020, 59, 1399–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganguly, P.; Alam, S.F. Role of homocysteine in the development of cardiovascular disease. Nutr. J. 2015, 14, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.-Y.; Tsai, Y.-W.; Chen, S.-Y.; Ho, C.-I.; Weng, Y.-M.; Hsiao, C.-T.; Li, W.-C. The association of leptin and homocysteine with renal function impairment in a population of Taiwanese adults. Clin. Nutr. 2015, 34, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Gorman, E.B.; Finell, R.H. Folate fortification for prevention of neural tube defects. In Vitamins in the Prevention of Human Diseases; Herrmann, W., Obeid, R., Eds.; De Gruyter: Berlin, Germany, 2011; pp. 114–125. [Google Scholar]
- Smith, A.D.; Refsum, H. Homocysteine, B Vitamins, and Cognitive Impairment. Annu. Rev. Nutr. 2016, 36, 211–239. [Google Scholar] [CrossRef]
- Fan, R.; Zhang, A.; Zhong, F. Association between Homocysteine Levels and All-cause Mortality: A Dose-Response Meta-Analysis of Prospective Studies. Sci. Rep. 2017, 7, 4769. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, W.; Obeid, R. Vitamins in the Prevention of Human Diseases; De Gruyter: Berlin, Germany, 2011. [Google Scholar]
- Christensen, K.E.; Mason, J.B.; Rozen, R. Folate and cancer risk. In Vitamins in the Prevention of Human Diseases; Herrmann, W., Obeid, R., Eds.; De Gruyter: Berlin, Germany, 2011; pp. 166–186. [Google Scholar]
- Herrmann, W.; Obeid, R. Utility and limitations of biochemical markers of vitamin B12 deficiency. Eur. J. Clin. Investig. 2013, 43, 231–237. [Google Scholar] [CrossRef]
- Hannibal, L.; Lysne, V.; Bjørke-Monsen, A.L.; Behringer, S.; Grünert, S.C.; Spiekerkoetter, U.; Jacobsen, D.W.; Blom, H.J. Biomarkers and Algorithms for the Diagnosis of Vitamin B12 Deficiency. Front. Mol. Biosci. 2016, 3, 27. [Google Scholar] [CrossRef] [Green Version]
- Pfeiffer, C.M.; Fazili, Z.; McCoy, L.; Zhang, M.; Gunter, E.W. Determination of folate vitamers in human serum by stable-is tope-dilution tandem mass spectrometry and comparison with radioassay and microbiologic assay. Clin. Chem. 2004, 50, 423–432. [Google Scholar] [CrossRef]
- Kirsch, S.H.; Knapp, J.-P.; Herrmann, W.; Obeid, R. Quantification of key folate forms in serum using stable-isotope dilution ultra performance liquid chromatography–tandem mass spectrometry. J. Chromatogr. B 2010, 878, 68–75. [Google Scholar] [CrossRef]
- Smulders, Y.M.; Smith, D.E.; Kok, R.M.; Teerlink, T.; Gellekink, H.; Vaes, W.H.; Stehouwer, C.D.; Jakobs, C. Red blood cell folate vitamer distribution in healthy subjects is determined by the methylenetetrahydrofolate reductase C677T polymorphism and by the total folate status. J. Nutr. Biochem. 2007, 18, 693–699. [Google Scholar] [CrossRef]
- Ward, M.; Hughes, C.F.; Strain, J.J.; Reilly, R.; Cunningham, C.; Molloy, A.M.; Horigan, G.; Casey, M.; McCarroll, K.; O’Kane, M.; et al. Impact of the common MTHFR 677C→T polymorphism on blood pressure in adulthood and role of riboflavin in modifying the genetic risk of hypertension: Evidence from the JINGO project. BMC Med. 2020, 18, 318. [Google Scholar] [CrossRef] [PubMed]
- Xiu, L.-L.; Lee, M.-S.; Wahlqvist, M.L.; Chen, R.C.-Y.; Huang, Y.-C.; Chen, K.-J.; Li, D. Low and high homocysteine are associated with mortality independent of B group vitamins but interactive with cognitive status in a free-living elderly cohort. Nutr. Res. 2012, 32, 928–939. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.-Y.; Man, C.-F.; Xu, J.; Fan, Y. Elevated homocysteine levels and risk of cardiovascular and all-cause mortality: A meta-analysis of prospective studies. J. Zhejiang Univ. Sci. B 2015, 16, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Refsum, H.; Nurk, E.; Smith, A.D.; Ueland, P.M.; Gjesdal, C.G.; Bjelland, I.; Tverdal, A.; Tell, G.S.; Nygaård, O.; Vollset, S.E. The Hordaland Homocysteine Study: A Community-Based Study of Homocysteine, Its Determinants, and Associations with Disease. J. Nutr. 2006, 136, S1731–S1740. [Google Scholar] [CrossRef] [Green Version]
- Vollset, S.E.; Refsum, H.; Tverdal, A.; Nygård, O.; Nordrehaug, J.E.; Tell, G.S.; Ueland, P.M. Plasma total homocysteine and cardiovascular and noncardiovascular mortality: The Hordaland Homocysteine Study. Am. J. Clin. Nutr. 2001, 74, 130–136. [Google Scholar] [CrossRef]
- Ueland, P.M.; Ulvik, A.; Rios-Avila, L.; Midttun, Ø.; Gregory, J.F. Direct and Functional Biomarkers of Vitamin B6 Status. Annu. Rev. Nutr. 2015, 35, 33–70. [Google Scholar] [CrossRef]
- Mayengbam, S.; Chleilat, F.; Reimer, R.A. Dietary Vitamin B6 Deficiency Impairs Gut Microbiota and Host and Microbial Metabolites in Rats. Biomedicines 2020, 8, 469. [Google Scholar] [CrossRef]
- Swart, K.M.; van Schoor, N.; Blom, H.J.; Smulders, Y.M.; Lips, P. Homocysteine and the risk of nursing home admission and mortality in older persons. Eur. J. Clin. Nutr. 2011, 66, 188–195. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Huang, Y.; Hu, Y.; Zhong, J.; He, Z.; Li, W.; Yang, Y.; Xu, D.; Wu, S. Hyperhomocysteinemia is an independent risk factor in young patients with coronary artery disease in southern China. Herz 2013, 38, 779–784. [Google Scholar] [CrossRef]
- Kardesoglu, E.; Uz, O.; Isilak, Z.; Cebeci, B.S. Homocysteine as a new risk factor for cardiovascular events in heart failure. Int. J. Cardiol. 2011, 146, 126–127. [Google Scholar] [CrossRef]
- Ulvik, A.; Pedersen, E.R.; Svingen, G.F.; McCann, A.; Midttun, Ø.; Nygård, O.; Ueland, P.M. Vitamin B-6 catabolism and long-term mortality risk in patients with coronary artery disease. Am. J. Clin. Nutr. 2016, 103, 1417–1425. [Google Scholar] [CrossRef] [PubMed]
- Patterson, C.C.; Blankenberg, S.; Ben-Shlomo, Y.; Heslop, L.; Bayer, A.; Lowe, G.; Zeller, T.; Gallacher, J.; Young, I.; Yarnell, J. Which biomarkers are predictive specifically for cardiovascular or for non-cardiovascular mortality in men? Evidence from the Caerphilly Prospective Study (CaPS). Int. J. Cardiol. 2015, 201, 113–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, K.S.; Kwok, T.C.; Celermajer, D.S. Vegan Diet, Subnormal Vitamin B-12 Status and Cardiovascular Health. Nutrients 2014, 6, 3259–3273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pusceddu, I.; Herrmann, W.; Kleber, M.E.; Scharnagl, H.; März, W.; Herrmann, M. Telomere length, vitamin B12 and mortality in persons undergoing coronary angiography: The Ludwigshafen risk and cardiovascular health study. Aging 2019, 11, 7083–7097. [Google Scholar] [CrossRef] [PubMed]
- Hoogeveen, E.K.; Kostense, P.J.; Jakobs, C.; Dekker, J.M.; Nijpels, G.; Heine, R.J.; Bouter, L.M.; Stehouwer, C.D. Hyperhomocysteinemia increases risk of death, especially in type 2 diabetes: 5-year follow-up of the Hoorn Study. Circulation 2000, 101, 1506–1511. [Google Scholar] [CrossRef] [Green Version]
- Stabler, S.P. Clinical practice. Vitamin B12 deficiency. N. Engl. J. Med. 2013, 368, 149–160. [Google Scholar] [CrossRef]
- Dali-Youcef, N.; Andres, E. An update on cobalamin deficiency in adults. QJM Int. J. Med. 2009, 102, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Corcoran, T.B.; O’Neill, M.P.; Webb, S.A.R.; Ho, K.M. Inflammation, Vitamin Deficiencies and Organ Failure in Critically Ill Patients. Anaesth. Intensive Care 2009, 37, 740–747. [Google Scholar] [CrossRef] [Green Version]
- Andrès, E.; Loukili, N.H.; Noel, E.; Kaltenbach, G.; Abdelgheni, M.B.; Perrin, A.E.; Noblet-Dick, M.; Maloisel, F.; Schlienger, J.-L.; Blicklé, J.-F. Vitamin B12 (cobalamin) deficiency in elderly patients. Can. Med. Assoc. J. 2004, 171, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Carmel, R. 10 Malabsorption of food cobalamin. Baillière’s Clin. Haematol. 1995, 8, 639–655. [Google Scholar] [CrossRef]
- Pannérec, A.; Migliavacca, E.; De Castro, A.; Michaud, J.; Karaz, S.; Goulet, L.; Rezzi, S.; Ng, T.P.; Bosco, N.; Larbi, A.; et al. Vitamin B12 deficiency and impaired expression of amnionless during aging. J. Cachex Sarcopenia Muscle 2018, 9, 41–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, C.; Ward, M.; Hoey, L.; McNulty, H. Vitamin B12 and ageing: Current issues and interaction with folate. Ann. Clin. Biochem. Int. J. Lab. Med. 2013, 50 Pt 4, 315–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sviri, S.; Khalaila, R.; Daher, S.; Bayya, A.; Linton, D.; Stav, I.; van Heerden, P. Increased Vitamin B12 levels are associated with mortality in critically ill medical patients. Clin. Nutr. 2012, 31, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Mendonça, N.; Jagger, C.; Granic, A.; Martin-Ruiz, C.; Mathers, J.C.; Seal, C.J.; Hill, T.R. Elevated Total Homocysteine in All Participants and Plasma Vitamin B12 Concentrations in Women Are Associated With All-Cause and Cardiovascular Mortality in the Very Old: The Newcastle 85+ Study. J. Gerontol. Ser. A 2018, 73, 1258–1264. [Google Scholar] [CrossRef]
- Argan, O.; Ural, D.; Karauzum, K.; Bozyel, S.; Aktaş, M.; Karauzum, I.Y.; Kozdag, G.; Agir, A.A. Elevated levels of vitamin B12 in chronic stable heart failure: A marker for subclinical liver damage and impaired prognosis. Ther. Clin. Risk Manag. 2018, 14, 1067–1073. [Google Scholar] [CrossRef] [Green Version]
- Callaghan, F.M.; Leishear, K.; Abhyankar, S.; Demner-Fushman, D.; McDonald, C.J. High vitamin B12 levels are not associated with increased mortality risk for ICU patients after adjusting for liver function: A cohort study. ESPEN J. 2014, 9, e76–e83. [Google Scholar] [CrossRef] [Green Version]
- Urbanski, G.; Hamel, J.-F.; Prouveur, B.; Annweiler, C.; Ghali, A.; Cassereau, J.; Lozac’H, P.; Lavigne, C.; Lacombe, V. Strength of the Association of Elevated Vitamin B12 and Solid Cancers: An Adjusted Case-Control Study. J. Clin. Med. 2020, 9, 474. [Google Scholar] [CrossRef] [Green Version]
- Strain, J.J.; Dowey, L.R.; Ward, M.; Pentieva, K.; McNulty, H. B-vitamins, homocysteine metabolism and CVD. Proc. Nutr. Soc. 2004, 63, 597–603. [Google Scholar] [CrossRef] [Green Version]
- Homocysteine Studies Collaboration. Homocysteine and risk of ischemic heart disease and stroke: A meta-analysis. JAMA 2002, 288, 2015–2022. [Google Scholar] [CrossRef]
- Wald, D.S.; Law, M.; Morris, J. Homocysteine and cardiovascular disease: Evidence on causality from a meta-analysis. BMJ 2002, 325, 1202–1206. [Google Scholar] [CrossRef] [Green Version]
- Frosst, P.; Blom, H.J.; Milos, R.; Goyette, P.; Sheppard, C.A.; Matthews, R.G.; Boers, G.J.H.; den Heijer, M.; Kluijtmans, L.A.J.; van den Heuve, L.P.; et al. A candidate genetic risk factor for vascular disease: A common mutation in methylenetetrahydrofolate reductase. Nat. Genet. 1995, 10, 111–113. [Google Scholar] [CrossRef] [PubMed]
- Brattström, L.; Zhang, Y.; Hurtig, M.; Refsum, H.; Östensson, S.; Fransson, L.; Jonés, K.; Landgren, F.; Brudin, L.; Ueland, P.M. A common methylenetetrahydrofolate reductase gene mutation and longevity. Atherosclerosis 1998, 141, 315–319. [Google Scholar] [CrossRef]
- Klerk, M.; Verhoef, P.; Clarke, R.; Blom, H.J.; Kok, F.J.; Schouten, E.G.; MTHFR Studies Collaboration Group. MTHFR 677C-->T polymorphism and risk of coronary heart disease: A meta-analysis. JAMA 2002, 288, 2023–2031. [Google Scholar] [CrossRef] [PubMed]
- Wald, D.S.; Morris, J.K.; Wald, N.J. Reconciling the Evidence on Serum Homocysteine and Ischaemic Heart Disease: A Meta-Analysis. PLoS ONE 2011, 6, e16473. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.P.; McNulty, H.; Ward, M.; Strain, J.J.; Trouton, T.G.; Hoeft, B.A.; Weber, P.; Roos, F.F.; Horigan, G.; McAnena, L.; et al. Blood pressure in treated hypertensive individuals with the MTHFR 677TT genotype is responsive to intervention with riboflavin: Findings of a targeted randomized trial. Hypertension 2013, 61, 1302–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kundu, S.; Sen, U. Endothelial Dysfunction: The Link Between Homocysteine and Hydrogen Sulfide. Curr. Med. Chem. 2014, 21, 3662–3672. [Google Scholar] [CrossRef]
- Selhub, J.; Jacques, P.F.; Bostom, A.G.; D’Agostino, R.B.; Wilson, P.W.; Belanger, A.J.; O’Leary, D.H.; Wolf, P.A.; Schaefer, E.J.; Rosenberg, I.H. Association between Plasma Homocysteine Concentrations and Extracranial Carotid-Artery Stenosis. N. Engl. J. Med. 1995, 332, 286–291. [Google Scholar] [CrossRef]
- Herrmann, M.; Whiting, M.J.; Veillard, A.-S.; Ehnholm, C.; Sullivan, D.R.; Keech, A.C.; for the FIELD Study Investigators. Plasma homocysteine and the risk of venous thromboembolism: Insights from the FIELD study. Clin. Chem. Lab. Med. 2012, 50, 2213–2219. [Google Scholar] [CrossRef] [Green Version]
- Ebbing, M.; Bønaa, K.H.; Arnesen, E.; Ueland, P.M.; Nordrehaug, J.E.; Rasmussen, K.; Njølstad, I.; Nilsen, D.W.; Refsum, H.; Tverdal, A.; et al. Combined analyses and extended follow-up of two randomized controlled homocysteine-lowering B-vitamin trials. J. Intern. Med. 2010, 268, 367–382. [Google Scholar] [CrossRef]
- Albert, C.M.; Cook, N.R.; Gaziano, J.M.; Zaharris, E.; MacFadyen, J.; Danielson, E.; Buring, J.E.; Manson, J.E. Effect of folic acid and B vitamins on risk of cardiovascular events and total mortality among women at high risk for cardiovascular disease: A randomized trial. JAMA 2008, 299, 2027–2036. [Google Scholar] [CrossRef] [Green Version]
- Lonn, E.; Yusuf, S.; Arnold, M.J.; Sheridan, P.; Pogue, J.; Micks, M.; McQueen, M.J.; Probstfield, J.; Fodor, G.; Held, C.; et al. Homocysteine lowering with folic acid and B vitamins in vascular disease. N. Engl. J. Med. 2006, 354, 1567–1577. [Google Scholar] [PubMed] [Green Version]
- Ebbing, M.; Bleie, Ø.; Ueland, P.M.; Nordrehaug, J.E.; Nilsen, D.W.; Vollset, S.E.; Refsum, H.; Pedersen, E.K.; Nygård, O. Mortality and cardiovascular events in patients treated with homocysteine-lowering B vitamins after coronary angiography: A randomized controlled trial. JAMA 2008, 300, 795–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bønaa, K.H.; Njølstad, I.; Ueland, P.M.; Schirmer, H.; Tverdal, A.; Steigen, T.; Wang, H.; Nordrehaug, J.E.; Arnesen, E.; Rasmussen, K. Homocysteine Lowering and Cardiovascular Events after Acute Myocardial Infarction. N. Engl. J. Med. 2006, 354, 1578–1588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toole, J.F.; Malinow, M.R.; Chambless, L.E.; Spence, J.D.; Pettigrew, L.C.; Howard, V.J.; Sides, E.G.; Wang, C.-H.; Stampfer, M.J. Lowering Homocysteine in Patients With Ischemic Stroke to Prevent Recurrent Stroke, Myocardial Infarction, and Death. JAMA J. Am. Med. Assoc. 2004, 291, 565–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VITATOPS Trial Study Group. The VITATOPS (Vitamins to Prevent Stroke) Trial: Rationale and design of an international, large, simple, randomised trial of homocysteine-lowering multivitamin therapy in patients with recent transient ischaemic attack or stroke. Cerebrov. Dis. 2002, 13, 120–126. [Google Scholar] [CrossRef]
- Armitage, J.M.; Bowman, L.; Clarke, R.J.; Wallendszus, K.; Bulbulia, R.; Rahimi, K.; Haynes, R.; Parish, S.; Sleight, P.; Peto, R.; et al. Effects of homocysteine-lowering with folic acid plus vitamin B12 vs placebo on mortality and major morbidity in myocardial infarction survivors: A randomized trial. JAMA 2010, 303, 2486–2494. [Google Scholar]
- Marchioli, R.; Barzi, F.; Bomba, E.; Chieffo, C.; Di Gregorio, D.; Di Mascio, R.; Franzosi, M.G.; Geraci, E.; Levantesi, G.; Maggioni, A.P.; et al. Early Protection Against Sudden Death by n-3 Polyunsaturated Fatty Acids After Myocardial Infarction. Circulation 2002, 105, 1897–1903. [Google Scholar] [CrossRef] [Green Version]
- Marchioli, R.; Levantesi, G.; Macchia, A.; Marfisi, R.M.; Nicolosi, G.L.; Tavazzi, L.; Tognoni, G.; Valagussa, F. Vitamin E increases the risk of developing heart failure after myocardial infarction: Results from the GISSI-Prevenzione trial. J. Cardiovasc. Med. 2006, 7, 347–350. [Google Scholar] [CrossRef]
- Stephens, N.G.; Parsons, A.; Schofield, P.M.; Kelly, F.; Cheeseman, K.; Mitchinson, M.J. Randomised controlled trial of vitamin E in patients with coronary disease: Cambridge Heart Antioxidant Study (CHAOS). Lancet 1996, 347, 781–786. [Google Scholar] [CrossRef]
- Heinz, J.; Kropf, S.; Domröse, U.; Westphal, S.; Borucki, K.; Luley, C.; Neumann, K.H.; Dierkes, J. B vitamins and the risk of total mortality and cardiovascular disease in end-stage renal disease: Results of a randomized controlled trial. Circulation 2010, 121, 1432–1438. [Google Scholar] [CrossRef] [Green Version]
- Bostom, A.G.; Carpenter, M.A.; Kusek, J.W.; Levey, A.S.; Hunsicker, L.; Pfeffer, M.A.; Selhub, J.; Jacques, P.F.; Cole, E.; Gravens-Mueller, L.; et al. Homocysteine-lowering and cardiovascular disease outcomes in kidney transplant recipients: Primary results from the Folic Acid for Vascular Outcome Reduction in Transplantation trial. Circulation 2011, 123, 1763–1770. [Google Scholar] [CrossRef] [PubMed]
- Potter, K.; Hankey, G.J.; Green, D.J.; Eikelboom, J.; Jamrozik, K.; Arnolda, L.F. The effect of long-term homocysteine-lowering on carotid intima-media thickness and flow-mediated vasodilation in stroke patients: A randomized controlled trial and meta-analysis. BMC Cardiovasc. Disord. 2008, 8, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, T.; Chen, Y.; Yang, B.; Yang, J.; Wahlqvist, M.L.; Li, D. Meta-analysis of B vitamin supplementation on plasma homocysteine, cardiovascular and all-cause mortality. Clin. Nutr. 2012, 31, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Martí-Carvajal, A.J.; Solà, I.; Lathyris, D.; Dayer, M. Homocysteine-lowering interventions for preventing cardiovascular events. Cochrane Database Syst. Rev. 2017, 8, CD006612. [Google Scholar]
- Ji, Y.; Tan, S.; Xu, Y.; Chandra, A.; Shi, C.; Song, B.; Qin, J.; Gao, Y. Vitamin B supplementation, homocysteine levels, and the risk of cerebrovascular disease: A meta-analysis. Neurology 2013, 81, 1298–1307. [Google Scholar] [CrossRef]
- Huo, Y.; Li, J.; Qin, X.; Huang, Y.; Wang, X.; Gottesman, R.F.; Tang, G.; Wang, B.; Chen, D.; He, M.; et al. Efficacy of folic acid therapy in primary prevention of stroke among adults with hypertension in China: The CSPPT randomized clinical trial. JAMA 2015, 313, 1325–1335. [Google Scholar] [CrossRef]
- Yang, Q.; Botto, L.D.; Erickson, J.D.; Berry, R.J.; Sambell, C.; Johansen, H.; Friedman, J. Improvement in Stroke Mortality in Canada and the United States, 1990 to 2002. Circulation 2006, 113, 1335–1343. [Google Scholar] [CrossRef]
- Sultan, S.; Murarka, S.; Jahangir, A.; Mookadam, F.; Tajik, A.J.; Jahangir, A. Vitamins for Cardiovascular Diseases: Is the Expense Justified? Cardiol. Rev. 2017, 25, 298–308. [Google Scholar] [CrossRef]
- Mursu, J.; Robien, K.; Harnack, L.J.; Park, K.; Jacobs, D.R., Jr. Dietary supplements and mortality rate in older women: The Iowa Women’s Health Study. Arch. Intern. Med. 2011, 171, 1625–1633. [Google Scholar] [CrossRef]
- Kushi, L.H.; Folsom, A.R.; Prineas, R.J.; Mink, P.J.; Wu, Y.; Bostick, R.M. Dietary antioxidant vitamins and death from coronary heart disease in postmenopausal women. N. Engl. J. Med. 1996, 334, 1156–1162. [Google Scholar] [CrossRef]
- Lichtenstein, A.H.; Appel, L.J.; Brands, M.; Carnethon, M.; Daniels, S.; Franch, H.A.; Franklin, B.; Kris-Etherton, P.; Harris, W.S.; Howard, B.; et al. Diet and lifestyle recommendations revision 2006: A scientific statement from the American Heart Association Nutrition Committee. Circulation 2006, 114, 82–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyagi, N.; Sedoris, K.C.; Steed, M.; Ovechkin, A.V.; Moshal, K.S.; Tyagi, S.C. Mechanisms of homocysteine-induced oxidative stress. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, H2649–H2656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Wang, X.; Kong, W. Hyperhomocysteinaemia and vascular injury: Advances in mechanisms and drug targets. Br. J. Pharmacol. 2018, 175, 1173–1189. [Google Scholar] [CrossRef] [PubMed]
- De SMoreira, D.; Figueiró, P.W.; Siebert, C.; Prezzi, C.A.; Rohden, F.; Guma, F.C.R.; Manfredini, V.; Wyse, A.T.S. Chronic Mild Hyperhomocysteinemia Alters Inflammatory and Oxidative/Nitrative Status and Causes Protein/DNA Damage, as well as Ultrastructural Changes in Cerebral Cortex: Is Acetylsalicylic Acid Neuroprotective? Neurotox. Res. 2018, 33, 580–592. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zeng, X.; Guo, J.; Wang, X. Effects of homocysteine on murine splenic B lymphocyte proliferation and its signal transduction mechanism. Cardiovasc. Res. 2001, 52, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Zeng, X.; Guo, J.; Wang, X. Oxidant stress mechanism of homocysteine potentiating Con A-induced proliferation in murine splenic T lymphocytes. Cardiovasc. Res. 2002, 53, 1035–1042. [Google Scholar] [CrossRef] [Green Version]
- Tarafdar, A.; Pula, G. The Role of NADPH Oxidases and Oxidative Stress in Neurodegenerative Disorders. Int. J. Mol. Sci. 2018, 19, 3824. [Google Scholar] [CrossRef] [Green Version]
- Welch, G.N.; Upchurch, G.R.; Farivar, R.S.; Pigazzi, A.; Vu, K.; Brecher, P.; Keaney, J.F.; Loscalzo, J. Homocysteine-induced nitric oxide production in vascular smooth-muscle cells by NF-kappa B-dependent transcriptional activation of Nos2. Proc. Assoc. Am. Physicians 1998, 110, 22–31. [Google Scholar]
- Gori, A.M.; Sofi, F.; Marcucci, R.; Giusti, B.; Gensini, G.F.; Abbate, R. Association between homocysteine, vitamin B6 concentrations and inflammation. Clin. Chem. Lab. Med. 2007, 45, 1728–1736. [Google Scholar] [CrossRef]
- Zeng, X.; Dai, J.; Remick, D.; Wang, X. Homocysteine Mediated Expression and Secretion of Monocyte Chemoattractant Protein-1 and Interleukin-8 in Human Monocytes. Circ. Res. 2003, 93, 311–320. [Google Scholar] [CrossRef]
- Zanin, R.F.; Bergamin, L.S.; Morrone, F.B.; Coutinho-Silva, R.; de Souza Wyse, A.T.; Battastini, A.M. Pathological concentrations of homocysteine increases IL-1β production in macrophages in a P2X7, NF-ĸB, and erk-dependent manner. Purinergic. Signal. 2015, 11, 463–470. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Li, W.; Chang, L.; Zhang, Z.; Tang, C.; Wang, N.; Zhu, Y.; Wang, X. Role of redox factor-1 in hyperhomocysteinemia-accelerated atherosclerosis. Free Radic. Biol. Med. 2006, 41, 1566–1577. [Google Scholar] [CrossRef] [PubMed]
- Prontera, C.; Martelli, N.; Evangelista, V.; D’Urbano, E.; Manarini, S.; Recchiuti, A.; Dragani, A.; Passeri, C.; Davì, G.; Romano, M. Homocysteine modulates the CD40/CD40L system. J. Am. Coll. Cardiol. 2007, 49, 2182–2190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grewal, I.S.; Flavell, R.A. CD40 and CD154 in cell-mediated immunity. Annu. Rev. Immunol. 1998, 16, 111–135. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Zhang, Z.; Li, W.; Dai, J.; Guan, Y.; Wang, X. Liver-X-receptor activator prevents homocysteine-induced production of IgG antibodies from murine B lymphocytes via the ROS-NF-kappaB pathway. Biochem. Biophys. Res. Commun. 2007, 357, 772–778. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Luo, H.; Zhang, L.; Huang, Y.; Liu, B.; Ma, K.; Feng, J.; Xie, J.; Zheng, J.; Hu, J.; et al. Hyperhomocysteinemia Exaggerates Adventitial Inflammation and Angiotensin II−Induced Abdominal Aortic Aneurysm in Mice. Circ. Res. 2012, 111, 1261–1273. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.-H.; Chen, F.-Y.; Wang, G.-S.; Chen, L.; Gao, W. Diet-induced hyperhomocysteinemia exacerbates vascular reverse remodeling of balloon-injured arteries in rat. Chin. Med. J. 2008, 121, 2265–2271. [Google Scholar] [CrossRef]
- Joshi, M.; Baipadithaya, G.; Balakrishnan, A.; Hegde, M.; Vohra, M.; Ahamed, R.; Nagri, S.K.; Ramachandra, L.; Satyamoorthy, K. Elevated homocysteine levels in type 2 diabetes induce constitutive neutrophil extracellular traps. Sci. Rep. 2016, 6, 36362. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Maqueda, M.; El Bekay, R.; Monteseirín, J.; Alba, G.; Chacón, P.; Vega, A.; Santa María, C.; Tejedo, J.R.; Martín-Nieto, J.; Bedoya, F.J.; et al. Homocysteine enhances superoxide anion release and NADPH oxidase assembly by human neutrophils. Effects on MAPK activation and neutrophil migration. Atherosclerosis 2004, 172, 229–238. [Google Scholar] [CrossRef]
- Qureshi, I.; Chen, H.; Brown, A.T.; Fitzgerald, R.; Zhang, X.; Breckenridge, J.; Kazi, R.; Crocker, A.J.; Stühlinger, M.C.; Lin, K.; et al. Homocysteine-induced vascular dysregulation is mediated by the NMDA receptor. Vasc. Med. 2005, 10, 215–223. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.S.; Lee, S.J.; Seo, K.W.; Bae, J.U.; Park, S.Y.; Kim, C.D. Homocysteine induces COX-2 expression in macrophages through ROS generated by NMDA receptor-calcium signaling pathways. Free Radic. Res. 2013, 47, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Binet, F.; Sapieha, P. ER Stress and Angiogenesis. Cell Metab. 2015, 22, 560–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, G.S.; van Thienen, J.V.; Werstuck, G.H.; Zhou, J.; Sood, S.K.; Dickhout, J.G.; de Koning, A.B.L.; Tang, D.; Wu, D.; Falk, E.; et al. TDAG51 Is Induced by Homocysteine, Promotes Detachment-mediated Programmed Cell Death, and Contributes to the Development of Atherosclerosis in Hyperhomocysteinemia. J. Biol. Chem. 2003, 278, 30317–30327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Outinen, P.A.; Sood, S.K.; Pfeifer, S.I.; Pamidi, S.; Podor, T.J.; Li, J.; Weitz, J.I.; Austin, R.C. Homocysteine-Induced Endoplasmic Reticulum Stress and Growth Arrest Leads to Specific Changes in Gene Expression in Human Vascular Endothelial Cells. Blood 1999, 94, 959–967. [Google Scholar] [CrossRef]
- Pusceddu, I.; Herrmann, M.; Kirsch, S.H.; Werner, C.; Hübner, U.; Bodis, M.; Laufs, U.; Wagenpfeil, S.; Geisel, J.; Herrmann, W. Prospective study of telomere length and LINE-1 methylation in peripheral blood cells: The role of B vitamins supplementation. Eur. J. Nutr. 2015, 55, 1863–1873. [Google Scholar] [CrossRef]
- Román, G.C.; Mancera-Páez, O.; Bernal, C. Epigenetic Factors in Late-Onset Alzheimer’s Disease: MTHFR and CTH Gene Polymorphisms, Metabolic Transsulfuration and Methylation Pathways, and B Vitamins. Int. J. Mol. Sci. 2019, 20, 319. [Google Scholar] [CrossRef] [Green Version]
- Van Dijk, S.C.; Enneman, A.W.; Swart, K.M.; van Wijngaarden, J.P.; Ham, A.C.; de Jonge, R.; Blom, H.J.; Feskens, E.J.; Geleijnse, J.M.; van Schoor, N.M.; et al. Effect of vitamin B12 and folic acid supplementation on biomarkers of endothelial function and inflammation among elderly individuals with hyperhomocysteinemia. Vasc. Med. 2016, 21, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Sun, X.; Liu, J.; Xie, X.; Cui, W.; Zhu, Y. Homocysteine Accelerates Senescence of Endothelial Cells via DNA Hypomethylation of Human Telomerase Reverse Transcriptase. Arter. Thromb. Vasc. Biol. 2015, 35, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Hu, H.; Fang, Z.; Cui, J.; Liu, F. CTRP6 inhibits PDGF-BB-induced vascular smooth muscle cell proliferation and migration. Biomed. Pharmacother. 2018, 103, 844–850. [Google Scholar] [CrossRef]
- Ren, R.; Horton, J.R.; Zhang, X.; Blumenthal, R.M.; Cheng, X. Detecting and interpreting DNA methylation marks. Curr. Opin. Struct. Biol. 2018, 53, 88–99. [Google Scholar] [CrossRef]
- Lu, L.-Y.; Kuang, H.; Korakavi, G.; Yu, X. Topoisomerase II Regulates the Maintenance of DNA Methylation. J. Biol. Chem. 2015, 290, 851–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arcidiacono, O.A.; Krejčí, J.; Suchánková, J.; Bártová, E. Deacetylation of Histone H4 Accompanying Cardiomyogenesis is Weakened in HDAC1-Depleted ES Cells. Int. J. Mol. Sci. 2018, 19, 2425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cong, G.; Yan, R.; Huang, H.; Wang, K.; Yan, N.; Jin, P.; Zhang, N.; Hou, J.; Chen, D.; Jia, S. Involvement of histone methylation in macrophage apoptosis and unstable plaque formation in methionine-induced hyperhomocysteinemic ApoE-/- mice. Life Sci. 2017, 173, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Jakubowski, H. Protein homocysteinylation: Possible mechanism underlying pathological consequences of elevated homocysteine levels. FASEB J. 1999, 13, 2277–2283. [Google Scholar] [CrossRef]
- Perła-Kaján, J.; Jakubowski, H. Paraoxonase 1 and homocysteine metabolism. Amino Acids 2012, 43, 1405–1417. [Google Scholar] [CrossRef]
- Moya, C.; Máñez, S. Paraoxonases: Metabolic role and pharmacological projection. Naunyn. Schmiedebergs Arch. Pharmacol. 2018, 391, 349–359. [Google Scholar] [CrossRef]
- Jakubowski, H. The Role of Paraoxonase 1 in the Detoxification of Homocysteine Thiolactone. Adv. Exp. Med. Biol. 2009, 660, 113–127. [Google Scholar] [CrossRef]
- Steele, M.L.; Fuller, S.; Maczurek, A.E.; Kersaitis, C.; Ooi, L.; Münch, G. Chronic Inflammation Alters Production and Release of Glutathione and Related Thiols in Human U373 Astroglial Cells. Cell. Mol. Neurobiol. 2012, 33, 19–30. [Google Scholar] [CrossRef]
- Christen, W.G.; Cook, N.R.; Van Denburgh, M.; Zaharris, E.; Albert, C.M.; Manson, J.E. Effect of Combined Treatment With Folic Acid, Vitamin B6, and Vitamin B12 on Plasma Biomarkers of Inflammation and Endothelial Dysfunction in Women. J. Am. Heart Assoc. 2018, 7, 8517. [Google Scholar] [CrossRef] [Green Version]
- Vermeulen, E.G.J.; Rauwerda, J.A.; Berg, M.V.D.; De Jong, S.C.; Schalkwijk, C.; Twisk, J.W.R.; Stehouwer, C.D.A. Homocysteine-lowering treatment with folic acid plus vitamin B6 lowers urinary albumin excretion but not plasma markers of endothelial function or C-reactive protein: Further analysis of secondary end-points of a randomized clinical trial. Eur. J. Clin. Investig. 2003, 33, 209–215. [Google Scholar] [CrossRef]
- Ulvik, A.; Midttun, Ø.; Pedersen, E.R.; Eussen, S.J.; Nygård, O.; Ueland, P.M. Evidence for increased catabolism of vitamin B-6 during systemic inflammation. Am. J. Clin. Nutr. 2014, 100, 250–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulvik, A.; Midttun, Ø.; Pedersen, E.R.; Nygård, O.; Ueland, P.M. Association of plasma B-6 vitamers with systemic markers of inflammation before and after pyridoxine treatment in patients with stable angina pectoris. Am. J. Clin. Nutr. 2012, 95, 1072–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balint, B.; Jepchumba, V.K.; Guéant, J.-L.; Guéant-Rodriguez, R.-M. Mechanisms of homocysteine-induced damage to the endothelial, medial and adventitial layers of the arterial wall. Biochimie 2020, 173, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Al-Maskari, M.Y.; Waly, M.; Ali, A.; Al-Shuaibi, Y.S.; Ouhtit, A. Folate and vitamin B12 deficiency and hyperhomocysteinemia promote oxidative stress in adult type 2 diabetes. Nutrition 2012, 28, e23–e26. [Google Scholar] [CrossRef]
- Lee, Y.-J.; Wang, M.-Y.; Lin, M.-C.; Lin, P.-T. Associations between Vitamin B-12 Status and Oxidative Stress and Inflammation in Diabetic Vegetarians and Omnivores. Nutrition 2016, 8, 118. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, M.; Pusceddu, I.; März, W.; Herrmann, W. Telomere biology and age-related diseases. Clin. Chem. Lab. Med. 2018, 56, 1210–1222. [Google Scholar] [CrossRef]
- Rode, L.; Nordestgaard, B.G.; Bojesen, S.E. Peripheral Blood Leukocyte Telomere Length and Mortality Among 64 637 Individuals From the General Population. JNCI J. Natl. Cancer Inst. 2015, 107, djv074. [Google Scholar] [CrossRef] [Green Version]
- Moores, C.; Fenech, M.; O’Callaghan, N.J. Telomere dynamics: The influence of folate and DNA methylation. Ann. N. Y. Acad. Sci. 2011, 1229, 76–88. [Google Scholar] [CrossRef]
- Fitzpatrick, A.L.; Kronmal, R.A.; Kimura, M.; Gardner, J.P.; Psaty, B.M.; Jenny, N.S.; Tracy, R.P.; Hardikar, S.; Aviv, A. Leukocyte Telomere Length and Mortality in the Cardiovascular Health Study. J. Gerontol. Ser. A 2011, 66, 421–429. [Google Scholar] [CrossRef] [Green Version]
- Pusceddu, I.; Kleber, M.; Delgado, G.; Herrmann, W.; März, W.; Herrmann, M. Telomere length and mortality in the Ludwigshafen Risk and Cardiovascular Health study. PLoS ONE 2018, 13, e0198373. [Google Scholar] [CrossRef] [Green Version]
- Boniewska-Bernacka, E.; Pańczyszyn, A.; Klinger, M. Telomeres and telomerase in risk assessment of cardiovascular diseases. Exp. Cell Res. 2020, 397, 112361. [Google Scholar] [CrossRef] [PubMed]
- Tan, R.; Lan, L. Guarding chromosomes from oxidative DNA damage to the very end. Acta Biochim. Biophys. Sin. 2016, 48, 617–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazidi, M.; Kengne, A.P.; Banach, M. Mineral and vitamin consumption and telomere length among adults in the United States. Pol. Arch. Intern. Med. 2017, 127, 87–90. [Google Scholar] [PubMed]
- O’Donovan, A.; Pantell, M.S.; Puterman, E.; Dhabhar, F.S.; Blackburn, E.H.; Yaffe, K.; Cawthon, R.M.; Opresko, P.L.; Hsueh, W.-C.; Satterfield, S.; et al. Cumulative Inflammatory Load Is Associated with Short Leukocyte Telomere Length in the Health, Aging and Body Composition Study. PLoS ONE 2011, 6, e19687. [Google Scholar] [CrossRef] [Green Version]
- Richards, J.; Valdes, A.; Gardner, J.; Kato, B.; Siva, A.; Kimura, M.; Lu, X.; Brown, M.; Aviv, A.; Spector, T. Homocysteine levels and leukocyte telomere length. Atherosclerosis 2008, 200, 271–277. [Google Scholar] [CrossRef]
- Rane, G.; Koh, W.-P.; Kanchi, M.M.; Wang, R.; Yuan, J.-M.; Wang, X. Association Between Leukocyte Telomere Length and Plasma Homocysteine in a Singapore Chinese Population. Rejuvenation Res. 2015, 18, 203–210. [Google Scholar] [CrossRef] [Green Version]
- Shin, C.; Baik, I. Leukocyte Telomere Length is Associated With Serum Vitamin B12 and Homocysteine Levels in Older Adults With the Presence of Systemic Inflammation. Clin. Nutr. Res. 2016, 5, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Paul, L.; Cattaneo, M.; D’Angelo, A.; Sampietro, F.; Fermo, I.; Razzari, C.; Fontana, G.; Eugene, N.; Jacques, P.F.; Selhub, J. Telomere Length in Peripheral Blood Mononuclear Cells Is Associated with Folate Status in Men. J. Nutr. 2009, 139, 1273–1278. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.J.; Prescott, J.; Giovannucci, E.; E Hankinson, S.; Rosner, B.; De Vivo, I. One-carbon metabolism factors and leukocyte telomere length. Am. J. Clin. Nutr. 2013, 97, 794–799. [Google Scholar] [CrossRef] [Green Version]
- Milne, E.; O’Callaghan, N.; Ramankutty, P.; de Klerk, N.; Greenop, K.R.; Armstrong, B.K.; Miller, M.; Fenech, M. Plasma micronutrient levels and telomere length in children. Nutrition 2015, 31, 331–336. [Google Scholar] [CrossRef] [Green Version]
- Nomura, S.J.; Robien, K.; Zota, A.R. Serum Folate, Vitamin B-12, Vitamin A, γ-Tocopherol, α-Tocopherol, and Carotenoids Do Not Modify Associations between Cadmium Exposure and Leukocyte Telomere Length in the General US Adult Population. J. Nutr. 2017, 147, 538–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Wen, X.; Wu, W.; Xu, E.; Zhang, Y.; Cui, W. Homocysteine-related hTERT DNA demethylation contributes to shortened leukocyte telomere length in atherosclerosis. Atherosclerosis 2013, 231, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.-H.; Wen, X.-M.; Zhang, L.; Cui, W. DNA Methylation of Human Telomerase Reverse Transcriptase Associated With Leukocyte Telomere Length Shortening in Hyperhomocysteinemia-Type Hypertension in Humans and in a Rat Model. Circ. J. 2014, 78, 1915–1923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenech, M. Folate (vitamin B9) and vitamin B12 and their function in the maintenance of nuclear and mitochondrial genome integrity. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2012, 733, 21–33. [Google Scholar] [CrossRef]
- Kolling, J.; Scherer, E.B.; da Cunha, A.A.; da Cunha, M.J.; Wyse, A.T.S. Homocysteine Induces Oxidative–Nitrative Stress in Heart of Rats: Prevention by Folic Acid. Cardiovasc. Toxicol. 2010, 11, 67–73. [Google Scholar] [CrossRef]
- Matté, C.; Mackedanz, V.; Stefanello, F.M.; Scherer, E.B.; Andreazza, A.C.; Zanotto, C.; Moro, A.M.; Garcia, S.C.; Gonçalves, C.A.; Erdtmann, B.; et al. Chronic hyperhomocysteinemia alters antioxidant defenses and increases DNA damage in brain and blood of rats: Protective effect of folic acid. Neurochem. Int. 2009, 54, 7–13. [Google Scholar] [CrossRef]
- Li, J.-J.; Li, Q.; Du, H.-P.; Wang, Y.-L.; You, S.-J.; Wang, F.; Xu, X.-S.; Cheng, J.; Cao, Y.-J.; Liu, C.-F.; et al. Homocysteine Triggers Inflammatory Responses in Macrophages through Inhibiting CSE-H2S Signaling via DNA Hypermethylation of CSE Promoter. Int. J. Mol. Sci. 2015, 16, 12560–12577. [Google Scholar] [CrossRef]
- Opresko, P.L.; Fan, J.; Danzy, S.; Wilson, D.M., 3rd; Bohr, V.A. Oxidative damage in telomeric DNA disrupts recognition by TRF1 and TRF. Nucleic Acids Res. 2005, 33, 1230–1239. [Google Scholar] [CrossRef]
- Prasad, K.N.; Wu, M.; Bondy, S.C. Telomere shortening during aging: Attenuation by antioxidants and anti-inflammatory agents. Mech. Ageing Dev. 2017, 164, 61–66. [Google Scholar] [CrossRef]
- Armanios, M. Telomeres and age-related disease: How telomere biology informs clinical paradigms. J. Clin. Investig. 2013, 123, 996–1002. [Google Scholar] [CrossRef] [Green Version]
- Friso, S.; Udali, S.; De Santis, D.; Choi, S.-W. One-carbon metabolism and epigenetics. Mol. Asp. Med. 2017, 54, 28–36. [Google Scholar] [CrossRef]
- Sibrian-Vazquez, M.; Escobedo, J.O.; Lim, S.; Samoei, G.K.; Strongin, R.M. Homocystamides promote free-radical and oxidative damage to proteins. Proc. Natl. Acad. Sci. USA 2009, 107, 551–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smulders, Y.M.; Blom, H. The homocysteine controversy. J. Inherit. Metab. Dis. 2011, 34, 93–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Qin, X.; Demirtas, H.; Li, J.; Mao, G.; Huo, Y.; Sun, N.; Liu, L.; Xu, X. Efficacy of folic acid supplementation in stroke prevention: A meta-analysis. Lancet 2007, 369, 1876–1882. [Google Scholar] [CrossRef]
- Giles, W.H.; Kittner, S.J.; Croft, J.B.; Anda, R.F.; Casper, M.L.; Ford, E.S. Serum Folate and Risk for Coronary Heart Disease: Results from a Cohort of US Adults. Ann. Epidemiol. 1998, 8, 490–496. [Google Scholar] [CrossRef]
- Van Halm, V.P.; Nurmohamed, M.T.; Twisk, J.W.; Dijkmans, B.A.; Voskuyl, A.E. Disease-modifying antirheumatic drugs are associated with a reduced risk for cardiovascular disease in patients with rheumatoid arthritis: A case control study. Arthritis Res. Ther. 2006, 8, R151. [Google Scholar] [CrossRef] [Green Version]
- Ridker, P.M.; Everett, B.M.; Pradhan, A.; MacFadyen, J.G.; Solomon, D.H.; Zaharris, E.; Mam, V.; Hasan, A.; Rosenberg, Y.; Iturriaga, E.; et al. Low-Dose Methotrexate for the Prevention of Atherosclerotic Events. N. Engl. J. Med. 2019, 380, 752–762. [Google Scholar] [CrossRef]
- Dayal, S.; Lentz, S.R. Murine Models of Hyperhomocysteinemia and Their Vascular Phenotypes. Arter. Thromb. Vasc. Biol. 2008, 28, 1596–1605. [Google Scholar] [CrossRef] [Green Version]
- Carnicer, R.; Navarro, M.A.; Arbonés-Mainar, J.M.; Acín, S.; Guzmán, M.A.; Surra, J.C.; Arnal, C.; Heras, M.D.L.; Blanco-Vaca, F.; Osada, J. Folic acid supplementation delays atherosclerotic lesion development in apoE-deficient mice. Life Sci. 2007, 80, 638–643. [Google Scholar] [CrossRef]
- Herrmann, W.; Herrmann, M. The Importance of Telomere Shortening for Atherosclerosis and Mortality. J. Cardiovasc. Dev. Dis. 2020, 7, 29. [Google Scholar] [CrossRef]
- Kalea, A.Z.; Drosatos, K.; Buxton, J.L. Nutriepigenetics and cardiovascular disease. Curr. Opin. Clin. Nutr. Metab. Care 2018, 21, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Rosato, V.; Temple, N.J.; La Vecchia, C.; Castellan, G.; Tavani, A.; Guercio, V. Mediterranean diet and cardiovascular disease: A systematic review and meta-analysis of observational studies. Eur. J. Nutr. 2019, 58, 173–191. [Google Scholar] [CrossRef] [PubMed]
- Tsoukalas, D.; Fragkiadaki, P.; Docea, A.O.; Alegakis, A.K.; Sarandi, E.; Vakonaki, E.; Salataj, E.; Kouvidi, E.; Nikitovic, D.; Kovatsi, L.; et al. Association of nutraceutical supplements with longer telomere length. Int. J. Mol. Med. 2019, 44, 218–226. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herrmann, W.; Herrmann, M. The Controversial Role of HCY and Vitamin B Deficiency in Cardiovascular Diseases. Nutrients 2022, 14, 1412. https://doi.org/10.3390/nu14071412
Herrmann W, Herrmann M. The Controversial Role of HCY and Vitamin B Deficiency in Cardiovascular Diseases. Nutrients. 2022; 14(7):1412. https://doi.org/10.3390/nu14071412
Chicago/Turabian StyleHerrmann, Wolfgang, and Markus Herrmann. 2022. "The Controversial Role of HCY and Vitamin B Deficiency in Cardiovascular Diseases" Nutrients 14, no. 7: 1412. https://doi.org/10.3390/nu14071412
APA StyleHerrmann, W., & Herrmann, M. (2022). The Controversial Role of HCY and Vitamin B Deficiency in Cardiovascular Diseases. Nutrients, 14(7), 1412. https://doi.org/10.3390/nu14071412