
Dietary Acid Load Correlates with Serum Amino Acid Concentrations after a Four-Week Intervention with Vegan vs. Meat-Rich Diets: A Secondary Data Analysis

 ,
,  ,
, 
Abstract
:1. Introduction
- (I)
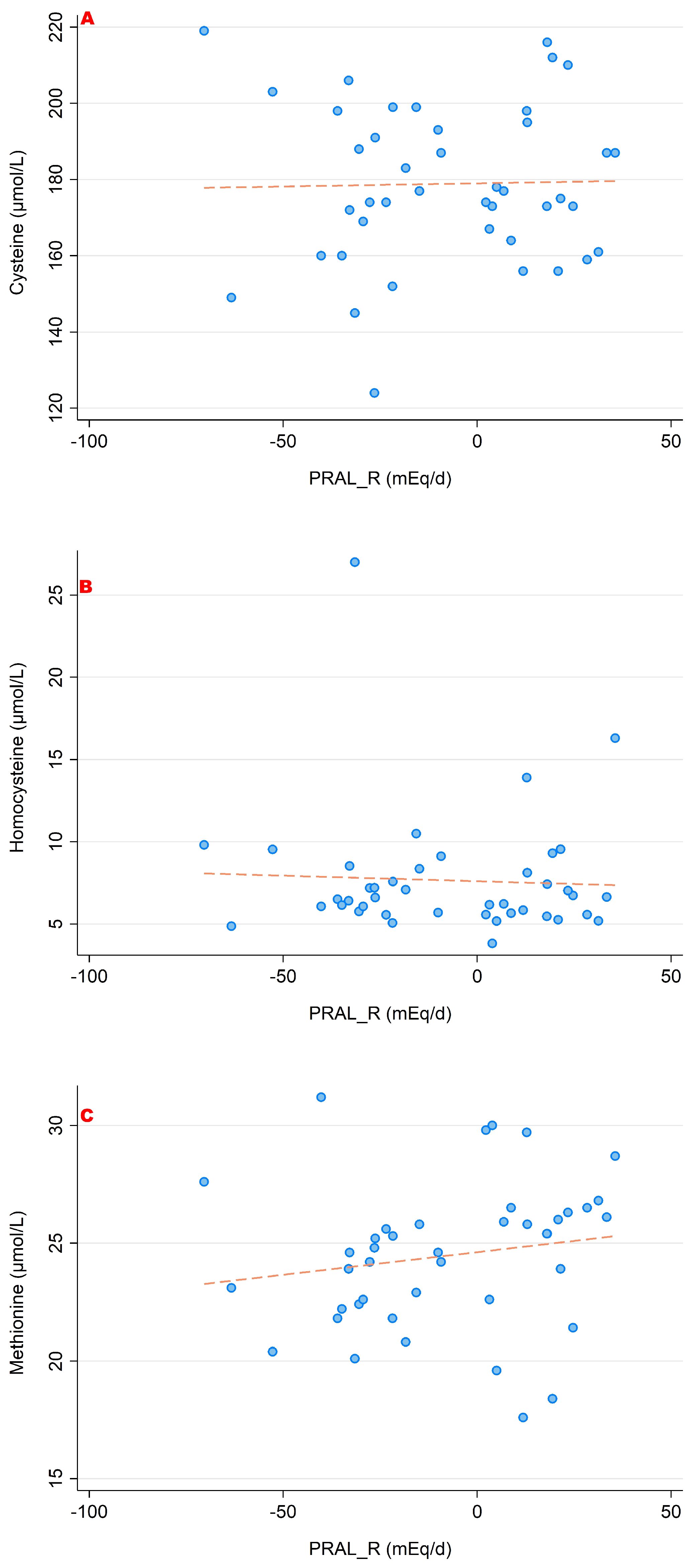
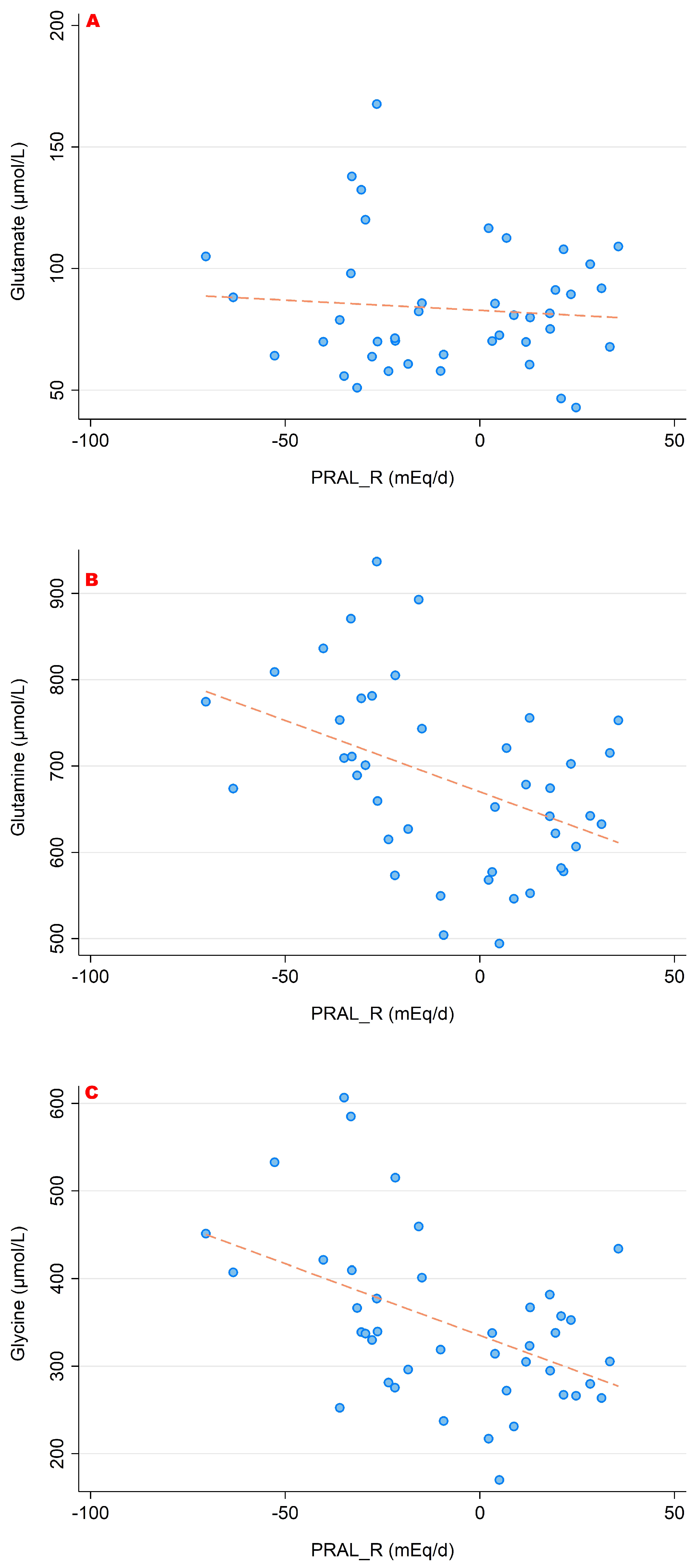
- lower concentrations of glutamine, glutamate and glycine (which are catabolized to buffer the acid load);
- (II)
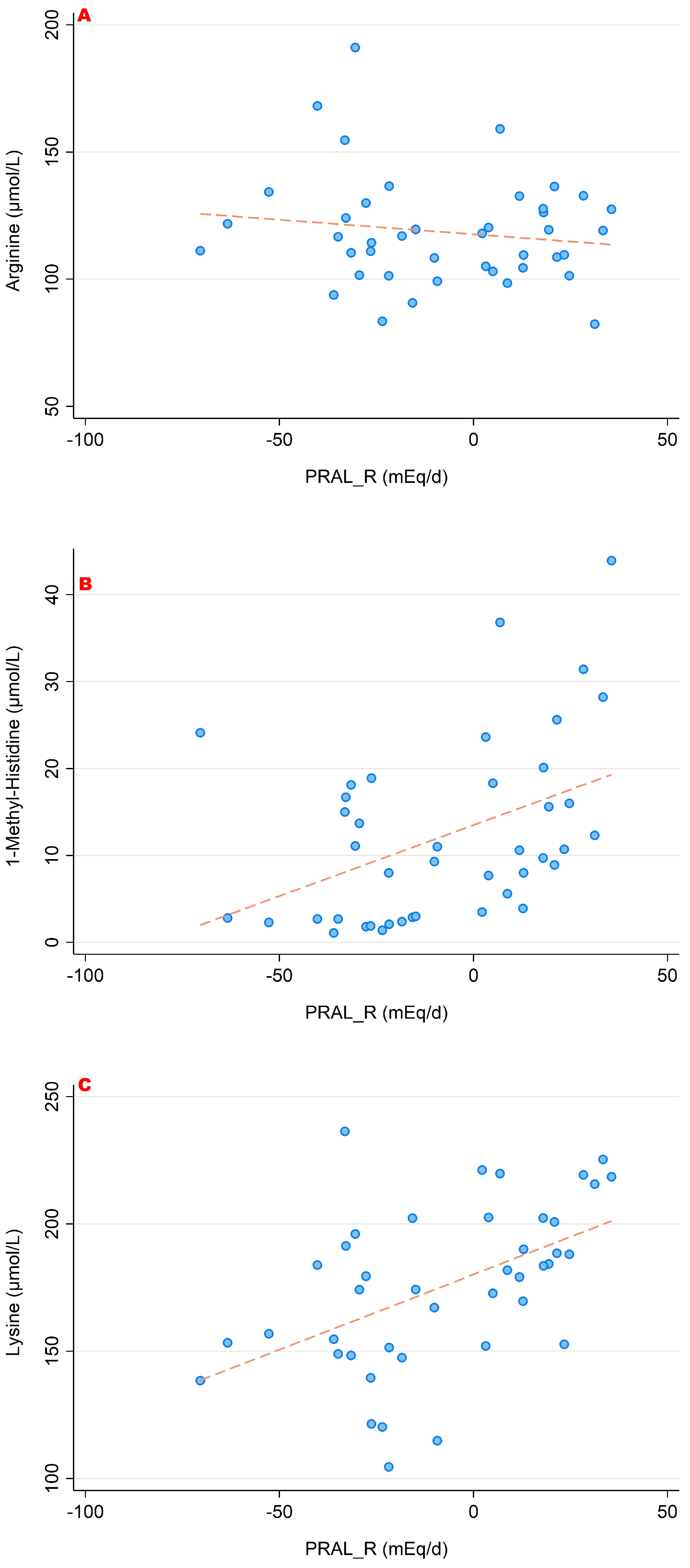
- higher concentrations of lysine, arginine and histidine (which are more abundant in acidic diets).
2. Materials and Methods
2.1. Study Population and Design
2.2. Dietary Acid Load Estimations
2.3. Dietary Intake
2.4. Amino Acid Profiles
2.5. Statistical Analysis and Analytical Considerations
2.6. Ethical Approval
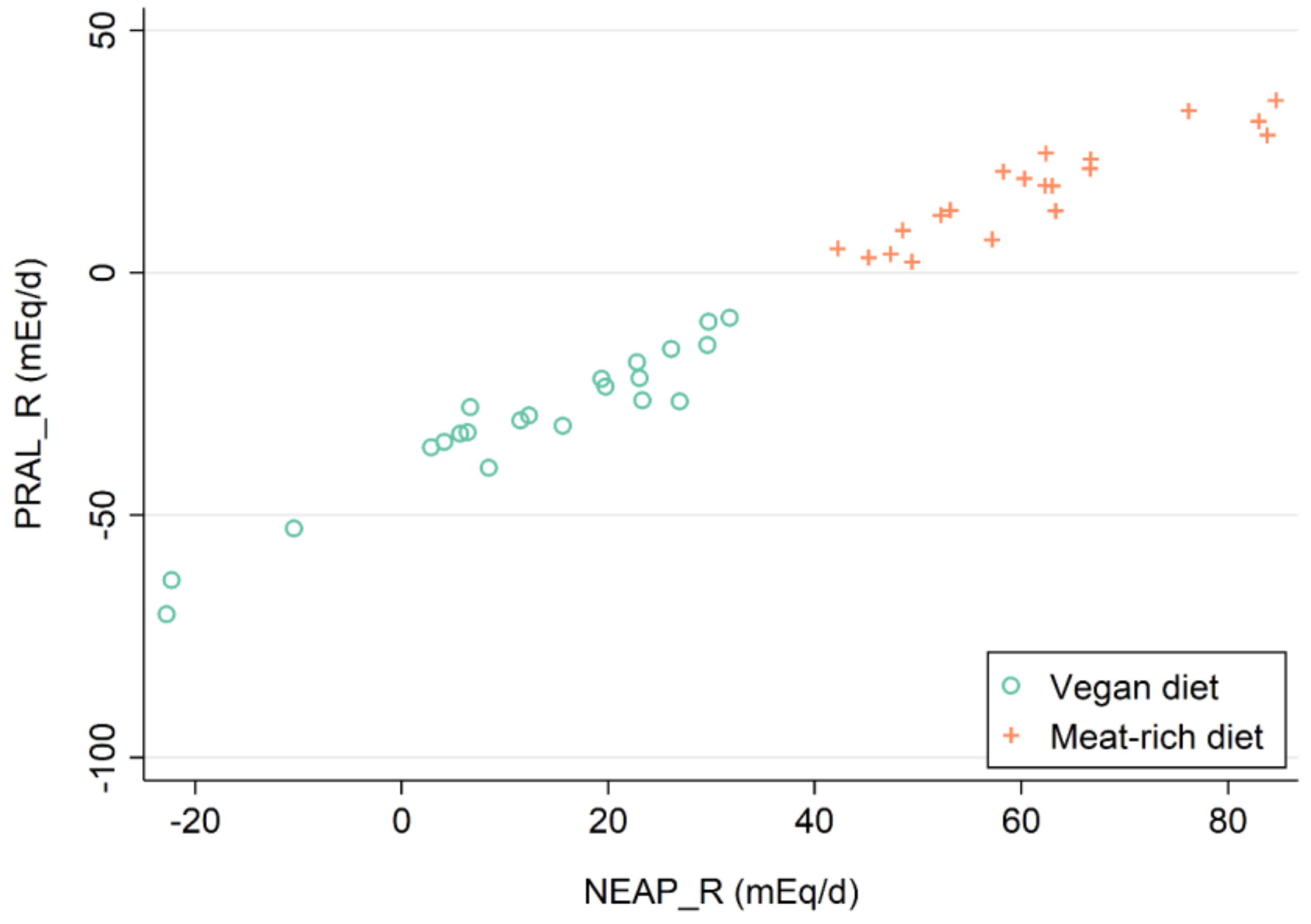
3. Results
3.1. PRALR and Its Association with Amino Acid Serum Concentrations
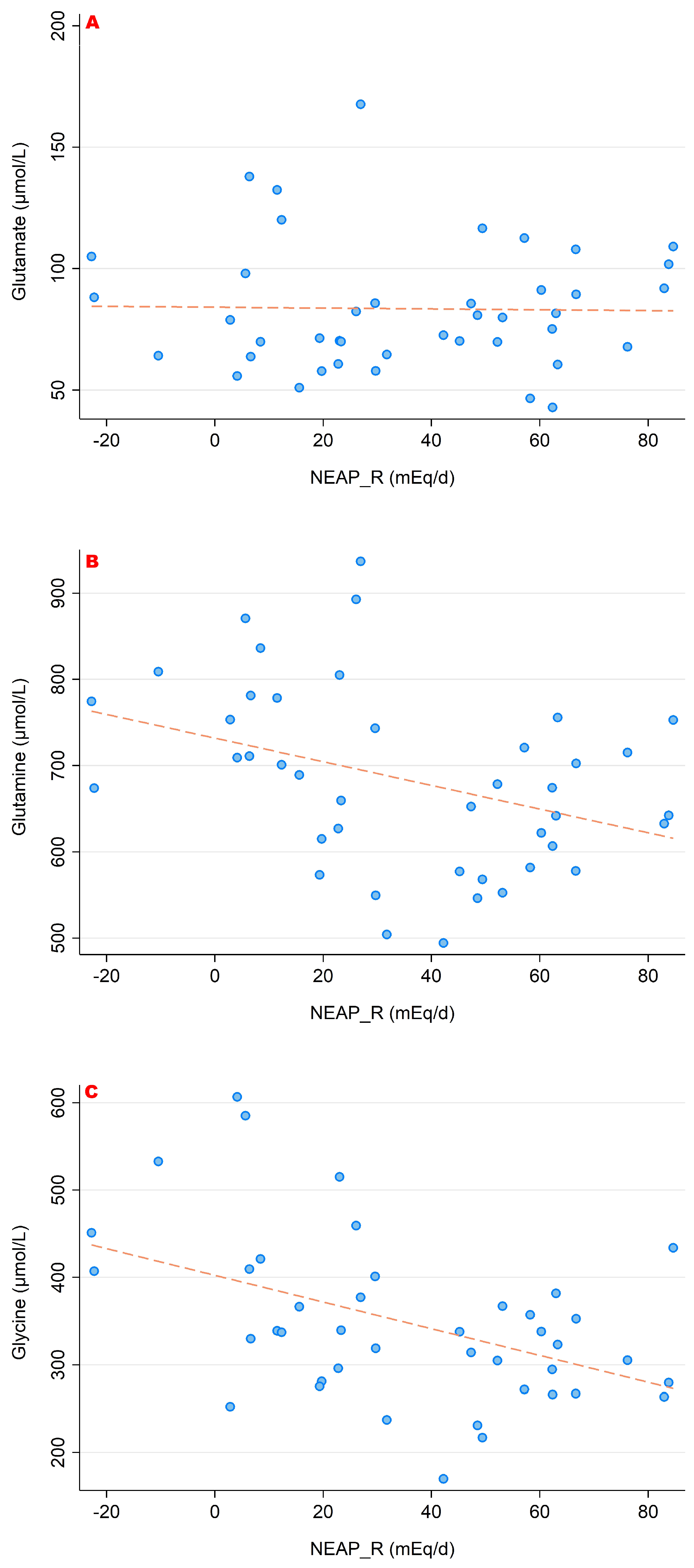
3.2. NEAPR and Its Association with Amino Acid Serum Concentrations
3.3. NEAPF and Its Association with Amino Acid Serum Concentrations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kahleova, H.; McCann, J.; Alwarith, J.; Rembert, E.; Tura, A.; Holubkov, R.; Barnard, N.D. A Plant-Based Diet in Overweight Adults in a 16-Week Randomized Clinical Trial: The Role of Dietary Acid Load. Clin. Nutr. ESPEN 2021, 44, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Wesson, D.E. The Importance of Recognizing and Addressing the Spectrum of Acid Stress. Adv. Chronic Kidney Dis. 2022, 29, 364–372. [Google Scholar] [CrossRef] [PubMed]
- DiNicolantonio, J.J.; O’Keefe, J. Low-Grade Metabolic Acidosis as a Driver of Chronic Disease: A 21st Century Public Health Crisis. Open Heart 2021, 8, e001730. [Google Scholar] [CrossRef]
- Robey, I.F. Examining the Relationship between Diet-Induced Acidosis and Cancer. Nutr. Metab. 2012, 9, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passey, C. Reducing the Dietary Acid Load: How a More Alkaline Diet Benefits Patients with Chronic Kidney Disease. J. Ren. Nutr. 2017, 27, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Tariq, A.; Chen, J.; Yu, B.; Boerwinkle, E.; Coresh, J.; Grams, M.E.; Rebholz, C.M. Metabolomics of Dietary Acid Load and Incident Chronic Kidney Disease. J. Ren. Nutr. 2022, 32, 292–300. [Google Scholar] [CrossRef]
- Storz, M.A.; Ronco, A.L.; Hannibal, L. Observational and Clinical Evidence That Plant-Based Nutrition Reduces Dietary Acid Load. J. Nutr. Sci. 2022, 11, e93. [Google Scholar] [CrossRef] [PubMed]
- Demigné, C.; Sabboh, H.; Puel, C.; Rémésy, C.; Coxam, V. Organic Anions and Potassium Salts in Nutrition and Metabolism. Nutr. Res. Rev. 2004, 17, 249–258. [Google Scholar] [CrossRef]
- Adeva, M.M.; Souto, G. Diet-Induced Metabolic Acidosis. Clin. Nutr. 2011, 30, 416–421. [Google Scholar] [CrossRef]
- Xiao, D.; Zeng, L.; Yao, K.; Kong, X.; Wu, G.; Yin, Y. The Glutamine-Alpha-Ketoglutarate (AKG) Metabolism and Its Nutritional Implications. Amino Acids 2016, 48, 2067–2080. [Google Scholar] [CrossRef]
- Chen, T.; Ni, Y.; Ma, X.; Bao, Y.; Liu, J.; Huang, F.; Hu, C.; Xie, G.; Zhao, A.; Jia, W.; et al. Branched-Chain and Aromatic Amino Acid Profiles and Diabetes Risk in Chinese Populations. Sci. Rep. 2016, 6, 20594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrannini, E.; Natali, A.; Camastra, S.; Nannipieri, M.; Mari, A.; Adam, K.-P.; Milburn, M.V.; Kastenmüller, G.; Adamski, J.; Tuomi, T.; et al. Early Metabolic Markers of the Development of Dysglycemia and Type 2 Diabetes and Their Physiological Significance. Diabetes 2013, 62, 1730–1737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Akter, S.; Kuwahara, K.; Matsushita, Y.; Nakagawa, T.; Konishi, M.; Honda, T.; Yamamoto, S.; Hayashi, T.; Noda, M.; et al. Serum Amino Acid Profiles and Risk of Type 2 Diabetes among Japanese Adults in the Hitachi Health Study. Sci. Rep. 2019, 9, 7010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lederer, A.-K.; Hannibal, L.; Hettich, M.; Behringer, S.; Spiekerkoetter, U.; Steinborn, C.; Gründemann, C.; Zimmermann-Klemd, A.M.; Müller, A.; Simmet, T.; et al. Vitamin B12 Status Upon Short-Term Intervention with a Vegan Diet-A Randomized Controlled Trial in Healthy Participants. Nutrients 2019, 11, 2815. [Google Scholar] [CrossRef] [Green Version]
- Lederer, A.-K.; Maul-Pavicic, A.; Hannibal, L.; Hettich, M.; Steinborn, C.; Gründemann, C.; Zimmermann-Klemd, A.M.; Müller, A.; Sehnert, B.; Salzer, U.; et al. Vegan Diet Reduces Neutrophils, Monocytes and Platelets Related to Branched-Chain Amino Acids—A Randomized, Controlled Trial. Clin. Nutr. 2020, 39, 3241–3250. [Google Scholar] [CrossRef]
- Müller, A.; Zimmermann-Klemd, A.M.; Lederer, A.-K.; Hannibal, L.; Kowarschik, S.; Huber, R.; Storz, M.A. A Vegan Diet Is Associated with a Significant Reduction in Dietary Acid Load: Post Hoc Analysis of a Randomized Controlled Trial in Healthy Individuals. Int. J. Environ. Res. Public Health 2021, 18, 9998. [Google Scholar] [CrossRef]
- Remer, T.; Manz, F. Estimation of the Renal Net Acid Excretion by Adults Consuming Diets Containing Variable Amounts of Protein. Am. J. Clin. Nutr. 1994, 59, 1356–1361. [Google Scholar] [CrossRef]
- Remer, T.; Manz, F. Potential Renal Acid Load of Foods and Its Influence on Urine PH. J. Am. Diet. Assoc. 1995, 95, 791–797. [Google Scholar] [CrossRef]
- Parmenter, B.H.; Dymock, M.; Banerjee, T.; Sebastian, A.; Slater, G.J.; Frassetto, L.A. Performance of Predictive Equations and Biochemical Measures Quantifying Net Endogenous Acid Production and the Potential Renal Acid Load. Kidney Int. Rep. 2020, 5, 1738–1745. [Google Scholar] [CrossRef]
- Frassetto, L.A.; Todd, K.M.; Morris, R.C.; Sebastian, A. Estimation of Net Endogenous Noncarbonic Acid Production in Humans from Diet Potassium and Protein Contents. Am. J. Clin. Nutr. 1998, 68, 576–583. [Google Scholar] [CrossRef] [Green Version]
- Kluthe, B. Freiburger Ernaehrungsprotokoll. Available online: www.ernaehrung.de/static/pdf/freiburger-ernaehrungsprotokoll.pdf (accessed on 20 April 2023).
- Willett, W. Nutritional Epidemiology, 3rd ed.; Oxford University Press: New York, NY, USA, 2013. [Google Scholar]
- Cosgrove, K.; Johnston, C.S. Examining the Impact of Adherence to a Vegan Diet on Acid-Base Balance in Healthy Adults. Plant Foods Hum. Nutr. 2017, 72, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Carnauba, R.A.; Baptistella, A.B.; Paschoal, V.; Hübscher, G.H. Diet-Induced Low-Grade Metabolic Acidosis and Clinical Outcomes: A Review. Nutrients 2017, 9, 538. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, J.A.; Rinaldi, S.; Scalbert, A.; Ferrari, P.; Achaintre, D.; Gunter, M.J.; Appleby, P.N.; Key, T.J.; Travis, R.C. Plasma Concentrations and Intakes of Amino Acids in Male Meat-Eaters, Fish-Eaters, Vegetarians and Vegans: A Cross-Sectional Analysis in the EPIC-Oxford Cohort. Eur. J. Clin. Nutr. 2016, 70, 306–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietrich, S.; Trefflich, I.; Ueland, P.M.; Menzel, J.; Penczynski, K.J.; Abraham, K.; Weikert, C. Amino Acid Intake and Plasma Concentrations and Their Interplay with Gut Microbiota in Vegans and Omnivores in Germany. Eur. J. Nutr. 2022, 61, 2103–2114. [Google Scholar] [CrossRef] [PubMed]
- Hayhoe, R.P.G.; Abdelhamid, A.; Luben, R.N.; Khaw, K.-T.; Welch, A.A. Dietary Acid-Base Load and Its Association with Risk of Osteoporotic Fractures and Low Estimated Skeletal Muscle Mass. Eur. J. Clin. Nutr. 2020, 74, 33–42. [Google Scholar] [CrossRef]
- Kataya, Y.; Murakami, K.; Kobayashi, S.; Suga, H.; Sasaki, S. Three-generation Study of Women on Diets and Health Study Group Higher Dietary Acid Load Is Associated with a Higher Prevalence of Frailty, Particularly Slowness/Weakness and Low Physical Activity, in Elderly Japanese Women. Eur. J. Nutr. 2018, 57, 1639–1650. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, A.L.; Friedrich, N.; Karlsson, M.K.; Ljunggren, Ö.; Lorentzon, M.; Nethander, M.; Wallaschofski, H.; Mellström, D.; Ohlsson, C. Serum Glycine Levels Are Associated with Cortical Bone Properties and Fracture Risk in Men. J. Clin. Endocrinol. Metab. 2021, 106, e5021–e5029. [Google Scholar] [CrossRef]
- Cruzat, V.; Macedo Rogero, M.; Noel Keane, K.; Curi, R.; Newsholme, P. Glutamine: Metabolism and Immune Function, Supplementation and Clinical Translation. Nutrients 2018, 10, 1564. [Google Scholar] [CrossRef] [Green Version]
- Durante, W. Glutamine Deficiency Promotes Immune and Endothelial Cell Dysfunction in COVID-19. Int. J. Mol. Sci. 2023, 24, 7593. [Google Scholar] [CrossRef]
- Deriemaeker, P.; Aerenhouts, D.; Hebbelinck, M.; Clarys, P. Nutrient Based Estimation of Acid-Base Balance in Vegetarians and Non-Vegetarians. Plant Foods Hum. Nutr. 2010, 65, 77–82. [Google Scholar] [CrossRef]
- Knurick, J.R.; Johnston, C.S.; Wherry, S.J.; Aguayo, I. Comparison of Correlates of Bone Mineral Density in Individuals Adhering to Lacto-Ovo, Vegan, or Omnivore Diets: A Cross-Sectional Investigation. Nutrients 2015, 7, 3416–3426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storz, M.A.; Ronco, A.L. Reduced Dietary Acid Load in U.S. Vegetarian Adults: Results from the National Health and Nutrition Examination Survey. Food Sci. Nutr. 2022, 10, 2091–2100. [Google Scholar] [CrossRef] [PubMed]
- Henriksson, J. Effect of Exercise on Amino Acid Concentrations in Skeletal Muscle and Plasma. J. Exp. Biol. 1991, 160, 149–165. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Result |
|---|---|
| Age (years) | 28.5 (12) |
| Gender | |
| Males | n = 15 (35.71%) |
| Females | n = 27 (64.29%) |
| Height (cm) | 173.24 ± 9.78 |
| Weight (kg) | 66.15 (15.2) |
| Body Mass Index (kg/m2) | 22.77 (3.28) |
| Nutrient and Energy Intake | |
|---|---|
| Total energy intake (kcal/d) | 2318 (850) |
| Magnesium intake (mg/d) | 382.79 (246.04) |
| Potassium intake (mg/d) | 3419.17 (1405.29) |
| Calcium intake (mg/d) | 691.38 (469.08) |
| Phosphorus intake (mg/d) | 1347.75 (618.05) |
| Protein intake (g/d) | 86.93 (41.98) |
| DAL scores | |
| PRALR (mEq/d) | −7.82 ± 27.41 |
| NEAPR (mEq/d) | 35.61 ± 28.51 |
| NEAPF (mEq/d) | 37.67 (30.29) |
| Amino Acid | |
|---|---|
| Arginine (μmol/L) | 116.85 (23.2) |
| Cysteine (μmol/L) | 178.88 ± 20.82 |
| Glutamate (μmol/L) | 79.4 (33.4) |
| Glutamine (μmol/L) | 683.07 ± 105.01 |
| Glycine (μmol/L) | 337.45 (121.7) |
| Histidine (μmol/L) | 10.15 (15.2) |
| Homocysteine (μmol/L) | 6.56 (2.7) |
| Lysine (μmol/L) | 175.55 ± 32.16 |
| Methionine (μmol/L) | 24.46 ± 3.10 |
| Amino Acid | EPIC–Oxford Cohort | RBVD Cohort | ||
|---|---|---|---|---|
| Vegans | Omnivores | Vegans | Omnivores | |
| Lysine | 2.82 (2.69, 2.95) a | 5.01 (4.78, 5.24) a | 41.0 (32.2–67.1) c,b | 78.9 (63.0–97.5) c,b |
| Glutamate/Glutamine | 14.06 (13.61, 14.52) a | 16.10 (15.59, 16.63) a | 206.7 (161.4–269.5) c,b | 250.1 (225.3–286.4) c,b |
| Glycine | 2.61 (2.50, 2.71) a | 3.12 (3.00, 3.25) a | 41.1 (32.2–57.0) c,b | 45.6 (40.3–61.3) c,b |
| Histidine | 1.52 (1.46, 1.57) a | 2.12 (2.04, 2.20) a | 20.0 (17.4–31.1) c,b | 32.0 (27.5–39.0) c,b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herter, J.; Lederer, A.-K.; Ronco, A.L.; Hannibal, L.; Huber, R.; Storz, M.A. Dietary Acid Load Correlates with Serum Amino Acid Concentrations after a Four-Week Intervention with Vegan vs. Meat-Rich Diets: A Secondary Data Analysis. Nutrients 2023, 15, 2942. https://doi.org/10.3390/nu15132942
Herter J, Lederer A-K, Ronco AL, Hannibal L, Huber R, Storz MA. Dietary Acid Load Correlates with Serum Amino Acid Concentrations after a Four-Week Intervention with Vegan vs. Meat-Rich Diets: A Secondary Data Analysis. Nutrients. 2023; 15(13):2942. https://doi.org/10.3390/nu15132942
Chicago/Turabian StyleHerter, Julian, Ann-Kathrin Lederer, Alvaro Luis Ronco, Luciana Hannibal, Roman Huber, and Maximilian Andreas Storz. 2023. "Dietary Acid Load Correlates with Serum Amino Acid Concentrations after a Four-Week Intervention with Vegan vs. Meat-Rich Diets: A Secondary Data Analysis" Nutrients 15, no. 13: 2942. https://doi.org/10.3390/nu15132942
APA StyleHerter, J., Lederer, A.-K., Ronco, A. L., Hannibal, L., Huber, R., & Storz, M. A. (2023). Dietary Acid Load Correlates with Serum Amino Acid Concentrations after a Four-Week Intervention with Vegan vs. Meat-Rich Diets: A Secondary Data Analysis. Nutrients, 15(13), 2942. https://doi.org/10.3390/nu15132942