
Effects of Flavonoid-Rich Orange Juice Intervention on Major Depressive Disorder in Young Adults: A Randomized Controlled Trial



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Participants
2.3. MDD Assessments
2.4. Treatment Drinks
2.5. Blood Parameters
2.6. Dietary Assessment
2.7. Fecal Sample Collection and Quantifications of Fecal Short-Chain Fatty Acids
2.8. Biochemical Assays
2.9. PCR and Sequencing
2.10. Gut Microbiome Sequencing Data Analysis
2.11. Differential Abundance of Taxa
2.12. Statistical Analysis
3. Results
3.1. Subjects and Baseline Characteristics
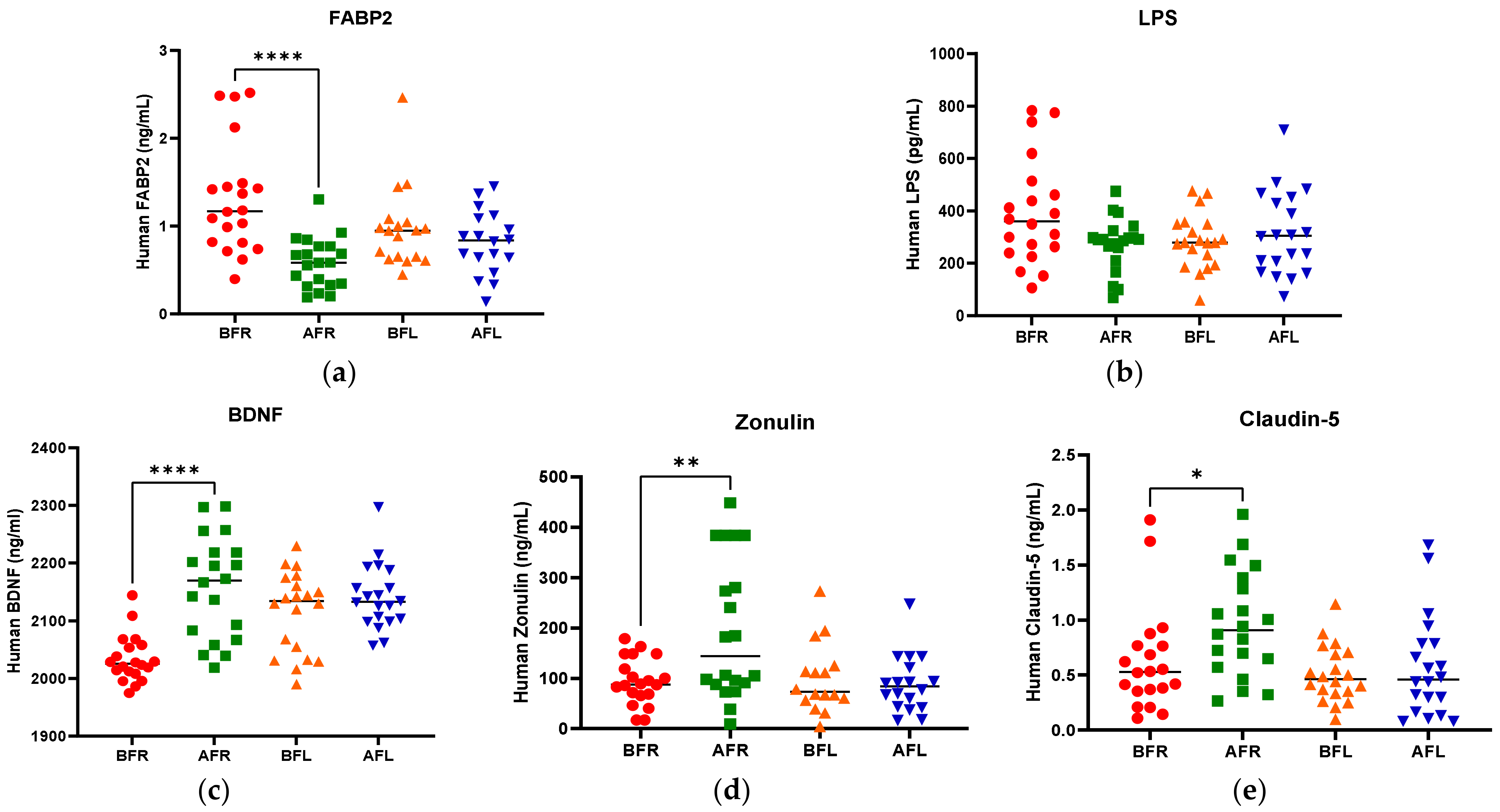
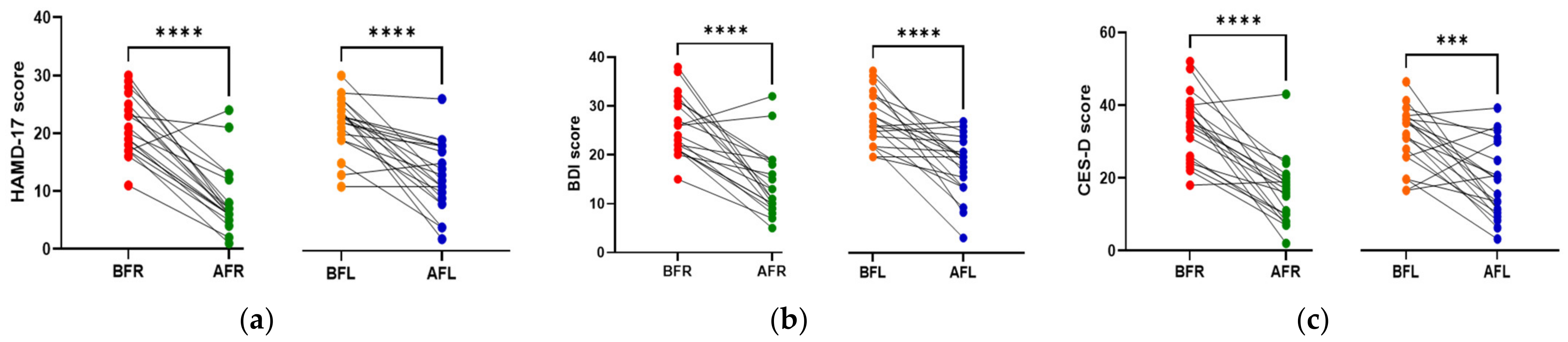
3.2. Anthropometric Data, Blood Parameters, and MDD Scores
3.3. Dietary Assessment
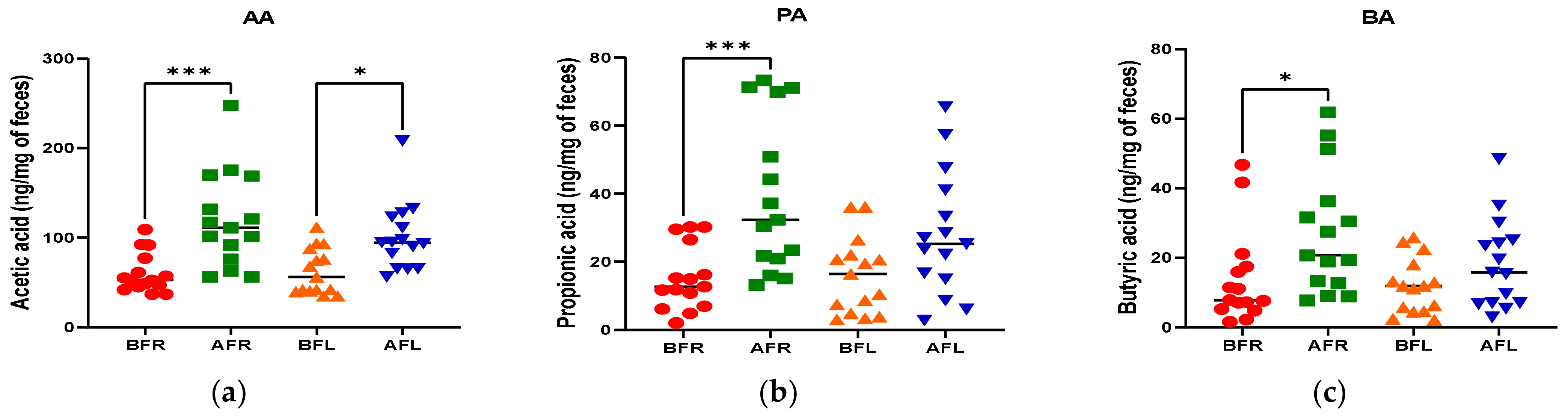
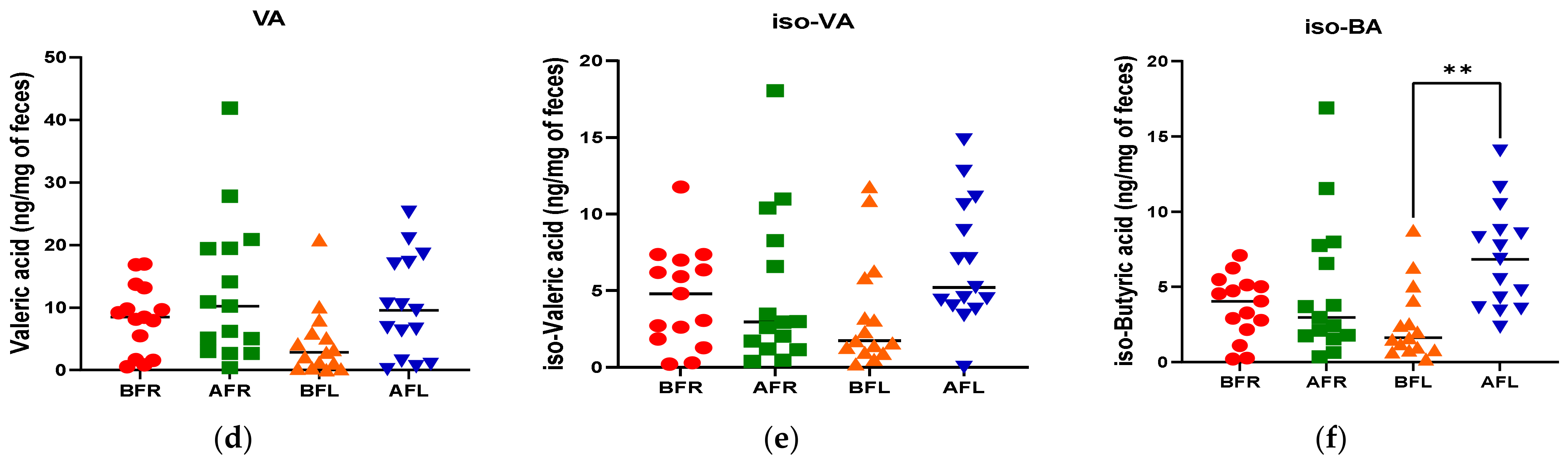
3.4. Fecal Short Chain Fatty Acids (SCFAs)
3.5. Gut Microbiome Diversity
3.6. Abundance of Bacterial Taxa
3.7. Correlations of Differences before and after Interventions in the FR Group
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kennis, M.; Gerritsen, L.; van Dalen, M.; Williams, A.; Cuijpers, P.; Bockting, C. Prospective biomarkers of major depressive disorder: A systematic review and meta-analysis. Mol. Psychiatry 2020, 25, 321–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malhi, G.S.; Mann, J.J. Depression. Lancet 2018, 392, 2299–2312. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Depression and Other Common Mental Disorders: Global Health Estimates; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Vlainić, J.V.; Šuran, J.; Vlainić, T.; Vukorep, A.L. Probiotics as an Adjuvant Therapy in Major Depressive Disorder. Curr. Neuropharmacol. 2016, 14, 952–958. [Google Scholar] [CrossRef] [PubMed]
- Cuijpers, P.; Sijbrandij, M.; Koole, S.L.; Andersson, G.; Beekman, A.T.; Reynolds, C.F., 3rd. Adding psychotherapy to antidepressant medication in depression and anxiety disorders: A meta-analysis. World Psychiatry 2014, 13, 56–67. [Google Scholar] [CrossRef] [Green Version]
- Lopez, J.P.; Kos, A.; Turecki, G. Major depression and its treatment: MicroRNAs as peripheral biomarkers of diagnosis and treatment response. Curr. Opin. Psychiatry 2018, 31, 7–16. [Google Scholar] [CrossRef]
- Hritcu, L.; Ionita, R.; Postu, P.A.; Gupta, G.K.; Turkez, H.; Lima, T.C.; Carvalho, C.U.S.; de Sousa, D.P. Antidepressant flavonoids and their relationship with oxidative stress. Oxidative Med. Cell. Longev. 2017, 2017, 5762172. [Google Scholar] [CrossRef] [Green Version]
- Jia, S.; Hou, Y.; Wang, D.; Zhao, X. Flavonoids for depression and anxiety: A systematic review and meta-analysis. Crit. Rev. Food Sci. Nutr. 2022, 9, 1–11. [Google Scholar] [CrossRef]
- Ali, S.; Corbi, G.; Maes, M.; Scapagnini, G.; Davinelli, S. Exploring the impact of flavonoids on symptoms of depression: A systematic review and meta-analysis. Antioxidants 2021, 10, 1644. [Google Scholar] [CrossRef]
- Ko, Y.-H.; Kim, S.-K.; Lee, S.-Y.; Jang, C.-G. Flavonoids as therapeutic candidates for emotional disorders such as anxiety and depression. Arch. Pharmacal Res. 2020, 43, 1128–1143. [Google Scholar] [CrossRef]
- Dayan, E.; Cohen, L.G. Neuroplasticity subserving motor skill learning. Neuron 2011, 72, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Duman, R.S.; Aghajanian, G.K. Synaptic dysfunction in depression: Potential therapeutic targets. Science 2012, 338, 68–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krystal, J.H.; Sanacora, G.; Duman, R.S. Rapid-acting glutamatergic antidepressants: The path to ketamine and beyond. Biol. Psychiatry 2013, 73, 1133–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraus, C.; Castrén, E.; Kasper, S.; Lanzenberger, R. Serotonin and neuroplasticity—Links between molecular, functional and structural pathophysiology in depression. Neurosci. Biobehav. Rev. 2017, 77, 317–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castricum, J.; Birkenhager, T.K.; Kushner, S.A.; Elgersma, Y.; Tulen, J.H.M. Cortical Inhibition and Plasticity in Major Depressive Disorder. Front. Psychiatry 2022, 13, 777422. [Google Scholar] [CrossRef] [PubMed]
- Glinert, A.; Turjeman, S.; Elliott, E.; Koren, O. Microbes, metabolites and (synaptic) malleability, oh my! The effect of the microbiome on synaptic plasticity. Biol. Rev. Camb. Philos. Soc. 2022, 97, 582–599. [Google Scholar] [CrossRef] [PubMed]
- Martinowich, K.; Lu, B. Interaction between BDNF and serotonin: Role in mood disorders. Neuropsychopharmacology 2008, 33, 73–83. [Google Scholar] [CrossRef]
- Khan, A.; Brown, W.A. Antidepressants versus placebo in major depression: An overview. World Psychiatry 2015, 14, 294–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaffe, D.H.; Rive, B.; Denee, T.R. The humanistic and economic burden of treatment-resistant depression in Europe: A cross-sectional study. BMC Psychiatry 2019, 19, 247. [Google Scholar] [CrossRef] [Green Version]
- Paris, J. The mistreatment of major depressive disorder. Can. J. Psychiatry. Rev. Can. Psychiatr. 2014, 59, 148–151. [Google Scholar] [CrossRef] [Green Version]
- Pennisi, E. Meet the psychobiome. Science 2020, 368, 570–573. [Google Scholar] [CrossRef]
- Matarazzo, I.; Toniato, E.; Robuffo, I. Psychobiome Feeding Mind: Polyphenolics in Depression and Anxiety. Curr. Top. Med. Chem. 2018, 18, 2108–2115. [Google Scholar] [CrossRef]
- Tian, P.; Zou, R.; Song, L.; Zhang, X.; Jiang, B.; Wang, G.; Lee, Y.K.; Zhao, J.; Zhang, H.; Chen, W. Ingestion of Bifidobacterium longum subspecies infantis strain CCFM687 regulated emotional behavior and the central BDNF pathway in chronic stress-induced depressive mice through reshaping the gut microbiota. Food Funct. 2019, 10, 7588–7598. [Google Scholar] [CrossRef]
- Ait-Belgnaoui, A.; Durand, H.; Cartier, C.; Chaumaz, G.; Eutamene, H.; Ferrier, L.; Houdeau, E.; Fioramonti, J.; Bueno, L.; Theodorou, V. Prevention of gut leakiness by a probiotic treatment leads to attenuated HPA response to an acute psychological stress in rats. Psychoneuroendocrinology 2012, 37, 1885–1895. [Google Scholar] [CrossRef] [PubMed]
- Bercik, P.; Verdu, E.F.; Foster, J.A.; Macri, J.; Potter, M.; Huang, X.; Malinowski, P.; Jackson, W.; Blennerhassett, P.; Neufeld, K.A.; et al. Chronic gastrointestinal inflammation induces anxiety-like behavior and alters central nervous system biochemistry in mice. Gastroenterology 2010, 139, 2102–2112.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strandwitz, P. Neurotransmitter modulation by the gut microbiota. Brain Res. 2018, 1693, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Ozogul, F.; Kuley, E.; Ozogul, Y.; Ozogul, I. The Function of Lactic Acid Bacteria on Biogenic Amines Production by Food-Borne Pathogens in Arginine Decarboxylase Broth. Food Sci. Technol. Res. 2012, 18, 795–804. [Google Scholar] [CrossRef] [Green Version]
- Stanaszek, P.M.; Snell, J.F.; Neill, J.J.O. Isolation, extraction, and measurement of acetylcholine from Lactobacillus plantarum. Appl. Environ. Microbiol. 1977, 34, 237–239. [Google Scholar] [CrossRef] [Green Version]
- Barrett, E.; Ross, R.P.; O’Toole, P.W.; Fitzgerald, G.F.; Stanton, C. γ-Aminobutyric acid production by culturable bacteria from the human intestine. J. Appl. Microbiol. 2012, 113, 411–417. [Google Scholar] [CrossRef]
- Tsavkelova, E.A.; Botvinko, I.V.; Kudrin, V.S.; Oleskin, A.V. Detection of neurotransmitter amines in microorganisms with the use of high-performance liquid chromatography. Dokl. Biochem. Proc. Acad. Sci. USSR Biochem. Sect. 2000, 372, 115–117. [Google Scholar]
- Swann, J.R.; Want, E.J.; Geier, F.M.; Spagou, K.; Wilson, I.D.; Sidaway, J.E.; Nicholson, J.K.; Holmes, E. Systemic gut microbial modulation of bile acid metabolism in host tissue compartments. Proc. Natl. Acad. Sci. USA 2011, 108, 4523–4530. [Google Scholar] [CrossRef] [Green Version]
- Quinn, M.; McMillin, M.; Galindo, C.; Frampton, G.; Pae, H.Y.; DeMorrow, S. Bile acids permeabilize the blood brain barrier after bile duct ligation in rats via Rac1-dependent mechanisms. Dig. Liver Dis. Off. J. Ital. Soc. Gastroenterol. Ital. Assoc. Study Liver 2014, 46, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Goyal, D.; Ali, S.A.; Singh, R.K. Emerging role of gut microbiota in modulation of neuroinflammation and neurodegeneration with emphasis on Alzheimer’s disease. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 106, 110112. [Google Scholar] [CrossRef]
- Karakula-Juchnowicz, H.; Rog, J.; Juchnowicz, D.; Łoniewski, I.; Skonieczna-Żydecka, K.; Krukow, P.; Futyma-Jedrzejewska, M.; Kaczmarczyk, M. The study evaluating the effect of probiotic supplementation on the mental status, inflammation, and intestinal barrier in major depressive disorder patients using gluten-free or gluten-containing diet (SANGUT study): A 12-week, randomized, double-blind, and placebo-controlled clinical study protocol. Nutr. J. 2019, 18, 50. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.C.; Cassidy, A.; Willett, W.C.; Rimm, E.B.; O’Reilly, E.J.; Okereke, O.I. Dietary flavonoid intake and risk of incident depression in midlife and older women. Am. J. Clin. Nutr. 2016, 104, 704–714. [Google Scholar] [CrossRef] [Green Version]
- Mihrshahi, S.; Dobson, A.J.; Mishra, G.D. Fruit and vegetable consumption and prevalence and incidence of depressive symptoms in mid-age women: Results from the Australian longitudinal study on women’s health. Eur. J. Clin. Nutr. 2015, 69, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Pase, M.P.; Scholey, A.B.; Pipingas, A.; Kras, M.; Nolidin, K.; Gibbs, A.; Wesnes, K.; Stough, C. Cocoa polyphenols enhance positive mood states but not cognitive performance: A randomized, placebo-controlled trial. J. Psychopharmacol. 2013, 27, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Caracci, F.; Harary, J.; Simkovic, S.; Pasinetti, G.M. Grape-Derived Polyphenols Ameliorate Stress-Induced Depression by Regulating Synaptic Plasticity. J. Agric. Food Chem. 2020, 68, 1808–1815. [Google Scholar] [CrossRef]
- Lamport, D.J.; Pal, D.; Macready, A.L.; Barbosa-Boucas, S.; Fletcher, J.M.; Williams, C.M.; Spencer, J.P.; Butler, L.T. The effects of flavanone-rich citrus juice on cognitive function and cerebral blood flow: An acute, randomised, placebo-controlled cross-over trial in healthy, young adults. Br. J. Nutr. 2016, 116, 2160–2168. [Google Scholar] [CrossRef] [Green Version]
- Kean, R.J.; Lamport, D.J.; Dodd, G.F.; Freeman, J.E.; Williams, C.M.; Ellis, J.A.; Butler, L.T.; Spencer, J.P. Chronic consumption of flavanone-rich orange juice is associated with cognitive benefits: An 8-wk, randomized, double-blind, placebo-controlled trial in healthy older adults. Am. J. Clin. Nutr. 2015, 101, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Alharbi, M.H.; Lamport, D.J.; Dodd, G.F.; Saunders, C.; Harkness, L.; Butler, L.T.; Spencer, J.P. Flavonoid-rich orange juice is associated with acute improvements in cognitive function in healthy middle-aged males. Eur. J. Nutr. 2016, 55, 2021–2029. [Google Scholar] [CrossRef] [Green Version]
- Park, K.; Jaekal, E.; Yoon, S.; Lee, S.-H.; Choi, K.-H. Diagnostic Utility and Psychometric Properties of the Beck Depression Inventory-II among Korean Adults. Front. Psychol. 2020, 10, 2934. [Google Scholar] [CrossRef] [PubMed]
- Helmreich, I.; Wagner, S.; König, J.; Kohnen, R.; Szegedi, A.; Hiemke, C.; Tadić, A. Hamilton depression rating subscales to predict antidepressant treatment outcome in the early course of treatment. J. Affect. Disord. 2015, 175, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Radloff, L.S. The CES-D Scale: A Self-Report Depression Scale for Research in the General Population. Appl. Psychol. Meas. 1977, 1, 385–401. [Google Scholar] [CrossRef]
- Beck, A.T.; Steer, R.A.; Ball, R.; Ranieri, W. Comparison of Beck Depression Inventories-IA and -II in psychiatric outpatients. J. Personal. Assess. 1996, 67, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.J.; Kim, K.H. Use of the Center for Epidemiologic Studies Depression (CES-D) Scale in Korea. J. Nerv. Ment. Dis. 1998, 186, 304–310. [Google Scholar] [CrossRef]
- Lee, K.H.; Jae, Y.M.; Choi, J.H.; Jang, S.H. Initial Response to Medication Predicts Early Improvement after 2 Weeks in Patients with Major Depressive Disorder. J. Korean Soc. Dep. Bip. Disord. 2014, 12, 37–42. [Google Scholar]
- Park, M.; Choi, J.; Lee, H.J. Flavonoid-Rich Orange Juice Intake and Altered Gut Microbiome in Young Adults with Depressive Symptom: A Randomized Controlled Study. Nutrients 2020, 12, 1815. [Google Scholar] [CrossRef] [PubMed]
- Estaki, M.; Jiang, L.; Bokulich, N.A.; McDonald, D.; González, A.; Kosciolek, T.; Martino, C.; Zhu, Q.; Birmingham, A.; Vázquez-Baeza, Y.; et al. QIIME 2 Enables Comprehensive End-to-End Analysis of Diverse Microbiome Data and Comparative Studies with Publicly Available Data. Curr. Protoc. Bioinform. 2020, 70, e100. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Kuma, K.-I.; Toh, H.; Miyata, T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res 2005, 33, 511–518. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree: Computing large minimum evolution trees with profiles instead of a distance matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Grases, G.; Colom, M.A.; Sanchis, P.; Grases, F. Possible relation between consumption of different food groups and depression. BMC Psychol. 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ljungberg, T.; Bondza, E.; Lethin, C. Evidence of the Importance of Dietary Habits Regarding Depressive Symptoms and Depression. Int. J. Environ. Res. Public Health 2020, 17, 1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Lv, M.R.; Wei, Y.J.; Sun, L.; Zhang, J.X.; Zhang, H.G.; Li, B. Dietary patterns and depression risk: A meta-analysis. Psychiatry Res. 2017, 253, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Hintikka, J.; Tolmunen, T.; Honkalampi, K.; Haatainen, K.; Koivumaa-Honkanen, H.; Tanskanen, A.; Viinamäki, H. Daily Tea Drinking Is Associated with a Low Level of Depressive Symptoms in the Finnish General Population. Eur. J. Epidemiol. 2005, 20, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Logan, A.C. Omega-3 fatty acids and major depression: A primer for the mental health professional. Lipids Health Dis. 2004, 3, 25. [Google Scholar] [CrossRef] [Green Version]
- Khalid, S.; Barfoot, K.L.; May, G.; Lamport, D.J.; Reynolds, S.A.; Williams, C.M. Effects of Acute Blueberry Flavonoids on Mood in Children and Young Adults. Nutrients 2017, 9, 158. [Google Scholar] [CrossRef] [PubMed]
- Firoozabadi, A.; Kolouri, S.; Zarshenas, M.M.; Salehi, A.; Mosavat, S.H.; Dastgheib, S.A. Efficacy of Nepeta Menthoides Boiss and Buhse Freeze-Dried Aqueous Extract on Anxiety of Patients with Depression: A Double-Blind Randomized Controlled Clinical Trial. Iran. J. Med. Sci. 2016, 41, S4. [Google Scholar] [CrossRef]
- Macready, A.L.; Kennedy, O.B.; Ellis, J.A.; Williams, C.M.; Spencer, J.P.; Butler, L.T. Flavonoids and cognitive function: A review of human randomized controlled trial studies and recommendations for future studies. Genes Nutr. 2009, 4, 227–242. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.G.; Shukitt-Hale, B. Berry fruit enhances beneficial signaling in the brain. J. Agric. Food Chem. 2012, 60, 5709–5715. [Google Scholar] [CrossRef] [PubMed]
- Pipingas, A.; Silberstein, R.B.; Vitetta, L.; Rooy, C.V.; Harris, E.V.; Young, J.M.; Frampton, C.M.; Sali, A.; Nastasi, J. Improved cognitive performance after dietary supplementation with a Pinus radiata bark extract formulation. Phytother. Res. PTR 2008, 22, 1168–1174. [Google Scholar] [CrossRef] [PubMed]
- van Praag, H.; Lucero, M.J.; Yeo, G.W.; Stecker, K.; Heivand, N.; Zhao, C.; Yip, E.; Afanador, M.; Schroeter, H.; Hammerstone, J.; et al. Plant-derived flavanol (-)epicatechin enhances angiogenesis and retention of spatial memory in mice. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 5869–5878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Field, D.T.; Williams, C.M.; Butler, L.T. Consumption of cocoa flavanols results in an acute improvement in visual and cognitive functions. Physiol. Behav. 2011, 103, 255–260. [Google Scholar] [CrossRef] [Green Version]
- Scholey, A.B.; French, S.J.; Morris, P.J.; Kennedy, D.O.; Milne, A.L.; Haskell, C.F. Consumption of cocoa flavanols results in acute improvements in mood and cognitive performance during sustained mental effort. J. Psychopharmacol. 2010, 24, 1505–1514. [Google Scholar] [CrossRef]
- Cassidy, A.; Minihane, A.M. The role of metabolism (and the microbiome) in defining the clinical efficacy of dietary flavonoids. Am. J. Clin. Nutr. 2017, 105, 10–22. [Google Scholar] [CrossRef] [Green Version]
- Tomás-Barberán, F.A.; Selma, M.V.; Espín, J.C. Interactions of gut microbiota with dietary polyphenols and consequences to human health. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 471–476. [Google Scholar] [CrossRef]
- Cueva, C.; Gil-Sánchez, I.; Ayuda-Durán, B.; González-Manzano, S.; González-Paramás, A.M.; Santos-Buelga, C.; Bartolomé, B.; Moreno-Arribas, M.V. An Integrated View of the Effects of Wine Polyphenols and Their Relevant Metabolites on Gut and Host Health. Molecules 2017, 22, 99. [Google Scholar] [CrossRef]
- O’Mahony, S.M.; Clarke, G.; Borre, Y.E.; Dinan, T.G.; Cryan, J.F. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav. Brain Res. 2015, 277, 32–48. [Google Scholar] [CrossRef]
- Jenkins, T.A.; Nguyen, J.C.; Polglaze, K.E.; Bertrand, P.P. Influence of Tryptophan and Serotonin on Mood and Cognition with a Possible Role of the Gut-Brain Axis. Nutrients 2016, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Dash, S.; Clarke, G.; Berk, M.; Jacka, F.N. The gut microbiome and diet in psychiatry: Focus on depression. Curr. Opin. Psychiatry 2015, 28, 1–6. [Google Scholar] [CrossRef]
- Kowiański, P.; Lietzau, G.; Czuba, E.; Waśkow, M.; Steliga, A.; Moryś, J. BDNF: A Key Factor with Multipotent Impact on Brain Signaling and Synaptic Plasticity. Cell. Mol. Neurobiol. 2018, 38, 579–593. [Google Scholar] [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids from Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flint, H.J.; Duncan, S.H.; Scott, K.P.; Louis, P. Links between diet, gut microbiota composition and gut metabolism. Proc. Nutr. Soc. 2014, 74, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, J.; Redondo-Blanco, S.; Gutiérrez-del-Río, I.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Colon microbiota fermentation of dietary prebiotics towards short-chain fatty acids and their roles as anti-inflammatory and antitumour agents: A review. J. Funct. Foods 2016, 25, 511–522. [Google Scholar] [CrossRef]
- Levy, M.; Thaiss, C.A.; Elinav, E. Metabolites: Messengers between the microbiota and the immune system. Genes Dev. 2016, 30, 1589–1597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erny, D.; Hrabě de Angelis, A.L.; Jaitin, D.; Wieghofer, P.; Staszewski, O.; David, E.; Keren-Shaul, H.; Mahlakoiv, T.; Jakobshagen, K.; Buch, T.; et al. Host microbiota constantly control maturation and function of microglia in the CNS. Nat. Neurosci. 2015, 18, 965–977. [Google Scholar] [CrossRef] [PubMed]
- El-Gedaily, A.; Paesold, G.; Chen, C.Y.; Guiney, D.G.; Krause, M. Plasmid virulence gene expression induced by short-chain fatty acids in Salmonella dublin: Identification of rpoS-dependent and rpo-S-independent mechanisms. J. Bacteriol. 1997, 179, 1409–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakkarach, A.; Foo, H.L.; Song, A.A.-L.; Mutalib, N.E.A.; Nitisinprasert, S.; Withayagiat, U. Anti-cancer and anti-inflammatory effects elicited by short chain fatty acids produced by Escherichia coli isolated from healthy human gut microbiota. Microb. Cell Factories 2021, 20, 36. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Fan, C.; Li, P.; Lu, Y.; Chang, X.; Qi, K. Short Chain Fatty Acids Prevent High-fat-diet-induced Obesity in Mice by Regulating G Protein-coupled Receptors and Gut Microbiota. Sci. Rep. 2016, 6, 37589. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; van Esch, B.C.A.M.; Henricks, P.A.J.; Folkerts, G.; Garssen, J. The Anti-inflammatory Effects of Short Chain Fatty Acids on Lipopolysaccharide- or Tumor Necrosis Factor α-Stimulated Endothelial Cells via Activation of GPR41/43 and Inhibition of HDACs. Front. Pharmacol. 2018, 9, 533. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Kim, M.; Kang, S.G.; Jannasch, A.H.; Cooper, B.; Patterson, J.; Kim, C.H. Short-chain fatty acids induce both effector and regulatory T cells by suppression of histone deacetylases and regulation of the mTOR–S6K pathway. Mucosal Immunol. 2015, 8, 80–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luu, M.; Pautz, S.; Kohl, V.; Singh, R.; Romero, R.; Lucas, S.; Hofmann, J.; Raifer, H.; Vachharajani, N.; Carrascosa, L.C.; et al. The short-chain fatty acid pentanoate suppresses autoimmunity by modulating the metabolic-epigenetic crosstalk in lymphocytes. Nat. Commun. 2019, 10, 760. [Google Scholar] [CrossRef] [Green Version]
- Stevens, B.R.; Goel, R.; Seungbum, K.; Richards, E.M.; Holbert, R.C.; Pepine, C.J.; Raizada, M.K. Increased human intestinal barrier permeability plasma biomarkers zonulin and FABP2 correlated with plasma LPS and altered gut microbiome in anxiety or depression. Gut 2018, 67, 1555–1557. [Google Scholar] [CrossRef]
- Eeckhaut, V.; Van Immerseel, F.; Teirlynck, E.; Pasmans, F.; Fievez, V.; Snauwaert, C.; Haesebrouck, F.; Ducatelle, R.; Louis, P.; Vandamme, P. Butyricicoccus pullicaecorum gen. nov., sp. nov., an anaerobic, butyrate-producing bacterium isolated from the caecal content of a broiler chicken. Int. J. Syst. Evol. Microbiol. 2008, 58, 2799–2802. [Google Scholar] [CrossRef]
- Boesmans, L.; Valles-Colomer, M.; Wang, J.; Eeckhaut, V.; Falony, G.; Ducatelle, R.; Immerseel, F.V.; Raes, J.; Verbeke, K.; Cotter, P.D. Butyrate Producers as Potential Next-Generation Probiotics: Safety Assessment of the Administration of Butyricicoccus pullicaecorum to Healthy Volunteers. mSystems 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Raina, S.K. Limitations of 24-hour Recall Method: Micronutrient Intake and the Presence of the Metabolic Syndrome. N. Am. J. Med. Sci. 2013, 5, 498. [Google Scholar] [CrossRef] [Green Version]
- Tang, Q.; Jin, G.; Wang, G.; Liu, T.; Liu, X.; Wang, B.; Cao, H. Current sampling methods for gut microbiota: A call for more precise devices. Front. Cell. Infect. Microbiol. 2020, 10, 151. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
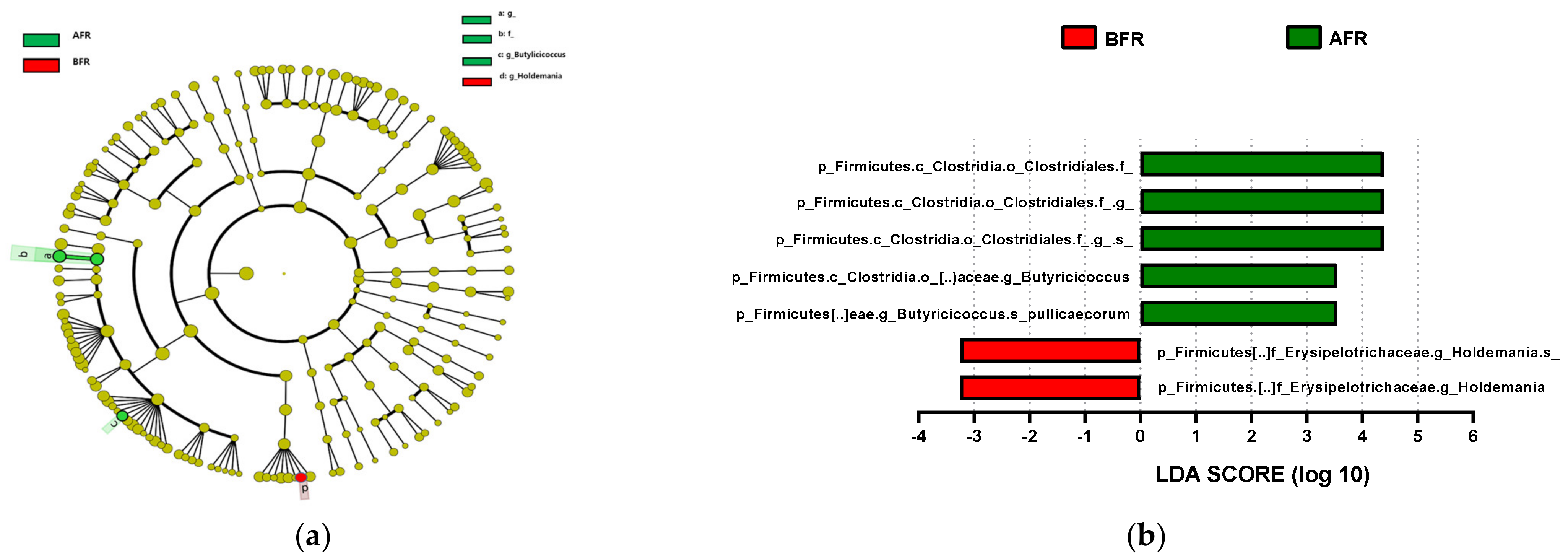{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | FR Group (n = 20) | FL Group (n = 20) | p-Value |
|---|---|---|---|
| Sex, N (%) | |||
| Male | 6 (30) | 6 (30) | 1.000 ‡ |
| Female | 14 (70) | 14 (70) | |
| Age, mean ± SD, year | 24.70 ± 2.830 | 23.10 ± 2.594 | 0.070 † |
| Residence type, N (%) | |||
| Living alone | 7 (35) | 6 (30) | 0.427 $ |
| With roommate | 1 (5) | 5 (25) | |
| With parents | 9 (45) | 6 (30) | |
| With parents and grandparents | 2 (10) | 1 (5) | |
| Others | 1 (5) | 2 (10) | |
| Household income, N (%) | |||
| <1,000,000 won per month | 1 (5) | 4 (20) | 0.227 $ |
| 1,000,000–2,999,999 won per month | 6 (30) | 7 (35) | |
| 3,000,000–4,999,999 won per month | 5 (25) | 1 (5) | |
| >5,000,000 won per month | 8 (40) | 8 (40) | |
| Regular physical activity, N (%) | |||
| Yes | 5 (25) | 5 (25) | 1.000 ‡ |
| No | 15 (75) | 15 (75) | |
| Alcohol drinking, N (%) | |||
| None | 4 (20) | 1 (5) | 0.511 $ |
| Less than once a month | 4 (20) | 4 (20) | |
| Once a month | 3 (15) | 6 (30) | |
| 2–4 times a month | 3 (15) | 5 (25) | |
| 2–3 times a week | 6 (30) | 4 (20) | |
| Heavy drinking frequency, N (%) | |||
| None | 11 (55) | 5 (25) | 0.030 $ |
| Less than once a month | 4 (20) | 6 (30) | |
| Once a month | 1 (5) | 7 (35) | |
| Once a week | 4 (20) | 1 (5) | |
| Almost everyday | 0 | 1 (5) | |
| Smoking, N (%) | |||
| None | 15 (75) | 11 (55) | 0.587 $ |
| Past smoking (no current smoking) | 1 (5) | 3 (15) | |
| Occasionally | 1 (5) | 1 (5) | |
| Always | 3 (15) | 5 (25) | |
| Stress, N (%) | |||
| Too much | 4 (20) | 4 (20) | 1.000 $ |
| Pretty much | 12 (60) | 11 (55) | |
| Little | 4 (20) | 5 (25) | |
| Variables | FR Group (n = 20) | p-Value | FL Group (n = 20) | p-Value | Δ Group Comparison $ | ||
|---|---|---|---|---|---|---|---|
| Baseline | After Intervention | Baseline | After Intervention | ||||
| Mean ± SE | Mean ± SE | ||||||
| Weight, kg | 66.33 ± 3.09 | 67.53 ± 3.13 | 0.001 † | 59.70 ± 2.91 | 60.62 ± 2.95 | 0.007 † | 0.512 |
| BMI, kg/m2 | 23.85 ± 0.80 | 24.29 ± 0.80 | 0.002 † | 22.00 ± 0.86 | 22.32 ± 0.87 | 0.016 † | 0.449 |
| Percent body fat, % | 31.42 ± 1.41 | 31.07 ± 1.40 | 0.737 ‡ | 28.48 ± 1.97 | 27.93 ± 1.81 | 0.338 ‡ | 0.754 |
| Skeletal muscle mass, kg | 24.87 ± 1.33 | 24.63 ± 1.43 | 0.376 ‡ | 23.16 ± 1.08 | 23.73 ± 1.09 | 0.285 ‡ | 0.346 |
| SBP, mmHg | 119.95 ± 3.39 | 120.00 ± 2.19 | 0.977 † | 113.21 ± 2.26 | 114.95 ± 2.14 | 0.524 † | 0.605 |
| DBP, mmHg | 76.47 ± 2.41 | 81.68 ± 1.83 | 0.064 † | 75.53 ± 2.75 | 77.21 ± 2.29 | 0.475 † | 0.321 |
| BDNF | 2033.61 ± 9.16 | 2157.80 ± 19.67 | 0.000 † | 2123.93 ± 16.13 | 2133.75 ± 9.87 | 0.670 † | 0.001 |
| Serotonin, ng/mL | 90.65 ± 10.98 | 111.24 ± 14.06 | 0.268 † | 94.64 ± 10.69 | 98.43 ± 14.78 | 0.846 † | 0.528 |
| Folate, ng/mL | 8.65 ± 1.24 | 9.03 ± 1.06 | 0.630 † | 7.50 ± 1.28 | 7.24 ± 0.84 | 0.970 ‡ | 0.646 |
| Homocysteine, μmol/L | 9.01 ± 0.92 | 8.31 ± 0.78 | 0.107 † | 8.73 ± 0.62 | 7.70 ± 0.51 | 0.090 † | 0.638 |
| hs-CRP, mg/L | 1.98 ± 0.69 | 1.03 ± 0.22 | 0.842 ‡ | 0.65 ± 1.67 | 2.21 ± 1.47 | 0.395 ‡ | 0.122 |
| Vitamin B12, pg/Ml | 624.70 ± 45.41 | 607.40 ± 54.55 | 0.639 † | 617.45 ± 87.21 | 609.15 ± 73.82 | 0.365 ‡ | 0.840 |
| BDI-II | 26.05 ± 1.38 | 14.10 ± 1.54 | <0.001 † | 26.55 ± 1.18 | 17.05 ± 1.33 | <0.001 † | 0.360 |
| CES-D score | 34.25 ± 2.06 | 15.90 ± 2.00 | <0.001 † | 30.30 ± 1.82 | 18.15 ± 2.24 | 0.000 † | 0.110 |
| HAMD-17 | 21.30 ± 1.11 | 8.30 ±1.29 | <0.001 † | 21.80 ± 1.05 | 12.40 ± 1.32 | <0.001 † | 0.114 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, J.; Kim, J.-H.; Park, M.; Lee, H.-J. Effects of Flavonoid-Rich Orange Juice Intervention on Major Depressive Disorder in Young Adults: A Randomized Controlled Trial. Nutrients 2023, 15, 145. https://doi.org/10.3390/nu15010145
Choi J, Kim J-H, Park M, Lee H-J. Effects of Flavonoid-Rich Orange Juice Intervention on Major Depressive Disorder in Young Adults: A Randomized Controlled Trial. Nutrients. 2023; 15(1):145. https://doi.org/10.3390/nu15010145
Chicago/Turabian StyleChoi, Jihee, Jong-Hoon Kim, Miey Park, and Hae-Jeung Lee. 2023. "Effects of Flavonoid-Rich Orange Juice Intervention on Major Depressive Disorder in Young Adults: A Randomized Controlled Trial" Nutrients 15, no. 1: 145. https://doi.org/10.3390/nu15010145
APA StyleChoi, J., Kim, J. -H., Park, M., & Lee, H. -J. (2023). Effects of Flavonoid-Rich Orange Juice Intervention on Major Depressive Disorder in Young Adults: A Randomized Controlled Trial. Nutrients, 15(1), 145. https://doi.org/10.3390/nu15010145