
miR-222 Is Involved in the Amelioration Effect of Genistein on Dexamethasone-Induced Skeletal Muscle Atrophy

 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Treatment
2.2. Muscle Atrophy Model
2.3. Cell Culture and Transfection
2.4. Skeletal Muscle Tissue Section and C2C12 Myotube Immunofluorescence Staining
2.5. Total RNA Extraction and RT-qPCR
2.6. Dual Luciferase Reporting System
2.7. Data Analysis in Public Databases
2.8. Statistical Analysis
3. Results
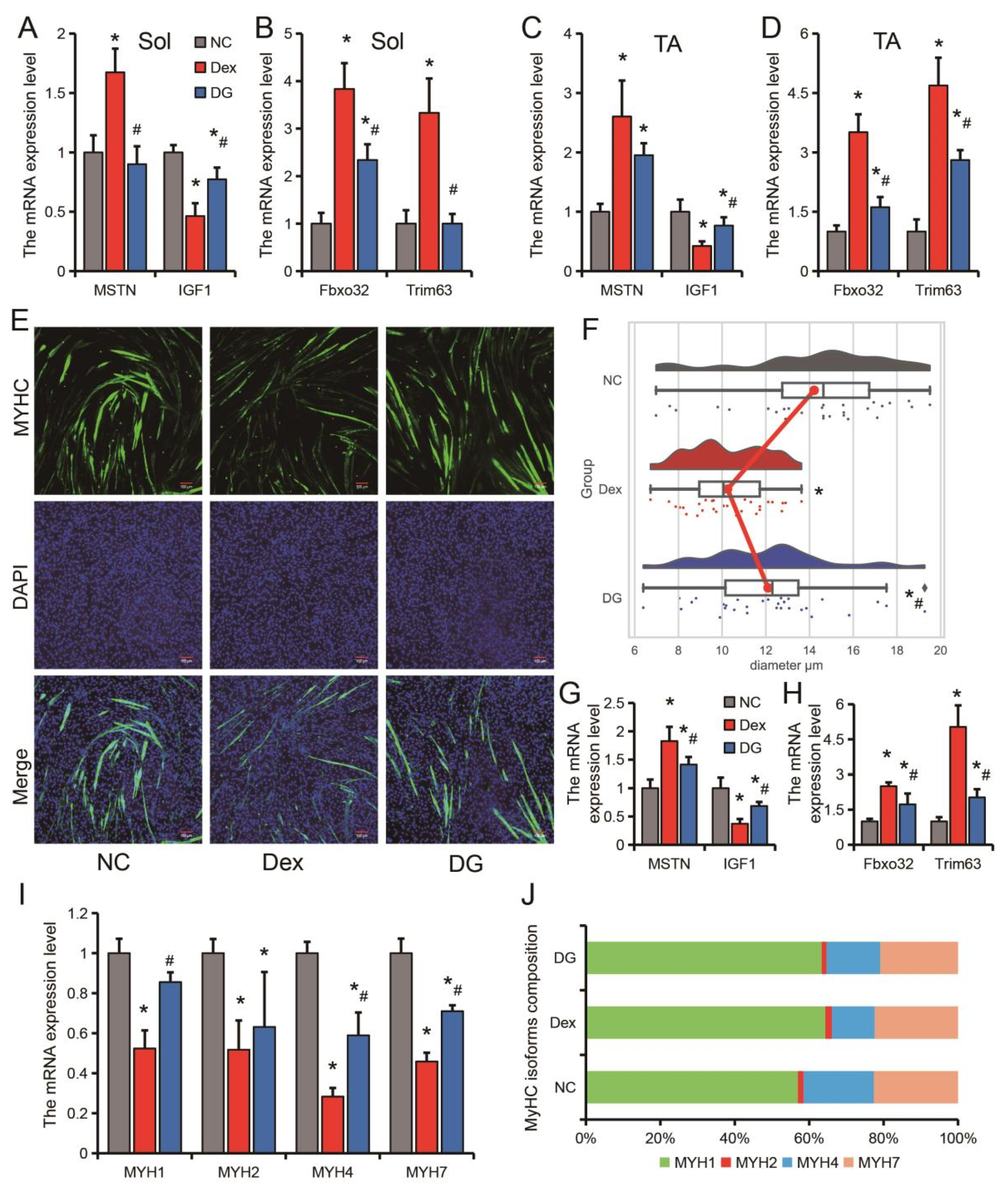
3.1. Genistein Alleviates Dexamethasone-Induced Skeletal Muscle Atrophy In Vivo
3.2. Genistein Alleviates Dexamethasone-Induced Skeletal Muscle Atrophy In Vitro
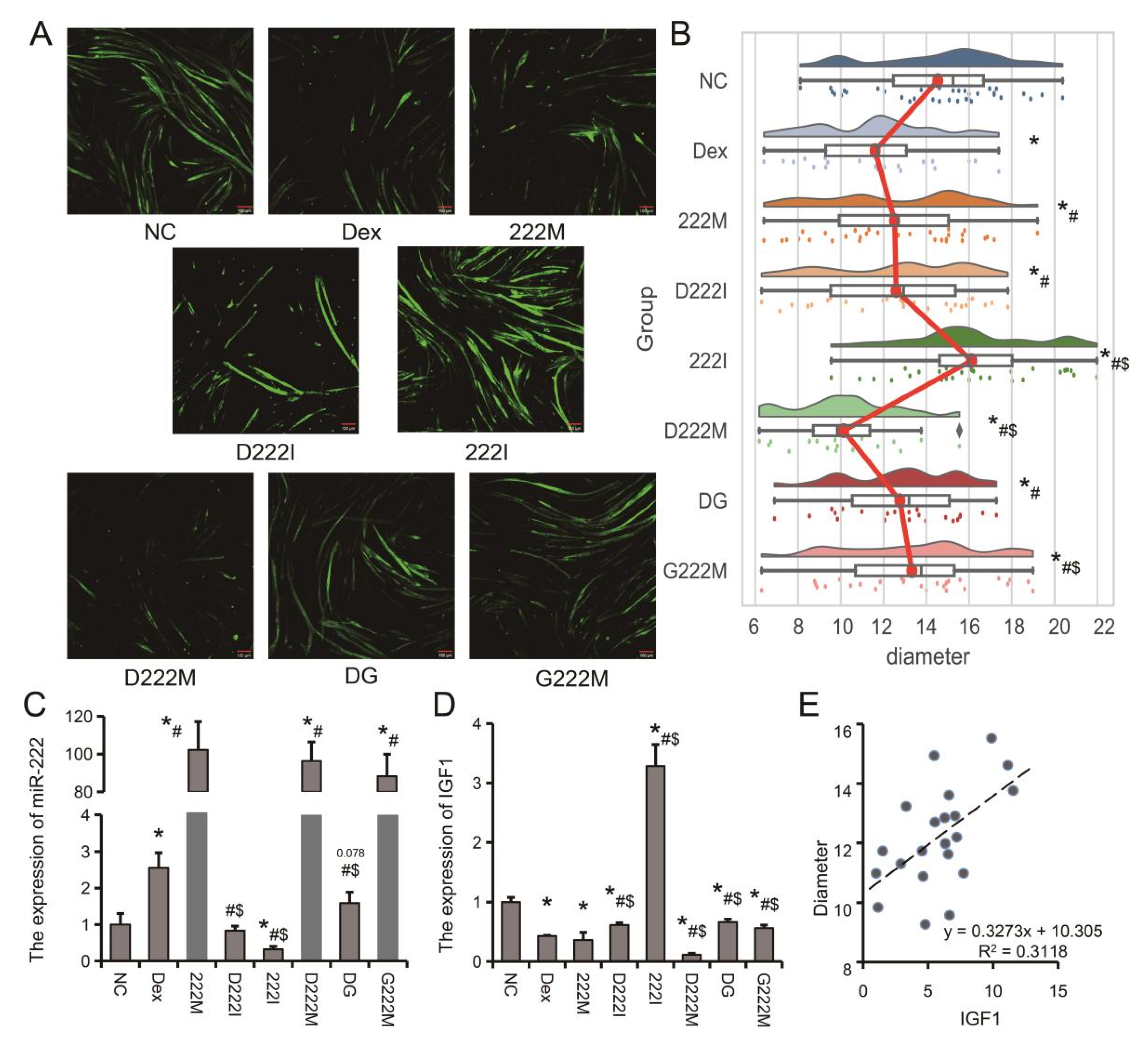
3.3. miR-222 Is One of the Core Regulators of Muscle Atrophy
3.4. IGF1 Is an Important Target Gene of miR-222 and Is Regulated by Genistein
3.5. Genistein Attenuates Dexamethasone-Induced Muscle Atrophy by Downregulating miR-222 In Vivo and In Vitro
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Shen, L.; Du, J.; Xia, Y.; Tan, Z.; Fu, Y.; Yang, Q.; Li, X.; Tang, G.; Jiang, Y.; Wang, J.; et al. Genome-wide landscape of DNA methylomes and their relationship with mrna and mirna transcriptomes in oxidative and glycolytic skeletal muscles. Sci. Rep. 2016, 6, 32186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, B.K.; Febbraio, M.A. Muscles, exercise and obesity: Skeletal muscle as a secretory organ. Nat. Rev. Endocrinol. 2012, 8, 457. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liang, J.; Chen, N. Do not neglect the role of circadian rhythm in muscle atrophy. Ageing Res. Rev. 2020, 63, 101155. [Google Scholar] [CrossRef] [PubMed]
- Siff, T.; Parajuli, P.; Razzaque, M.; Atfi, A. Cancer-mediated muscle cachexia: Etiology and clinical management. Trends Endocrinol. Metab. TEM 2021, 32, 382–402. [Google Scholar] [CrossRef]
- Yin, L.; Li, N.; Jia, W.; Wang, N.; Liang, M.; Yang, X.; Du, G. Skeletal muscle atrophy: From mechanisms to treatments. Pharmacol. Res. 2021, 172, 105807. [Google Scholar] [CrossRef]
- Jang, Y.; Rodriguez, K.; Lustgarten, M.; Muller, F.; Bhattacharya, A.; Pierce, A.; Choi, J.; Lee, N.; Chaudhuri, A.; Richardson, A.; et al. Superoxide-mediated oxidative stress accelerates skeletal muscle atrophy by synchronous activation of proteolytic systems. GeroScience 2020, 42, 1579–1591. [Google Scholar] [CrossRef]
- Stitt, T.; Drujan, D.; Clarke, B.; Panaro, F.; Timofeyva, Y.; Kline, W.; Gonzalez, M.; Yancopoulos, G.; Glass, D. The igf-1/pi3k/akt pathway prevents expression of muscle atrophy-induced ubiquitin ligases by inhibiting foxo transcription factors. Mol. Cell 2004, 14, 395–403. [Google Scholar] [CrossRef]
- Mc Pherron, A.; Lawler, A.; Lee, S. Regulation of skeletal muscle mass in mice by a new tgf-beta superfamily member. Nature 1997, 387, 83–90. [Google Scholar] [CrossRef]
- Cid-Díaz, T.; Santos-Zas, I.; González-Sánchez, J.; Gurriarán-Rodríguez, U.; Mosteiro, C.; Casabiell, X.; García-Caballero, T.; Mouly, V.; Pazos, Y.; Camiña, J. Obestatin controls the ubiquitin-proteasome and autophagy-lysosome systems in glucocorticoid-induced muscle cell atrophy. J. Cachexia Sarcopenia Muscle 2017, 8, 974–990. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Zhai, B.; Gygi, S.; Goldberg, A. Mtor inhibition activates overall protein degradation by the ubiquitin proteasome system as well as by autophagy. Proc. Natl. Acad. Sci. USA 2015, 112, 15790–15797. [Google Scholar] [CrossRef] [Green Version]
- Sukari, A.; Muqbil, I.; Mohammad, R.; Philip, P.; Azmi, A. F-box proteins in cancer cachexia and muscle wasting: Emerging regulators and therapeutic opportunities. Semin. Cancer Biol. 2016, 36, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Martín, A.; Priego, T.; López-Calderón, A. Hormones and muscle atrophy. Adv. Exp. Med. Biol. 2018, 1088, 207–233. [Google Scholar]
- Schakman, O.; Kalista, S.; Barbé, C.; Loumaye, A.; Thissen, J. Glucocorticoid-induced skeletal muscle atrophy. Int. J. Biochem. Cell Biol. 2013, 45, 2163–2172. [Google Scholar] [CrossRef]
- Su, Z.; Hu, L.; Cheng, J.; Klein, J.; Hassounah, F.; Cai, H.; Li, M.; Wang, H.; Wang, X. Acupuncture plus low-frequency electrical stimulation (acu-lfes) attenuates denervation-induced muscle atrophy. J. Appl. Physiol. 2016, 120, 426–436. [Google Scholar]
- Zhao, H.; Zhou, L.; Li, L.; Coon, V.J.; Chatterton, R.; Brooks, D.; Jiang, E.; Liu, L.; Xu, X.; Dong, Z.; et al. Shift from androgen to estrogen action causes abdominal muscle fibrosis, atrophy, and inguinal hernia in a transgenic male mouse model. Proc. Natl. Acad. Sci. USA 2018, 115, E10427–E10436. [Google Scholar] [CrossRef] [Green Version]
- Aoyama, S.; Jia, H.; Nakazawa, K.; Yamamura, J.; Saito, K.; Kato, H. Dietary genistein prevents denervation-induced muscle atrophy in male rodents via effects on estrogen receptor-α. J. Nutr. 2016, 146, 1147–1154. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Deng, J.; Qiu, Y.; Gao, J.; Li, J.; Guan, L.; Lee, H.; Zhou, Q.; Xiao, J. Non-coding rna basis of muscle atrophy. Mol. Ther. Nucleic Acids 2021, 26, 1066–1078. [Google Scholar] [CrossRef]
- Majid, S.; Dar, A.; Saini, S.; Chen, Y.; Shahryari, V.; Liu, J.; Zaman, M.; Hirata, H.; Yamamura, S.; Ueno, K.; et al. Regulation of minichromosome maintenance gene family by microrna-1296 and genistein in prostate cancer. Cancer Res. 2010, 70, 2809–2818. [Google Scholar] [CrossRef] [Green Version]
- Gan, M.; Shen, L.; Liu, L.; Guo, Z.; Wang, S.; Chen, L.; Zheng, T.; Fan, Y.; Tan, Y.; Jiang, D.; et al. Mir-222 is involved in the regulation of genistein on skeletal muscle fiber type. J. Nutr. Biochem. 2020, 80, 108320. [Google Scholar] [CrossRef]
- Gan, M.; Shen, L.; Fan, Y.; Tan, Y.; Zheng, T.; Tang, G.; Niu, L.; Zhao, Y.; Chen, L.; Jiang, D.; et al. Microrna-451 and genistein ameliorate nonalcoholic steatohepatitis in mice. Int. J. Mol. Sci. 2019, 20, 6084. [Google Scholar] [CrossRef] [Green Version]
- Gan, M.; Shen, L.; Wang, S.; Guo, Z.; Zheng, T.; Tan, Y.; Fan, Y.; Liu, L.; Chen, L.; Jiang, A.; et al. Genistein inhibits high fat diet-induced obesity through mir-222 by targeting btg2 and adipor1. Food Funct. 2020, 11, 2418–2426. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Chan, M.C.; Yan, Y.; Bei, Y.; Ping, C.; Zhou, Q.; Cheng, L.; Lei, C.; Ziegler, O.; Rowe, G.C. Mir-29b contributes to multiple types of muscle atrophy. Nat. Commun. 2017, 8, 15201. [Google Scholar]
- Son, Y.H.; Jang, E.J.; Kim, Y.W.; Lee, J.H. Sulforaphane prevents dexamethasone-induced muscle atrophy via regulation of the akt/foxo1 axis in c2c12 myotubes. Biomed. Pharmacother. 2017, 95, 1486. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.; Nam, J.; Bartel, D. Predicting effective microrna target sites in mammalian mrnas. Elife 2015, 4, e05005. [Google Scholar] [CrossRef] [PubMed]
- Krüger, J.; Rehmsmeier, M. Rnahybrid: Microrna target prediction easy, fast and flexible. Nucleic Acids Res. 2006, 34, W451–W454. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, X. Mirdb: An online database for prediction of functional microrna targets. Nucleic Acids Res. 2020, 48, D127–D131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gheller, B.; Riddle, E.; Lem, M.; Thalacker-Mercer, A. Understanding age-related changes in skeletal muscle metabolism: Differences between females and males. Annu. Rev. Nutr. 2016, 36, 129–156. [Google Scholar] [CrossRef] [PubMed]
- Praestholm, S.M.; Correia, C.M.; Grntved, L. Multifaceted control of gr signaling and its impact on hepatic transcriptional networks and metabolism. Front. Endocrinol. 2020, 11, 572981. [Google Scholar] [CrossRef]
- Vandewalle, J.; Luypaert, A.; De Bosscher, K.; Libert, C. Therapeutic mechanisms of glucocorticoids. Trends Endocrinol. Metab. TEM 2018, 29, 42–54. [Google Scholar] [CrossRef]
- Mehta, H.; An, H.; Andersen, K.; Mansour, O.; Madhira, V.; Rashidi, E.; Bates, B.; Setoguchi, S.; Joseph, C.; Kocis, P.; et al. Use of hydroxychloroquine, remdesivir, and dexamethasone among adults hospitalized with covid-19 in the united states: A retrospective cohort study. Ann. Intern. Med. 2021, 174, 1395–1403. [Google Scholar] [CrossRef]
- Yoshioka, Y.; Samukawa, Y.; Yamashita, Y.; Ashida, H. 4-hydroxyderricin and xanthoangelol isolated from angelica keiskei prevent dexamethasone-induced muscle loss. Food Funct. 2020, 11, 5498–5512. [Google Scholar] [CrossRef]
- Fitts, R.; Widrick, J. Muscle mechanics: Adaptations with exercise-training. Exerc. Sport Sci. Rev. 1996, 24, 427–473. [Google Scholar] [CrossRef]
- Seene, T.; Viru, A. The catabolic effect of glucocorticoids on different types of skeletal muscle fibres and its dependence upon muscle activity and interaction with anabolic steroids. J. Steroid Biochem. Mol. Biol. 1982, 16, 349–352. [Google Scholar] [CrossRef]
- Dayton, W.; White, M. Meat science and muscle biology symposium-role of satellite cells in anabolic steroid-induced muscle growth in feedlot steers. J. Anim. Sci. 2014, 92, 30–38. [Google Scholar] [CrossRef]
- Wiik, A.; Ekman, M.; Johansson, O.; Jansson, E.; Rnsson, M. Expression of both oestrogen receptor alpha and beta in human skeletal muscle tissue. Histochem. Cell Biol. 2009, 131, 181. [Google Scholar] [CrossRef]
- Wiik, A.T.; Gustafsson, T.; Esbjörnsson, M.; Johansson, O.; Jansson, E. Expression of oestrogen receptor alpha and beta is higher in skeletal muscle of highly endurance-trained than of moderately active men. Acta Physiol. 2010, 184, 105–112. [Google Scholar] [CrossRef]
- Barros, R.; Machado, U.F.; Warner, M.; Gustafsson, J.A. Muscle glut4 regulation by estrogen receptors erβ and erα. Proc. Natl. Acad. Sci. USA 2006, 103, 1605–1608. [Google Scholar] [CrossRef] [Green Version]
- Hevener, A.; Ribas, V.; Moore, T.; Zhou, Z. The impact of skeletal muscle erα on mitochondrial function and metabolic health. Endocrinology 2020, 161, bqz017. [Google Scholar] [CrossRef]
- Park, K.-S.; Kim, H.; Kim, H.J.; Lee, K.-I.; Lee, S.-Y.; Kim, J. Paeoniflorin alleviates skeletal muscle atrophy in ovariectomized mice through the erα/nrf1 mitochondrial biogenesis pathway. Pharmaceuticals 2022, 15, 390. [Google Scholar] [CrossRef]
- Soleymani, T.; Daraei, P.; Ribas, V.; Drew, B.; Hevener, A. Erα Is Elevated with Endurance Exercise and Is Critical for Normal Skeletal Muscle Oxidative Metabolism and Insulin Action; American College of Sports Medicine, Southwestern Regional Meeting: Los Angeles, CA, USA, 2009. [Google Scholar]
- Yin, X.U.; Guan, J. Research progress on biological function of genistein. Med. Recapitul. 2010, 16, 735–737. [Google Scholar]
- Zilahi, E.; Adamecz, Z.; Bodoki, L.; Griger, Z.; Póliska, S.; Nagy-Vincze, M.; Dankó, K. Dysregulated expression profile of myomirs in the skeletal muscle of patients with polymyositis. Ejifcc 2019, 30, 237–245. [Google Scholar] [PubMed]
- Srivastava, S.; Rathor, R.; Singh, S.; Suryakumar, G. Emerging role of myomirs as biomarkers and therapeutic targets in skeletal muscle diseases. Am. J. Physiol. Cell Physiol. 2021, 321, C859–C875. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, J. The myomir network in skeletal muscle plasticity. Exerc. Sport Sci. Rev. 2011, 39, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Marinho, R.; Alcântara, P.; Ottoch, J.; Seelaender, M. Role of exosomal micrornas and myomirs in the development of cancer cachexia-associated muscle wasting. Front. Nutr. 2017, 4, 69. [Google Scholar] [CrossRef] [Green Version]
- Freire, P.; Cury, S.; Lopes, L.; Fernandez, G.; Liu, J.; de Moraes, L.; de Oliveira, G.; Oliveira, J.; de Moraes, D.; Cabral-Marques, O.; et al. Decreased mir-497-5p suppresses il-6 induced atrophy in muscle cells. Cells 2021, 10, 3527. [Google Scholar] [CrossRef]
- Yang, X.; Li, Z.; Wang, Z.; Yu, J.; Ma, M.; Nie, Q. Mir-27b-3p attenuates muscle atrophy by targeting cbl-b in skeletal muscles. Biomolecules 2022, 12, 191. [Google Scholar] [CrossRef]
- Chen, T.; Kuo, T.; Dandan, M.; Lee, R.; Chang, M.; Villivalam, S.; Liao, S.; Costello, D.; Shankaran, M.; Mohammed, H.; et al. The role of striated muscle pik3r1 in glucose and protein metabolism following chronic glucocorticoid exposure. J. Biol. Chem. 2021, 296, 100395. [Google Scholar] [CrossRef]
- Schiaffino, S.; Dyar, K.; Ciciliot, S.; Blaauw, B.; Sandri, M. Mechanisms regulating skeletal muscle growth and atrophy. FEBS J. 2013, 280, 4294–4314. [Google Scholar] [CrossRef]
- Brooks, N.; Myburgh, K. Skeletal muscle wasting with disuse atrophy is multi-dimensional: The response and interaction of myonuclei, satellite cells and signaling pathways. Front. Physiol. 2014, 5, 99. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gan, M.; Ma, J.; Chen, J.; Chen, L.; Zhang, S.; Zhao, Y.; Niu, L.; Li, X.; Zhu, L.; Shen, L. miR-222 Is Involved in the Amelioration Effect of Genistein on Dexamethasone-Induced Skeletal Muscle Atrophy. Nutrients 2022, 14, 1861. https://doi.org/10.3390/nu14091861
Gan M, Ma J, Chen J, Chen L, Zhang S, Zhao Y, Niu L, Li X, Zhu L, Shen L. miR-222 Is Involved in the Amelioration Effect of Genistein on Dexamethasone-Induced Skeletal Muscle Atrophy. Nutrients. 2022; 14(9):1861. https://doi.org/10.3390/nu14091861
Chicago/Turabian StyleGan, Mailin, Jianfeng Ma, Jingyun Chen, Lei Chen, Shunhua Zhang, Ye Zhao, Lili Niu, Xuewei Li, Li Zhu, and Linyuan Shen. 2022. "miR-222 Is Involved in the Amelioration Effect of Genistein on Dexamethasone-Induced Skeletal Muscle Atrophy" Nutrients 14, no. 9: 1861. https://doi.org/10.3390/nu14091861
APA StyleGan, M., Ma, J., Chen, J., Chen, L., Zhang, S., Zhao, Y., Niu, L., Li, X., Zhu, L., & Shen, L. (2022). miR-222 Is Involved in the Amelioration Effect of Genistein on Dexamethasone-Induced Skeletal Muscle Atrophy. Nutrients, 14(9), 1861. https://doi.org/10.3390/nu14091861