
Controlling Chronic Diseases and Acute Infections with Vitamin D Sufficiency


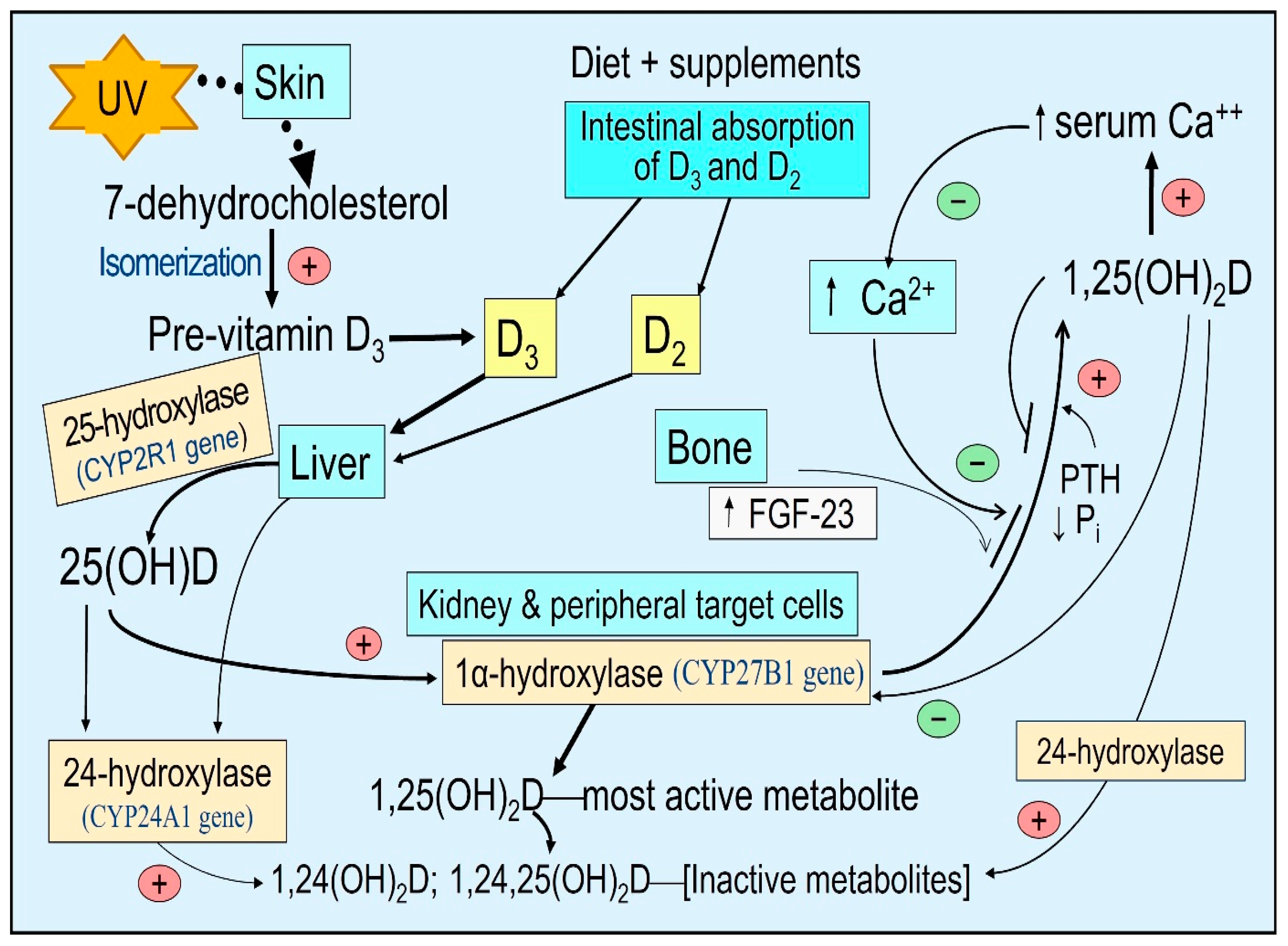{kind=link}
{kind=link}
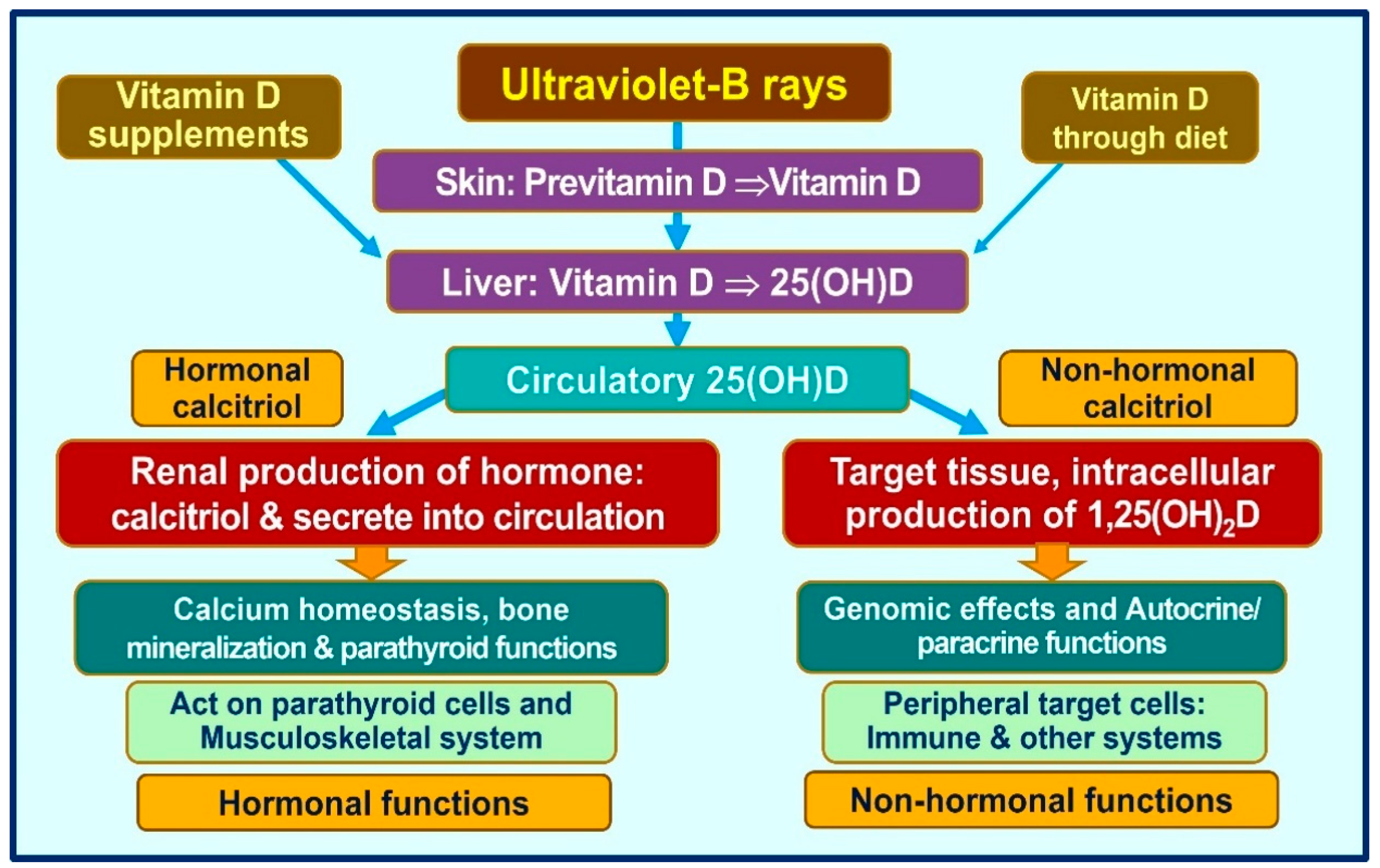{kind=link}
{kind=link}
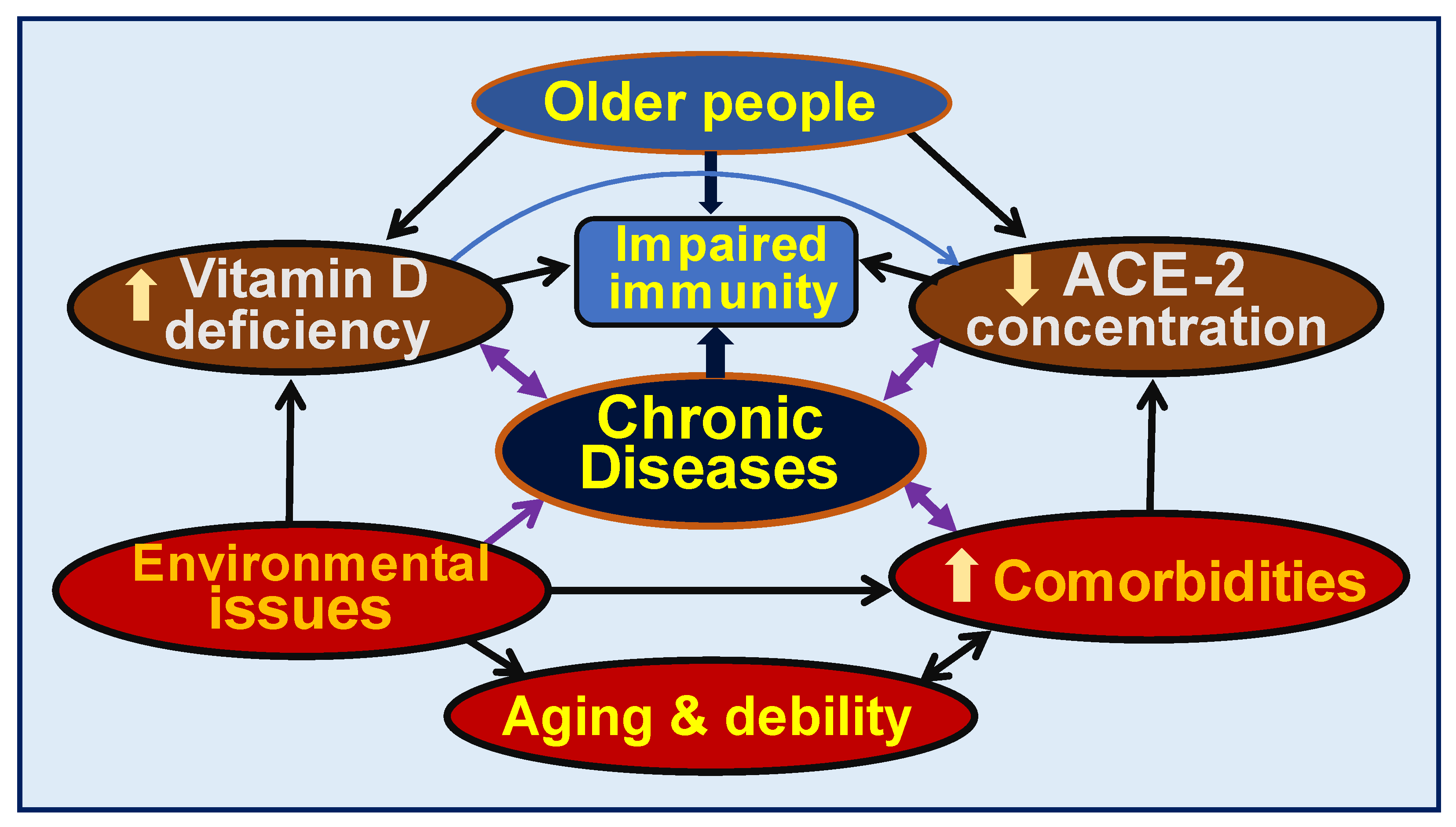{kind=link}
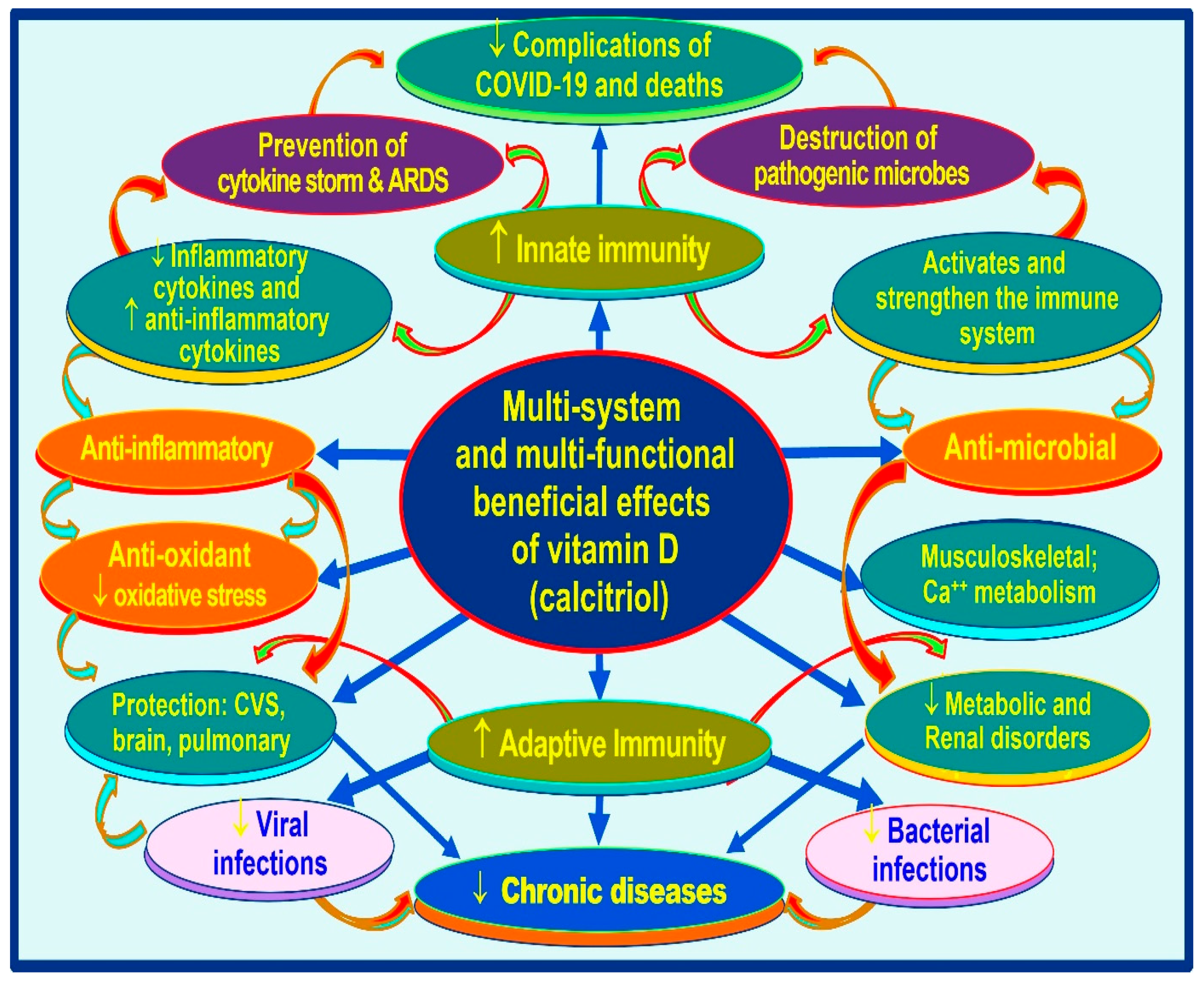{kind=link}
Abstract
:1. Introduction
2. Vitamin D—Brief History
3. Generation/Obtaining Vitamin D and Transportation in Humans
4. Consequences of Vitamin D Deficiency
5. Muscular–Skeletal Benefits of Vitamin D
6. Hypovitaminosis D and Viral Respiratory Infections
7. Extra-Skeletal Benefits of Vitamin D
8. Importance of Circulatory Vitamin D and 25(OH)D for Target Cell Generation of Calcitriol
9. Clinical Study Outcomes Using Higher Doses of Vitamin D
10. Entry of D and 25(OH)D into Peripheral Target Cells
11. Vitamin D, Epithelial Barriers, and Gap Junction Stability
12. What Has Changed over the Years Related to Vitamin D?
13. Vitamin D Intake Should Depend on Body Weight and Target Serum 25(OH)D Concentration
14. Vitamin D Is Essential for Activating Immune Cells
15. Discussions
16. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zdrenghea, M.T.; Makrinioti, H.; Bagacean, C.; Bush, A.; Johnston, S.L.; Stanciu, L.A. Vitamin D modulation of innate immune responses to respiratory viral infections. Rev. Med. Virol. 2017, 27, e1909. [Google Scholar] [CrossRef]
- Gotelli, E.; Soldano, S.; Hysa, E.; Paolino, S.; Campitiello, R.; Pizzorni, C.; Sulli, A.; Smith, V.; Cutolo, M. Vitamin D and COVID-19: Narrative Review after 3 Years of Pandemic. Nutrients 2022, 14, 4907. [Google Scholar] [CrossRef] [PubMed]
- Hollis, B.W.; Marshall, D.T.; Savage, S.J.; Garrett-Mayer, E.; Kindy, M.S.S.; Gattoni-Celli, S. Vitamin D3 supplementation, low-risk prostate cancer, and health disparities. J. Steroid Biochem. Mol. Biol. 2013, 136, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Xiaoyu, Z.; Payal, B.; Melissa, O.; Zanello, L.P. 1alpha,25(OH)2-vitamin D3 membrane-initiated calcium signaling modulates exocytosis and cell survival. J. Steroid Biochem. Mol. Biol. 2007, 103, 457–461. [Google Scholar] [CrossRef]
- Liang, F.; Liu, C.; Li, L.; Guo, Y.; Bai, L. Effects of gastrin on rat intestinal epithelial 1,25(OH)2D3-membrane associated rapid response steroid binding protein. Nan Fang Yi Ke Da Xue Xue Bao 2013, 33, 990–993. [Google Scholar] [PubMed]
- Vieth, R. Why the optimal requirement for Vitamin D3 is probably much higher than what is officially recommended for adults. J. Steroid Biochem. Mol. Biol. 2004, 89–90, 575–579. [Google Scholar] [CrossRef]
- Wimalawansa, S.J. Vitamin D in the new millennium. Curr. Osteoporos. Rep. 2012, 10, 4–15. [Google Scholar] [CrossRef]
- Bouillon, R. Genetic and racial differences in the vitamin D endocrine system. Endocrinol. Metab. Clin. N. Am. 2017, 46, 1119–1135. [Google Scholar] [CrossRef]
- Holick, M.F.; Chen, T.C.; Lu, Z.; Sauter, E. Vitamin D and skin physiology: A D-lightful story. J. Bone Miner. Res. 2007, 22 (Suppl. S2), V28–V33. [Google Scholar] [CrossRef]
- Engelsen, O. The relationship between ultraviolet radiation exposure and vitamin D status. Nutrients 2010, 2, 482–495. [Google Scholar] [CrossRef]
- Webb, A.R.; Alghamdi, R.; Kift, R.; Rhodes, L.E. 100 Years of Vitamin D: Dose-response for change in 25-hydroxyvitamin D after UV exposure: Outcome of a systematic review. Endocr. Connect. 2021, 10, R248–R266. [Google Scholar] [CrossRef] [PubMed]
- Wimalawansa, S.J. Public health interventions for chronic diseases: Cost-benefit modelizations for eradicating chronic kidney disease of multifactorial origin (CKDmfo/CKDu) from tropical countries. Heliyon 2019, 5, e02309. [Google Scholar] [CrossRef] [PubMed]
- Wimalawansa, S.J. Biology of vitamin D. J. Steroids Horm Sci. 2019, 10, 2. [Google Scholar] [CrossRef]
- Adams, J.S.; Clemens, T.L.; Parrish, J.A.; Holick, M.F. Vitamin-D synthesis and metabolism after ultraviolet irradiation of normal and vitamin-D-deficient subjects. N. Engl. J. Med. 1982, 306, 722–725. [Google Scholar] [CrossRef]
- Vieth, R. Vitamin D supplementation, 25-hydroxyvitamin D concentrations, and safety. Am. J. Clin. Nutr. 1999, 69, 842–856. [Google Scholar] [CrossRef]
- Hollis, B.W.; Johnson, D.; Hulsey, T.C.; Ebeling, M.; Wagner, C.L. Vitamin D supplementation during pregnancy: Double-blind, randomized clinical trial of safety and effectiveness. J. Bone Miner. Res. 2011, 26, 2341–2357. [Google Scholar] [CrossRef]
- Nykjaer, A.; Dragun, D.; Walther, D.; Vorum, H.; Jacobsen, C.; Herz, J.; Melsen, F.; Christensen, E.I.; Willnow, T.E. An endocytic pathway essential for renal uptake and activation of the steroid 25-(OH) vitamin D3. Cell 1999, 96, 507–515. [Google Scholar] [CrossRef]
- Aatsinki, S.M.; Elkhwanky, M.S.; Kummu, O.; Karpale, M.; Buler, M.; Viitala, P.; Rinne, V.; Mutikainen, M.; Tavi, P.; Franko, A.; et al. Fasting-Induced Transcription Factors Repress Vitamin D Bioactivation, a Mechanism for Vitamin D Deficiency in Diabetes. Diabetes 2019, 68, 918–931. [Google Scholar] [CrossRef]
- Elkhwanky, M.S.; Kummu, O.; Piltonen, T.T.; Laru, J.; Morin-Papunen, L.; Mutikainen, M.; Tavi, P.; Hakkola, J. Obesity represses CYP2R1, the vitamin D 25-Hydroxylase, in the liver and extrahepatic tissues. JBMR Plus 2020, 4, e10397. [Google Scholar] [CrossRef]
- Haddad, J.G.; Hillman, L.; Rojanasathit, S. Human serum binding capacity and affinity for 25-hydroxyergocalciferol and 25-hydroxycholecalciferol. J. Clin. Endocrinol. Metab. 1976, 43, 86–91. [Google Scholar] [CrossRef]
- Keenan, M.J.; Holmes, R.P. The uptake and metabolism of 25-hydroxyvitamin D3 and vitamin D binding protein by cultured porcine kidney cells (LLC-PK1). Int. J. Biochem. 1991, 23, 1225–1230. [Google Scholar] [CrossRef]
- Kissmeyer, A.; Mathiasen, I.S.; Latini, S.; Binderup, L. Pharmacokinetic studies of vitamin D analogues: Relationship to vitamin D binding protein (DBP). Endocrine 1995, 3, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Vieth, R.; Kessler, M.J.; Pritzker, K.P. Species differences in the binding kinetics of 25-hydroxyvitamin D3 to vitamin D binding protein. Can. J. Physiol. Pharmacol. 1990, 68, 1368–1371. [Google Scholar] [CrossRef] [PubMed]
- Wimalawansa, S.J. Non-musculoskeletal benefits of vitamin D. J. Steroid Biochem. Mol. Biol. 2018, 175, 60–81. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, M.; Han, C.; Wen, Z.; Meng, X.; Qi, D.; Wang, N.; Du, H.; Wang, J.; Lu, L.; et al. Intraduodenal Delivery of Exosome-Loaded SARS-CoV-2 RBD mRNA Induces a Neutralizing Antibody Response in Mice. Vaccines 2023, 11, 673. [Google Scholar] [CrossRef]
- Jiang, Y.; Chen, L.; Taylor, R.N.; Li, C.; Zhou, X. Physiological and pathological implications of retinoid action in the endometrium. J. Endocrinol. 2018, 236, R169–R188. [Google Scholar] [CrossRef]
- Keane, K.N.; Cruzat, V.F.; Calton, E.K.; Hart, P.H.; Soares, M.J.; Newsholme, P.; Yovich, J.L. Molecular actions of vitamin D in reproductive cell biology. Reproduction 2017, 153, R29–R42. [Google Scholar] [CrossRef]
- Holick, M.F. The cutaneous photosynthesis of previtamin D3: A unique photoendocrine system. J. Investig. Dermatol. 1981, 77, 51–58. [Google Scholar] [CrossRef]
- Haddad, J.G.; Matsuoka, L.Y.; Hollis, B.W.; Hu, Y.Z.; Wortsman, J. Human plasma transport of vitamin D after its endogenous synthesis. J. Clin. Investig. 1993, 91, 2552–2555. [Google Scholar] [CrossRef]
- Ponchon, G.; Kennan, A.L.; DeLuca, H.F. “Activation” of vitamin D by the liver. J. Clin. Investig. 1969, 48, 2032–2037. [Google Scholar] [CrossRef]
- Hosseinpour, F.; Wikvall, K. Porcine microsomal vitamin D(3) 25-hydroxylase (CYP2D25). Catalytic properties, tissue distribution, and comparison with human CYP2D6. J. Biol. Chem. 2000, 275, 34650–34655. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, J.N.; Young, M.V.; Persons, K.S.; Wang, L.; Mathieu, J.S.; Whitlatch, L.W.; Holick, M.F.; Chen, T.C. Vitamin D metabolism in human prostate cells: Implications for prostate cancer chemoprevention by vitamin D. Anticancer Res. 2006, 26, 2567–2572. [Google Scholar] [PubMed]
- Zhu, P.; Ren, M.; Yang, C.; Hu, Y.X.; Ran, J.M.; Yan, L. Involvement of RAGE, MAPK and NF-kappaB pathways in AGEs-induced MMP-9 activation in HaCaT keratinocytes. Exp. Dermatol. 2012, 21, 123–129. [Google Scholar] [CrossRef]
- McBrearty, N.; Cho, C.; Chen, J.; Zahedi, F.; Peck, A.R.; Radaelli, E.; Assenmacher, C.A.; Pavlak, C.; Devine, A.; Yu, P.; et al. Tumor-Suppressive and Immune-Stimulating Roles of Cholesterol 25-hydroxylase in Pancreatic Cancer Cells. Mol. Cancer Res. 2023, 21, 228–239. [Google Scholar] [CrossRef]
- Karlgren, M.; Miura, S.; Ingelman-Sundberg, M. Novel extrahepatic cytochrome P450s. Toxicol. Appl. Pharmacol. 2005, 207, 57–61. [Google Scholar] [CrossRef]
- Smith, J.E.; Goodman, D.S. The turnover and transport of vitamin D and of a polar metabolite with the properties of 25-hydroxycholecalciferol in human plasma. J. Clin. Investig. 1971, 50, 2159–2167. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.W.; Paris, P.W.; Clemens, T.L.; Nolan, J.; Holick, M.F. Vitamin D absorption in healthy subjects and in patients with intestinal malabsorption syndromes. Am. J. Clin. Nutr. 1985, 42, 644–649. [Google Scholar] [CrossRef]
- Heaney, R.P.; Recker, R.R.; Grote, J.; Horst, R.L.; Armas, L.A. Vitamin D(3) is more potent than vitamin D(2) in humans. J. Clin. Endocrinol. Metab. 2011, 96, E447–E452. [Google Scholar] [CrossRef]
- Mangin, M.; Sinha, R.; Fincher, K. Inflammation and vitamin D: The infection connection. Inflamm. Res. 2014, 63, 803–819. [Google Scholar] [CrossRef]
- Chung, M.K.; Karnik, S.; Saef, J.; Bergmann, C.; Barnard, J.; Lederman, M.M.; Tilton, J.; Cheng, F.; Harding, C.V.; Young, J.B.; et al. SARS-CoV-2 and ACE2: The biology and clinical data settling the ARB and ACEI controversy. EBioMedicine 2020, 58, 102907. [Google Scholar] [CrossRef]
- Wimalawansa, S.J. Physiological basis for using vitamin D to improve health. Biomedicines 2023, 11, 1542. [Google Scholar] [CrossRef] [PubMed]
- Arai, Y.; Kanda, E.; Iimori, S.; Naito, S.; Noda, Y.; Kawasaki, T.; Sato, H.; Ando, R.; Sasaki, S.; Sohara, E.; et al. The use of vitamin D analogs is independently associated with the favorable renal prognosis in chronic kidney disease stages 4-5: The CKD-ROUTE study. Clin. Exp. Nephrol. 2017, 21, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Oh, T.R.; Kim, C.S.; Bae, E.H.; Ma, S.K.K.; Han, S.H.; Sung, S.A.; Lee, K.; Oh, K.H.; Ahn, C.; Kim, S.W.; et al. Association between vitamin D deficiency and health-related quality of life in patients with chronic kidney disease from the KNOW-CKD study. PLoS ONE 2017, 12, e0174282. [Google Scholar] [CrossRef]
- Zittermann, A.; Iodice, S.; Pilz, S.; Grant, W.B.; Bagnardi, V.; Gandini, S. Vitamin D deficiency and mortality risk in the general population: A meta-analysis of prospective cohort studies. Am. J. Clin. Nutr. 2012, 95, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Jean, G.; Souberbielle, J.C.; Chazot, C. Vitamin D in Chronic Kidney Disease and Dialysis Patients. Nutrients 2017, 9, 328. [Google Scholar] [CrossRef] [PubMed]
- Waziri, B.; Duarte, R.; Naicker, S. Chronic Kidney Disease-Mineral and Bone Disorder (CKD-MBD): Current Perspectives. Int. J. Nephrol. Renovasc. Dis. 2019, 12, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Urena-Torres, P.; Metzger, M.; Haymann, J.P.; Karras, A.; Boffa, J.J.; Flamant, M.; Vrtovsnik, F.; Gauci, C.; Froissart, M.; Houillier, P.; et al. Association of kidney function, vitamin D deficiency, and circulating markers of mineral and bone disorders in CKD. Am. J. Kidney Dis. 2011, 58, 544–553. [Google Scholar] [CrossRef]
- Kanai, G.; Fukagawa, M. CKD-MBD (Chronic Kidney Disease-Mineral and Bone Disorder). Gene therapy for secondary hyperparathyroidism. Clin. Calcium 2010, 20, 1052–1059. [Google Scholar]
- Zelnick, L.R.; de Boer, I.H.; Kestenbaum, B.R.; Chonchol, M.; Kendrick, J. Comparative Effects of Cholecalciferol and Calcitriol on Circulating Markers of CKD Mineral Bone Disorder: A Randomized Clinical Trial. Clin. J. Am. Soc. Nephrol. 2018, 13, 927–928. [Google Scholar] [CrossRef]
- Bansal, N.; Katz, R.; Appel, L.; Denburg, M.; Feldman, H.; Go, A.S.; He, J.; Hoofnagle, A.; Isakova, T.; Kestenbaum, B.; et al. Vitamin D Metabolic Ratio and Risks of Death and CKD Progression. Kidney Int. Rep. 2019, 4, 1598–1607. [Google Scholar] [CrossRef]
- Durazo-Arvizu, R.A.; Dawson-Hughes, B.; Kramer, H.; Cao, G.; Merkel, J.; Coates, P.M.; Sempos, C.T. The reverse J-shaped association between serum total 25-hydroxyvitamin D concentration and all-cause mortality: The impact of assay standardization. Am. J. Epidemiol. 2017, 185, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Durup, D.; Jorgensen, H.L.; Christensen, J.; Schwarz, P.; Heegaard, A.M.; Lind, B. A reverse J-shaped association of all-cause mortality with serum 25-hydroxyvitamin D in general practice: The CopD study. J. Clin. Endocrinol. Metab. 2012, 97, 2644–2652. [Google Scholar] [CrossRef] [PubMed]
- Grant, W.B. Letter to the Editor: The J-shaped 25-hydroxyvitamin D concentration-cardiovascular disease mortality relation is very likely due to starting vitamin D supplementation late in life. J. Clin. Endocrinol. Metab. 2015, 100, L49–L50. [Google Scholar] [CrossRef] [PubMed]
- Marzolo, M.P.; Farfan, P. New insights into the roles of megalin/LRP2 and the regulation of its functional expression. Biol. Res. 2011, 44, 89–105. [Google Scholar] [CrossRef]
- Wimalawansa, S. Overcoming infections including COVID-19, by maintaining circulating 25(OH)D concentrations above 50 ng/mL. Pathol. Lab. Med. Int. 2022, 14, 37–60. [Google Scholar] [CrossRef]
- Wimalawansa, S.J. Rapidly Increasing Serum 25(OH)D Boosts the Immune System, against Infections-Sepsis and COVID-19. Nutrients 2022, 14, 2997. [Google Scholar] [CrossRef]
- Ianevski, A.; Zusinaite, E.; Shtaida, N.; Kallio-Kokko, H.; Valkonen, M.; Kantele, A.; Telling, K.; Lutsar, I.; Letjuka, P.; Metelitsa, N.; et al. Low Temperature and Low UV Indexes Correlated with Peaks of Influenza Virus Activity in Northern Europe during 2010–2018. Viruses 2019, 11, 207. [Google Scholar] [CrossRef]
- Imai, C.M.; Halldorsson, T.I.; Eiriksdottir, G.; Cotch, M.F.; Steingrimsdottir, L.; Thorsdottir, I.; Launer, L.J.; Harris, T.; Gudnason, V.; Gunnarsdottir, I. Depression and serum 25-hydroxyvitamin D in older adults living at northern latitudes—AGES-Reykjavik Study. J. Nutr. Sci. 2015, 4, e37. [Google Scholar] [CrossRef]
- Devaraj, S.; Jialal, G.; Cook, T.; Siegel, D.; Jialal, I. Low vitamin D levels in Northern American adults with the metabolic syndrome. Horm. Metab. Res. 2011, 43, 72–74. [Google Scholar] [CrossRef]
- Reichrath, J.; Saternus, R.; Vogt, T. Challenge and perspective: The relevance of ultraviolet (UV) radiation and the vitamin D endocrine system (VDES) for psoriasis and other inflammatory skin diseases. Photochem. Photobiol. Sci. 2017, 16, 433–444. [Google Scholar] [CrossRef]
- Premkumar, M.; Sable, T.; Dhanwal, D.; Dewan, R. Vitamin D homeostasis, bone mineral metabolism, and seasonal affective disorder during 1 year of Antarctic residence. Arch. Osteoporos. 2013, 8, 129. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, G.L.; Cendoroglo, M.S.; Ramos, L.R.; Araujo, L.M.; Vieira, J.G.; Kunii, I.; Hayashi, L.F.; Correa, M.P.; Lazaretti-Castro, M. Influence of ultraviolet radiation on the production of 25 hydroxyvitamin D in the elderly population in the city of Sao Paulo (23 degrees 34′S), Brazil. Osteoporos. Int. 2005, 16, 1649–1654. [Google Scholar] [CrossRef] [PubMed]
- Pletz, M.W.; Terkamp, C.; Schumacher, U.; Rohde, G.; Schutte, H.; Welte, T.; Bals, R.; Group, C.A.-S. Vitamin D deficiency in community-acquired pneumonia: Low levels of 1,25(OH)2 D are associated with disease severity. Respir. Res. 2014, 15, 53. [Google Scholar] [CrossRef]
- Ginde, A.A.; Mansbach, J.M.; Camargo, C.A., Jr. Vitamin D, respiratory infections, and asthma. Curr. Allergy Asthma Rep. 2009, 9, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Czaja, A.J. Factoring the intestinal microbiome into the pathogenesis of autoimmune hepatitis. World J. Gastroenterol. 2016, 22, 9257–9278. [Google Scholar] [CrossRef]
- Jin, D.; Wu, S.; Zhang, Y.G.; Lu, R.; Xia, Y.; Dong, H.; Sun, J. Lack of Vitamin D Receptor Causes Dysbiosis and Changes the Functions of the Murine Intestinal Microbiome. Clin. Ther. 2015, 37, 996–1009 e1007. [Google Scholar] [CrossRef]
- Zhou, Y.F.; Luo, B.A.; Qin, L.L. The association between vitamin D deficiency and community-acquired pneumonia: A meta-analysis of observational studies. Medicine 2019, 98, e17252. [Google Scholar] [CrossRef]
- Laplana, M.; Royo, J.L.L.; Fibla, J. Vitamin D Receptor polymorphisms and risk of enveloped virus infection: A meta-analysis. Gene 2018, 678, 384–394. [Google Scholar] [CrossRef]
- Platitsyna, N.G.; Bolotnova, T.V. Vitamin D deficiency as a risk factor for chronic non-infectious diseases. Adv. Gerontol. 2017, 30, 873–879. [Google Scholar]
- Grant, W.B.; Lahore, H.; McDonnell, S.L.; Baggerly, C.A.; French, C.B.; Aliano, J.L.; Bhattoa, H.P. Evidence that Vitamin D Supplementation Could Reduce Risk of Influenza and COVID-19 Infections and Deaths. Nutrients 2020, 12, 1626. [Google Scholar] [CrossRef]
- Eroglu, C.; Demir, F.; Erge, D.; Uysal, P.; Kirdar, S.; Yilmaz, M.; Kurt Omurlu, I. The relation between serum vitamin D levels, viral infections and severity of attacks in children with recurrent wheezing. Allergol. Immunopathol. (Madr.) 2019, 47, 591–597. [Google Scholar] [CrossRef]
- Arihiro, S.; Nakashima, A.; Matsuoka, M.; Suto, S.; Uchiyama, K.; Kato, T.; Mitobe, J.; Komoike, N.; Itagaki, M.; Miyakawa, Y.; et al. Randomized Trial of Vitamin D Supplementation to Prevent Seasonal Influenza and Upper Respiratory Infection in Patients with Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2019, 25, 1088–1095. [Google Scholar] [CrossRef]
- Jolliffe, D.A.; Greiller, C.L.; Mein, C.A.; Hoti, M.; Bakhsoliani, E.; Telcian, A.G.; Simpson, A.; Barnes, N.C.; Curtin, J.A.; Custovic, A.; et al. Vitamin D receptor genotype influences risk of upper respiratory infection. Br. J. Nutr. 2018, 120, 891–900. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.L.; Hollis, B.W.; Kotsa, K.; Fakhoury, H.; Karras, S.N. Vitamin D administration during pregnancy as prevention for pregnancy, neonatal and postnatal complications. Rev. Endocr. Metab. Disord. 2017, 18, 307–322. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.K.; Pell, L.G.; Rahman, M.Z.; Dimitris, M.C.; Mahmud, A.; Islam, M.M.; Ahmed, T.; Pullenayegum, E.; Kashem, T.; Shanta, S.S.; et al. Maternal vitamin D supplementation during pregnancy and lactation to prevent acute respiratory infections in infancy in Dhaka, Bangladesh (MDARI trial): Protocol for a prospective cohort study nested within a randomized controlled trial. BMC Pregnancy Childbirth 2016, 16, 309. [Google Scholar] [CrossRef] [PubMed]
- Wimalawansa, S.J.; Polonowita, A. Boosting immunity with vitamin D for preventing complications and deaths from COVID-19. In Proceedings of the COVID 19: Impact, Mitigation, Opportunities and Building Resilience “From Adversity to Serendipity”, Perspectives of Global Relevance Based on Research, Experience and Successes in Combating COVID-19 in Sri Lanka, Colombo, Sri Lanka, 27–28 January 2021; pp. 171–198. [Google Scholar]
- Kone-Paut, I.; Cimaz, R. Is it Kawasaki shock syndrome, Kawasaki-like disease or pediatric inflammatory multisystem disease? The importance of semantic in the era of COVID-19 pandemic. RMD Open 2020, 6, e001333. [Google Scholar] [CrossRef]
- Pouletty, M.; Borocco, C.; Ouldali, N.; Caseris, M.; Basmaci, R.; Lachaume, N.; Bensaid, P.; Pichard, S.; Kouider, H.; Morelle, G.; et al. Paediatric multisystem inflammatory syndrome temporally associated with SARS-CoV-2 mimicking Kawasaki disease (Kawa-COVID-19): A multicentre cohort. Ann. Rheum. Dis. 2020, 79, 999–1006. [Google Scholar] [CrossRef]
- Stagi, S.; Rigante, D.; Lepri, G.; Matucci Cerinic, M.; Falcini, F. Severe vitamin D deficiency in patients with Kawasaki disease: A potential role in the risk to develop heart vascular abnormalities? Clin. Rheumatol. 2016, 35, 1865–1872. [Google Scholar] [CrossRef]
- Torpoco Rivera, D.; Misra, A.; Sanil, Y.; Sabzghabaei, N.; Safa, R.; Garcia, R.U. Vitamin D and morbidity in children with Multisystem inflammatory syndrome related to COVID-19. Prog Pediatr Cardiol. 2022, 66, 101507. Available online: https://www.ncbi.nlm.nih.gov/pubmed/35250251 (accessed on 14 August 2023). [CrossRef]
- Oliver, S.M. The immune system and new therapies for inflammatory joint disease. Musculoskelet. Care 2003, 1, 44–57. [Google Scholar] [CrossRef]
- Wimalawansa, S.J. Global epidemic of coronavirus—COVID-19: What can we do to minimize risks? Eur. J. Biomed. Pharma Sci. 2020, 7, 432–438. [Google Scholar]
- Wimalawansa, S.J.; Razzaque, M.S.; Al-Daghri, N.M. Calcium and vitamin D in human health: Hype or real? J. Steroid Biochem. Mol. Biol. 2018, 180, 4–14. [Google Scholar] [CrossRef]
- Soltani-Zangbar, M.S.; Mahmoodpoor, A.; Dolati, S.; Shamekh, A.; Valizadeh, S.; Yousefi, M.; Sanaie, S. Serum levels of vitamin D and immune system function in patients with COVID-19 admitted to intensive care unit. Gene Rep. 2022, 26, 101509. [Google Scholar] [CrossRef]
- Arora, J.; Wang, J.; Weaver, V.; Zhang, Y.; Cantorna, M.T. Novel insight into the role of the vitamin D receptor in the development and function of the immune system. J. Steroid Biochem. Mol. Biol. 2022, 219, 106084. [Google Scholar] [CrossRef] [PubMed]
- Quraishi, S.A.; Bittner, E.A.; Blum, L.; McCarthy, C.M.; Bhan, I.; Camargo, C.A., Jr. Prospective study of vitamin D status at initiation of care in critically ill surgical patients and risk of 90-day mortality. Crit. Care Med. 2014, 42, 1365–1371. [Google Scholar] [CrossRef]
- Chauss, D.; Freiwald, T.; McGregor, R.; Yan, B.; Wang, L.; Nova-Lamperti, E.; Kumar, D.; Zhang, Z.; Teague, H.; West, E.E.; et al. Autocrine vitamin D signaling switches off pro-inflammatory programs of T(H)1 cells. Nat. Immunol. 2022, 23, 62–74. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Arbesman, J.; Piliang, M. Vitamin D, autoimmunity and immune-related adverse events of immune checkpoint inhibitors. Arch. Dermatol. Res. 2021, 313, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.R.; Thacher, T.D. Vitamin D: Immune function, inflammation, infections and auto-immunity. Paediatr. Int. Child Health 2023, 1–11. [Google Scholar] [CrossRef] [PubMed]
- McCartney, D.M.; Byrne, D.G. Optimisation of vitamin D status for enhanced immuno-protection against COVID-19. Ir. Med. J. 2020, 113, 58. [Google Scholar] [PubMed]
- Tsujino, I.; Ushikoshi-Nakayama, R.; Yamazaki, T.; Matsumoto, N.; Saito, I. Pulmonary activation of vitamin D(3) and preventive effect against interstitial pneumonia. J. Clin. Biochem. Nutr. 2019, 65, 245–251. [Google Scholar] [CrossRef]
- Holick, M.F.; Binkley, N.C.; Bischoff-Ferrari, H.A.; Gordon, C.M.; Hanley, D.A.; Heaney, R.P.; Murad, M.H.; Weaver, C.M.; Endocrine, S. Evaluation, treatment, and prevention of vitamin D deficiency: An Endocrine Society clinical practice guideline. J. Clin. Endocrinol. Metab. 2011, 96, 1911–1930. [Google Scholar] [CrossRef] [PubMed]
- Anonymus. Vitamin D for COVID-19: Real-Time Analysis of All 300 Studies. Available online: https://c19vitamind.com/ (accessed on 25 January 2023).
- Adams, J.S.; Modlin, R.L.; Diz, M.M.M.; Barnes, P.F. Potentiation of the macrophage 25-hydroxyvitamin D-1-hydroxylation reaction by human tuberculous pleural effusion fluid. J. Clin. Endocrinol. Metab. 1989, 69, 457–460. [Google Scholar] [CrossRef] [PubMed]
- Antal, A.S.; Dombrowski, Y.; Koglin, S.; Ruzicka, T.; Schauber, J. Impact of vitamin D3 on cutaneous immunity and antimicrobial peptide expression. Dermatoendocrinology 2011, 3, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Ahorsu, D.K.; Imani, V.; Lin, C.Y.; Timpka, T.; Brostrom, A.; Updegraff, J.A.; Arestedt, K.; Griffiths, M.D.; Pakpour, A.H. Associations between Fear of COVID-19, Mental Health, and Preventive Behaviours Across Pregnant Women and Husbands: An Actor-Partner Interdependence Modelling. Int. J. Ment. Health Addict. 2022, 20, 68–82. [Google Scholar] [CrossRef] [PubMed]
- Aloia, J.F.; Li-Ng, M. Re: Epidemic influenza and vitamin D. Epidemiol. Infect. 2007, 135, 1095–1096. [Google Scholar] [CrossRef]
- Fleming, D.M.; Elliot, A.J. Epidemic influenza and vitamin D. Epidemiol. Infect. 2007, 135, 1091–1092. [Google Scholar] [CrossRef]
- Ali, M.; Uddin, Z. Factors associated with vitamin D deficiency among patients with musculoskeletal disorders seeking physiotherapy intervention: A hospital-based observational study. BMC Musculoskelet. Disord. 2022, 23, 817. [Google Scholar] [CrossRef]
- Dawodu, A.; Saadi, H.F.F.; Bekdache, G.; Javed, Y.; Altaye, M.; Hollis, B.W. Randomized controlled trial (RCT) of vitamin D supplementation in pregnancy in a population with endemic vitamin D deficiency. J. Clin. Endocrinol. Metab. 2013, 98, 2337–2346. [Google Scholar] [CrossRef]
- Camargo, C.A., Jr.; Ganmaa, D.; Frazier, A.L.; Kirchberg, F.F.; Stuart, J.J.; Kleinman, K.; Sumberzul, N.; Rich-Edwards, J.W. Randomized trial of vitamin D supplementation and risk of acute respiratory infection in Mongolia. Pediatrics 2012, 130, e561–e567. [Google Scholar] [CrossRef]
- Bergman, P.; Norlin, A.C.; Hansen, S.; Rekha, R.S.; Agerberth, B.; Bjorkhem-Bergman, L.; Ekstrom, L.; Lindh, J.D.; Andersson, J. Vitamin D3 supplementation in patients with frequent respiratory tract infections: A randomised and double-blind intervention study. BMJ Open 2012, 2, e001663. [Google Scholar] [CrossRef]
- Hutchinson, M.S.; Grimnes, G.; Joakimsen, R.M.; Figenschau, Y.; Jorde, R. Low serum 25-hydroxyvitamin D levels are associated with increased all-cause mortality risk in a general population: The Tromso study. Eur. J. Endocrinol. 2010, 162, 935–942. [Google Scholar] [CrossRef]
- Murdoch, D.R.; Slow, S.; Chambers, S.T.; Jennings, L.C.; Stewart, A.W.; Priest, P.C.; Florkowski, C.M.; Livesey, J.H.; Camargo, C.A.; Scragg, R. Effect of vitamin D3 supplementation on upper respiratory tract infections in healthy adults: The VIDARIS randomized controlled trial. JAMA 2012, 308, 1333–1339. [Google Scholar] [CrossRef]
- Manaseki-Holland, S.; Maroof, Z.; Bruce, J.; Mughal, M.Z.Z.; Masher, M.I.I.; Bhutta, Z.A.; Walraven, G.; Chandramohan, D. Effect on the incidence of pneumonia of vitamin D supplementation by quarterly bolus dose to infants in Kabul: A randomised controlled superiority trial. Lancet 2012, 379, 1419–1427. [Google Scholar] [CrossRef] [PubMed]
- Martineau, A.R.; Timms, P.M.; Bothamley, G.H.; Hanifa, Y.; Islam, K.; Claxton, A.P.; Packe, G.E.; Moore-Gillon, J.C.; Darmalingam, M.; Davidson, R.N.; et al. High-dose vitamin D(3) during intensive-phase antimicrobial treatment of pulmonary tuberculosis: A double-blind randomised controlled trial. Lancet 2011, 377, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.C.; Manson, J.E.; Abrams, S.A.; Aloia, J.F.; Brannon, P.M.; Clinton, S.K.; Durazo-Arvizu, R.A.; Gallagher, J.C.; Gallo, R.L.; Jones, G.; et al. The 2011 report on dietary reference intakes for calcium and vitamin D from the Institute of Medicine: What clinicians need to know. J. Clin. Endocrinol. Metab. 2011, 96, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Kimball, S.; Vieth, R.; Dosch, H.M.; Bar-Or, A.; Cheung, R.; Gagne, D.; O’Connor, P.; D’Souza, C.; Ursell, M.; Burton, J.M. Cholecalciferol plus calcium suppresses abnormal PBMC reactivity in patients with multiple sclerosis. J. Clin. Endocrinol. Metab. 2011, 96, 2826–2834. [Google Scholar] [CrossRef]
- Heaney, R.P.; Davies, K.M.; Chen, T.C.; Holick, M.F.; Barger-Lux, M.J. Human serum 25-hydroxycholecalciferol response to extended oral dosing with cholecalciferol. Am. J. Clin. Nutr. 2003, 77, 204–210. [Google Scholar] [CrossRef]
- Sanders, K.M.; Stuart, A.L.; Williamson, E.J.; Simpson, J.A.; Kotowicz, M.A.; Young, D.; Nicholson, G.C. Annual high-dose oral vitamin D and falls and fractures in older women: A randomized controlled trial. JAMA 2010, 303, 1815–1822. [Google Scholar] [CrossRef]
- Nowson, C.A.; McGrath, J.J.; Ebeling, P.R.; Haikerwal, A.; Daly, R.M.; Sanders, K.M.; Seibel, M.J.; Mason, R.S.; Working Group of Australian and New Zealand Bone and Mineral Society, Endocrine Society of Australia and Osteoporosis Australia. Vitamin D and health in adults in Australia and New Zealand: A position statement. Med. J. Aust. 2012, 196, 686–687. [Google Scholar] [CrossRef]
- Schwalfenberg, G.K.; Whiting, S.J. A Canadian response to the 2010 Institute of Medicine vitamin D and calcium guidelines. Public Health Nutr. 2011, 14, 746–748. [Google Scholar] [CrossRef]
- Lappe, J.M.; Travers-Gustafson, D.; Davies, K.M.; Recker, R.R.; Heaney, R.P. Vitamin D and calcium supplementation reduces cancer risk: Results of a randomized trial. Am. J. Clin. Nutr. 2007, 85, 1586–1591. [Google Scholar] [CrossRef]
- Marshall, D.T.; Savage, S.J.; Garrett-Mayer, E.; Keane, T.E.; Hollis, B.W.; Horst, R.L.; Ambrose, L.H.; Kindy, M.S.S.; Gattoni-Celli, S. Vitamin D3 supplementation at 4000 international units per day for one year results in a decrease of positive cores at repeat biopsy in subjects with low-risk prostate cancer under active surveillance. J. Clin. Endocrinol. Metab. 2012, 97, 2315–2324. [Google Scholar] [CrossRef] [PubMed]
- Forman, J.P.; Scott, J.B.; Ng, K.; Drake, B.F.; Suarez, E.G.; Hayden, D.L.; Bennett, G.G.; Chandler, P.D.; Hollis, B.W.; Emmons, K.M.; et al. Effect of vitamin D supplementation on blood pressure in blacks. Hypertension 2013, 61, 779–785. [Google Scholar] [CrossRef] [PubMed]
- Mitri, J.; Dawson-Hughes, B.; Hu, F.B.; Pittas, A.G. Effects of vitamin D and calcium supplementation on pancreatic beta cell function, insulin sensitivity, and glycemia in adults at high risk of diabetes: The Calcium and Vitamin D for Diabetes Mellitus (CaDDM) randomized controlled trial. Am. J. Clin. Nutr. 2011, 94, 486–494. [Google Scholar] [CrossRef]
- von Hurst, P.R.; Stonehouse, W.; Coad, J. Vitamin D supplementation reduces insulin resistance in South Asian women living in New Zealand who are insulin resistant and vitamin D deficient—A randomised, placebo-controlled trial. Br. J. Nutr. 2010, 103, 549–555. [Google Scholar] [CrossRef]
- Belenchia, A.M.; Tosh, A.K.; Hillman, L.S.; Peterson, C.A. Correcting vitamin D insufficiency improves insulin sensitivity in obese adolescents: A randomized controlled trial. Am. J. Clin. Nutr. 2013, 97, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Derakhshandi, H.; Etemadifar, M.; Feizi, A.; Abtahi, S.H.; Minagar, A.; Abtahi, M.A.; Abtahi, Z.A.; Dehghani, A.; Sajjadi, S.; Tabrizi, N. Preventive effect of vitamin D3 supplementation on conversion of optic neuritis to clinically definite multiple sclerosis: A double blind, randomized, placebo-controlled pilot clinical trial. Acta Neurol. Belg. 2013, 113, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Hollis, B.W.; Wagner, C.L. Vitamin D requirements during lactation: High-dose maternal supplementation as therapy to prevent hypovitaminosis D for both the mother and the nursing infant. Am. J. Clin. Nutr. 2004, 80, 1752S–1758S. [Google Scholar] [CrossRef]
- Trivedi, D.; Doll, R.; Khaw, K.T. Effect of four monthly oral vitamin D3 (cholecalciferol) supplementation on fractures and mortality in men and women living in the community: Randomised double blind controlled trial. BMJ 2003, 326, 469. [Google Scholar] [CrossRef]
- Johansson, H.; Oden, A.; Kanis, J.; McCloskey, E.; Lorentzon, M.; Ljunggren, O.; Karlsson, M.K.; Thorsby, P.M.; Tivesten, A.; Barrett-Connor, E.; et al. Low serum vitamin D is associated with increased mortality in elderly men: MrOS Sweden. Osteoporos. Int. 2012, 23, 991–999. [Google Scholar] [CrossRef]
- Hollis, B.W.; Wagner, C.L. Clinical review: The role of the parent compound vitamin D with respect to metabolism and function: Why clinical dose intervals can affect clinical outcomes. J. Clin. Endocrinol. Metab. 2013, 98, 4619–4628. [Google Scholar] [CrossRef]
- Morris, H.A.; Anderson, P.H. Autocrine and paracrine actions of vitamin d. Clin. Biochem. Rev. 2010, 31, 129–138. [Google Scholar]
- Atkinson, R.L. Viruses as an etiology of obesity. Mayo Clin. Proc. 2007, 82, 1192–1198. [Google Scholar] [CrossRef] [PubMed]
- McGregor, R.; Chauss, D.; Freiwald, T.; Yan, B.; Wang, L.; Nova-Lamperti, E.; Zhang, Z.; Teague, H.; West, E.E.E.; Bibby, J.; et al. An autocrine Vitamin D-driven Th1 shutdown program can be exploited for COVID-19. bioRxiv 2020. [Google Scholar] [CrossRef]
- Gibson, C.C.; Davis, C.T.; Zhu, W.; Bowman-Kirigin, J.A.; Walker, A.E.; Tai, Z.; Thomas, K.R.; Donato, A.J.; Lesniewski, L.A.; Li, D.Y. Dietary Vitamin D and Its Metabolites Non-Genomically Stabilize the Endothelium. PLoS ONE 2015, 10, e0140370. [Google Scholar] [CrossRef]
- Moromizato, T.; Litonjua, A.A.; Braun, A.B.; Gibbons, F.K.; Giovannucci, E.; Christopher, K.B. Association of low serum 25-hydroxyvitamin D levels and sepsis in the critically ill. Crit. Care Med. 2014, 42, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Tolppanen, A.M.; Fraser, A.; Fraser, W.D.; Lawlor, D.A. Risk factors for variation in 25-hydroxyvitamin D(3) and D(2) concentrations and vitamin D deficiency in children. J. Clin. Endocrinol. Metab. 2012, 97, 1202–1210. [Google Scholar] [CrossRef]
- Bae, J.H.; Choe, H.J.; Holick, M.F.; Lim, S. Association of vitamin D status with COVID-19 and its severity: Vitamin D and COVID-19: A narrative review. Rev. Endocr. Metab. Disord. 2022, 23, 579–599. [Google Scholar] [CrossRef]
- Holick, M.F. Sunlight, UV Radiation, Vitamin D, and Skin Cancer: How Much Sunlight Do We Need? Adv. Exp. Med. Biol. 2020, 1268, 19–36. [Google Scholar] [CrossRef]
- Heaney, R.; Armas, L.; Shary, J.; Bell, N.; Binkley, N.; Hollis, B. 25-Hydroxylation of vitamin D3: Relation to circulating vitamin D3 under various input conditions. Am. J. Clin. Nutr. 2008, 87, 1738–1742. [Google Scholar] [CrossRef]
- Hollis, B.W. Short-term and long-term consequences and concerns regarding valid assessment of vitamin D deficiency: Comparison of recent food supplementation and clinical guidance reports. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 598–604. [Google Scholar] [CrossRef] [PubMed]
- Urashima, M.; Segawa, T.; Okazaki, M.; Kurihara, M.; Wada, Y.; Ida, H. Randomized trial of vitamin D supplementation to prevent seasonal influenza A in schoolchildren. Am. J. Clin. Nutr. 2010, 91, 1255–1260. [Google Scholar] [CrossRef] [PubMed]
- Bergman, P.; Lindh, A.U.; Bjorkhem-Bergman, L.; Lindh, J.D. Vitamin D and Respiratory Tract Infections: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. PLoS ONE 2013, 8, e65835. [Google Scholar] [CrossRef] [PubMed]
- Borel, P.; Caillaud, D.; Cano, N.J. Vitamin D bioavailability: State of the art. Crit. Rev. Food Sci. Nutr. 2015, 55, 1193–1205. [Google Scholar] [CrossRef] [PubMed]
- Greer, F.R.; Hollis, B.W.; Napoli, J.L. High concentrations of vitamin D2 in human milk associated with pharmacologic doses of vitamin D2. J. Pediatr. 1984, 105, 61–64. [Google Scholar] [CrossRef]
- Greer, F.R.; Hollis, B.W.; Cripps, D.J.; Tsang, R.C. Effects of maternal ultraviolet B irradiation on vitamin D content of human milk. J. Pediatr. 1984, 105, 431–433. [Google Scholar] [CrossRef]
- Ziegler, E.E.; Hollis, B.W.; Nelson, S.E.; Jeter, J.M. Vitamin D deficiency in breastfed infants in Iowa. Pediatrics 2006, 118, 603–610. [Google Scholar] [CrossRef]
- Wagner, C.L.; Greer, F.R.; American Academy of Pediatrics Section on Breastfeeding; American Academy of Pediatrics Committee on Nutrition. Prevention of rickets and vitamin D deficiency in infants, children, and adolescents. Pediatrics 2008, 122, 1142–1152. [Google Scholar] [CrossRef]
- Hollis, B.W.; Pittard, W.B., 3rd. Evaluation of the total fetomaternal vitamin D relationships at term: Evidence for racial differences. J. Clin. Endocrinol. Metab. 1984, 59, 652–657. [Google Scholar] [CrossRef]
- Hillman, L.S.; Haddad, J.G. Human perinatal vitamin D metabolism. I. 25-Hydroxyvitamin D in maternal and cord blood. J. Pediatr. 1974, 84, 742–749. [Google Scholar] [CrossRef]
- Heyden, E.L.; Wimalawansa, S.J. Vitamin D: Effects on human reproduction, pregnancy, and fetal well-being. J. Steroid Biochem. Mol. Biol. 2018, 180, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Hanel, A.; Bendik, I.; Carlberg, C. Transcriptome-Wide Profile of 25-Hydroxyvitamin D(3) in Primary Immune Cells from Human Peripheral Blood. Nutrients 2021, 13, 4100. [Google Scholar] [CrossRef] [PubMed]
- Aygun, H. Vitamin D can prevent COVID-19 infection-induced multiple organ damage. Naunyn Schmiedebergs Arch. Pharmacol. 2020, 393, 1157–1160. [Google Scholar] [CrossRef] [PubMed]
- Wimalawansa, S. Commonsense approaches to minimizing risks from COVID-19. J. Pulmonol. Resp. Med. 2020, 2, 28–37. [Google Scholar] [CrossRef]
- Grant, W.B. Variations in vitamin D production could possibly explain the seasonality of childhood respiratory infections in Hawaii. Pediatr. Infect. Dis. J. 2008, 27, 853. [Google Scholar] [CrossRef] [PubMed]
- Martineau, A.R.; Jolliffe, D.A.; Hooper, R.L.; Greenberg, L.; Aloia, J.F.; Bergman, P.; Dubnov-Raz, G.; Esposito, S.; Ganmaa, D.; Ginde, A.A.; et al. Vitamin D supplementation to prevent acute respiratory tract infections: Systematic review and meta-analysis of individual participant data. BMJ 2017, 356, i6583. [Google Scholar] [CrossRef] [PubMed]
- Wimalawansa, S.J. Fighting against COVID-19: Boosting the immunity with micronutrients, stress reduction, physical activity, and vitamin D. Nutr. Food Sci. J. (Sci. Lit.) 2020, 3, 126. [Google Scholar]
- Chetty, V.; Chetty, M. Potential benefit of vitamin D supplementation in people with respiratory illnesses, during the COVID-19 pandemic. Clin. Transl. Sci. 2021, 14, 2111–2116. [Google Scholar] [CrossRef]
- D’Avolio, A.; Avataneo, V.; Manca, A.; Cusato, J.; De Nicolo, A.; Lucchini, R.; Keller, F.; Cantu, M. 25-Hydroxyvitamin D Concentrations Are Lower in Patients with Positive PCR for SARS-CoV-2. Nutrients 2020, 12, 1359. [Google Scholar] [CrossRef]
- Kaufman, H.W.; Niles, J.K.; Kroll, M.H.; Bi, C.; Holick, M.F. SARS-CoV-2 positivity rates associated with circulating 25-hydroxyvitamin D levels. PLoS ONE 2020, 15, e0239252. [Google Scholar] [CrossRef]
- DiNicolantonio, J.; O’Keefe, J. Magnesium and vitamin D deficiency as a potential cause of immune dysfunction, cytokine storm and disseminated intravascular coagulation in covid-19 patients. Mol. Med. 2021, 118, 68–73. [Google Scholar]
- Richardson, S.; Hirsch, J.S.; Narasimhan, M.; Crawford, J.M.; McGinn, T.; Davidson, K.W.; the Northwell, C.-R.C.; Barnaby, D.P.; Becker, L.B.; Chelico, J.D.; et al. Presenting Characteristics, Co-morbidities, and Outcomes among 5700 Patients Hospitalized with COVID-19 in the New York City Area. JAMA 2020, 323, 2052–2059. [Google Scholar] [CrossRef]
- Kurylowicz, A.; Bednarczuk, T.; Nauman, J. The influence of vitamin D deficiency on cancers and autoimmune diseases development. Endokrynol. Pol. 2007, 58, 140–152. [Google Scholar]
- Quraishi, S.A.; Bittner, E.A.; Blum, L.; Hutter, M.M.; Camargo, C.A., Jr. Association between preoperative 25-hydroxyvitamin D level and hospital-acquired infections following Roux-en-Y gastric bypass surgery. JAMA Surg. 2014, 149, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Quraishi, S.A.; De Pascale, G.; Needleman, J.S.; Nakazawa, H.; Kaneki, M.; Bajwa, E.K.; Camargo, C.A., Jr.; Bhan, I. Effect of Cholecalciferol Supplementation on Vitamin D Status and Cathelicidin Levels in Sepsis: A Randomized, Placebo-Controlled Trial. Crit. Care Med. 2015, 43, 1928–1937. [Google Scholar] [CrossRef] [PubMed]
- Borsche, L.; Glauner, B.; von Mendel, J. COVID-19 Mortality Risk Correlates Inversely with Vitamin D3 Status, and a Mortality Rate Close to Zero Could Theoretically Be Achieved at 50 ng/mL 25(OH)D3: Results of a Systematic Review and Meta-Analysis. Nutrients 2021, 13, 3596. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, N.K.; Shrestha, P.; Burke, P.C.; Nowacki, A.S.; Terpeluk, P.; Gordon, S.M. Coronavirus Disease 2019 Vaccine Boosting in Previously Infected or Vaccinated Individuals. Clin. Infect. Dis. 2022, 75, 2169–2177. [Google Scholar] [CrossRef]
- Uraki, R.; Ito, M.; Furusawa, Y.; Yamayoshi, S.; Iwatsuki-Horimoto, K.; Adachi, E.; Saito, M.; Koga, M.; Tsutsumi, T.; Yamamoto, S.; et al. Humoral immune evasion of the omicron subvariants BQ.1.1 and XBB. Lancet Infect. Dis. 2023, 23, 30–32. [Google Scholar] [CrossRef]
- Dingemans, J.; van der Veer, B.; Gorgels, K.M.F.; Hackert, V.; den Heijer, C.D.J.; Hoebe, C.; Savelkoul, P.H.M.; van Alphen, L.B. Investigating SARS-CoV-2 breakthrough infections per variant and vaccine type. Front. Microbiol. 2022, 13, 1027271. [Google Scholar] [CrossRef]
- Wimalawansa, S.J. Controlling COVID-19 pandemic with cholecalciferol. World J. Adv. Heathc. Res. 2020, 5, 155–165. [Google Scholar] [CrossRef]
- Arabi, Y.M.M.; Fowler, R.; Hayden, F.G. Critical care management of adults with community-acquired severe respiratory viral infection. Intensive Care Med. 2020, 46, 315–328. [Google Scholar] [CrossRef]
- Wimalawansa, S.J. Effective and practical ways to overcome vitamin D deficiency. J. Family Med. Community Health 2021, 8, 1185. [Google Scholar]
- Aslam, S.; Danziger-Isakov, L.; Mehra, M.R. COVID-19 vaccination immune paresis in heart and lung transplantation. J. Heart Lung Transplant. 2021, 40, 763–766. [Google Scholar] [CrossRef] [PubMed]
- Biesalski, H.K.; Aggett, P.J.; Anton, R.; Bernstein, P.S.; Blumberg, J.; Heaney, R.P.; Henry, J.; Nolan, J.M.; Richardson, D.P.; van Ommen, B.; et al. 26th Hohenheim Consensus Conference, September 11, 2010 Scientific substantiation of health claims: Evidence-based nutrition. Nutrition 2011, 27, S1–S20. [Google Scholar] [CrossRef]
- Tsai, F.; Coyle, W.J. The microbiome and obesity: Is obesity linked to our gut flora? Curr. Gastroenterol. Rep. 2009, 11, 307–313. [Google Scholar] [CrossRef]
- Dai, Q.; Zhu, X.; Manson, J.E.; Song, Y.; Li, X.; Franke, A.A.; Costello, R.B.; Rosanoff, A.; Nian, H.; Fan, L.; et al. Magnesium status and supplementation influence vitamin D status and metabolism: Results from a randomized trial. Am. J. Clin. Nutr. 2018, 108, 1249–1258. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wimalawansa, S.J. Controlling Chronic Diseases and Acute Infections with Vitamin D Sufficiency. Nutrients 2023, 15, 3623. https://doi.org/10.3390/nu15163623
Wimalawansa SJ. Controlling Chronic Diseases and Acute Infections with Vitamin D Sufficiency. Nutrients. 2023; 15(16):3623. https://doi.org/10.3390/nu15163623
Chicago/Turabian StyleWimalawansa, Sunil J. 2023. "Controlling Chronic Diseases and Acute Infections with Vitamin D Sufficiency" Nutrients 15, no. 16: 3623. https://doi.org/10.3390/nu15163623
APA StyleWimalawansa, S. J. (2023). Controlling Chronic Diseases and Acute Infections with Vitamin D Sufficiency. Nutrients, 15(16), 3623. https://doi.org/10.3390/nu15163623