
Selected Flavonols in Breast and Gynecological Cancer: A Systematic Review


Abstract
:1. Introduction
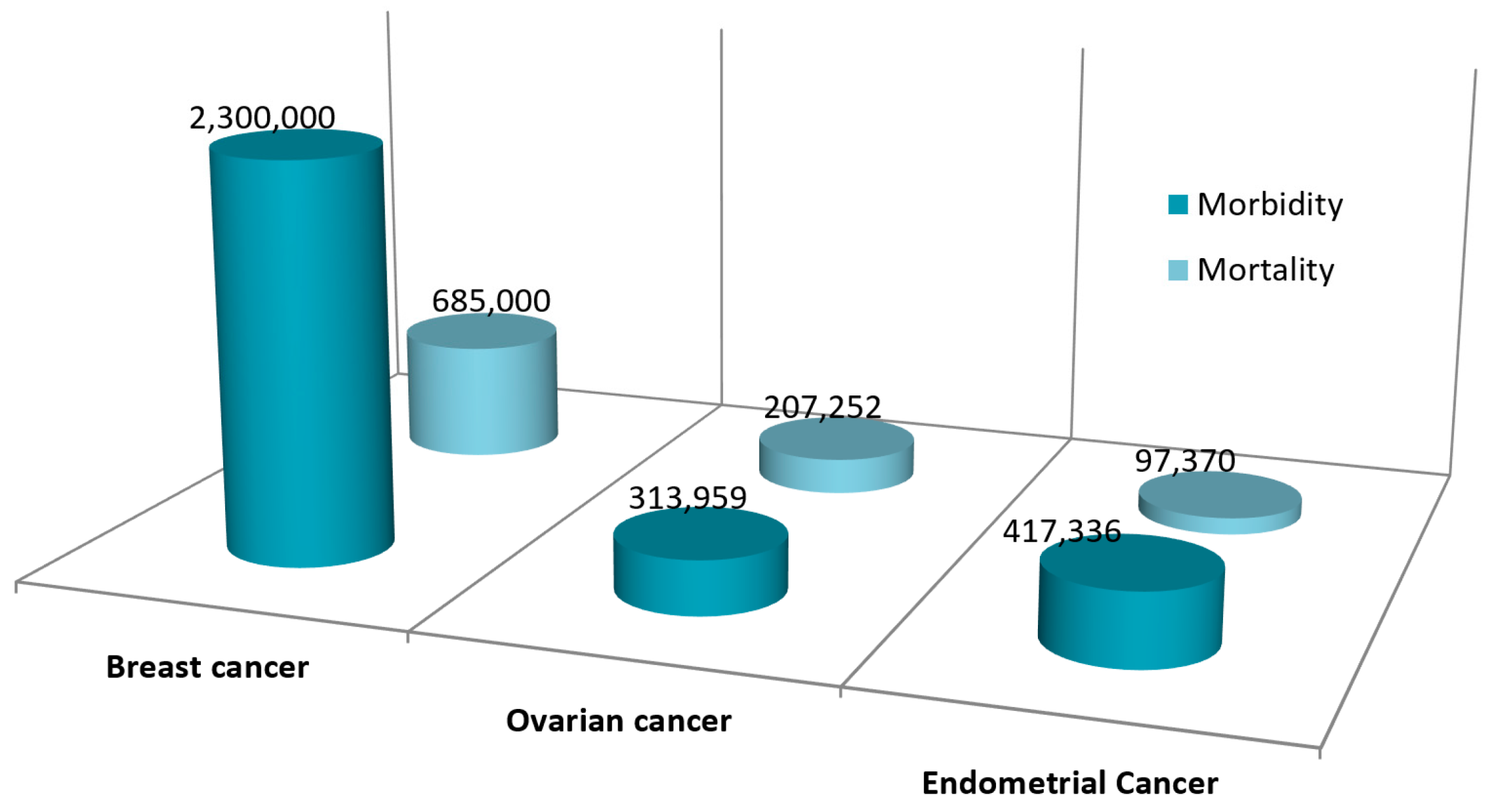
1.1. Breast Cancer
1.2. Ovarian Cancer
1.3. Endometrial Cancer
2. Sources of Flavonoids
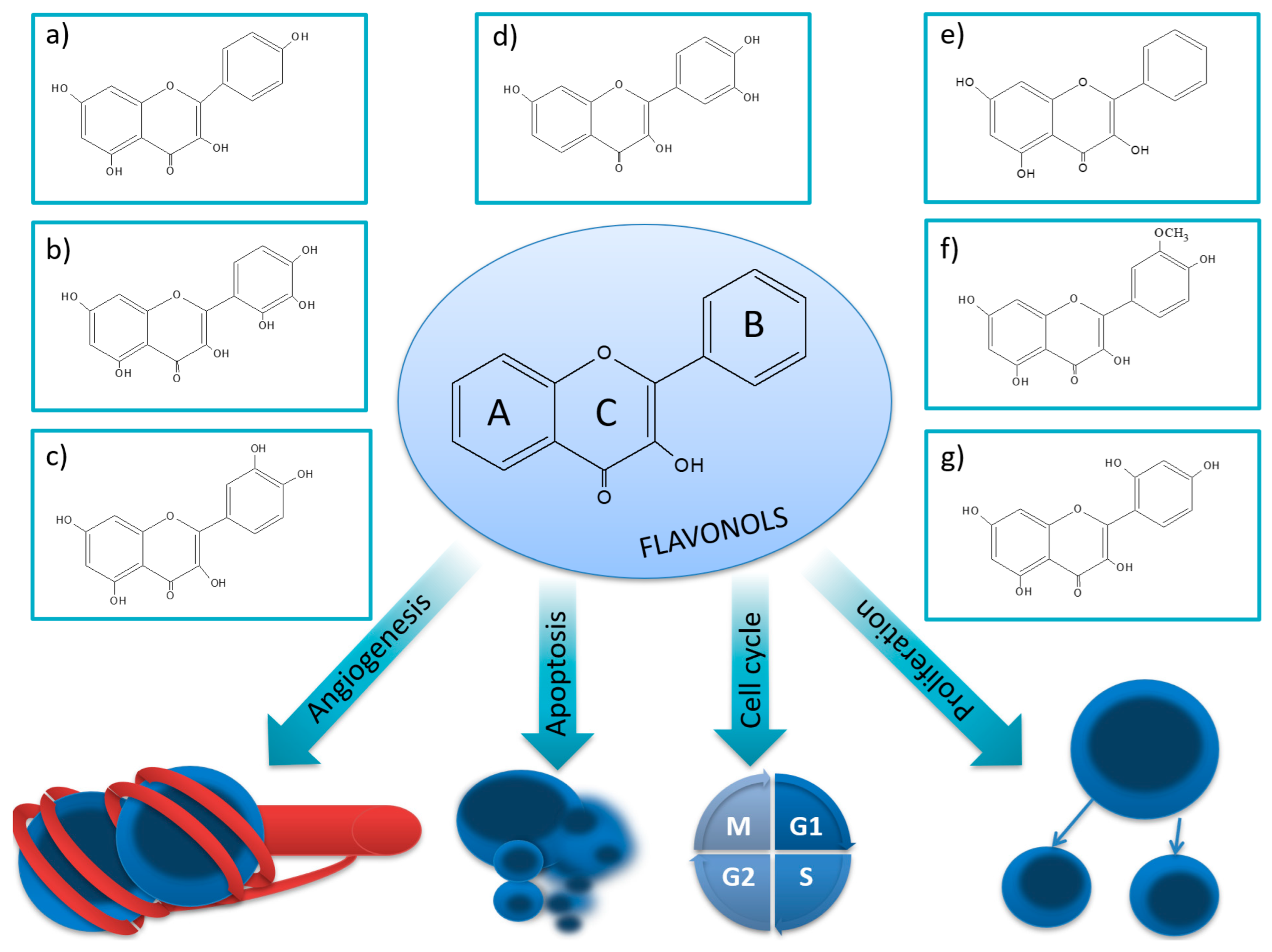
3. Flavonols
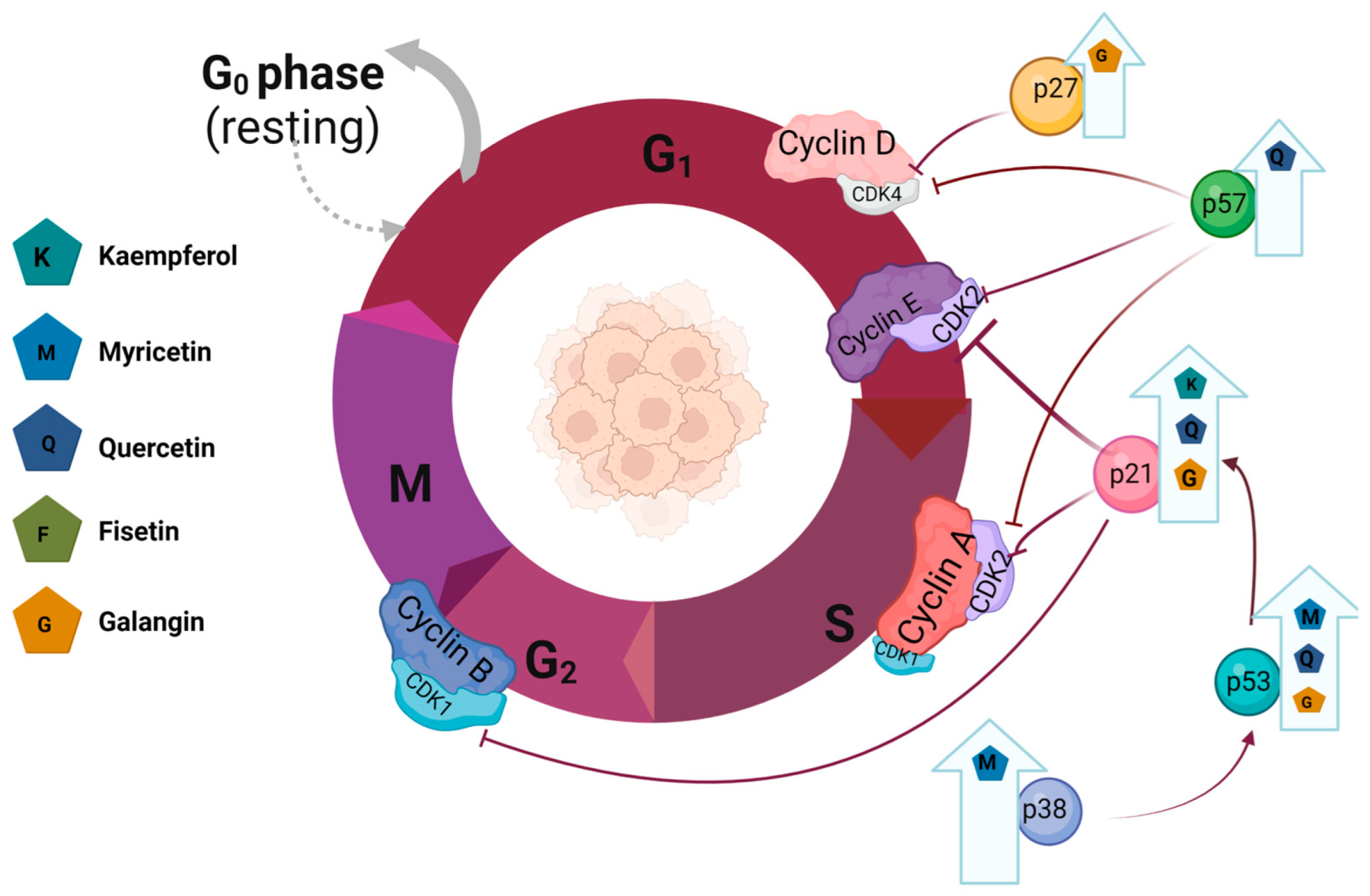
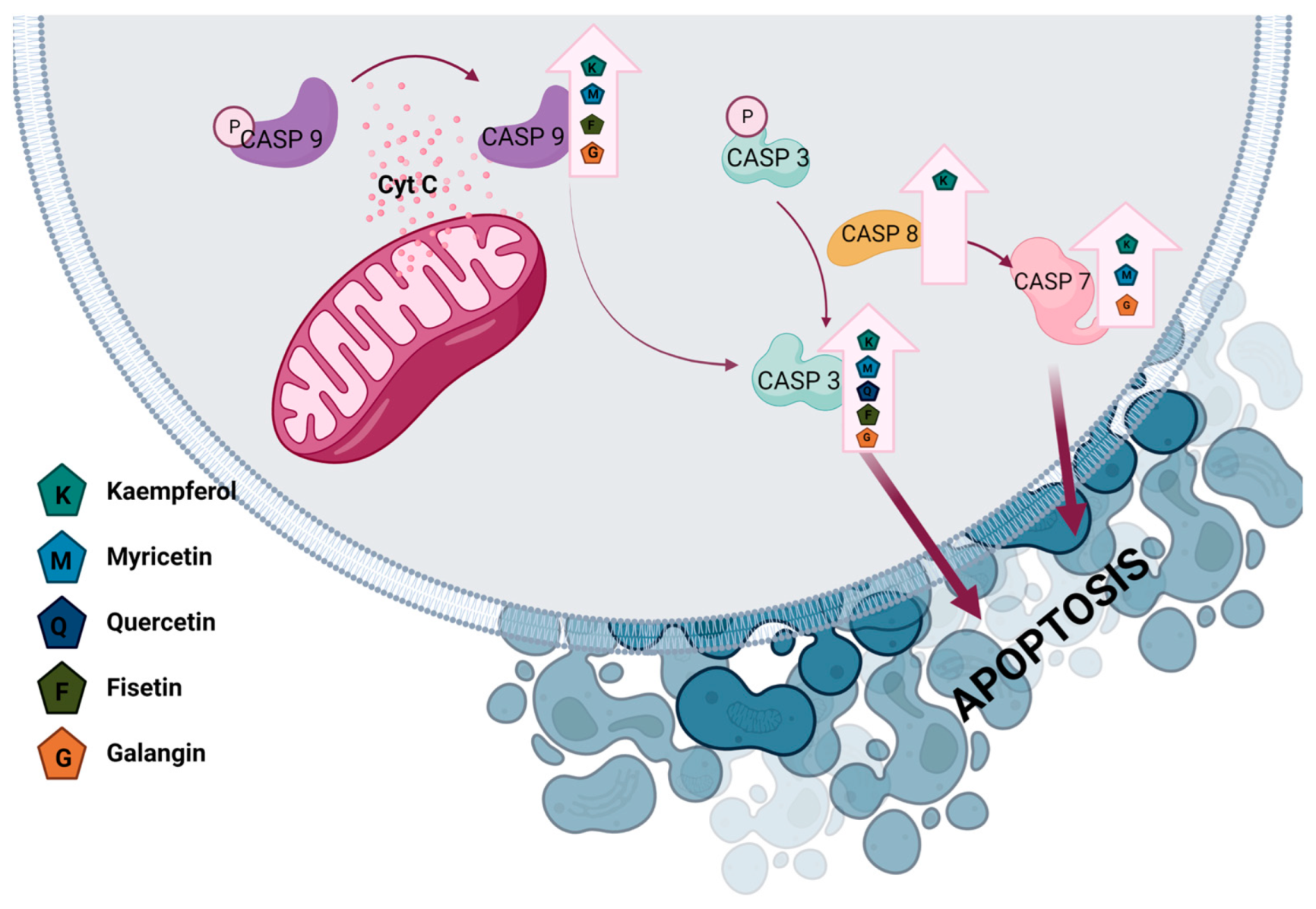
3.1. Kaempferol
3.1.1. Effect of Kaempferol on Breast Cancer
3.1.2. Effect of Kaempferol on Ovarian Cancer
3.1.3. Effect of Kaempferol on Endometrial Cancer

| Flavonoid | Cell Line | Mechanism | Result | Refs. |
|---|---|---|---|---|
| Kaempferol | MCF-7 |
| Antiproliferative activity | [44] |
| Antiproliferative activity | [44] | ||
| Antiproliferative activity | [44] | ||
| Induced apoptosis | [44,46] | ||
| VM7LUC4E2 |
| Induced apoptosis | [47] | |
| Induced apoptosis | [47] | ||
| MDA-MB-231 |
| Anti-invasion activity | [49] | |
| Anti-invasion activity | [50] | ||
| Induced apoptosis | [51] | ||
| Antimigration activity Anti-invasion activity | [52] | ||
| MDA-MB-453 |
| Antimigration activity Anti-invasion activity | [52] | |
| Myricetin | T47-D |
| Induced apoptosis | [64] |
| Induced apoptosis | [64] | ||
| Induced apoptosis | [64] | ||
| MCF-7 |
| Induced apoptosis | [65] | |
| Induced apoptosis | [65,66] | ||
| Induced apoptosis | [65] | ||
| Induced apoptosis | [65] | ||
| Anti-invasion activity | [66] | ||
| Anti-invasion activity | [66] | ||
| Induced apoptosis | [66] | ||
| Induced apoptosis | [66] | ||
| Induced apoptosis | [66] | ||
| MDA-MB-231 |
| Induced apoptosis | [67] | |
| Induced apoptosis | [67] | ||
| MDA-MB-231Br |
| Anti-invasion activity | [68] | |
| MDA-MB-468 |
| Induced apoptosis | [67] | |
| Induced apoptosis | [67] | ||
| SK-BR-3 |
| Induced apoptosis | [69] | |
| Induced apoptosis | [69] | ||
| Induced apoptosis, | [69] | ||
| Induced autophagy | [69] | ||
| Induced autophagy | [69] | ||
| Quercetin | MCF-7 |
| Antiproliferative activity | [70] |
| Antiproliferative activity | [70,71] | ||
| Antiproliferative activity | [70] | ||
| Antiproliferative activity | [70,72,73] | ||
| Induced apoptosis | [70,74,75,76,77] | ||
| Antiproliferative activity and Induced apoptosis | [72,73,78,79] | ||
| Antiproliferative activity | [76] | ||
| Inhibited cancer growth | [72] | ||
| Anti-invasion activity | [71] | ||
| Anti-invasion activity | [71] | ||
| Induced apoptosis | [73,74,75,76,78,80] | ||
| Induced necroptosis | [74] | ||
| Promoted apoptosis | [81] | ||
| MDA-MB-231 |
| Antimigration activity and antiproliferative activity | [71,82] | |
| Induced apoptosis | [76,77] | ||
| Induced apoptosis | [76,77] | ||
| Antiproliferative activity | [83] | ||
| Antiproliferative activity | [76] | ||
| Suppressed edepitheliale–mesenchymal transition (EMT) | [84] | ||
| Anti-invasion activity | [85] | ||
| Anti-invasion activity | [71,84] | ||
| Anti-invasion activity | [71] | ||
| Induced apoptosis | [83] | ||
| MDA-MB-435 |
| Antimigration activity and antiproliferative activity | [82] | |
| BT474 |
| Induced apoptosis | [86] | |
| BT 20 |
| Induced apoptosis | [80] | |
| Fisetin | 4T1 |
| Anti-invasion activity | [87] |
| Antiproliferative activity | [88] | ||
| Antiproliferative activity | [88] | ||
| Antiproliferative activity | [88] | ||
| Antiproliferative activity | [88] | ||
| MCF-7 |
| Anti-invasion activity | [89] | |
| Induced apoptosis | [90] | ||
| MDA-MB-231 |
| Anti-invasion activity | [89] | |
| Antiproliferative activity | [91] | ||
| MDA-MB-453 |
| Antiproliferative activity | [92] | |
| Antiproliferative activity | [92] | ||
| Galangin | T47D |
| Induced apoptosis | [93] |
| Antiproliferative activity | [93] | ||
| Induced apoptosis | [93] | ||
| MCF-7 |
| Induced apoptosis | [93,94] | |
| Antiproliferative activity | [93] | ||
| Antiproliferative activity | [94] | ||
| Antiproliferative activity | [94] | ||
| Antiproliferative activity and induced apoptosis | [94] | ||
| Induced apoptosis | [93] | ||
| Anti-invasion activity | [95] | ||
| ER- Hs578T |
| Antiproliferative activity | [96] | |
| MDA-MB-231 |
| Anti-invasion activity | [95] | |
| Isorhamnetin | MCF-7 |
| Antiproliferative activity and induced apoptosis | [97] |
| Induced apoptosis | [98] | ||
| Induced apoptosis | [98] | ||
| MDA-MB-231 |
| Antiproliferative activity and induced apoptosis | [97] | |
| MDA-MB-468 |
| Induced apoptosis | [98] | |
| Induced apoptosis | [98] | ||
| Morin | MDA-MB-231 |
| Anti-invasion activity | [99] |
| Anti-invasion activity | [100] | ||
| Antiproliferative activity | [100] | ||
| Suppressed epitheliale–mesenchymal transition (EMT) | [100] |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
3.2. Myricetin
3.2.1. Effect of Myricetin on Breast Cancer
3.2.2. Effect of Myricetin on Ovarian Cancer
3.3. Quercetin
3.3.1. Effect of Quercetin on Breast Cancer
3.3.2. Effect of Quercetin on Ovarian Cancer
3.3.3. Effect of Quercetin on Endometrial Cancer
| Flavonoid | Cell line | Mechanism | Result | Ref. |
|---|---|---|---|---|
| Kaempferol | OVCAR-3 |
| Induced apoptosis | [54,55] |
| Induced apoptosis | [54] | ||
| Induced apoptosis | [54,55,57] | ||
| Induced apoptosis | [57] | ||
| Anti-invasion activity | [59] | ||
| Induced apoptosis | [57] | ||
| Induced apoptosis, | [55] | ||
| A2780/CP70 |
| Induced apoptosis | [58] | |
| Induced apoptosis | [58] | ||
| Induced apoptosis, | [56] | ||
| Cell arrest | [56] | ||
| Anti-invasion activity and antiangiogenesis | [59] | ||
| Induced apoptosis | [58] | ||
| SKOV3 |
| Induced apoptosis | [57] | |
| Induced apoptosis | [57] | ||
| Induced apoptosis | [55] | ||
| A2780 |
| Induced autophagy | [60] | |
| Induced enoplasmic reticulum stress (ER stress) | [60] | ||
| Myricetin | OVCAR-3 |
| Induced apoptosis | [112,113] |
| Induced apoptosis | [112] | ||
| Induced apoptosis | [112] | ||
| Cell cycle arrest | [112] | ||
| Induced apoptosis | [113] | ||
| Anti-invasion activity, antiangiogenesis | [112,116] | ||
| Antiangiogenesis | [112,116] | ||
| Antiangiogenesis | [112,116] | ||
| SKOV3 |
| Anti-invasion activity | [111] | |
| Induced apoptosis | [111,115] | ||
| Antiproliferative activity | [111] | ||
| Induced Endoplasmic reticulum stress | [115] | ||
| A2780/CP70 |
| Induced apoptosis | [112,114,116] | |
| Induced apoptosis | [112] | ||
| Cell cycle arrest | [112] | ||
| Induced apoptosis | [112,114] | ||
| Antiproliferation activity | [112,114] | ||
| Induced apoptosis | [114] | ||
| Cell cycle arrest | [114] | ||
| Anti-invasion activity, antiangiogenesis | [116] | ||
| Antiangiogenesis | [116] | ||
| Antiangiogenesis | [116] | ||
| Quercetin | PA-1 |
| Antiproliferation activity | [145] |
| Antiproliferation activity | [145] | ||
| Antiproliferation activity | [145] | ||
| Antiproliferation activity | [145] | ||
| Anti-invasion activity | [145] | ||
| Antiadherent activity | [145] | ||
| CaOV3 |
| Induced autophagy | [150] | |
| Induced apoptosis | [150] | ||
| Induced apoptosis | [150] | ||
| A2780 |
| Antiproliferation activity | [148] | |
| Induced apoptosis | [148] | ||
| SKOV-3 |
| Antiproliferation activity | [148] | |
| Induced apoptosis | [148] | ||
| Antiproliferation activity | [151] | ||
| Antiproliferation activity | [151] | ||
| Fisetin | SKOV-3 |
| Induced apoptosis | [158] |
| Induced apoptosis | [158] | ||
| OVCAR-3 |
| Induced necrosis | [159] | |
| A2780 |
| Induced necrosis | [159] | |
| Induced apoptosis | [160] | ||
| Induced apoptosis | [160] | ||
| Galangin | OVCAR-3 |
| Induced apoptosis | [161] |
| Induced apoptosis | [161] | ||
| Antiproliferation activity | [161] | ||
| Induced apoptosis | [161] | ||
| Anti-invasion activity and antiangiogenesis | [161] | ||
| A2780/CP70 |
| Induced apoptosis | [161] | |
| Induced apoptosis | [161] | ||
| Induced apoptosis | [161] | ||
| Antiproliferation activity | [161] | ||
| Isorhamnetin | SKOV3 |
| Anti-invasion activity | [162] |
| Anti-invasion activity | [162] | ||
| Morin | A2780 |
| Antimigration | [163] |
| Antiadhesion | [163] | ||
| Antimigration | [163] | ||
| SKOV-3 |
| Antimigration | [163] | |
| Antiadhesion | [163] | ||
| Antimigration | [163] |
3.4. Fisetin
3.4.1. Effect of Fisetin on Breast Cancer
3.4.2. Effect of Fisetin on Ovarian Cancer
3.5. Galangin
3.5.1. Effect of Galangin on Breast Cancer
3.5.2. Effect of Galangin on Ovarian Cancer
3.5.3. Effect of Galangin on Endometrial Cancer
3.6. Isorhamnetin
3.6.1. Effect of Isorhamnetin on Breast Cancer
3.6.2. Effect of Isorhamnetin on Ovarian Cancer
3.6.3. Effect of Isorhamnetin on Endometrial Cancer
3.7. Morin
3.7.1. Effect of Morin on Breast Cancer
3.7.2. Effect of Morin on Ovarian Cancer
4. Summary
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hailu, H.E.; Mondul, A.M.; Rozek, L.S.; Geleta, T. Descriptive Epidemiology of Breast and Gynecological Cancers among Patients Attending Saint Paul’s Hospital Millennium Medical College, Ethiopia. PLoS ONE 2020, 15, e0230625. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Li, Z.; Tang, L.; Shen, M.; Zhou, Z.; Wei, Y.; Zhao, Y.; Bai, S.; Song, L. Associations of Dietary Intakes with Gynecological Cancers: Findings from a Cross-Sectional Study. Nutrients 2022, 14, 5026. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Arnold, M.; Morgan, E.; Rumgay, H.; Mafra, A.; Singh, D.; Laversanne, M.; Vignat, J.; Gralow, J.R.; Cardoso, F.; Siesling, S.; et al. Current and Future Burden of Breast Cancer: Global Statistics for 2020 and 2040. Breast 2022, 66, 15–23. [Google Scholar] [CrossRef] [PubMed]
- McDonald, E.S.; Clark, A.S.; Tchou, J.; Zhang, P.; Freedman, G.M. Clinical Diagnosis and Management of Breast Cancer. J. Nucl. Med. 2016, 57, 9S–16S. [Google Scholar] [CrossRef] [Green Version]
- Shien, T.; Iwata, H. Adjuvant and Neoadjuvant Therapy for Breast Cancer. Jpn. J. Clin. Oncol. 2020, 50, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Zamwar, U.M.; Anjankar, A.P. Aetiology, Epidemiology, Histopathology, Classification, Detailed Evaluation, and Treatment of Ovarian Cancer. Cureus 2022, 14, e30561. [Google Scholar] [CrossRef]
- Eisenhauer, E.A. Real-World Evidence in the Treatment of Ovarian Cancer. Ann. Oncol. 2017, 28, viii61–viii65. [Google Scholar] [CrossRef]
- Elias, K.M.; Guo, J.; Bast, R.C. Early Detection of Ovarian Cancer. Hematol. Oncol. Clin. N. Am. 2018, 32, 903–914. [Google Scholar] [CrossRef] [Green Version]
- Makker, V.; MacKay, H.; Ray-Coquard, I.; Levine, D.A.; Westin, S.N.; Aoki, D.; Oaknin, A. Endometrial Cancer. Nat. Rev. Dis. Primers 2021, 7, 88. [Google Scholar] [CrossRef]
- van den Heerik, A.S.V.M.; Horeweg, N.; de Boer, S.M.; Bosse, T.; Creutzberg, C.L. Adjuvant Therapy for Endometrial Cancer in the Era of Molecular Classification: Radiotherapy, Chemoradiation and Novel Targets for Therapy. Int. J. Gynecol. Cancer 2021, 31, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Trojano, G.; Olivieri, C.; Tinelli, R.; Damiani, G.R.; Pellegrino, A.; Cicinelli, E. Conservative Treatment in Early Stage Endometrial Cancer: A Review. Acta Biomed. 2019, 90, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska, A.; Rudzki, M.; Rudzki, S.; Lewandowski, T.; Laskowska, B. Environmental Risk Factors for Cancer—Review Paper. Ann. Agric. Environ. Med. 2019, 26, 1–7. [Google Scholar] [CrossRef]
- Forni, C.; Rossi, M.; Borromeo, I.; Feriotto, G.; Platamone, G.; Tabolacci, C.; Mischiati, C.; Beninati, S. Flavonoids: A Myth or a Reality for Cancer Therapy? Molecules 2021, 26, 3583. [Google Scholar] [CrossRef]
- Chambers, C.S.; Viktorová, J.; Řehořová, K.; Biedermann, D.; Turková, L.; Macek, T.; Křen, V.; Valentová, K. Defying Multidrug Resistance! Modulation of Related Transporters by Flavonoids and Flavonolignans. J. Agric. Food Chem. 2020, 68, 1763–1779. [Google Scholar] [CrossRef]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as Anticancer Agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef] [Green Version]
- Tao, H.; Li, L.; He, Y.; Zhang, X.; Zhao, Y.; Wang, Q.; Hong, G. Flavonoids in Vegetables: Improvement of Dietary Flavonoids by Metabolic Engineering to Promote Health. Crit. Rev. Food Sci. Nutr. 2022, Oct 11, 1–15. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids: An Overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Feng, Y.; Yu, S.; Fan, Z.; Li, X.; Li, J.; Yin, H. The Flavonoid Biosynthesis Network in Plants. Int. J. Mol. Sci. 2021, 22, 12824. [Google Scholar] [CrossRef]
- Peniche-Pavía, H.A.; Guzmán, T.J.; Magaña-Cerino, J.M.; Gurrola-Díaz, C.M.; Tiessen, A. Maize Flavonoid Biosynthesis, Regulation, and Human Health Relevance: A Review. Molecules 2022, 27, 5166. [Google Scholar] [CrossRef] [PubMed]
- Rudrapal, M.; Khan, J.; Dukhyil, A.A.B.; Alarousy, R.M.I.I.; Attah, E.I.; Sharma, T.; Khairnar, S.J.; Bendale, A.R. Chalcone Scaffolds, Bioprecursors of Flavonoids: Chemistry, Bioactivities, and Pharmacokinetics. Molecules 2021, 26, 7177. [Google Scholar] [CrossRef]
- Testai, L.; Calderone, V. Nutraceutical Value of Citrus Flavanones and Their Implications in Cardiovascular Disease. Nutrients 2017, 9, 502. [Google Scholar] [CrossRef] [Green Version]
- Espíndola, C. Some Nanocarrier’s Properties and Chemical Interaction Mechanisms with Flavones. Molecules 2023, 28, 2864. [Google Scholar] [CrossRef]
- Luo, Y.; Jian, Y.; Liu, Y.; Jiang, S.; Muhammad, D.; Wang, W. Flavanols from Nature: A Phytochemistry and Biological Activity Review. Molecules 2022, 27, 719. [Google Scholar] [CrossRef]
- Gervasi, T.; Calderaro, A.; Barreca, D.; Tellone, E.; Trombetta, D.; Ficarra, S.; Smeriglio, A.; Mandalari, G.; Gattuso, G. Biotechnological Applications and Health-Promoting Properties of Flavonols: An Updated View. Int. J. Mol. Sci. 2022, 23, 1710. [Google Scholar] [CrossRef]
- Saito, A. Challenges and Complexity of Functionality Evaluation of Flavan-3-Ol Derivatives. Biosci. Biotechnol. Biochem. 2017, 81, 1055–1060. [Google Scholar] [CrossRef] [Green Version]
- Di Pede, G.; Mena, P.; Bresciani, L.; Almutairi, T.M.; Del Rio, D.; Clifford, M.N.; Crozier, A. Human Colonic Catabolism of Dietary Flavan-3-Ol Bioactives. Mol. Aspects Med. 2023, 89, 101107. [Google Scholar] [CrossRef]
- Chagas, M.D.S.S.; Behrens, M.D.; Moragas-Tellis, C.J.; Penedo, G.X.M.; Silva, A.R.; Gonçalves-de-Albuquerque, C.F. Flavonols and Flavones as Potential Anti-Inflammatory, Antioxidant, and Antibacterial Compounds. Oxid. Med. Cell. Longev. 2022, 2022, 9966750. [Google Scholar] [CrossRef]
- Imran, M.; Salehi, B.; Sharifi-Rad, J.; Aslam Gondal, T.; Saeed, F.; Imran, A.; Shahbaz, M.; Tsouh Fokou, P.V.; Umair Arshad, M.; Khan, H.; et al. Kaempferol: A Key Emphasis to Its Anticancer Potential. Molecules 2019, 24, 2277. [Google Scholar] [CrossRef] [Green Version]
- Bhosale, P.B.; Ha, S.E.; Vetrivel, P.; Kim, H.H.; Kim, S.M.; Kim, G.S. Functions of Polyphenols and Its Anticancer Properties in Biomedical Research: A Narrative Review. Transl. Cancer Res. TCR 2020, 9, 7619–7631. [Google Scholar] [CrossRef] [PubMed]
- Kubina, R.; Iriti, M.; Kabała-Dzik, A. Anticancer Potential of Selected Flavonols: Fisetin, Kaempferol, and Quercetin on Head and Neck Cancers. Nutrients 2021, 13, 845. [Google Scholar] [CrossRef]
- Ren, J.; Lu, Y.; Qian, Y.; Chen, B.; Wu, T.; Ji, G. Recent Progress Regarding Kaempferol for the Treatment of Various Diseases (Review). Exp. Ther. Med. 2019, 18, 2759–2776. [Google Scholar] [CrossRef] [Green Version]
- Dabeek, W.M.; Marra, M.V. Dietary Quercetin and Kaempferol: Bioavailability and Potential Cardiovascular-Related Bioactivity in Humans. Nutrients 2019, 11, 2288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, W.; Khan, H.; Shah, M.A.; Cauli, O.; Saso, L. Kaempferol as a Dietary Anti-Inflammatory Agent: Current Therapeutic Standing. Molecules 2020, 25, 4073. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; Chen, L.; Ma, X.; Jiao, R.; Li, X.; Wang, Y. Protective Effects of Kaempferol against Reactive Oxygen Species-Induced Hemolysis and Its Antiproliferative Activity on Human Cancer Cells. Eur. J. Med. Chem. 2016, 114, 24–32. [Google Scholar] [CrossRef]
- Saw, C.L.L.; Guo, Y.; Yang, A.Y.; Paredes-Gonzalez, X.; Ramirez, C.; Pung, D.; Kong, A.-N.T. The Berry Constituents Quercetin, Kaempferol, and Pterostilbene Synergistically Attenuate Reactive Oxygen Species: Involvement of the Nrf2-ARE Signaling Pathway. Food Chem. Toxicol. 2014, 72, 303–311. [Google Scholar] [CrossRef]
- Kashyap, D.; Sharma, A.; Tuli, H.S.; Sak, K.; Punia, S.; Mukherjee, T.K. Kaempferol—A Dietary Anticancer Molecule with Multiple Mechanisms of Action: Recent Trends and Advancements. J. Funt. Foods 2017, 30, 203–219. [Google Scholar] [CrossRef] [PubMed]
- Periferakis, A.; Periferakis, K.; Badarau, I.A.; Petran, E.M.; Popa, D.C.; Caruntu, A.; Costache, R.S.; Scheau, C.; Caruntu, C.; Costache, D.O. Kaempferol: Antimicrobial Properties, Sources, Clinical, and Traditional Applications. Int. J. Mol. Sci. 2022, 23, 15054. [Google Scholar] [CrossRef]
- Crocetto, F.; di Zazzo, E.; Buonerba, C.; Aveta, A.; Pandolfo, S.D.; Barone, B.; Trama, F.; Caputo, V.F.; Scafuri, L.; Ferro, M.; et al. Kaempferol, Myricetin and Fisetin in Prostate and Bladder Cancer: A Systematic Review of the Literature. Nutrients 2021, 13, 3750. [Google Scholar] [CrossRef]
- Wang, F.; Wang, L.; Qu, C.; Chen, L.; Geng, Y.; Cheng, C.; Yu, S.; Wang, D.; Yang, L.; Meng, Z.; et al. Kaempferol Induces ROS-Dependent Apoptosis in Pancreatic Cancer Cells via TGM2-Mediated Akt/MTOR Signaling. BMC Cancer 2021, 21, 396. [Google Scholar] [CrossRef]
- Saraei, R.; Rahman, H.S.; Soleimani, M.; Asghari-Jafarabadi, M.; Naimi, A.; Hassanzadeh, A.; Solali, S. Kaempferol Sensitizes Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand-Resistance Chronic Myelogenous Leukemia Cells to Apoptosis. Mol. Biol. Rep. 2022, 49, 19–29. [Google Scholar] [CrossRef]
- Kim, T.W.; Lee, S.Y.; Kim, M.; Cheon, C.; Ko, S.-G. Kaempferol Induces Autophagic Cell Death via IRE1-JNK-CHOP Pathway and Inhibition of G9a in Gastric Cancer Cells. Cell. Death Dis. 2018, 9, 875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qattan, M.Y.; Khan, M.I.; Alharbi, S.H.; Verma, A.K.; Al-Saeed, F.A.; Abduallah, A.M.; Al Areefy, A.A. Therapeutic Importance of Kaempferol in the Treatment of Cancer through the Modulation of Cell Signalling Pathways. Molecules 2022, 27, 8864. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-H.; Hwang, K.-A.; Choi, K.-C. Treatment with Kaempferol Suppresses Breast Cancer Cell Growth Caused by Estrogen and Triclosan in Cellular and Xenograft Breast Cancer Models. J. Nutr. Biochem. 2016, 28, 70–82. [Google Scholar] [CrossRef]
- Ackland, M.L. Synergistic Antiproliferative Action of the Flavonols Quercetin and Kaempferol in Cultured Human Cancer Cell Lines. In Vivo 2005, 19, 69–76. [Google Scholar]
- Yi, X.; Zuo, J.; Tan, C.; Xian, S.; Luo, C.; Chen, S.; Yu, L.; Luo, Y. Kaempferol, a flavonoid compound from gynura medica induced apoptosis and growth inhibition in mcf-7 breast cancer cell. Afr. J. Tradit. Complement. Altern. Med. 2016, 13, 210–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.-A.; Choi, K.-C.; Hwang, K.-A. Treatment with Phytoestrogens Reversed Triclosan and Bisphenol A-Induced Anti-Apoptosis in Breast Cancer Cells. Biomol. Ther. 2018, 26, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Ayob, Z.; Mohd Bohari, S.P.; Abd Samad, A.; Jamil, S. Cytotoxic Activities against Breast Cancer Cells of Local Justicia Gendarussa Crude Extracts. Evid. Based Complement. Alternat. Med. 2014, 2014, 732980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phromnoi, K.; Yodkeeree, S.; Anuchapreeda, S.; Limtrakul, P. Inhibition of MMP-3 Activity and Invasion of the MDA-MB-231 Human Invasive Breast Carcinoma Cell Line by Bioflavonoids. Acta Pharmacol. Sin. 2009, 30, 1169–1176. [Google Scholar] [CrossRef] [Green Version]
- Chenglin Li 1, Yuanwei Zhao, Dan Yang, Yanyan Yu, Hao Guo, Ziming Zhao, Bei Zhang, Xiaoxing Yin Inhibitory Effects of Kaempferol on the Invasion of Human Breast Carcinoma Cells by Downregulating the Expression and Activity of Matrix Metalloproteinase-9. Biochem. Cell. Biol. 2015, 93, 16–27. [CrossRef]
- Zhu, L.; Xue, L. Kaempferol Suppresses Proliferation and Induces Cell Cycle Arrest, Apoptosis, and DNA Damage in Breast Cancer Cells. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2019, 27, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yan, T.; Deng, R.; Jiang, X.; Xiong, H.; Wang, Y.; Yu, Q.; Wang, X.; Chen, C.; Zhu, Y. Low Dose of Kaempferol Suppresses the Migration and Invasion of Triple-Negative Breast Cancer Cells by Downregulating the Activities of RhoA and Rac1. Onco Targets Ther. 2017, 10, 4809–4819. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Zhang, Y.; Xin, X.; Huang, G.; Zhang, N.; Zeng, Q.; Tang, L.; Attaribo, T.; Lee, K.S.; Jin, B.R.; et al. Dual-Targeting Antiproliferation Hybrids Derived from 1-Deoxynojirimycin and Kaempferol Induce MCF-7 Cell Apoptosis through the Mitochondria-Mediated Pathway. J. Nat. Prod. 2021, 84, 1534–1543. [Google Scholar] [CrossRef]
- Luo, H.; Rankin, G.O.; Li, Z.; DePriest, L.; Chen, Y.C. Kaempferol Induces Apoptosis in Ovarian Cancer Cells through Activating P53 in the Intrinsic Pathway. Food Chem. 2011, 128, 513–519. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Si, L.; Jia, Y.; Jian, W.; Yu, Q.; Wang, M.; Lin, R. Kaempferol Exerts Anti-Proliferative Effects on Human Ovarian Cancer Cells by Inducing Apoptosis, G0/G1 Cell Cycle Arrest and Modulation of MEK/ERK and STAT3 Pathways. J. BUON 2019, 24, 975–981. [Google Scholar]
- Gao, Y.; Yin, J.; Rankin, G.; Chen, Y. Kaempferol Induces G2/M Cell Cycle Arrest via Checkpoint Kinase 2 and Promotes Apoptosis via Death Receptors in Human Ovarian Carcinoma A2780/CP70 Cells. Molecules 2018, 23, 1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Tian, B.; Wang, Y.; Ding, H. Kaempferol Sensitizes Human Ovarian Cancer Cells-OVCAR-3 and SKOV-3 to Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand (TRAIL)-Induced Apoptosis via JNK/ERK-CHOP Pathway and Up-Regulation of Death Receptors 4 and 5. Med. Sci. Monit. 2017, 23, 5096–5105. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; Daddysman, M.K.; Rankin, G.O.; Jiang, B.-H.; Chen, Y.C. Kaempferol Enhances Cisplatin’s Effect on Ovarian Cancer Cells through Promoting Apoptosis Caused by down Regulation of CMyc. Cancer Cell. Int. 2010, 10, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, H.; Rankin, G.O.; Liu, L.; Daddysman, M.K.; Jiang, B.-H.; Chen, Y.C. Kaempferol Inhibits Angiogenesis and VEGF Expression Through Both HIF Dependent and Independent Pathways in Human Ovarian Cancer Cells. Nutr. Cancer 2009, 61, 554–563. [Google Scholar] [CrossRef]
- El-Kott, A.F.; Shati, A.A.; Al-Kahtani, M.A.; Alharbi, S.A. Kaempferol Induces Cell Death in A2780 Ovarian Cancer Cells and Increases Their Sensitivity to Cisplatin by Activation of Cytotoxic Endoplasmic Reticulum-Mediated Autophagy and Inhibition of Protein Kinase B. Folia Biol. 2020, 66, 36–46. [Google Scholar]
- Lei, X.; Guo, J.; Wang, Y.; Cui, J.; Feng, B.; Su, Y.; Zhao, H.; Yang, W.; Hu, Y. Inhibition of Endometrial Carcinoma by Kaempferol Is Interceded through Apoptosis Induction, G2/M Phase Cell Cycle Arrest, Suppression of Cell Invasion and Upregulation of m-TOR/PI3K Signalling Pathway. J. BUON 2019, 24, 1555–1561. [Google Scholar] [PubMed]
- Chuwa, A.; Sone, K.; Oda, K.; Tanikawa, M.; Kukita, A.; Kojima, M.; Oki, S.; Fukuda, T.; Takeuchi, M.; Miyasaka, A.; et al. Kaempferol, a Natural Dietary Flavonoid, Suppresses 17β-estradiol-induced Survivin Expression and Causes Apoptotic Cell Death in Endometrial Cancer. Oncol. Lett. 2018, 16, 6195–6201. [Google Scholar] [CrossRef] [Green Version]
- Ruan, G.-Y.; Ye, L.-X.; Lin, J.-S.; Lin, H.-Y.; Yu, L.-R.; Wang, C.-Y.; Mao, X.-D.; Zhang, S.-H.; Sun, P.-M. An Integrated Approach of Network Pharmacology, Molecular Docking, and Experimental Verification Uncovers Kaempferol as the Effective Modulator of HSD17B1 for Treatment of Endometrial Cancer. J. Transl. Med. 2023, 21, 204. [Google Scholar] [CrossRef]
- Soleimani, M.; Sajedi, N. Myricetin Apoptotic Effects on T47D Breast Cancer Cells Is a P53-Independent Approach. Asian. Pac. J. Cancer Prev. 2020, 21, 3697–3704. [Google Scholar] [CrossRef]
- Sajedi, N.; Homayoun, M.; Mohammadi, F.; Soleimani, M. Myricetin Exerts Its Apoptotic Effects on MCF-7 Breast Cancer Cells through Evoking the BRCA1-GADD45 Pathway. Asian Pac. J. Cancer Prev. 2020, 21, 3461–3468. [Google Scholar] [CrossRef] [PubMed]
- Jiao, D.; Zhang, X.D. Myricetin Suppresses P21-Activated Kinase 1 in Human Breast Cancer MCF-7 Cells through Downstream Signaling of the β-Catenin Pathway. Oncol. Rep. 2016, 36, 342–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knickle, A.; Fernando, W.; Greenshields, A.L.; Rupasinghe, H.P.V.; Hoskin, D.W. Myricetin-Induced Apoptosis of Triple-Negative Breast Cancer Cells Is Mediated by the Iron-Dependent Generation of Reactive Oxygen Species from Hydrogen Peroxide. Food Chem. Toxicol. 2018, 118, 154–167. [Google Scholar] [CrossRef]
- Ci, Y.; Zhang, Y.; Liu, Y.; Lu, S.; Cao, J.; Li, H.; Zhang, J.; Huang, Z.; Zhu, X.; Gao, J.; et al. Myricetin Suppresses Breast Cancer Metastasis through Down-Regulating the Activity of Matrix Metalloproteinase (MMP)-2/9. Phytother. Res. 2018, 32, 1373–1381. [Google Scholar] [CrossRef]
- Han, S.-H.; Lee, J.-H.; Woo, J.-S.; Jung, G.-H.; Jung, S.-H.; Han, E.-J.; Park, Y.-S.; Kim, B.-S.; Kim, S.-K.; Park, B.-K.; et al. Myricetin Induces Apoptosis through the MAPK Pathway and Regulates JNK-mediated Autophagy in SK-BR-3 Cells. Int. J. Mol. Med. 2022, 49, 54. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhou, N.; Wang, J.; Liu, Z.; Wang, X.; Zhang, Q.; Liu, Q.; Gao, L.; Wang, R. Quercetin Suppresses Breast Cancer Stem Cells (CD44+/CD24−) by Inhibiting the PI3K/Akt/MTOR-Signaling Pathway. Life Sci. 2018, 196, 56–62. [Google Scholar] [CrossRef]
- Jia, L.; Huang, S.; Yin, X.; Zan, Y.; Guo, Y.; Han, L. Quercetin Suppresses the Mobility of Breast Cancer by Suppressing Glycolysis through Akt-MTOR Pathway Mediated Autophagy Induction. Life Sci. 2018, 208, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Ranganathan, S.; Halagowder, D.; Sivasithambaram, N.D. Quercetin Suppresses Twist to Induce Apoptosis in MCF-7 Breast Cancer Cells. PLoS ONE 2015, 10, e0141370. [Google Scholar] [CrossRef] [Green Version]
- Chou, C.-C.; Yang, J.-S.; Lu, H.-F.; Ip, S.-W.; Lo, C.; Wu, C.-C.; Lin, J.-P.; Tang, N.-Y.; Chung, J.-G.; Chou, M.-J.; et al. Quercetin-Mediated Cell Cycle Arrest and Apoptosis Involving Activation of a Caspase Cascade through the Mitochondrial Pathway in Human Breast Cancer MCF-7 Cells. Arch. Pharm. Res. 2010, 33, 1181–1191. [Google Scholar] [CrossRef] [PubMed]
- Khorsandi, L.; Orazizadeh, M.; Niazvand, F.; Abbaspour, M.R.; Mansouri, E.; Khodadadi, A. Quercetin Induces Apoptosis and Necroptosis in MCF-7 Breast Cancer Cells. Bratisl. Lek. Listy 2017, 118, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Rhman, M.A.; Devnarain, N.; Khan, R.; Owira, P.M.O. Synergism Potentiates Oxidative Antiproliferative Effects of Naringenin and Quercetin in MCF-7 Breast Cancer Cells. Nutrients 2022, 14, 3437. [Google Scholar] [CrossRef]
- Tao, S.; He, H.; Chen, Q. Quercetin Inhibits Proliferation and Invasion Acts by Up-Regulating MiR-146a in Human Breast Cancer Cells. Mol. Cell. Biochem. 2015, 402, 93–100. [Google Scholar] [CrossRef]
- Khan, M.A.; Singh, R.; Siddiqui, S.; Ahmad, I.; Ahmad, R.; Upadhyay, S.; Barkat, A.; Ali, A.M.A.; Zia, Q.; Srivastava, A.; et al. Anticancer Potential of Phoenix Dactylifera L. Seed Extract in Human Cancer Cells and pro-Apoptotic Effects Mediated through Caspase-3 Dependent Pathway in Human Breast Cancer MDA-MB-231 Cells: An in Vitro and in Silico Investigation. BMC Complement. Med. Ther. 2022, 22, 68. [Google Scholar] [CrossRef]
- Alhakamy, N.A.; Fahmy, U.A.; Eldin, S.M.B.; Ahmed, O.A.A.; Aldawsari, H.M.; Okbazghi, S.Z.; Alfaleh, M.A.; Abdulaal, W.H.; Alamoudi, A.J.; Mady, F.M. Scorpion Venom-Functionalized Quercetin Phytosomes for Breast Cancer Management: In Vitro Response Surface Optimization and Anticancer Activity against MCF-7 Cells. Polymers 2021, 14, 93. [Google Scholar] [CrossRef] [PubMed]
- Karimian, A.; Majidinia, M.; Moliani, A.; Alemi, F.; Asemi, Z.; Yousefi, B.; Naghibi, A.F. The Modulatory Effects of Two Bioflavonoids, Quercetin and Thymoquinone on the Expression Levels of DNA Damage and Repair Genes in Human Breast, Lung and Prostate Cancer Cell Lines. Pathol. Res. Pract. 2022, 240, 154143. [Google Scholar] [CrossRef]
- Manouchehri, J.M.; Turner, K.A.; Kalafatis, M. TRAIL-Induced Apoptosis in TRAIL-Resistant Breast Carcinoma Through Quercetin Cotreatment. Breast Cancer 2018, 12, 117822341774985. [Google Scholar] [CrossRef]
- Qiu, D.; Yan, X.; Xiao, X.; Zhang, G.; Wang, Y.; Cao, J.; Ma, R.; Hong, S.; Ma, M. To Explore Immune Synergistic Function of Quercetin in Inhibiting Breast Cancer Cells. Cancer Cell. Int. 2021, 21, 632. [Google Scholar] [CrossRef]
- Rivera Rivera, A.; Castillo-Pichardo, L.; Gerena, Y.; Dharmawardhane, S. Anti-Breast Cancer Potential of Quercetin via the Akt/AMPK/Mammalian Target of Rapamycin (MTOR) Signaling Cascade. PLoS ONE 2016, 11, e0157251. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Lee, Y.-H.; Sharma, A.R.; Park, J.-B.; Jagga, S.; Sharma, G.; Lee, S.-S.; Nam, J.-S. Quercetin Induces Apoptosis and Cell Cycle Arrest in Triple-Negative Breast Cancer Cells through Modulation of Foxo3a Activity. Korean J. Physiol. Pharmacol. 2017, 21, 205. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.-J.; Tsai, J.-H.; Hsu, L.-S.; Lin, C.-L.; Hong, H.-M.; Pan, M.-H. Quercetin Blocks the Aggressive Phenotype of Triple Negative Breast Cancer by Inhibiting IGF1/IGF1R-Mediated EMT Program. J. Food Drug. Anal. 2021, 29, 98–112. [Google Scholar] [CrossRef]
- Wang, L.; Du, X.; Yue, D.; Chen, X. Catechin, Rutin and Quercetin in Quercus Mongolica Fisch Leaves Exert Inhibitory Effects on Multiple Cancer Cells. J. Food Biochem. 2022, 46, e14486. [Google Scholar] [CrossRef]
- Seo, H.-S.; Ku, J.M.; Choi, H.-S.; Choi, Y.K.; Woo, J.-K.; Kim, M.; Kim, I.; Na, C.H.; Hur, H.; Jang, B.-H.; et al. Quercetin Induces Caspase-Dependent Extrinsic Apoptosis through Inhibition of Signal Transducer and Activator of Transcription 3 Signaling in HER2-Overexpressing BT-474 Breast Cancer Cells. Oncol. Rep. 2016, 36, 31–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.-F.; Chen, J.-H.; Chang, C.-N.; Lu, D.-Y.; Chang, P.-C.; Wang, S.-L.; Yeh, W.-L. Fisetin Inhibits Cell Migration via Inducing HO-1 and Reducing MMPs Expression in Breast Cancer Cell Lines. Food Chem. Toxicol. 2018, 120, 528–535. [Google Scholar] [CrossRef]
- Sun, X.; Ma, X.; Li, Q.; Yang, Y.; Xu, X.; Sun, J.; Yu, M.; Cao, K.; Yang, L.; Yang, G.; et al. Anti-cancer Effects of Fisetin on Mammary Carcinoma Cells via Regulation of the PI3K/Akt/MTOR Pathway: In vitro and In vivo Studies. Int. J. Mol. Med. 2018, 42, 811–820. [Google Scholar] [CrossRef] [Green Version]
- Hosseini, S.S.; Ebrahimi, S.O.; Haji Ghasem Kashani, M.; Reiisi, S. Study of Quercetin and Fisetin Synergistic Effect on Breast Cancer and Potentially Involved Signaling Pathways. Cell. Biol. Int. 2023, 47, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.L.; Murphy, K.; Doucette, C.D.; Greenshields, A.L.; Hoskin, D.W. The Dietary Flavonoid Fisetin Causes Cell Cycle Arrest, Caspase-Dependent Apoptosis, and Enhanced Cytotoxicity of Chemotherapeutic Drugs in Triple-Negative Breast Cancer Cells. J. Cell. Biochem. 2016, 117, 1913–1925. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Gong, X.; Jiang, R.; Lin, D.; Zhou, T.; Zhang, A.; Li, H.; Zhang, X.; Wan, J.; Kuang, G.; et al. Fisetin Inhibited Growth and Metastasis of Triple-Negative Breast Cancer by Reversing Epithelial-to-Mesenchymal Transition via PTEN/Akt/GSK3β Signal Pathway. Front. Pharmacol. 2018, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, G.; Zhang, W.; Dang, M.; Yan, M.; Chen, Z. Fisetin Induces Apoptosis in Breast Cancer MDA-MB-453 Cells through Degradation of HER2/Neu and via the PI3K/Akt Pathway. J. Biochem. Mol. Toxicol. 2019, 33, e22268. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Yan, C.; Zhou, Q.; Zhen, L. Galangin Potentiates Human Breast Cancer to Apoptosis Induced byTRAIL through Activating AMPK. Biomed. Pharmacother. 2017, 89, 845–856. [Google Scholar] [CrossRef]
- Liu, D.; You, P.; Luo, Y.; Yang, M.; Liu, Y. Galangin Induces Apoptosis in MCF-7 Human Breast Cancer Cells Through Mitochondrial Pathway and Phosphatidylinositol 3-Kinase/Akt Inhibition. Pharmacology 2018, 102, 58–66. [Google Scholar] [CrossRef]
- Qaddoori, M.H.; Al-Shmgani, H.S. Galangin-Loaded Gold Nanoparticles: Molecular Mechanisms of Antiangiogenesis Properties in Breast Cancer. Int. J. Breast Cancer. 2023, 2023, 3251211. [Google Scholar] [CrossRef]
- Murray, T.J.; Yang, X.; Sherr, D.H. Growth of a Human Mammary Tumor Cell Line Is Blocked by Galangin, a Naturally Occurring Bioflavonoid, and Is Accompanied by down-Regulation of Cyclins D3, E, and A. Breast Cancer Res. 2006, 8, R17. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.-W.; Hu, J.-J.; Fu, R.-Q.; Liu, X.; Zhang, Y.-H.; Li, J.; Liu, L.; Li, Y.-N.; Deng, Q.; Luo, Q.-S.; et al. Flavonoids Inhibit Cell Proliferation and Induce Apoptosis and Autophagy through Downregulation of PI3Kγ Mediated PI3K/AKT/MTOR/P70S6K/ULK Signaling Pathway in Human Breast Cancer Cells. Sci. Rep. 2018, 8, 11255. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Huang, L.; Meng, L.; Sun, H.; Zhang, W.; Xu, Y. Isorhamnetin Inhibits Cell Proliferation and Induces Apoptosis in Breast Cancer via Akt and Mitogen-Activated Protein Kinase Kinase Signaling Pathways. Mol. Med. Rep. 2015, 12, 6745–6751. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-H.; Jin, H.; Lee, W.S.; Nagappan, A.; Hyun, Y.; Kim, G.; Jung, J.-M.; Ryu, C.H.; Shin, S.C.; Hong, C.; et al. Morin, a Flavonoid from Moraceae, Inhibits Cancer Cell Adhesion to Endothelial Cells and EMT by Down-Regulating VCAM-1 and N-Cadherin. Asian Pac. J. Cancer Prev. 2016, 17, 3071–3075. [Google Scholar] [PubMed]
- Jin, H.; Lee, W.S.; Eun, S.Y.; Jung, J.H.; Park, H.-S.; Kim, G.; Choi, Y.H.; Ryu, C.H.; Jung, J.M.; Hong, S.C.; et al. Morin, a Flavonoid from Moraceae, Suppresses Growth and Invasion of the Highly Metastatic Breast Cancer Cell Line MDA-MB-231 Partly through Suppression of the Akt Pathway. Int. J. Oncol. 2014, 45, 1629–1637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agraharam, G.; Girigoswami, A.; Girigoswami, K. Myricetin: A Multifunctional Flavonol in Biomedicine. Curr. Pharmacol. Rep. 2022, 8, 48–61. [Google Scholar] [CrossRef]
- Arafah, A.; Rehman, M.U.; Ahmad, A.; AlKharfy, K.M.; Alqahtani, S.; Jan, B.L.; Almatroudi, N.M. Myricetin (3,3′,4′,5,5′,7-Hexahydroxyflavone) Prevents 5-Fluorouracil-Induced Cardiotoxicity. ACS Omega 2022, 7, 4514–4524. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Saeed, F.; Hussain, G.; Imran, A.; Mehmood, Z.; Gondal, T.A.; El-Ghorab, A.; Ahmad, I.; Pezzani, R.; Arshad, M.U.; et al. Myricetin: A Comprehensive Review on Its Biological Potentials. Food. Sci. Nutr. 2021, 9, 5854–5868. [Google Scholar] [CrossRef] [PubMed]
- Semwal, D.; Semwal, R.; Combrinck, S.; Viljoen, A. Myricetin: A Dietary Molecule with Diverse Biological Activities. Nutrients 2016, 8, 90. [Google Scholar] [CrossRef] [Green Version]
- Javed, Z.; Khan, K.; Herrera-Bravo, J.; Naeem, S.; Iqbal, M.J.; Raza, Q.; Sadia, H.; Raza, S.; Bhinder, M.; Calina, D.; et al. Myricetin: Targeting Signaling Networks in Cancer and Its Implication in Chemotherapy. Cancer Cell. Int. 2022, 22, 239. [Google Scholar] [CrossRef]
- Qian, J.; Zhang, J.; Chen, Y.; Dai, C.; Fan, J.; Guo, H. Hypoglycemic Activity and Mechanisms of Myricetin. Nat. Prod. Res. 2022, 36, 6177–6180. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Mao, W.; Li, Y.; Qi, C.; He, Y. Myricetin Inhibits Breast Tumor Growth and Angiogenesis by Regulating VEGF/VEGFR2 and P38MAPK Signaling Pathways. Anat. Rec. 2019, 302, 2186–2192. [Google Scholar] [CrossRef]
- Benarba, B.; Elmallah, A.; Pandiella, A. Bryoniadioica Aqueous Extract Induces Apoptosis and G2/M Cell Cycle Arrest in MDA-MB 231 Breast Cancer Cells. Mol. Med. Rep. 2019, 20, 73–80. [Google Scholar] [CrossRef]
- Maroufi, N.F.; Vahedian, V.; Mazrakhondi, S.A.M.; Kooti, W.; Khiavy, H.A.; Bazzaz, R.; Ramezani, F.; Pirouzpanah, S.M.; Ghorbani, M.; Akbarzadeh, M.; et al. Sensitization of MDA-MBA231 Breast Cancer Cell to Docetaxel by Myricetin Loaded into Biocompatible Lipid Nanoparticles via Sub-G1 Cell Cycle Arrest Mechanism. Naunyn-Schmiedeberg’s Arch. Pharm. 2020, 393, 1–11. [Google Scholar] [CrossRef]
- Mohan, U.P.; Sriram, B.; Panneerselvam, T.; Devaraj, S.; MubarakAli, D.; Parasuraman, P.; Palanisamy, P.; Premanand, A.; Arunachalam, S.; Kunjiappan, S. Utilization of Plant-Derived Myricetin Molecule Coupled with Ultrasound for the Synthesis of Gold Nanoparticles against Breast Cancer. Naunyn-Schmiedeberg’s Arch. Pharm. 2020, 393, 1963–1976. [Google Scholar] [CrossRef]
- Li, Q.; Tan, Q.; Ma, Y.; Gu, Z.; Chen, S. Myricetin Suppresses Ovarian Cancer In Vitro by Activating the P38/Sapla Signaling Pathway and Suppressing Intracellular Oxidative Stress. Front. Oncol. 2022, 12, 903394. [Google Scholar] [CrossRef]
- Huang, H.; Chen, A.Y.; Ye, X.; Li, B.; Rojanasakul, Y.; Rankin, G.O.; Chen, Y.C. Myricetin Inhibits Proliferation of Cisplatin-Resistant Cancer Cells through a P53-Dependent Apoptotic Pathway. Int. J. Oncol. 2015, 47, 1494–1502. [Google Scholar] [CrossRef] [Green Version]
- Zheng, A.-W.; Chen, Y.-Q.; Zhao, L.-Q.; Feng, J.-G. Myricetin Induces Apoptosis and Enhances Chemosensitivity in Ovarian Cancer Cells. Oncol. Lett. 2017, 13, 4974–4978. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, S.; Wei, C.; Rankin, G.O.; Ye, X.; Chen, Y.C. Flavonoids from Chinese Bayberry Leaves Induced Apoptosis and G1 Cell Cycle Arrest via Erk Pathway in Ovarian Cancer Cells. Eur. J. Med. Chem. 2018, 147, 218–226. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Xie, Q.; Wu, S.; Yi, D.; Yu, Y.; Liu, S.; Li, S.; Li, Z. Myricetin Induces Apoptosis via Endoplasmic Reticulum Stress and DNA Double-Strand Breaks in Human Ovarian Cancer Cells. Mol. Med. Rep. 2016, 13, 2094–2100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Chen, A.Y.; Rojanasakul, Y.; Ye, X.; Rankin, G.O.; Chen, Y.C. Dietary Compounds Galangin and Myricetin Suppress Ovarian Cancer Cell Angiogenesis. J. Funct. Foods 2015, 15, 464–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varela-Rodríguez, L.; Sánchez-Ramírez, B.; Hernández-Ramírez, V.I.; Varela-Rodríguez, H.; Castellanos-Mijangos, R.D.; González-Horta, C.; Chávez-Munguía, B.; Talamás-Rohana, P. Effect of Gallic Acid and Myricetin on Ovarian Cancer Models: A Possible Alternative Antitumoral Treatment. BMC Complement. Med. Ther. 2020, 20, 110. [Google Scholar] [CrossRef]
- Andres, S.; Pevny, S.; Ziegenhagen, R.; Bakhiya, N.; Schäfer, B.; Hirsch-Ernst, K.I.; Lampen, A. Safety Aspects of the Use of Quercetin as a Dietary Supplement. Mol. Nutr. Food Res. 2018, 62, 1700447. [Google Scholar] [CrossRef]
- Alizadeh, S.R.; Ebrahimzadeh, M.A. Quercetin Derivatives: Drug Design, Development, and Biological Activities, a Review. Eur. J. Med. Chem. 2022, 229, 114068. [Google Scholar] [CrossRef]
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M.; Wang, S.; Liu, H.; Yin, Y. Quercetin, Inflammation and Immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauf, A.; Imran, M.; Khan, I.A.; ur-Rehman, M.-; Gilani, S.A.; Mehmood, Z.; Mubarak, M.S. Anticancer Potential of Quercetin: A Comprehensive Review: Quercetin as an Anticancer Agent. Phytother. Res. 2018, 32, 2109–2130. [Google Scholar] [CrossRef]
- Sato, S.; Mukai, Y. Modulation of Chronic Inflammation by Quercetin: The Beneficial Effects on Obesity. J. Inflamm. Res. 2020, 13, 421–431. [Google Scholar] [CrossRef]
- Hashemzaei, M.; Far, A.D.; Yari, A.; Heravi, R.E.; Tabrizian, K.; Taghdisi, S.M.; Sadegh, S.E.; Tsarouhas, K.; Kouretas, D.; Tzanakakis, G.; et al. Anticancer and Apoptosis-Inducing Effects of Quercetin in Vitro and in Vivo. Oncol. Rep. 2017, 38, 819–828. [Google Scholar] [CrossRef] [Green Version]
- Aghapour, F.; Moghadamnia, A.A.; Nicolini, A.; Kani, S.N.M.; Barari, L.; Morakabati, P.; Rezazadeh, L.; Kazemi, S. Quercetin Conjugated with Silica Nanoparticles Inhibits Tumor Growth in MCF-7 Breast Cancer Cell Lines. Biochem. Biophys. Res. Commun. 2018, 500, 860–865. [Google Scholar] [CrossRef] [PubMed]
- Kabała-Dzik, A.; Rzepecka-Stojko, A.; Kubina, R.; Iriti, M.; Wojtyczka, R.D.; Buszman, E.; Stojko, J. Flavonoids, Bioactive Components of Propolis, Exhibit Cytotoxic Activity and Induce Cell Cycle Arrest and Apoptosis in Human Breast Cancer Cells MDA-MB-231 and MCF-7—A Comparative Study. Cell. Mol. Biol. 2018, 64, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Q.; Yang, L.; Yang, H.; Han, Y.; Chen, Y.; He, Y. Quercetin Alleviates the Progression of Breast Cancer-Related Depression via Inhibiting the Pyroptosis and Promoting the Immune Response. Mediat. Inflamm. 2022, 2022, 8011988. [Google Scholar] [CrossRef]
- Kıyga, E.; Şengelen, A.; Adıgüzel, Z.; Önay Uçar, E. Investigation of the Role of Quercetin as a Heat Shock Protein Inhibitor on Apoptosis in Human Breast Cancer Cells. Mol. Biol. Rep. 2020, 47, 4957–4967. [Google Scholar] [CrossRef]
- An, S.; Hu, M. Quercetin Promotes TFEB Nuclear Translocation and Activates Lysosomal Degradation of Ferritin to Induce Ferroptosis in Breast Cancer Cells. Comput. Intell. Neurosci. 2022, 2022, 5299218. [Google Scholar] [CrossRef]
- Wang, R.; Yang, L.; Li, S.; Ye, D.; Yang, L.; Liu, Q.; Zhao, Z.; Cai, Q.; Tan, J.; Li, X. Quercetin Inhibits Breast Cancer Stem Cells via Downregulation of Aldehyde Dehydrogenase 1A1 (ALDH1A1), Chemokine Receptor Type 4 (CXCR4), Mucin 1 (MUC1), and Epithelial Cell Adhesion Molecule (EpCAM). Med. Sci. Monit. 2018, 24, 412–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Kroon, P.A.; Shao, H.; Needs, P.W.; Yang, X. Differential Effects of Quercetin and Two of Its Derivatives, Isorhamnetin and Isorhamnetin-3-Glucuronide, in Inhibiting the Proliferation of Human Breast-Cancer MCF-7 Cells. J. Agric. Food Chem. 2018, 66, 7181–7189. [Google Scholar] [CrossRef] [PubMed]
- Azizi, E.; Fouladdel, S.; Komeili Movahhed, T.; Modaresi, F.; Barzegar, E.; Ghahremani, M.H.; Ostad, S.N.; Atashpour, S. Quercetin Effects on Cell Cycle Arrest and Apoptosis and Doxorubicin Activity in T47D Cancer Stem Cells. Asian Pac. J. Cancer Prev. 2022, 23, 4145–4154. [Google Scholar] [CrossRef]
- Ruidas, B.; Sur, T.K.; Das Mukhopadhyay, C.; Sinha, K.; Som Chaudhury, S.; Sharma, P.; Bhowmick, S.; Majumder, R.; Saha, A. Quercetin: A Silent Retarder of Fatty Acid Oxidation in Breast Cancer Metastasis through Steering of Mitochondrial CPT1. Breast Cancer 2022, 29, 748–760. [Google Scholar] [CrossRef]
- Mostafavi-Pour, Z.; Ramezani, F.; Keshavarzi, F.; Samadi, N. The Role of Quercetin and Vitamin C in Nrf2-Dependent Oxidative Stress Production in Breast Cancer Cells. Oncol. Lett. 2017, 13, 1965–1973. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, F.; Ijaz, B.; Ahmad, Z.; Farooq, N.; Sarwar, M.B.; Husnain, T. Modification of MiRNA Expression through Plant Extracts and Compounds against Breast Cancer: Mechanism and Translational Significance. Phytomedicine 2020, 68, 153168. [Google Scholar] [CrossRef] [PubMed]
- Mekkawy, A.I.; Eleraky, N.E.; Soliman, G.M.; Elnaggar, M.G.; Elnaggar, M.G. Combinatorial Therapy of Letrozole- and Quercetin-Loaded Spanlastics for Enhanced Cytotoxicity against MCF-7 Breast Cancer Cells. Pharmaceutics 2022, 14, 1727. [Google Scholar] [CrossRef]
- Mawalizadeh, F.; Mohammadzadeh, G.; Khedri, A.; Rashidi, M. Quercetin Potentiates the Chemosensitivity of MCF-7 Breast Cancer Cells to 5-Fluorouracil. Mol. Biol. Rep. 2021, 48, 7733–7742. [Google Scholar] [CrossRef] [PubMed]
- Safi, A.; Heidarian, E.; Ahmadi, R. Quercetin Synergistically Enhances the Anticancer Efficacy of Docetaxel through Induction of Apoptosis and Modulation of PI3K/AKT, MAPK/ERK, and JAK/STAT3 Signaling Pathways in MDA-MB-231 Breast Cancer Cell Line. Int. J. Mol. Cell. Med. 2021, 10, 11–22. [Google Scholar] [CrossRef]
- Prieto-Vila, M.; Shimomura, I.; Kogure, A.; Usuba, W.; Takahashi, R.; Ochiya, T.; Yamamoto, Y. Quercetin Inhibits Lef1 and Resensitizes Docetaxel-Resistant Breast Cancer Cells. Molecules 2020, 25, 2576. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhao, Q.; Wang, B.; Yuan, S.; Wang, X.; Li, K. Quercetin Reversed MDR in Breast Cancer Cells through Down-Regulating P-Gp Expression and Eliminating Cancer Stem Cells Mediated by YB-1 Nuclear Translocation: The Chemosensitization of Quercetin. Phytother. Res. 2018, 32, 1530–1536. [Google Scholar] [CrossRef]
- Song, Y.-K.; Yoon, J.-H.; Woo, J.K.; Kang, J.-H.; Lee, K.-R.; Oh, S.H.; Chung, S.-J.; Maeng, H.-J. Quercetin Is a Flavonoid Breast Cancer Resistance Protein Inhibitor with an Impact on the Oral Pharmacokinetics of Sulfasalazine in Rats. Pharmaceutics 2020, 12, 397. [Google Scholar] [CrossRef]
- Hosseinzadeh, R.; Khorsandi, K.; Esfahani, H.S.; Habibi, M.; Hosseinzadeh, G. Preparation of Cerium-Curcumin and Cerium-Quercetin Complexes and Their LEDs Irradiation Assisted Anticancer Effects on MDA-MB-231 and A375 Cancer Cell Lines. Photodiagnosis Photodyn. Ther. 2021, 34, 102326. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Nie, S.; Pan, X.; Zhang, R.; Fan, Z.; Wang, S. Quercetin-Nanostructured Lipid Carriers: Characteristics and Anti-Breast Cancer Activities in Vitro. Colloids Surf. B Biointerfaces 2014, 113, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Niazvand, F.; Orazizadeh, M.; Khorsandi, L.; Abbaspour, M.; Mansouri, E.; Khodadadi, A. Effects of Quercetin-Loaded Nanoparticles on MCF-7 Human Breast Cancer Cells. Medicina 2019, 55, 114. [Google Scholar] [CrossRef] [Green Version]
- Askar, M.A.; El-Nashar, H.A.; Al-Azzawi, M.A.; Rahman, S.S.A.; Elshawi, O.E. Synergistic Effect of Quercetin Magnetite Nanoparticles and Targeted Radiotherapy in Treatment of Breast Cancer. Breast Cancer 2022, 16, 117822342210867. [Google Scholar] [CrossRef]
- Dhanaraj, T.; Mohan, M.; Arunakaran, J. Quercetin Attenuates Metastatic Ability of Human Metastatic Ovarian Cancer Cells via Modulating Multiple Signaling Molecules Involved in Cell Survival, Proliferation, Migration and Adhesion. Arch. Biochem. Biophys. 2021, 701, 108795. [Google Scholar] [CrossRef]
- Ren, M.-X.; Deng, X.-H.; Ai, F.; Yuan, G.-Y.; Song, H.-Y. Effect of Quercetin on the Proliferation of the Human Ovarian Cancer Cell Line SKOV-3 in Vitro. Exp. Ther. Med. 2015, 10, 579–583. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Li, B.; Wang, T.; Wan, J.; Zhang, Y.; Huang, J.; Shen, Y. Enhancing the Anti-Ovarian Cancer Activity of Quercetin Using a Self-Assembling Micelle and Thermosensitive Hydrogel Drug Delivery System. RSC Adv. 2018, 8, 21229–21242. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Gong, J.; Ding, C.; Chen, G. Quercetin Induces the Apoptosis of Human Ovarian Carcinoma Cells by Upregulating the Expression of MicroRNA-145. Mol. Med. Rep. 2015, 12, 3127–3131. [Google Scholar] [CrossRef] [Green Version]
- Ramalingam, V.; Muthukumar Sathya, P.; Srivalli, T.; Mohan, H. Synthesis of Quercetin Functionalized Wurtzite Type Zinc Oxide Nanoparticles and Their Potential to Regulate Intrinsic Apoptosis Signaling Pathway in Human Metastatic Ovarian Cancer. Life. Sci. 2022, 309, 121022. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gong, W.; Yang, Z.Y.; Zhou, X.S.; Gong, C.; Zhang, T.R.; Wei, X.; Ma, D.; Ye, F.; Gao, Q.L. Quercetin Induces Protective Autophagy and Apoptosis through ER Stress via the P-STAT3/Bcl-2 Axis in Ovarian Cancer. Apoptosis 2017, 22, 544–557. [Google Scholar] [CrossRef] [PubMed]
- Hasan, A.A.S.; Kalinina, E.V.; Tatarskiy, V.V.; Volodina, Y.L.; Petrova, A.S.; Novichkova, M.D.; Zhdanov, D.D.; Shtil, A.A. Suppression of the Antioxidant System and PI3K/Akt/MTOR Signaling Pathway in Cisplatin-Resistant Cancer Cells by Quercetin. Bull. Exp. Biol. Med. 2022, 173, 760–764. [Google Scholar] [CrossRef]
- Gong, C.; Yang, Z.; Zhang, L.; Wang, Y.; Gong, W.; Liu, Y. Quercetin Suppresses DNA Double-Strand Break Repair and Enhances the Radiosensitivity of Human Ovarian Cancer Cells via P53-Dependent Endoplasmic Reticulum Stress Pathway. Onco Targets Ther. 2017, 11, 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, L.; Zongyuan, Y.; Cheng, G.; Lingyun, Z.; GuiLian, Y.; Wei, G. Quercetin Enhances Apoptotic Effect of Tumor Necrosis Factor-related Apoptosis-inducing Ligand (TRAIL) in Ovarian Cancer Cells through Reactive Oxygen Species (ROS) Mediated CCAAT Enhancer-binding Protein Homologous Protein (CHOP)-death Receptor 5 Pathway. Cancer Sci. 2014, 105, 520–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, S.G.; Gala, K.; Khattar, E. Telomerase Inhibitor MST-312 and Quercetin Synergistically Inhibit Cancer Cell Proliferation by Promoting DNA Damage. Transl. Oncol. 2023, 27, 101569. [Google Scholar] [CrossRef]
- Bandera, E.V.; Williams, M.G.; Sima, C.; Bayuga, S.; Pulick, K.; Wilcox, H.; Soslow, R.; Zauber, A.G.; Olson, S.H. Phytoestrogen Consumption and Endometrial Cancer Risk: A Population-Based Case–Control Study in New Jersey. Cancer Causes Control. 2009, 20, 1117–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estany, S.; Palacio, J.R.; Barnadas, R.; Sabes, M.; Iborra, A.; Martínez, P. Antioxidant Activity of N-Acetylcysteine, Flavonoids and α-Tocopherol on Endometrial Cells in Culture. J. Reprod. Immunol. 2007, 75, 1–10. [Google Scholar] [CrossRef]
- Li, X.; Zhu, Q.; Ma, M.; Guo, H. Quercetin Inhibits the Progression of Endometrial HEC-1-A Cells by Regulating Ferroptosis—A Preliminary Study. Eur. J. Med. Res. 2022, 27, 292. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.B.; Xiao, C.; Chen, X.L.; Bai, P.; Yao, Y.; Wang, H.; Xiao, X. The Antitumor Effects of Fisetin on Ovarian Cancer in Vitro and in Vivo. Sichuan Univ. Med. Sci. Ed. 2016, 47, 830–836. [Google Scholar]
- Liu, Y.; Cao, H.; Zhao, Y.; Shan, L.; Lan, S. Fisetin-Induced Cell Death in Human Ovarian Cancer Cell Lines via Zbp1-Mediated Necroptosis. J. Ovarian Res. 2022, 15, 57. [Google Scholar] [CrossRef]
- Jafarzadeh, S.; Baharara, J.; Tehranipour, M. Apoptosis Induction with Combined Use of Cisplatin and Fisetin in Cisplatin- Resistant Ovarian Cancer Cells (A2780). Avicenna J. Med. Biotechnol. 2021, 13, 176. [Google Scholar] [CrossRef]
- Huang, H.; Chen, A.Y.; Ye, X.; Guan, R.; Rankin, G.O.; Chen, Y.C. Galangin, a Flavonoid from Lesser Galangal, Induced Apoptosis via P53-Dependent Pathway in Ovarian Cancer Cells. Molecules 2020, 25, 1579. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Xu, Z.; Cai, Q.; Deng, Y.; Shi, W.; Zhou, H.; Wang, D.; Li, J. Isorhamnetin Inhibits Progression of Ovarian Cancer by Targeting ESR1. Ann. Transl. Med. 2022, 10, 1216. [Google Scholar] [CrossRef]
- Nowak, E.; Sypniewski, D.; Bednarek, I. Morin Exerts Anti-Metastatic, Anti-Proliferative and Anti-Adhesive Effect in Ovarian Cancer Cells: An in Vitro Studies. Mol. Biol. Rep. 2020, 47, 1965–1978. [Google Scholar] [CrossRef] [Green Version]
- Kashyap, D.; Sharma, A.; Sak, K.; Tuli, H.S.; Buttar, H.S.; Bishayee, A. Fisetin: A Bioactive Phytochemical with Potential for Cancer Prevention and Pharmacotherapy. Life Sci. 2018, 194, 75–87. [Google Scholar] [CrossRef]
- Yarla, N.S.; Bishayee, A.; Sethi, G.; Reddanna, P.; Kalle, A.M.; Dhananjaya, B.L.; Dowluru, K.S.V.G.K.; Chintala, R.; Duddukuri, G.R. Targeting Arachidonic Acid Pathway by Natural Products for Cancer Prevention and Therapy. Semin. Cancer Biol. 2016, 40–41, 48–81. [Google Scholar] [CrossRef]
- Rahmani, A.H.; Almatroudi, A.; Allemailem, K.S.; Khan, A.A.; Almatroodi, S.A. The Potential Role of Fisetin, a Flavonoid in Cancer Prevention and Treatment. Molecules 2022, 27, 9009. [Google Scholar] [CrossRef]
- Imran, M.; Saeed, F.; Gilani, S.A.; Shariati, M.A.; Imran, A.; Afzaal, M.; Atif, M.; Tufail, T.; Anjum, F.M. Fisetin: An Anticancer Perspective. Food Sci. Nutr. 2021, 9, 3–16. [Google Scholar] [CrossRef]
- Farooqi, A.A.; Naureen, H.; Zahid, R.; Youssef, L.; Attar, R.; Xu, B. Cancer Chemopreventive Role of Fisetin: Regulation of Cell Signaling Pathways in Different Cancers. Pharmacol. Res. 2021, 172, 105784. [Google Scholar] [CrossRef]
- Kubina, R.; Krzykawski, K.; Kabała-Dzik, A.; Wojtyczka, R.D.; Chodurek, E.; Dziedzic, A. Fisetin, a Potent Anticancer Flavonol Exhibiting Cytotoxic Activity against Neoplastic Malignant Cells and Cancerous Conditions: A Scoping, Comprehensive Review. Nutrients 2022, 14, 2604. [Google Scholar] [CrossRef]
- Khozooei, S.; Lettau, K.; Barletta, F.; Jost, T.; Rebholz, S.; Veerappan, S.; Franz-Wachtel, M.; Macek, B.; Iliakis, G.; Distel, L.V.; et al. Fisetin Induces DNA Double-Strand Break and Interferes with the Repair of Radiation-Induced Damage to Radiosensitize Triple Negative Breast Cancer Cells. J. Exp. Clin. Cancer Res. 2022, 41, 256. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, D.-Z.; Wang, Y.-X. Bioflavonoid Fisetin Loaded α-Tocopherol-Poly(Lactic Acid)-Based Polymeric Micelles for Enhanced Anticancer Efficacy in Breast Cancers. Pharm. Res. 2017, 34, 453–461. [Google Scholar] [CrossRef]
- Xiao, X.; Zou, J.; Fang, Y.; Meng, Y.; Xiao, C.; Fu, J.; Liu, S.; Bai, P.; Yao, Y. Fisetin and Polymeric Micelles Encapsulating Fisetin Exhibit Potent Cytotoxic Effects towards Ovarian Cancer Cells. BMC Complement. Altern. Med. 2018, 18, 91. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.S.; Kim, M.K.; Choi, Y.K.; Shin, Y.C.; Cho, S.-G.; Ko, S.-G. Rhus Verniciflua Stokes (RVS) and Butein Induce Apoptosis of Paclitaxel-Resistant SKOV-3/PAX Ovarian Cancer Cells through Inhibition of AKT Phosphorylation. BMC Complement. Altern. Med. 2016, 16, 122. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.; Feng, Z.; Chen, A.; Qi, Q.; Han, M.; Wang, S.; Zhang, Y.; Zhang, X.; Yang, N.; Wang, J.; et al. The Natural Flavonoid Galangin Elicits Apoptosis, Pyroptosis, and Autophagy in Glioblastoma. Front. Oncol. 2019, 9, 942. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Ma, Y.; Yin, J.; Li, Y.; Cao, Y.; Zhang, L. Analysis of Galangin and Its In Vitro/In Vivo Metabolites via Ultra-High-Performance Liquid Chromatography/Quadrupole Time-of-Flight Mass Spectrometry. Metabolites 2022, 12, 1032. [Google Scholar] [CrossRef]
- Rampogu, S.; Gajula, R.G.; Lee, K.W. A Comprehensive Review on Chemotherapeutic Potential of Galangin. Biomed. Pharmacother. 2021, 141, 111808. [Google Scholar] [CrossRef]
- Su, L.; Chen, X.; Wu, J.; Lin, B.; Zhang, H.; Lan, L.; Luo, H. Galangin Inhibits Proliferation of Hepatocellular Carcinoma Cells by Inducing Endoplasmic Reticulum Stress. Food Chem. Toxicol. 2013, 62, 810–816. [Google Scholar] [CrossRef]
- Zhu, Y.; Rao, Q.; Zhang, X.; Zhou, X. Galangin Induced Antitumor Effects in Human Kidney Tumor Cells Mediated via Mitochondrial Mediated Apoptosis, Inhi- Bition of Cell Migration and Invasion and Targeting PI3K/ AKT/MTOR Signalling Pathway. J. BUON 2018, 23, 795–799. [Google Scholar]
- Liang, X.; Wang, P.; Yang, C.; Huang, F.; Wu, H.; Shi, H.; Wu, X. Galangin Inhibits Gastric Cancer Growth Through Enhancing STAT3 Mediated ROS Production. Front. Pharmacol. 2021, 12, 646628. [Google Scholar] [CrossRef]
- van Zanden, J.J.; Geraets, L.; Wortelboer, H.M.; Bladeren, P.J.; van Rietjens, I.M.C.M.; Cnubben, N.H.P. Structural Requirements for the Flavonoid-Mediated Modulation of Glutathione S-Transferase P1-1 and GS-X Pump Activity in MCF7 Breast Cancer Cells. Biochem. Pharmacol. 2004, 67, 1607–1617. [Google Scholar] [CrossRef]
- Seyhan, M.F.; Yılmaz, E.; Timirci-Kahraman, Ö.; Saygılı, N.; Kısakesen, H.İ.; Gazioğlu, S.; Gören, A.C.; Eronat, A.P.; Begüm Ceviz, A.; Öztürk, T.; et al. Different Propolis Samples, Phenolic Content, and Breast Cancer Cell Lines: Variable Cytotoxicity Ranging from Ineffective to Potent. IUBMB Life 2019, 71, 619–631. [Google Scholar] [CrossRef]
- Aryappalli, P.; Shabbiri, K.; Masad, R.J.; Al-Marri, R.H.; Haneefa, S.M.; Mohamed, Y.A.; Arafat, K.; Attoub, S.; Cabral-Marques, O.; Ramadi, K.B.; et al. Inhibition of Tyrosine-Phosphorylated STAT3 in Human Breast and Lung Cancer Cells by Manuka Honey Is Mediated by Selective Antagonism of the IL-6 Receptor. Int. J. Mol. Sci. 2019, 20, 4340. [Google Scholar] [CrossRef] [Green Version]
- Abbas, Z.S.; Sulaiman, G.M.; Jabir, M.S.; Mohammed, S.A.A.; Khan, R.A.; Mohammed, H.A.; Al-Subaiyel, A. Galangin/β-Cyclodextrin Inclusion Complex as a Drug-Delivery System for Improved Solubility and Biocompatibility in Breast Cancer Treatment. Molecules 2022, 27, 4521. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.; Sohel, M.; Rahman, M.H.; Hasan, M.I.; Khan, S.; Amin, A.; Islam, Z.; Peng, S. Bioinformatics and In Silico Approaches to Identify Novel Biomarkers and Key Pathways for Cancers That Are Linked to the Progression of Female Infertility: A Comprehensive Approach for Drug Discovery. PLoS ONE 2023, 18, e0265746. [Google Scholar] [CrossRef] [PubMed]
- Gong, G.; Guan, Y.-Y.; Zhang, Z.-L.; Rahman, K.; Wang, S.-J.; Zhou, S.; Luan, X.; Zhang, H. Isorhamnetin: A Review of Pharmacological Effects. Biomed. Pharmacother. 2020, 128, 110301. [Google Scholar] [CrossRef] [PubMed]
- González-Arceo, M.; Gomez-Lopez, I.; Carr-Ugarte, H.; Eseberri, I.; González, M.; Cano, M.P.; Portillo, M.P.; Gómez-Zorita, S. Anti-Obesity Effects of Isorhamnetin and Isorhamnetin Conjugates. Int. J. Mol. Sci. 2022, 24, 299. [Google Scholar] [CrossRef]
- Antunes-Ricardo, M.; Rodríguez-Rodríguez, C.; Gutiérrez-Uribe, J.; Cepeda-Cañedo, E.; Serna-Saldívar, S. Bioaccessibility, Intestinal Permeability and Plasma Stability of Isorhamnetin Glycosides from Opuntia ficus-indica (L.). Int. J. Mol. Sci. 2017, 18, 1816. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Zhang, Y.; Jiang, X.; Zhang, H.; Gao, Z.; Li, Y.; Fu, R.; Li, L.; Li, J.; Cui, H.; et al. ROS-Mediated Activation and Mitochondrial Translocation of CaMKII Contributes to Drp1-Dependent Mitochondrial Fission and Apoptosis in Triple-Negative Breast Cancer Cells by Isorhamnetin and Chloroquine. J. Exp. Clin. Cancer Res. 2019, 38, 225. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wang, X.; Hong, W.; Zhang, S.; Yang, Y.; Xia, Y.; Yang, R. The Pharmacological Mechanism of Chinese Herbs Effective in Treating Advanced Ovarian Cancer: Integrated Meta-Analysis and Network Pharmacology Analysis. Front. Pharmacol. 2022, 13, 1040641. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Ma, R.-H.; Zhang, X.-X.; Thakur, K.; Zhang, J.-G.; Khan, M.R.; Busquets, R.; Wei, Z.-J. Isorhamnetin Induces Apoptosis and Suppresses Metastasis of Human Endometrial Carcinoma Ishikawa Cells via Endoplasmic Reticulum Stress Promotion and Matrix Metalloproteinase-2/9 Inhibition In Vitro and In Vivo. Foods 2022, 11, 3415. [Google Scholar] [CrossRef] [PubMed]
- Rajput, S.A.; Wang, X.; Yan, H.-C. Morin Hydrate: A Comprehensive Review on Novel Natural Dietary Bioactive Compound with Versatile Biological and Pharmacological Potential. Biomed. Pharmacother. 2021, 138, 111511. [Google Scholar] [CrossRef]
- Bieg, D.; Sypniewski, D.; Nowak, E.; Bednarek, I. Morin Decreases Galectin-3 Expression and Sensitizes Ovarian Cancer Cells to Cisplatin. Arch. Gynecol. Obstet. 2018, 298, 1181–1194. [Google Scholar] [CrossRef] [Green Version]




| Flavonoid | Structure | Bioactivities | Sources | Ref. |
|---|---|---|---|---|
| Chalcones |  | Antioxidant, antimalarial, anti-inflammatory, antimicrobial, antiosteoporosis, antiplasmodial, anticancer, antifungal, and antihyperglycemic | Plants: Buteamonospermia, Humuluslupulus, Helichrysumrugulosum, Neoraputiamagnifica, Angelicakeiskei, Piperhispidum, Tarennaattenuata, and Calythropisaurea | [21] |
| Flavanones |  | Antioxidant, anti-inflammatory, and anti-ischemic | Citrus (grapefruit, orange, and lemon) and tomatoes | [22] |
| Flavones |  | Antitumor, antiviral, antimicrobial, anti-inflammatory, antioxidant, neuroprotective, anti-IR-insulin resistance activity, and hepatoprotective | Plants: Godmaniaaesculifolia, Tridaxprocumbens, Primulafarinosa L., and Chrysanthemummorifolium | [23] |
| Leucoanthocyanidin Flavan-3,4-ol |  | Antioxidation, anti-inflammatory, anticancer, antiviral, and protective cardiovascular properties | Plants: Acaciapeuce, A. carneorum, and A. crombiei | [24] |
| Flavonols |  | Antioxidant, anticancer, anti-inflammatory, hepatoprotective, neuroprotective, cardioprotective, antimicrobial, and iron-chelating | Berries, citrus fruits, spices, black or green tea, capers, arugula, cabbage, kale, cress, watercress, sea buckthorn, parsley, and carob | [25] |
| Proanthocyanidin flavan-3-ol |  | Antifungal, antiviral, anti-inflammatory, anticancer, antiangiogenic, and protective against neurological and heart diseases | Tea, grapes, and wine | [26,27] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wendlocha, D.; Krzykawski, K.; Mielczarek-Palacz, A.; Kubina, R. Selected Flavonols in Breast and Gynecological Cancer: A Systematic Review. Nutrients 2023, 15, 2938. https://doi.org/10.3390/nu15132938
Wendlocha D, Krzykawski K, Mielczarek-Palacz A, Kubina R. Selected Flavonols in Breast and Gynecological Cancer: A Systematic Review. Nutrients. 2023; 15(13):2938. https://doi.org/10.3390/nu15132938
Chicago/Turabian StyleWendlocha, Dominika, Kamil Krzykawski, Aleksandra Mielczarek-Palacz, and Robert Kubina. 2023. "Selected Flavonols in Breast and Gynecological Cancer: A Systematic Review" Nutrients 15, no. 13: 2938. https://doi.org/10.3390/nu15132938
APA StyleWendlocha, D., Krzykawski, K., Mielczarek-Palacz, A., & Kubina, R. (2023). Selected Flavonols in Breast and Gynecological Cancer: A Systematic Review. Nutrients, 15(13), 2938. https://doi.org/10.3390/nu15132938