
Association between Gut Microbiota and Digestive System Cancers: A Bidirectional Two-Sample Mendelian Randomization Study

 and
and
Abstract
:1. Introduction
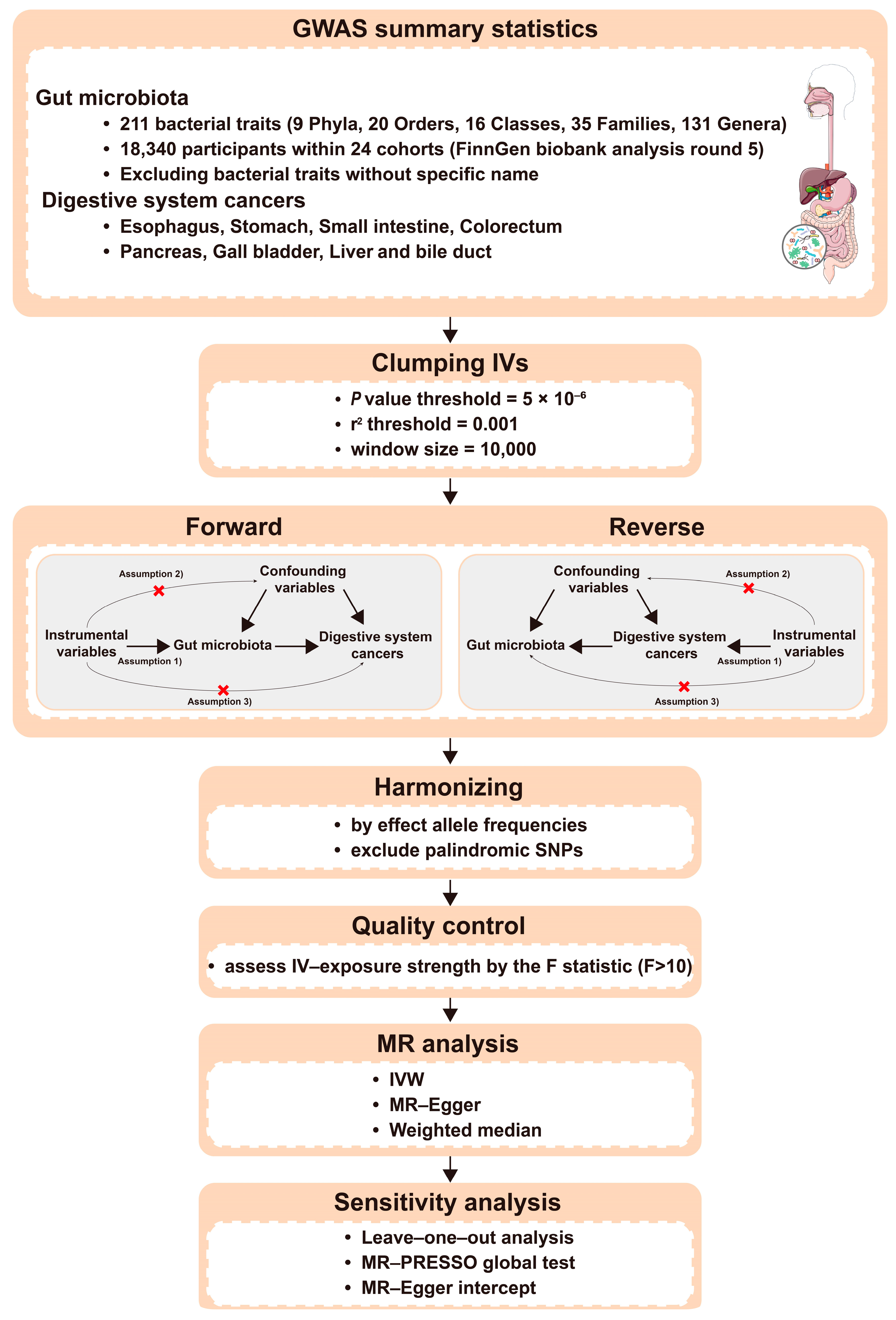
2. Method
2.1. Study Design
2.2. Data Sources and Study Population
2.3. Selection of IVs
3. Statistical Analysis
4. Results
4.1. Instrumental Variables
4.2. The Bidirectional Causal Associations between Gut Microbiota and DSCs
4.2.1. Gastric Cancer
4.2.2. Small Intestinal Cancer
4.2.3. Colorectal Cancer
4.2.4. Pancreatic Cancer
4.2.5. Liver Cancer
4.3. Sensitivity Analysis
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Jordan, B.F. Gut microbiota-mediated inflammation in obesity: A link with gastrointestinal cancer. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Smet, A.; Kupcinskas, J.; Link, A.; Hold, G.L.; Bornschein, J. The Role of Microbiota in Gastrointestinal Cancer and Cancer Treatment: Chance or Curse? Cell Mol. Gastroenterol. Hepatol. 2022, 13, 857–874. [Google Scholar] [CrossRef] [PubMed]
- de Vos, W.M.; Tilg, H.; Van Hul, M.; Cani, P.D. Gut microbiome and health: Mechanistic insights. Gut 2022, 71, 1020–1032. [Google Scholar] [CrossRef]
- Fernandes, M.R.; Aggarwal, P.; Costa, R.G.F.; Cole, A.M.; Trinchieri, G. Targeting the gut microbiota for cancer therapy. Nat. Rev. Cancer 2022, 22, 703–722. [Google Scholar] [CrossRef]
- Mima, K.; Nakagawa, S.; Sawayama, H.; Ishimoto, T.; Imai, K.; Iwatsuki, M.; Hashimoto, D.; Baba, Y.; Yamashita, Y.I.; Yoshida, N.; et al. The microbiome and hepatobiliary-pancreatic cancers. Cancer Lett. 2017, 402, 9–15. [Google Scholar] [CrossRef]
- Tilg, H.; Adolph, T.E.; Gerner, R.R.; Moschen, A.R. The Intestinal Microbiota in Colorectal Cancer. Cancer Cell 2018, 33, 954–964. [Google Scholar] [CrossRef] [Green Version]
- Okumura, S.; Konishi, Y.; Narukawa, M.; Sugiura, Y.; Yoshimoto, S.; Arai, Y.; Sato, S.; Yoshida, Y.; Tsuji, S.; Uemura, K.; et al. Gut bacteria identified in colorectal cancer patients promote tumourigenesis via butyrate secretion. Nat. Commun. 2021, 12, 5674. [Google Scholar] [CrossRef]
- Ma, J.; Li, J.; Jin, C.; Yang, J.; Zheng, C.; Chen, K.; Xie, Y.; Yang, Y.; Bo, Z.; Wang, J.; et al. Association of gut microbiome and primary liver cancer: A two-sample Mendelian randomization and case-control study. Liver Int. 2023, 43, 221–233. [Google Scholar] [CrossRef]
- Ma, C.; Han, M.; Heinrich, B.; Fu, Q.; Zhang, Q.; Sandhu, M.; Agdashian, D.; Terabe, M.; Berzofsky, J.A.; Fako, V.; et al. Gut microbiome-mediated bile acid metabolism regulates liver cancer via NKT cells. Science 2018, 360, eaan5931. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Guo, F.; Yu, Y.; Sun, T.; Ma, D.; Han, J.; Qian, Y.; Kryczek, I.; Sun, D.; Nagarsheth, N.; et al. Fusobacterium nucleatum Promotes Chemoresistance to Colorectal Cancer by Modulating Autophagy. Cell 2017, 170, 548–563 e516. [Google Scholar] [CrossRef] [Green Version]
- Mager, L.F.; Burkhard, R.; Pett, N.; Cooke, N.C.A.; Brown, K.; Ramay, H.; Paik, S.; Stagg, J.; Groves, R.A.; Gallo, M.; et al. Microbiome-derived inosine modulates response to checkpoint inhibitor immunotherapy. Science 2020, 369, 1481–1489. [Google Scholar] [CrossRef]
- Davey Smith, G.; Hemani, G. Mendelian randomization: Genetic anchors for causal inference in epidemiological studies. Hum. Mol. Genet. 2014, 23, R89–R98. [Google Scholar] [CrossRef] [Green Version]
- Grover, S.; Del Greco, M.F.; Stein, C.M.; Ziegler, A. Mendelian Randomization. Methods Mol. Biol. 2017, 1666, 581–628. [Google Scholar] [CrossRef]
- Davies, N.M.; Holmes, M.V.; Davey Smith, G. Reading Mendelian randomisation studies: A guide, glossary, and checklist for clinicians. BMJ 2018, 362, k601. [Google Scholar] [CrossRef] [Green Version]
- Hemani, G.; Tilling, K.; Davey Smith, G. Orienting the causal relationship between imprecisely measured traits using GWAS summary data. PLoS Genet. 2017, 13, e1007081. [Google Scholar] [CrossRef] [Green Version]
- Hemani, G.; Zheng, J.; Elsworth, B.; Wade, K.H.; Haberland, V.; Baird, D.; Laurin, C.; Burgess, S.; Bowden, J.; Langdon, R.; et al. The MR-Base platform supports systematic causal inference across the human phenome. Elife 2018, 7, e34408. [Google Scholar] [CrossRef]
- Verbanck, M.; Chen, C.Y.; Neale, B.; Do, R. Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nat. Genet. 2018, 50, 693–698. [Google Scholar] [CrossRef]
- Park, E.M.; Chelvanambi, M.; Bhutiani, N.; Kroemer, G.; Zitvogel, L.; Wargo, J.A. Targeting the gut and tumor microbiota in cancer. Nat. Med. 2022, 28, 690–703. [Google Scholar] [CrossRef]
- He, Z.; Gharaibeh, R.Z.; Newsome, R.C.; Pope, J.L.; Dougherty, M.W.; Tomkovich, S.; Pons, B.; Mirey, G.; Vignard, J.; Hendrixson, D.R.; et al. Campylobacter jejuni promotes colorectal tumorigenesis through the action of cytolethal distending toxin. Gut 2019, 68, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.J.; Tao, H.; Carloni, E.; Leung, W.K.; Graham, D.Y.; Sepulveda, A.R. Helicobacter pylori impairs DNA mismatch repair in gastric epithelial cells. Gastroenterology 2002, 123, 542–553. [Google Scholar] [CrossRef] [PubMed]
- Gur, C.; Ibrahim, Y.; Isaacson, B.; Yamin, R.; Abed, J.; Gamliel, M.; Enk, J.; Bar-On, Y.; Stanietsky-Kaynan, N.; Coppenhagen-Glazer, S.; et al. Binding of the Fap2 protein of Fusobacterium nucleatum to human inhibitory receptor TIGIT protects tumors from immune cell attack. Immunity 2015, 42, 344–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davar, D.; Zarour, H.M. Facts and Hopes for Gut Microbiota Interventions in Cancer Immunotherapy. Clin. Cancer Res. 2022, 28, 4370–4384. [Google Scholar] [CrossRef] [PubMed]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; Deroos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef] [Green Version]
- Trompette, A.; Gollwitzer, E.S.; Pattaroni, C.; Lopez-Mejia, I.C.; Riva, E.; Pernot, J.; Ubags, N.; Fajas, L.; Nicod, L.P.; Marsland, B.J. Dietary Fiber Confers Protection against Flu by Shaping Ly6c(−) Patrolling Monocyte Hematopoiesis and CD8(+) T Cell Metabolism. Immunity 2018, 48, 992–1005 e1008. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Li, F.; Liang, B.; Liang, Y.; Chen, S.; Mo, X.; Ju, Y.; Zhao, H.; Jia, H.; Spector, T.D.; et al. A metagenome-wide association study of gut microbiota in asthma in UK adults. BMC Microbiol. 2018, 18, 114. [Google Scholar] [CrossRef] [Green Version]
- Cekanaviciute, E.; Yoo, B.B.; Runia, T.F.; Debelius, J.W.; Singh, S.; Nelson, C.A.; Kanner, R.; Bencosme, Y.; Lee, Y.K.; Hauser, S.L.; et al. Gut bacteria from multiple sclerosis patients modulate human T cells and exacerbate symptoms in mouse models. Proc. Natl. Acad. Sci. USA 2017, 114, 10713–10718. [Google Scholar] [CrossRef] [Green Version]
- Xiang, K.; Wang, P.; Xu, Z.; Hu, Y.Q.; He, Y.S.; Chen, Y.; Feng, Y.T.; Yin, K.J.; Huang, J.X.; Wang, J.; et al. Causal Effects of Gut Microbiome on Systemic Lupus Erythematosus: A Two-Sample Mendelian Randomization Study. Front Immunol. 2021, 12, 667097. [Google Scholar] [CrossRef]
- Chen, J.; Wright, K.; Davis, J.M.; Jeraldo, P.; Marietta, E.V.; Murray, J.; Nelson, H.; Matteson, E.L.; Taneja, V. An expansion of rare lineage intestinal microbes characterizes rheumatoid arthritis. Genome Med. 2016, 8, 43. [Google Scholar] [CrossRef] [Green Version]
- Bai, X.; Wei, H.; Liu, W.; Coker, O.O.; Gou, H.; Liu, C.; Zhao, L.; Li, C.; Zhou, Y.; Wang, G.; et al. Cigarette smoke promotes colorectal cancer through modulation of gut microbiota and related metabolites. Gut 2022, 71, 2439–2450. [Google Scholar] [CrossRef]
- Noto, J.M.; Piazuelo, M.B.; Shah, S.C.; Romero-Gallo, J.; Hart, J.L.; Di, C.; Carmichael, J.D.; Delgado, A.G.; Halvorson, A.E.; Greevy, R.A.; et al. Iron deficiency linked to altered bile acid metabolism promotes Helicobacter pylori-induced inflammation-driven gastric carcinogenesis. J. Clin. Investig. 2022, 132, e147822. [Google Scholar] [CrossRef]
- Lavelle, A.; Nancey, S.; Reimund, J.M.; Laharie, D.; Marteau, P.; Treton, X.; Allez, M.; Roblin, X.; Malamut, G.; Oeuvray, C.; et al. Fecal microbiota and bile acids in IBD patients undergoing screening for colorectal cancer. Gut Microbes 2022, 14, 2078620. [Google Scholar] [CrossRef]
- Yinhang, W.; Jing, Z.; Jie, Z.; Yin, J.; Xinyue, W.; Yifei, S.; Zhiqing, F.; Wei, W.; Shuwen, H. Prediction model of colorectal cancer (CRC) lymph node metastasis based on intestinal bacteria. Clin. Transl. Oncol. 2023, 25, 1661–1672. [Google Scholar] [CrossRef]
- Gao, J.; Yan, K.T.; Wang, J.X.; Dou, J.; Wang, J.; Ren, M.; Ma, J.; Zhang, X.; Liu, Y. Gut microbial taxa as potential predictive biomarkers for acute coronary syndrome and post-STEMI cardiovascular events. Sci. Rep. 2020, 10, 2639. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; Wang, Q.; Li, L. A genome-wide systems analysis reveals strong link between colorectal cancer and trimethylamine N-oxide (TMAO), a gut microbial metabolite of dietary meat and fat. BMC Genom. 2015, 16 (Suppl. S7), S4. [Google Scholar] [CrossRef] [Green Version]
- Chan, C.W.H.; Law, B.M.H.; Waye, M.M.Y.; Chan, J.Y.W.; So, W.K.W.; Chow, K.M. Trimethylamine-N-oxide as One Hypothetical Link for the Relationship between Intestinal Microbiota and Cancer—Where We Are and Where Shall We Go? J. Cancer 2019, 10, 5874–5882. [Google Scholar] [CrossRef]
- Zhang, X.; Yu, D.; Wu, D.; Gao, X.; Shao, F.; Zhao, M.; Wang, J.; Ma, J.; Wang, W.; Qin, X.; et al. Tissue-resident Lachnospiraceae family bacteria protect against colorectal carcinogenesis by promoting tumor immune surveillance. Cell Host Microbe 2023, 31, 418–432 e418. [Google Scholar] [CrossRef]
- Flemer, B.; Warren, R.D.; Barrett, M.P.; Cisek, K.; Das, A.; Jeffery, I.B.; Hurley, E.; O′Riordain, M.; Shanahan, F.; O′Toole, P.W. The oral microbiota in colorectal cancer is distinctive and predictive. Gut 2018, 67, 1454–1463. [Google Scholar] [CrossRef] [Green Version]
- Li, J.Y.; Gillilland, M., 3rd; Lee, A.A.; Wu, X.; Zhou, S.Y.; Owyang, C. Secondary bile acids mediate high-fat diet-induced upregulation of R-spondin 3 and intestinal epithelial proliferation. JCI Insight 2022, 7, e148309. [Google Scholar] [CrossRef]
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672. [Google Scholar] [CrossRef]
- Liu, W.; Crott, J.W.; Lyu, L.; Pfalzer, A.C.; Li, J.; Choi, S.W.; Yang, Y.; Mason, J.B.; Liu, Z. Diet- and Genetically-induced Obesity Produces Alterations in the Microbiome, Inflammation and Wnt Pathway in the Intestine of Apc(+/1638N) Mice: Comparisons and Contrasts. J. Cancer 2016, 7, 1780–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Wu, S.D.; Chen, Y.; Li, X.Y.; Zhu, Q.; Nakayama, K.; Zhang, W.Q.; Weng, C.Z.; Zhang, J.; Wang, H.K.; et al. Alterations in gut microbiome and metabolomics in chronic hepatitis B infection-associated liver disease and their impact on peripheral immune response. Gut Microbes 2023, 15, 2155018. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Diaz, C.; Taminiau, B.; Garcia-Garcia, A.; Cueto, A.; Robles-Diaz, M.; Ortega-Alonso, A.; Martin-Reyes, F.; Daube, G.; Sanabria-Cabrera, J.; Jimenez-Perez, M.; et al. Microbiota diversity in nonalcoholic fatty liver disease and in drug-induced liver injury. Pharmacol. Res. 2022, 182, 106348. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Ng, S.K.; Ding, Y.; Lin, Y.; Liu, W.; Wong, S.H.; Sung, J.J.; Yu, J. Meta-analysis of mucosal microbiota reveals universal microbial signatures and dysbiosis in gastric carcinogenesis. Oncogene 2022, 41, 3599–3610. [Google Scholar] [CrossRef]
- Coker, O.O.; Dai, Z.; Nie, Y.; Zhao, G.; Cao, L.; Nakatsu, G.; Wu, W.K.; Wong, S.H.; Chen, Z.; Sung, J.J.Y.; et al. Mucosal microbiome dysbiosis in gastric carcinogenesis. Gut 2018, 67, 1024–1032. [Google Scholar] [CrossRef]
- Liu, B.; Ye, D.; Yang, H.; Song, J.; Sun, X.; Mao, Y.; He, Z. Two-Sample Mendelian Randomization Analysis Investigates Causal Associations Between Gut Microbial Genera and Inflammatory Bowel Disease, and Specificity Causal Associations in Ulcerative Colitis or Crohn′s Disease. Front Immunol. 2022, 13, 921546. [Google Scholar] [CrossRef]
- Matson, V.; Fessler, J.; Bao, R.; Chongsuwat, T.; Zha, Y.; Alegre, M.L.; Luke, J.J.; Gajewski, T.F. The commensal microbiome is associated with anti-PD-1 efficacy in metastatic melanoma patients. Science 2018, 359, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Zaidi, A.H.; Pratama, M.Y.; Omstead, A.N.; Gorbonova, A.; Mansoor, R.; Melton-Kreft, R.; Jobe, B.A.; Wagner, P.L.; Kelly, R.J.; Goel, A. A blood-based circulating microbial metagenomic panel for early diagnosis and prognosis of oesophageal adenocarcinoma. Br. J. Cancer 2022, 127, 2016–2024. [Google Scholar] [CrossRef]
- McCulloch, J.A.; Davar, D.; Rodrigues, R.R.; Badger, J.H.; Fang, J.R.; Cole, A.M.; Balaji, A.K.; Vetizou, M.; Prescott, S.M.; Fernandes, M.R.; et al. Intestinal microbiota signatures of clinical response and immune-related adverse events in melanoma patients treated with anti-PD-1. Nat. Med. 2022, 28, 545–556. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Qian, Y.; Xie, Y.H.; Jiang, S.S.; Kang, Z.R.; Chen, Y.X.; Chen, Z.F.; Fang, J.Y. Alterations in the oral and gut microbiome of colorectal cancer patients and association with host clinical factors. Int. J. Cancer 2021, 149, 925–935. [Google Scholar] [CrossRef]
- Li, H.; Du, X.; Yan, L.; Tang, Z.; Zhang, L.; Zheng, Q.; Zeng, X.; Chen, G.; Yue, H.; Fu, X. Low Abundance of Lactococcus lactis in Human Colorectal Cancer Is Associated with Decreased Natural Killer Cells. Nutr. Cancer 2022, 74, 938–946. [Google Scholar] [CrossRef]
- Gao, Z.; Guo, B.; Gao, R.; Zhu, Q.; Qin, H. Microbiota disbiosis is associated with colorectal cancer. Front Microbiol. 2015, 6, 20. [Google Scholar] [CrossRef]
- Cheng, W.Y.; Wu, C.Y.; Yu, J. The role of gut microbiota in cancer treatment: Friend or foe? Gut 2020, 69, 1867–1876. [Google Scholar] [CrossRef]
- Wang, Y.; Wiesnoski, D.H.; Helmink, B.A.; Gopalakrishnan, V.; Choi, K.; DuPont, H.L.; Jiang, Z.D.; Abu-Sbeih, H.; Sanchez, C.A.; Chang, C.C.; et al. Fecal microbiota transplantation for refractory immune checkpoint inhibitor-associated colitis. Nat. Med. 2018, 24, 1804–1808. [Google Scholar] [CrossRef]
- Salemi, R.; Vivarelli, S.; Ricci, D.; Scillato, M.; Santagati, M.; Gattuso, G.; Falzone, L.; Libra, M. Lactobacillus rhamnosus GG cell-free supernatant as a novel anti-cancer adjuvant. J. Transl. Med. 2023, 21, 195. [Google Scholar] [CrossRef]
- Si, W.; Liang, H.; Bugno, J.; Xu, Q.; Ding, X.; Yang, K.; Fu, Y.; Weichselbaum, R.R.; Zhao, X.; Wang, L. Lactobacillus rhamnosus GG induces cGAS/STING-dependent type I interferon and improves response to immune checkpoint blockade. Gut 2022, 71, 521–533. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| GWAS_ID | Disease | Consortium | Sample Size | Case | Control | Population |
|---|---|---|---|---|---|---|
| ieu-b-4960 | Oesophageal cancer | UK Biobank | 372,756 | 740 | 372,016 | European |
| finn-b-C3_STOMACH | GC | Finngen | 218,792 | 633 | 218,159 | European |
| finn-b-C3_SMALL_INTESTINE | Small intestine cancer | Finngen | 218,792 | 252 | 218,540 | European |
| finn-b-C3_COLORECTAL | CRC | Finngen | 218,792 | 3022 | 215,770 | European |
| ieu-a-822 | Pancreatic cancer | PanScan1 | 3835 | 1896 | 1939 | European |
| ieu-a-1057 | Gallbladder cancer | NA | 907 | 41 | 866 | East Asian |
| finn-b-C3_LIVER_INTRAHEPATIC_BILE_DUCTS | Liver cancer | Finngen | 218,792 | 304 | 218,488 | European |
| Outcome | Exposure | SNPs (n) | p-Val | OR | 95%CI |
|---|---|---|---|---|---|
| GC | family Bacteroaceae | 3 | 0.001 | 0.156 | (0.053, 0.459) |
| family Enterobacteriaceae | 3 | 0.006 | 0.206 | (0.067, 0.636) | |
| genus Bacteroes | 3 | 0.001 | 0.156 | (0.053, 0.459) | |
| genus Eggerthella | 5 | 0.005 | 0.464 | (0.270, 0.796) | |
| genus Lachnospira | 1 | 0.026 | 0.079 | (0.008, 0.739) | |
| order Enterobacteriales | 3 | 0.006 | 0.206 | (0.067, 0.636) | |
| genus Escherichia Shigella | 4 | 0.025 | 3.099 | (1.152, 8.338) | |
| genus Eubacterium fissicatena group | 5 | 0.038 | 1.648 | (1.028, 2.642) | |
| genus Ruminococcaceae UCG014 | 6 | 0.047 | 1.911 | (1.010, 3.615) | |
| Small intestine cancer | family Clostriales vadin BB60 group | 10 | 0.030 | 0.422 | (0.194, 0.918) |
| family Peptostreptococcaceae | 8 | 0.035 | 0.292 | (0.093, 0.918) | |
| genus Anaerofilum | 6 | 0.033 | 0.444 | (0.211, 0.936) | |
| genus Streptococcus | 8 | 0.045 | 0.266 | (0.073, 0.969) | |
| order Lactobacillales | 9 | 0.032 | 0.314 | (0.109, 0.903) | |
| genus Candatus Soleaferrea | 2 | 0.030 | 5.166 | (1.171, 22.797) | |
| CRC | family Clostriales vadin BB60 group | 10 | 0.044 | 0.751 | (0.569, 0.992) |
| genus Lachnospiraceae FCS020 group | 7 | 0.003 | 0.607 | (0.439, 0.840) | |
| phylum Euryarchaeota | 5 | 0.036 | 0.801 | (0.650, 0.986) | |
| phylum Proteobacteria | 4 | 0.037 | 0.613 | (0.387, 0.971) | |
| Pancreatic cancer | genus Bilophila | 1 | 0.031 | 0.187 | (0.041, 0.855) |
| genus Streptococcus | 2 | 0.032 | 0.253 | (0.072, 0.888) | |
| genus Ruminococcaceae UCG014 | 1 | 0.029 | 6.101 | (1.206, 30.873) | |
| Liver cancer | family Rhodospirillaceae | 7 | 0.014 | 0.357 | (0.157, 0.809) |
| genus Escherichia Shigella | 4 | 0.026 | 0.198 | (0.048, 0.821) | |
| genus Eubacterium nodatum group | 3 | 0.009 | 0.382 | (0.186, 0.783) | |
| genus Family XIII AD3011 group | 7 | 0.030 | 0.261 | (0.078, 0.876) | |
| genus Turicibacter | 5 | 0.005 | 0.271 | (0.109, 0.676) | |
| genus Dorea | 5 | 0.010 | 8.102 | (1.643, 39.965) | |
| genus Lachnospiraceae UCG004 | 7 | 0.039 | 3.199 | (1.059, 9.662) | |
| genus Oscillibacter | 12 | 0.021 | 2.129 | (1.123, 4.035) | |
| genus Paraprevotella | 9 | 0.032 | 1.961 | (1.058, 3.636) |
| Exposure | Outcome | SNPs (n) | p-Val | Beta | 95%CI |
|---|---|---|---|---|---|
| GC | class Methanobacteria | 5 | 0.023 | −0.087 | (−0.161, −0.012) |
| family Methanobacteriaceae | 5 | 0.023 | −0.087 | (−0.161, −0.012) | |
| family Oxalobacteraceae | 5 | 0.014 | −0.076 | (−0.137, −0.015) | |
| genus Methanobrevibacter | 5 | 0.005 | −0.109 | (−0.184, −0.033) | |
| genus Oxalobacter | 5 | 0.032 | −0.070 | (−0.134, −0.006) | |
| order Methanobacteriales | 5 | 0.023 | −0.087 | (−0.161, −0.012) | |
| phylum Euryarchaeota | 5 | 0.047 | −0.074 | (−0.147, −0.001) | |
| genus Dialister | 5 | 0.001 | 0.070 | (0.029, 0.110) | |
| genus Eubacterium ventriosum group | 5 | 0.032 | 0.039 | (0.003, 0.075) | |
| Small intestine cancer | genus Intestinibacter | 5 | 0.022 | −0.033 | (−0.060, −0.005) |
| genus Lachnoclostrium | 5 | 0.023 | −0.024 | (−0.044, −0.003) | |
| genus Peptococcus | 5 | 0.004 | −0.051 | (−0.086, −0.016) | |
| genus Collinsella | 5 | 4.80 × 10-4 | 0.040 | (0.018, 0.063) | |
| genus Erysipelotrichaceae UCG003 | 1 | 0.046 | 0.057 | (0.001, 0.113) | |
| genus Eubacterium ruminantium group | 5 | 0.026 | 0.038 | (0.005, 0.072) | |
| genus Howardella | 4 | 0.002 | 0.085 | (0.031, 0.138) | |
| genus Lachnospiraceae UCG008 | 5 | 0.002 | 0.051 | (0.018, 0.084) | |
| CRC | genus Lactococcus | 9 | 0.004 | −0.157 | (−0.263, −0.051) |
| genus Marvinbryantia | 9 | 0.016 | −0.081 | (−0.148, −0.015) | |
| genus Eggerthella | 9 | 0.045 | 0.100 | (0.002, 0.197) | |
| genus Eisenbergiella | 9 | 0.021 | 0.106 | (0.016, 0.196) | |
| Pancreatic cancer | genus Eubacterium ruminantium group | 3 | 0.013 | −0.104 | (−0.186, −0.022) |
| genus Lachnospiraceae NC2004 group | 3 | 0.013 | −0.116 | (−0.208, −0.025) | |
| genus Ruminococcaceae UCG014 | 3 | 0.004 | 0.088 | (0.028, 0.148) | |
| Liver cancer | genus Butyricimonas | 2 | 0.032 | −0.056 | (−0.107, −0.005) |
| genus Eubacterium nodatum group | 2 | 0.029 | −0.104 | (−0.197, −0.010) | |
| family Actinomycetaceae | 2 | 0.027 | 0.071 | (0.008, 0.135) | |
| family Lactobacillaceae | 2 | 0.033 | 0.069 | (0.006, 0.132) | |
| genus Eubacterium brachy group | 2 | 0.044 | 0.087 | (0.002, 0.171) | |
| genus Lactobacillus | 2 | 0.036 | 0.068 | (0.004, 0.132) | |
| order Actinomycetales | 2 | 0.024 | 0.071 | (0.009, 0.133) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, N.; Wang, Z.; Shu, Q.; Liang, X.; Wang, J.; Wu, K.; Nie, Y.; Shi, Y.; Fan, D.; Wu, J. Association between Gut Microbiota and Digestive System Cancers: A Bidirectional Two-Sample Mendelian Randomization Study. Nutrients 2023, 15, 2937. https://doi.org/10.3390/nu15132937
Xie N, Wang Z, Shu Q, Liang X, Wang J, Wu K, Nie Y, Shi Y, Fan D, Wu J. Association between Gut Microbiota and Digestive System Cancers: A Bidirectional Two-Sample Mendelian Randomization Study. Nutrients. 2023; 15(13):2937. https://doi.org/10.3390/nu15132937
Chicago/Turabian StyleXie, Ning, Ziwei Wang, Qiuai Shu, Xiru Liang, Jinhai Wang, Kaichun Wu, Yongzhan Nie, Yongquan Shi, Daiming Fan, and Jian Wu. 2023. "Association between Gut Microbiota and Digestive System Cancers: A Bidirectional Two-Sample Mendelian Randomization Study" Nutrients 15, no. 13: 2937. https://doi.org/10.3390/nu15132937
APA StyleXie, N., Wang, Z., Shu, Q., Liang, X., Wang, J., Wu, K., Nie, Y., Shi, Y., Fan, D., & Wu, J. (2023). Association between Gut Microbiota and Digestive System Cancers: A Bidirectional Two-Sample Mendelian Randomization Study. Nutrients, 15(13), 2937. https://doi.org/10.3390/nu15132937