
Whey Protein Hydrolysate Ameliorated High-Fat-Diet Induced Bone Loss via Suppressing Oxidative Stress and Regulating GSK-3β/Nrf2 Signaling Pathway


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Preparation of Whey Protein Hydrolysate (WPH)
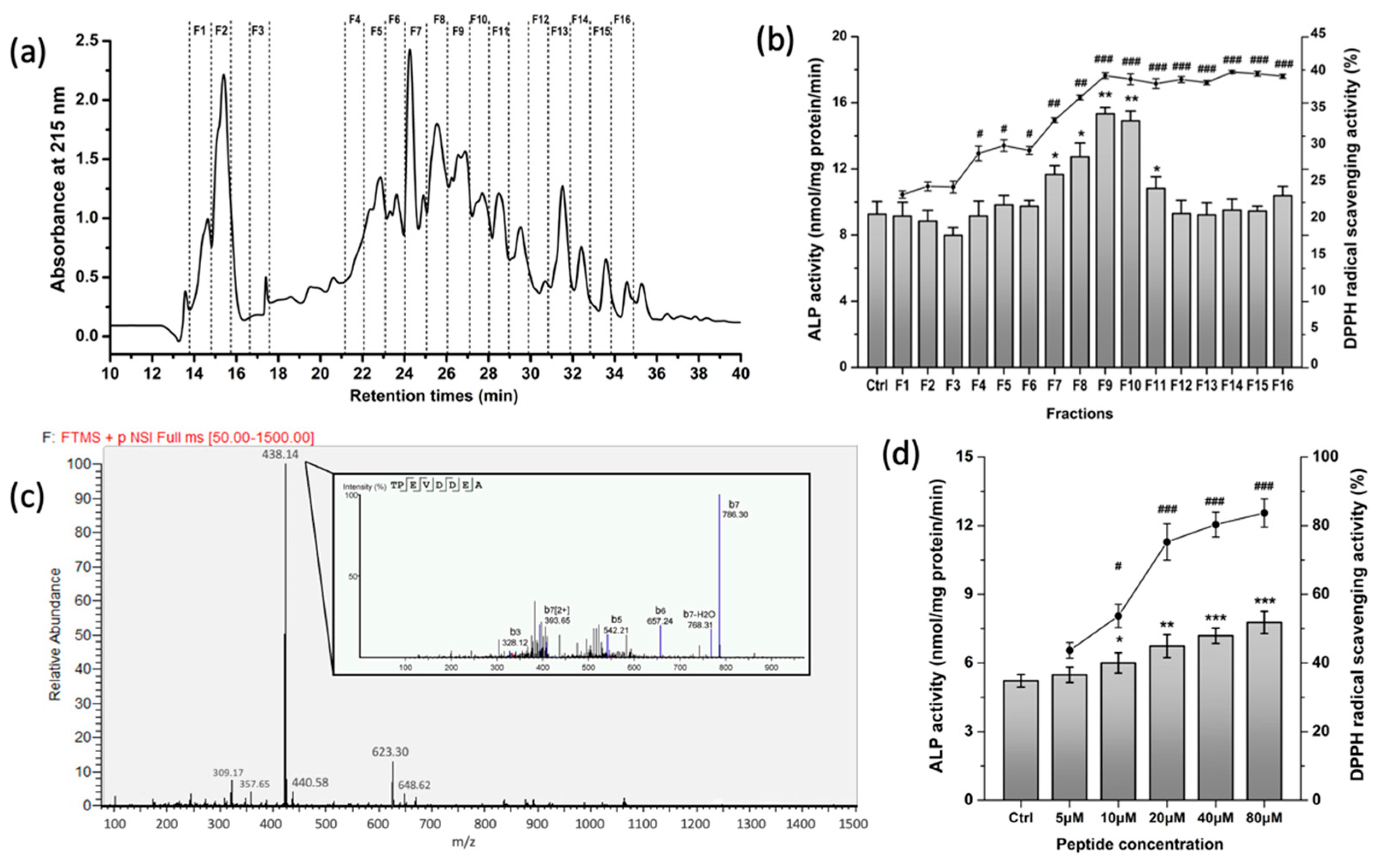
2.3. Characterization of the Peptide Sequences
2.4. Alkaline Phosphatase (ALP) Activity Assay
2.5. Capacity of Scavenging DPPH Radicals
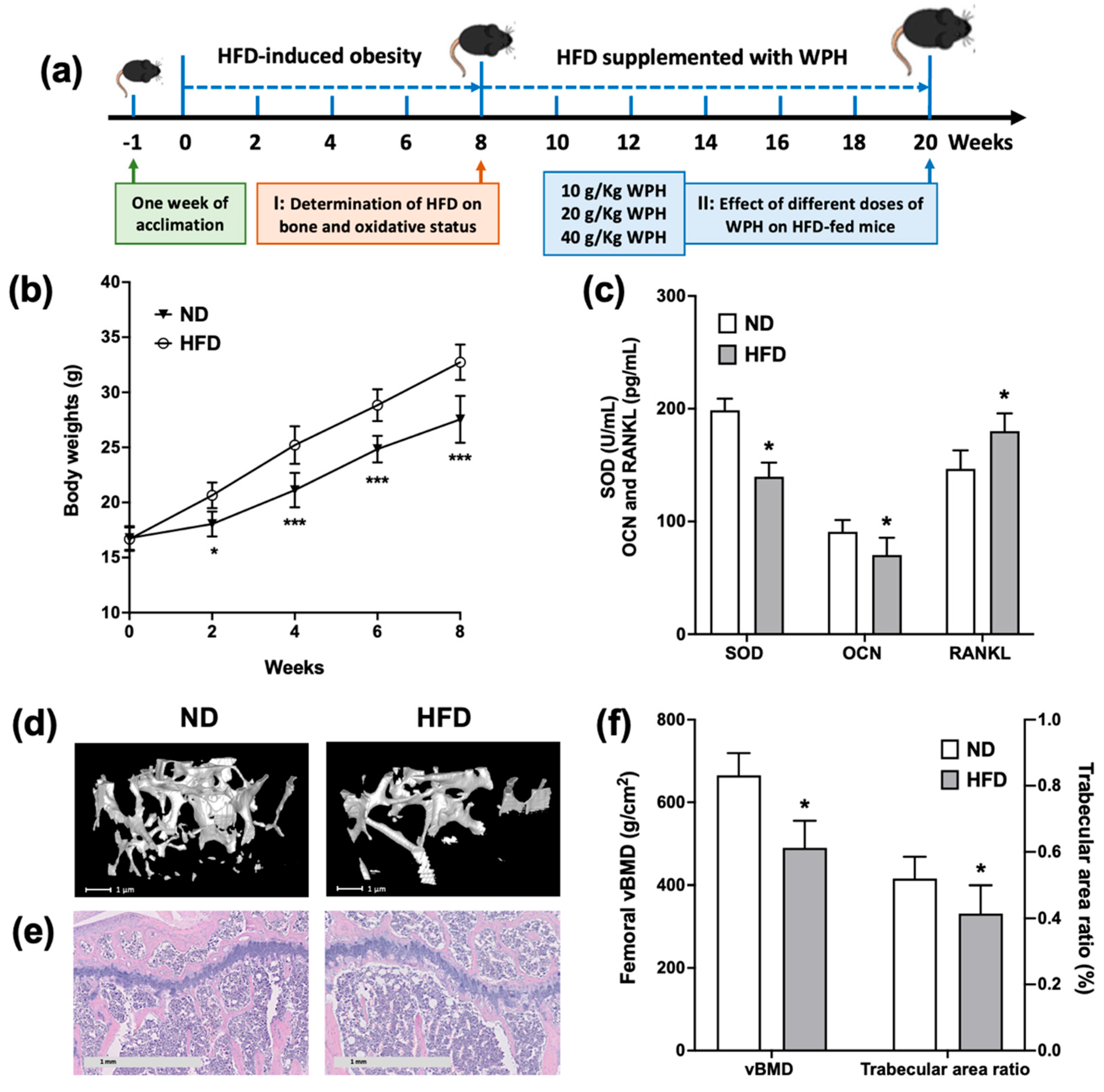
2.6. Animals and Treatment
2.7. Sample Collection
2.8. Serum Lipid and Glucose Status
2.9. Mechanical Testing and Bone Composition
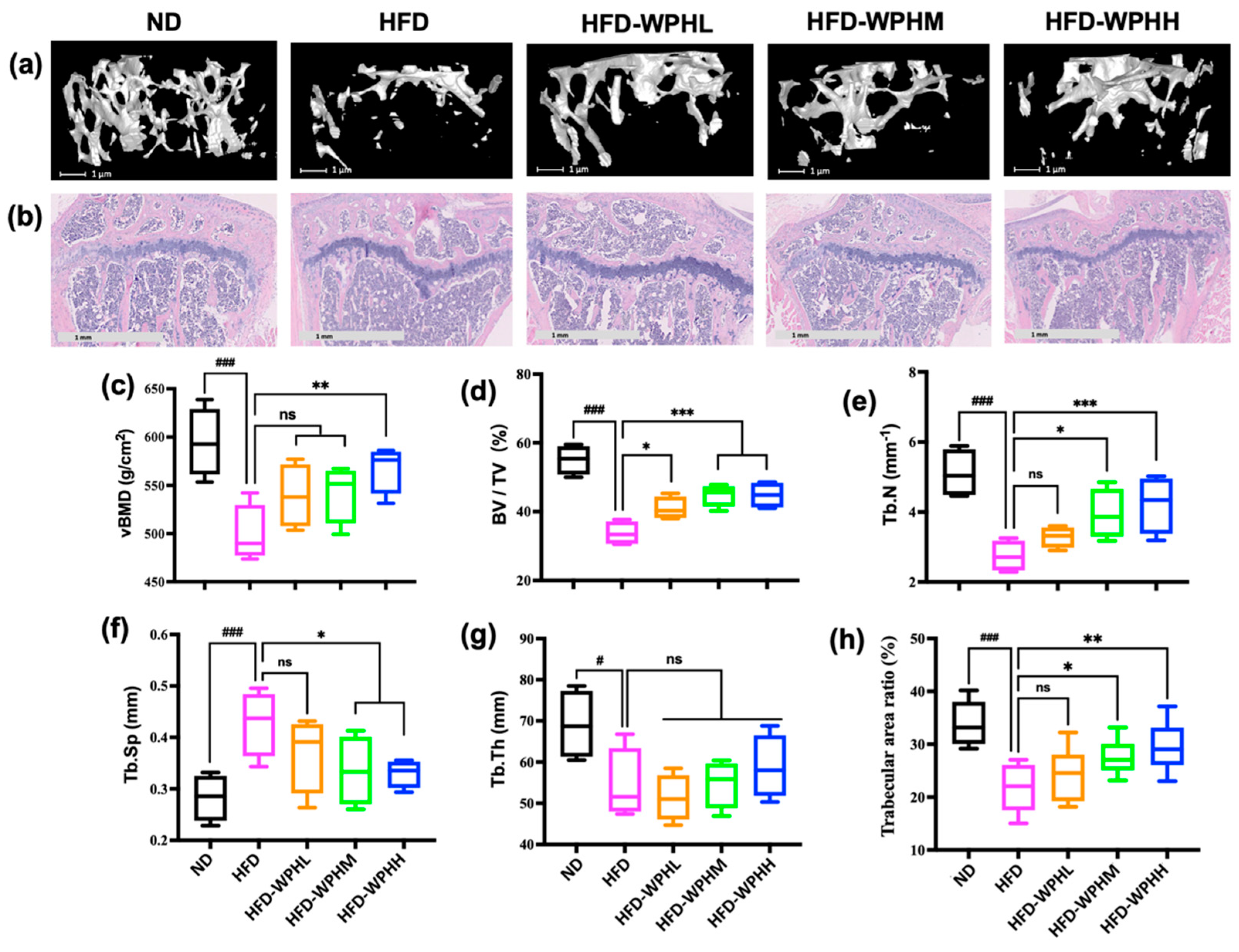
2.10. Micro-CT Analysis
2.11. H&E Staining
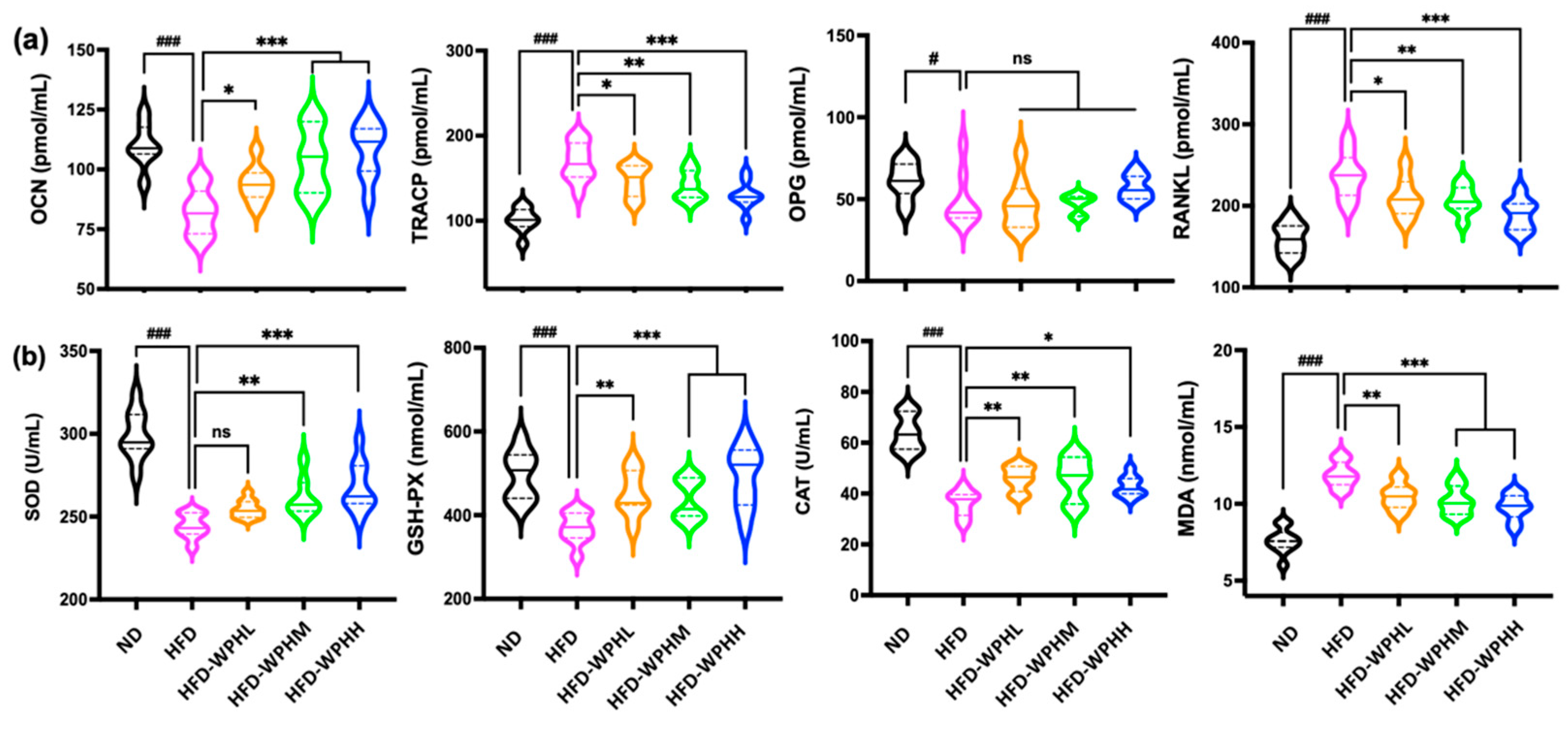
2.12. Bone Metabolism Biomarkers and Antioxidant Enzymes in Serum
2.13. Western Blotting
2.14. Statistical Analysis
3. Results
3.1. Purification and Characterization of Osteogenic and Antioxidative Peptides from WPH
3.2. HFD-Induced Obese Mice Exhibited Bone Loss and Oxidative Damages
3.3. WPH Restored Fat Mass and Improved Serum Lipid Levels in HFD-Fed Mice
3.4. WPH Improved Bone Minerals and Mechanical Properties in HFD-Fed Mice
3.5. WPH Repaired Bone Microstructure in HFD-Fed Mice
3.6. WPH Readjusted Bone Remodeling in HFD-Fed Mice
3.7. WPH Alleviated Oxidative Stress in HFD-Fed Mice
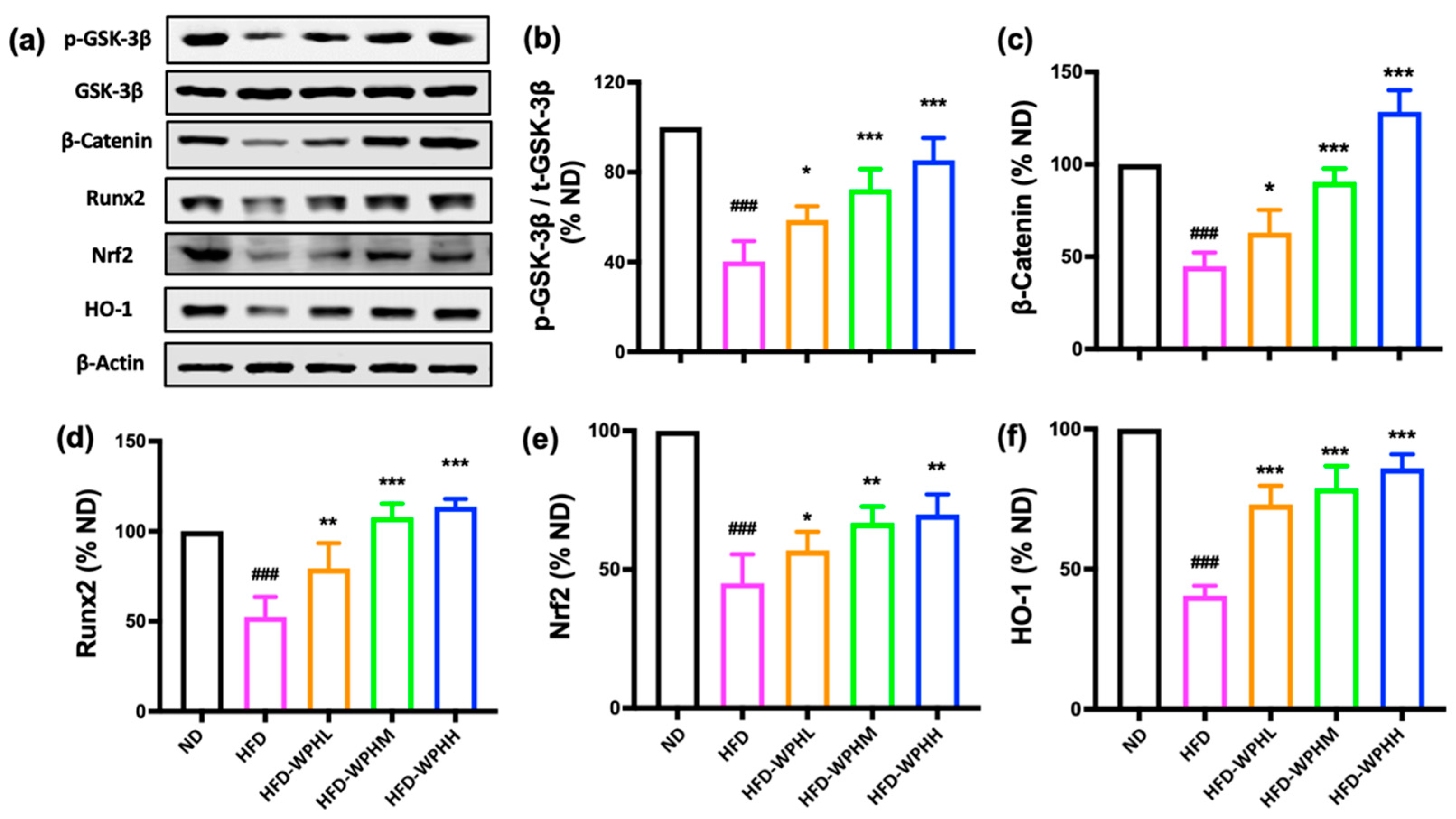
3.8. WPH Regulated GSK-3β/Nrf2/HO-1 in HFD-Fed Mice
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Longo, M.; Zatterale, F.; Naderi, J.; Parrillo, L.; Formisano, P.; Raciti, G.A.; Beguinot, F.; Miele, C. Adipose Tissue Dysfunction as Determinant of Obesity-Associated Metabolic figureComplications. Int. J. Mol. Sci. 2019, 20, 2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z. The metabolic syndrome. Lancet 2005, 365, 1415–1428. [Google Scholar] [CrossRef] [PubMed]
- Migliaccio, S.; Greco, E.A.; Fornari, R.; Donini, L.M.; Lenzi, A. Is obesity in women protective against osteoporosis? Diabetes Metab. Syndr. Obes. 2011, 4, 273–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, J.; Wu, Y.W.; Ren, Y.Z. The impact of a high fat diet on bones: Potential mechanisms. Food Funct. 2021, 12, 963–975. [Google Scholar] [CrossRef]
- Xiao, Y.; Cui, J.; Li, Y.X.; Shi, Y.H.; Wang, B.; Le, G.W.; Wang, Z.P. Dyslipidemic high-fat diet affects adversely bone metabolism in mice associated with impaired antioxidant capacity. Nutrition 2011, 27, 214–220. [Google Scholar] [CrossRef]
- Manna, P.; Jain, S.K. Obesity, Oxidative Stress, Adipose Tissue Dysfunction, and the Associated Health Risks: Causes and Therapeutic Strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444. [Google Scholar] [CrossRef] [Green Version]
- Tencerova, M.; Figeac, F.; Ditzel, N.; Taipaleenmaki, H.; Nielsen, T.K.; Kassem, M. High-Fat Diet-Induced Obesity Promotes Expansion of Bone Marrow Adipose Tissue and Impairs Skeletal Stem Cell Functions in Mice. J. Bone Miner. Res. 2018, 33, 1154–1165. [Google Scholar] [CrossRef] [Green Version]
- Guicheux, J.; Wittrant, Y.; Wauquier, F.; Léotoing, L.; Coxam, V. Oxidative stress in bone remodelling and disease. Trends Mol. Med. 2009, 15, 468–477. [Google Scholar]
- Shu, L.; Beier, E.; Sheu, T.; Zhang, H.W.; Zuscik, M.J.; Puzas, E.J.; Boyce, B.F.; Mooney, R.A.; Xing, L.P. High-Fat Diet Causes Bone Loss in Young Mice by Promoting Osteoclastogenesis through Alteration of the Bone Marrow Environment. Calcif. Tissue Int. 2015, 96, 313–323. [Google Scholar] [CrossRef] [Green Version]
- Chowdhry, S.; Zhang, Y.; McMahon, M.; Sutherland, C.; Cuadrado, A.; Hayes, J.D. Nrf2 is controlled by two distinct beta-TrCP recognition motifs in its Neh6 domain, one of which can be modulated by GSK-3 activity. Oncogene 2013, 32, 3765–3781. [Google Scholar] [CrossRef] [Green Version]
- Cuadrado, A. Structural and functional characterization of Nrf2 degradation by glycogen synthase kinase 3/beta-TrCP. Free. Radic. Biol. Med. 2015, 88, 147–157. [Google Scholar] [CrossRef]
- Ibanez, L.; Ferrandiz, M.L.; Brines, R.; Guede, D.; Cuadrado, A.; Alcaraz, M.J. Effects of Nrf2 Deficiency on Bone Microarchitecture in an Experimental Model of Osteoporosis. Oxidative Med. Cell. Longev. 2014, 2014, 726590. [Google Scholar] [CrossRef] [Green Version]
- Gaur, T.; Lengner, C.J.; Hovhannisyan, H.; Bhat, R.A.; Bodine, P.V.N.; Komm, B.S.; Javed, A.; van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; et al. Canonical WNT signaling promotes osteogenesis by directly stimulating Runx2 gene expression. J. Biol. Chem. 2005, 280, 33132–33140. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Ma, S.H.; Li, Y.R.; Sun, Y.J.; Zhang, K.; Zhou, Q.; Yu, R.Q. Evaluating the Activity of Sodium Butyrate to Prevent Osteoporosis in Rats by Promoting Osteal GSK-3 beta/Nrf2 Signaling and Mitochondrial Function. J. Agric. Food Chem. 2020, 68, 6588–6603. [Google Scholar] [CrossRef]
- Guo, D.J.; Zhao, M.G.; Xu, W.; He, H.; Li, B.; Hou, T. Dietary interventions for better management of osteoporosis: An overview. Crit. Rev. Food Sci. Nutr. 2023, 63, 125–144. [Google Scholar] [CrossRef]
- Shen, C.L.; Chanjaplammootil, S.; Yeh, J.K.; Cao, J.J.; Chyu, M.C.; Dagda, R.Y.; Wang, J.S. Anti-obesity and osteo-protective effect of green tea polyphenols on long-term high-fat-diet-induced obesity in rats. Faseb J. 2011, 25, 776–782. [Google Scholar]
- Qi, S.S.; He, J.; Han, H.; Zheng, H.X.; Jiang, H.; Hu, C.Y.; Zhang, Z.J.; Li, X.S. Anthocyanin-rich extract from black rice (Oryza sativa L. Japonica) ameliorates diabetic osteoporosis in rats. Food Funct. 2019, 10, 5350–5360. [Google Scholar] [CrossRef]
- Wang, Y.J.; Sun, J.H.; Zhang, Y.Q.; Liu, W.Z.; Yang, S.; Yang, J.F. Stichopus japonicus Polysaccharide Stimulates Osteoblast Differentiation through Activation of the Bone Morphogenetic Protein Pathway in MC3T3-E1 Cells. J. Agric. Food Chem. 2021, 69, 2576–2584. [Google Scholar] [CrossRef]
- Yue, H.; Bo, Y.Y.; Tian, Y.Y.; Mao, L.; Xue, C.H.; Dong, P.; Wang, J.F. Docosahexaenoic Acid-Enriched Phosphatidylcholine Exerted Superior Effects to Triglyceride in Ameliorating Obesity-Induced Osteoporosis through up-Regulating the Wnt/β–Catenin Pathway. J. Agric. Food Chem. 2022, 70, 13904–13912. [Google Scholar] [CrossRef]
- Bu, T.T.; Zheng, J.X.; Liu, L.; Li, S.S.; Wu, J.P. Milk proteins and their derived peptides on bone health: Biological functions, mechanisms, and prospects. Compr. Rev. Food Sci. Food Saf. 2021, 20, 2234–2262. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Tian, Y.Y.; Feng, X.X.; Bo, Y.Y.; Xue, C.H.; Dong, P.; Wang, J.F. Novel Peptides Derived from Sea Cucumber Intestine Promotes Osteogenesis by Upregulating Integrin-Mediated Transdifferentiation of Growth Plate Chondrocytes to Osteoblasts. J. Agric. Food Chem. 2022, 70, 13212–13222. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, S.S.; Zheng, J.X.; Bu, T.T.; He, G.Q.; Wu, J.P. Safety considerations on food protein-derived bioactive peptides. Trends Food Sci. Technol. 2020, 96, 199–207. [Google Scholar] [CrossRef]
- Madureira, A.R.; Tavares, T.; Gomes, A.M.P.; Pintado, M.E.; Malcata, F.X. Invited review: Physiological properties of bioactive peptides obtained from whey proteins. J. Dairy Sci. 2010, 93, 437–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, M.; Kapila, S.; Kapila, R.; Trivedi, R.; Karvande, A. Evaluation of the osteoprotective potential of whey derived-antioxidative (YVEEL) and angiotensin-converting enzyme inhibitory (YLLF) bioactive peptides in ovariectomised rats. Food Funct. 2018, 9, 4791–4801. [Google Scholar] [CrossRef] [PubMed]
- Bu, T.T.; Ren, Y.T.; Yu, S.F.; Zheng, J.X.; Liu, L.; Sun, P.L.; Wu, J.P.; Yang, K. A Low-Phenylalanine-Containing Whey Protein Hydrolysate Stimulates Osteogenic Activity through the Activation of p38/Runx2 Signaling in Osteoblast Cells. Nutrients 2022, 14, 3135. [Google Scholar] [CrossRef]
- Bu, T.T.; Zhou, M.J.; Zheng, J.X.; Yang, P.; Song, H.L.; Li, S.S.; Wu, J.P. Preparation and characterization of a low-phenylalanine whey hydrolysate using two-step enzymatic hydrolysis and macroporous resin adsorption. Lwt Food Sci. Technol. 2020, 132, 109753. [Google Scholar] [CrossRef]
- Bu, T.T.; Zhang, L.; Liu, L.; Yu, S.F.; Zheng, J.X.; Wu, J.P.; Yang, K. Evaluation of the anti-osteoporotic effect of a low-phenylalanine whey protein hydrolysate in an ovariectomized mice model. Food Funct. 2022, 13, 3957–3967. [Google Scholar] [CrossRef]
- Sawada, Y.; Sakamoto, Y.; Toh, M.; Ohara, N.; Hatanaka, Y.; Naka, A.; Kishimoto, Y.; Kondo, K.; Iida, K. Milk-derived peptide Val-Pro-Pro (VPP) inhibits obesity-induced adipose inflammation via an angiotensin-converting enzyme (ACE) dependent cascade. Mol. Nutr. Food Res. 2015, 59, 2502–2510. [Google Scholar] [CrossRef]
- Yoda, K.; Sun, X.; Kawase, M.; Kubota, A.; Miyazawa, K.; Harata, G.; Hosoda, M.; Hiramatsu, M.; He, F.; Zemel, M.B. A combination of probiotics and whey proteins enhances anti-obesity effects of calcium and dairy products during nutritional energy restriction in aP2-agouti transgenic mice. Br. J. Nutr. 2015, 113, 1689–1696. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.X.; Bu, T.T.; Liu, L.; He, G.Q.; Li, S.S.; Wu, J.P. Naturally occurring low molecular peptides identified in egg white show antioxidant activity. Food Res. Int. 2020, 138, 109766. [Google Scholar] [CrossRef]
- Vinholes, J.; Grosso, C.; Andrade, P.B.; Gil-Izquierdo, A.; Valentao, P.; Pinho, P.G.; Ferreres, F. In vitro studies to assess the antidiabetic, anti-cholinesterase and antioxidant potential of Spergularia rubra. Food Chem. 2011, 129, 454–462. [Google Scholar] [CrossRef]
- Brandelli, A.; Daroit, D.J.; Correa, A.P.F. Whey as a source of peptides with remarkable biological activities. Food Res. Int. 2015, 73, 149–161. [Google Scholar] [CrossRef] [Green Version]
- Jiang, B.; Na, J.; Wang, L.; Li, D.; Liu, C.; Feng, Z. Separation and Enrichment of Antioxidant Peptides from Whey Protein Isolate Hydrolysate by Aqueous Two-Phase Extraction and Aqueous Two-Phase Flotation. Foods 2019, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- Chougule, R.A.; Salimath, B.P.; Sosalegowda, A.H. Buffalo colostrum beta-lactoglobulin inhibits VEGF-induced angiogenesis by interacting with G protein-coupled receptor kinase. Appl. Biochem. Biotechnol. 2013, 171, 366–381. [Google Scholar] [CrossRef]
- Matsuzawa-Nagata, N.; Takamura, T.; Ando, H.; Nakamura, S.; Kurita, S.; Misu, H.; Ota, T.; Yokoyama, M.; Honda, M.; Miyamoto, K.; et al. Increased oxidative stress precedes the onset of high-fat diet-induced insulin resistance and obesity. Metabolism 2008, 57, 1071–1077. [Google Scholar] [CrossRef] [Green Version]
- Bittencourt, A.; Brum, P.O.; Ribeiro, C.T.; Gasparotto, J.; Bortolin, R.C.; de Vargas, A.R.; Heimfarth, L.; de Almeida, R.F.; Moreira, J.C.F.; de Oliveira, J.; et al. High fat diet-induced obesity causes a reduction in brain tyrosine hydroxylase levels and non-motor features in rats through metabolic dysfunction, neuroinflammation and oxidative stress. Nutr. Neurosci. 2022, 25, 1026–1040. [Google Scholar] [CrossRef]
- Rosen, C.J.; Bouxsein, M.L. Mechanisms of disease: Is osteoporosis the obesity of bone? Nat. Clin. Pract. Rheumatol. 2006, 2, 35–43. [Google Scholar] [CrossRef]
- Cortez, M.; Carmo, L.S.; Rogero, M.M.; Borelli, P.; Fock, R.A. A high-fat diet increases IL-1, IL-6, and TNF-alpha production by increasing NF–κB and attenuating PPAR-gamma expression in bone marrow mesenchymal stem cells. Inflammation 2013, 36, 379–386. [Google Scholar] [CrossRef]
- An, S.; Ling, J.; Gao, Y.; Xiao, Y. Effects of varied ionic calcium and phosphate on the proliferation, osteogenic differentiation and mineralization of human periodontal ligament cells in vitro. J. Periodontal Res. 2012, 47, 374–382. [Google Scholar] [CrossRef] [Green Version]
- Ashtar, M.; Tenshin, H.; Teramachi, J.; Bat-Erdene, A.; Hiasa, M.; Oda, A.; Tanimoto, K.; Shimizu, S.; Higa, Y.; Harada, T.; et al. The Roles of ROS Generation in RANKL-Induced Osteoclastogenesis: Suppressive Effects of Febuxostat. Cancers 2020, 12, 929. [Google Scholar] [CrossRef] [Green Version]
- Roy, B.; Curtis, M.E.; Fears, L.S.; Nahashon, S.N.; Fentress, H.M. Molecular Mechanisms of Obesity-Induced Osteoporosis and Muscle Atrophy. Front. Physiol. 2016, 7, 439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.K.; Chen, J.; Tie, H.Y.; Tian, W.; Zhao, Y.L.; Qin, L.Y.; Guo, S.Y.; Li, Q.F.; Bao, C. Eriodictyol regulated ferroptosis, mitochondrial dysfunction, and cell viability via Nrf2/HO-1/NQO1 signaling pathway in ovarian cancer cells. J. Biochem. Mol. Toxicol. 2023, 23368. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.X.; Wong, S.W.; Liang, K.X.; Wu, T.H.; Wang, S.; Wang, L.F.; Liu, J.; Yamauchi, M.; Foster, B.L.; Ting, J.P.; et al. Optineurin regulates NRF2-mediated antioxidant response in a mouse model of Paget’s disease of bone. Sci. Adv. 2023, 9, eade6998. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.R.; Zhu, X.B.; Wei, A.; Chen, F.; Gao, Q.; Lu, K.; Jiang, Q.; Cao, W.S. Nrf2 epigenetic derepression induced by running exercise protects against osteoporosis. Bone Res. 2021, 9, 15. [Google Scholar] [CrossRef]
- Rojo, A.I.; Sagarra, M.R.; Cuadrado, A. GSK-3beta down-regulates the transcription factor Nrf2 after oxidant damage: Relevance to exposure of neuronal cells to oxidative stress. J. Neurochem. 2008, 105, 192–202. [Google Scholar] [CrossRef]
- Zahoor, M.; Cha, P.H.; Choi, K.Y. Indirubin-3′–oxime, an activator of Wnt/β–catenin signaling, enhances osteogenic commitment of ST2 cells and restores bone loss in high-fat diet-induced obese male mice. Bone 2014, 65, 60–68. [Google Scholar] [CrossRef]
- Sun, X.L.; Xia, T.S.; Zhang, S.Y.; Zhang, J.B.; Xu, L.C.; Han, T.; Xin, H.L. Hops extract and xanthohumol ameliorate bone loss induced by iron overload via activating Akt/GSK3β/Nrf2 pathway. J. Bone Miner. Metab. 2022, 40, 375–388. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient (g/kg) | ND | HFD | HFD-WPHL | HFD-WPHM | HFD-WPHH |
|---|---|---|---|---|---|
| Casein | 200 | 200 | 190 | 180 | 160 |
| WPH † | / | / | 12 | 24 | 48 |
| Lard oil | 20 | 177.5 | 177.5 | 177.5 | 177.5 |
| l–Cysteine | 3 | 3 | 3 | 3 | 3 |
| Corn starch | 452.2 | 72.8 | 72.8 | 72.8 | 72.8 |
| Maltodextrin | 75 | 100 | 100 | 100 | 100 |
| Sucrose | 172.8 | 172.8 | 172.8 | 172.8 | 172.8 |
| Cellulose $ | 50 | 50 | 48 | 46 | 42 |
| Corn oil | 25 | 25 | 25 | 25 | 25 |
| Mineral mixture | 45 | 45 | 45 | 45 | 45 |
| Vitamin mixture | 10 | 10 | 10 | 10 | 10 |
| Choline | 2 | 2 | 2 | 2 | 2 |
| Ratio of fat/total calorie (%) | 15 | 45 | 45 | 45 | 45 |
| No. | Peptide Sequence | m/z | MW # | Source (Protein Accession) |
|---|---|---|---|---|
| 1 | SSRQP | 287.65 | 573.29 | Glycosylation-dependent cell adhesion molecule 1 (P80195) |
| 2 | LNENK | 309.17 | 616.318 | Beta-lactoglobulin (P02754) |
| 3 | TPKAKDKNKH | 389.56 | 1165.65 | Osteopontin (P31096) |
| 4 | TPEVDDEALEK | 415.19 | 1245.58 | Beta-lactoglobulin (P02754) |
| 5 | TEAQEDGQSTSE | 427.17 | 1281.50 | Endoplasmin (Q95M18) |
| 6 | TPEVDDEA | 438.18 | 874.36 | Beta-lactoglobulin (P02754) |
| 7 | VPYPQRDMPIQ | 447.89 | 1343.68 | Beta-casein |
| 8 | TPKAKDKNKHSN | 456.58 | 1368.73 | Osteopontin (A0A077KSH2) |
| 9 | VSNAEGSQPDDSSS | 459.85 | 1379.56 | Perilipin (A1L5C2) |
| 10 | YLYEIAR | 464.25 | 926.49 | Albumin (P02769) |
| 11 | ENSAEPEQS | 495.70 | 989.39 | Beta-lactoglobulin (P02754) |
| 12 | NKPEDETH | 485.22 | 968.42 | Glycosylation-dependent cell adhesion molecule 1 (P80195) |
| 13 | KVPQVSTPTLVEVSR | 547.32 | 1638.93 | Albumin (P02769) |
| 14 | RNAVPITPTLN | 598.34 | 1194.67 | Alpha-S2-casein (P02663) |
| 15 | QSEEQQQTEDE | 675.26 | 1350.53 | Beta-casein (A0A452DHW7) |
| 16 | TEAQEDGQSTSE | 427.17 | 1281.50 | Endoplasmin (Q95M18) |
| ND | HFD | HFD-WPHL | HFD-WPHM | HFD-WPHH | |
|---|---|---|---|---|---|
| Body and tissue weight (g) | |||||
| 36.63 ± 3.25 a | 50.52 ± 3.24 b | 49.74 ± 4.02 b | 48.49 ± 3.37 b | 48.16 ± 3.76 b |
| 1.13 ± 0.11 a | 1.67 ± 0.20 b | 1.60 ± 0.18 b | 1.58 ± 0.21 b | 1.56 ± 0.22 b |
| 0.25 ± 0.05 a | 0.72 ± 0.13 b | 0.73 ± 0.14 b | 0.70 ± 0.06 b | 0.68 ± 0.10 b |
| 0.11 ± 0.05 a | 0.84 ± 0.10 b | 0.80 ± 0.12 b | 0.82 ± 0.09 b | 0.81 ± 0.10 b |
| 0.40 ± 0.06 a | 1.57 ± 0.08 c | 1.55 ± 0.06 c | 1.44 ± 0.07 b | 1.38 ± 0.09 b |
| Metabolic profile | |||||
| 3.28 ± 0.10 a | 7.64 ± 0.13 d | 7.33 ± 0.12 d | 6.27 ± 0.13 c | 5.83 ± 0.13 b |
| 1.01 ± 0.03 a | 1.80 ± 0.08 d | 1.75 ± 0.04 d | 1.46 ± 0.11 c | 1.28 ± 0.04 b |
| 0.78 ± 0.05 a | 1.45 ± 0.08 b | 1.43 ± 0.06 b | 1.40 ± 0.05 b | 1.41 ± 0.06 b |
| Femoral composition and mechanical properties | |||||
| 65.18 ± 3.14 c | 47.33 ± 2.28 a | 49.40 ± 2.09 a | 53.29 ± 2.51 b | 55.47 ± 2.67 b |
| 134.01 ± 9.84 c | 108.66 ± 5.37 a | 110.43 ± 6.52 ab | 112.41 ± 4.28 b | 116.54 ± 6.33 b |
| 46.76 ± 2.07 b | 35.28 ± 4.31 a | 35.54 ± 3.58 a | 36.50 ± 3.63 a | 35.33 ± 3.75 a |
| 102.54 ± 8.86 c | 79.53 ± 5.26 a | 82.23 ± 5.04 ab | 85.34 ± 4.67 ab | 88.23 ± 5.30 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bu, T.; Huang, J.; Yu, Y.; Sun, P.; Yang, K. Whey Protein Hydrolysate Ameliorated High-Fat-Diet Induced Bone Loss via Suppressing Oxidative Stress and Regulating GSK-3β/Nrf2 Signaling Pathway. Nutrients 2023, 15, 2863. https://doi.org/10.3390/nu15132863
Bu T, Huang J, Yu Y, Sun P, Yang K. Whey Protein Hydrolysate Ameliorated High-Fat-Diet Induced Bone Loss via Suppressing Oxidative Stress and Regulating GSK-3β/Nrf2 Signaling Pathway. Nutrients. 2023; 15(13):2863. https://doi.org/10.3390/nu15132863
Chicago/Turabian StyleBu, Tingting, Ju Huang, Yue Yu, Peilong Sun, and Kai Yang. 2023. "Whey Protein Hydrolysate Ameliorated High-Fat-Diet Induced Bone Loss via Suppressing Oxidative Stress and Regulating GSK-3β/Nrf2 Signaling Pathway" Nutrients 15, no. 13: 2863. https://doi.org/10.3390/nu15132863
APA StyleBu, T., Huang, J., Yu, Y., Sun, P., & Yang, K. (2023). Whey Protein Hydrolysate Ameliorated High-Fat-Diet Induced Bone Loss via Suppressing Oxidative Stress and Regulating GSK-3β/Nrf2 Signaling Pathway. Nutrients, 15(13), 2863. https://doi.org/10.3390/nu15132863