
Immunomodulation of B Lymphocytes by Prebiotics, Probiotics and Synbiotics: Application in Pathologies
Abstract
:1. Introduction
1.1. Probiotics, Prebiotics and Synbiotics: Definitions and Effects
1.2. B Lymphocytes: Ontogeny, Function and Localization
- -
- “prebiotics and B cells”: we found 18 relevant publications in a health context and 14 relevant publications in pathological context.
- -
- “probiotics and B cells”: we found 10 relevant publications in a health context and 10 relevant publications in pathological context.
- -
- “synbiotics and B cells”: we found 3 relevant publications in a health context and 2 relevant publications in pathological context.
2. Immunomodulation of B Lymphocytes by Prebiotics, Probiotics and Synbiotics Supplementation in Healthy Individuals
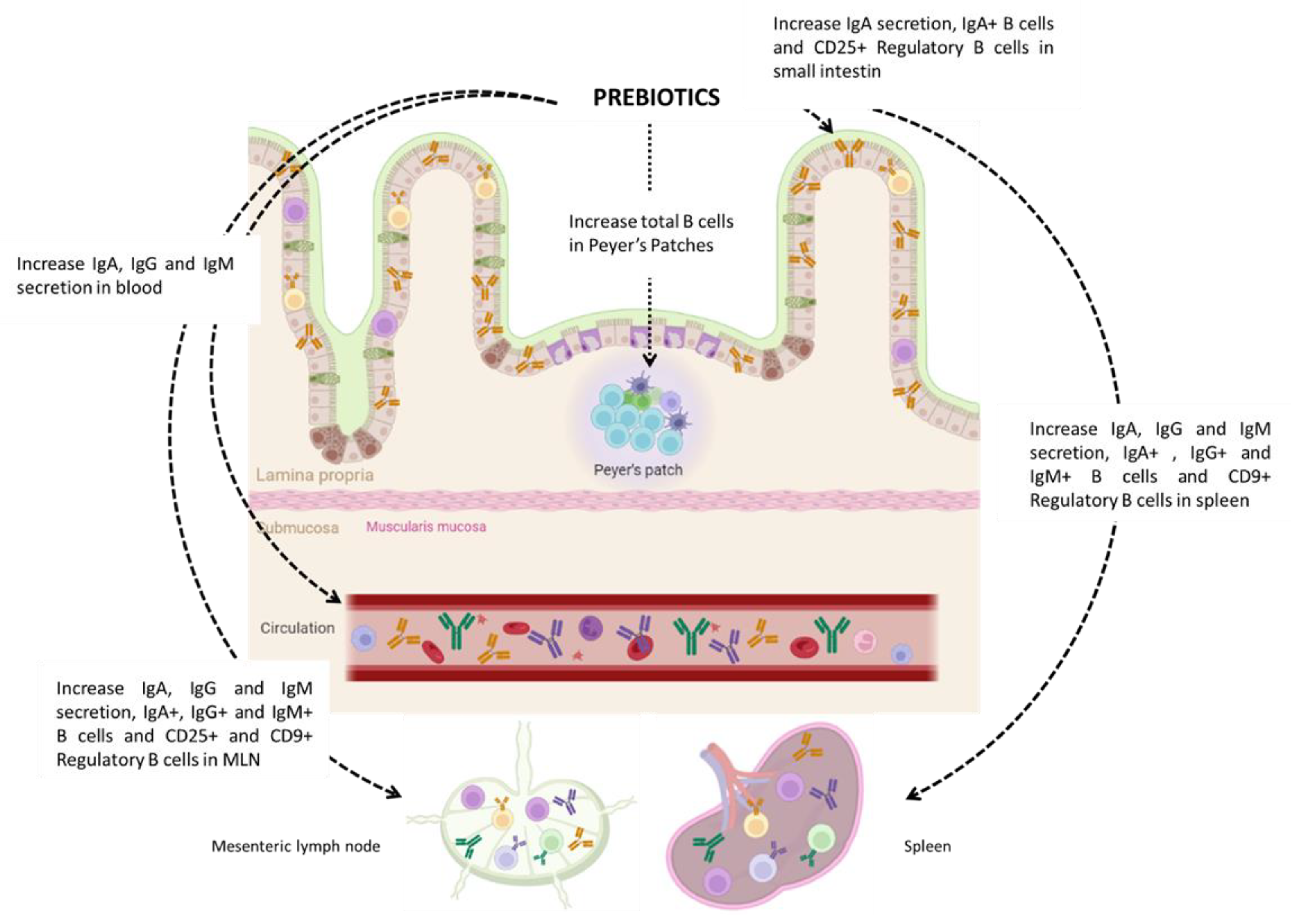
2.1. Effects of Prebiotic Supplementation on B-Cell Immunomodulation (Table 1)
2.1.1. Prebiotic Supplementation during Gestation
2.1.2. Prebiotic Supplementation during Lactation
2.1.3. Prebiotic Supplementation during Adulthood
- -
- Increase the expression of genes associated with B cells in utero.
- -
- Increase Breg frequency in the gestational tissues and in the fetus, leading to the establishment of tolerogenic immune imprinting in the child.
- -
- Promote B cell activation in neonates.
- -
- Foster the humoral response.

{kind=link}
{kind=link}
{kind=link}
| Period of Exposure | Models | Type of Prebiotics | Results | Refences |
|---|---|---|---|---|
| Gestation | Preclinical study Mice | GOS/Inulin (ratio 9/1) during 4 weeks | Increase CD9+, CD25+ Breg in dams uterus and placenta and in fetal bone marrow (B CD9+) and intestine (B CD25+) | Brosseau et al. [16] |
| Increase CD9+, CD25+ Breg in dams and CD9+ in pups at 7 weeks of age | Selle et al. [17] | |||
| Lactation neonatal period | Core-fucosylated oligosaccharides during 3 weeks | Fut8+/+ increase total and activated B cell in spleen and thymus | Li et al. [20] | |
| FOS (5% of diet) during 5 weeks | Increase total IgA in intestine and increase B cell percentage in PPs | Nakamura et al. [24] | ||
| Preclinical study Dogs | scFOS (1% of diet) during 7 weeks | Increase IgM but not IgG or IgA in colostrum and milk | Adogony et al. [21] | |
| Preclinical study Neonatal piglets | scGOS (0.8% of diet) during 3 weeks | Increase IgA concentration in saliva | Alizadeh et al. [22] | |
| Preclinical study Ckicken eggs and chickens | RFO (3 doses: 1.5; 3.0; and 4.5 mg) 1 injection in eggs at day 12 | Increase B cell marker ChB6 in small intestine | Berrocoso et al. [18] | |
| Zinc Bacitracin (ZnB, 0.05 g/kg), FOS (5 g/kg) and MOS (5 g/kg) during 3 weeks | ZnB, or MOS, or FOS: no effect on B cell percentage or Ig+ B cells FOS alone: Increase IgM and IgG concentration in plasma | Janardhana et al. [23] | ||
| Clinical trial Infants | scGOS/lcFOS (6 g/L) during the first 6 months of life | No effect on total IgE, IgA, IgM, or IgG concentration but decrease CD23+ B cells | Raes et al. [25] | |
| Adulthood | Preclinical study Dogs | Beet pulp, FOS and gum arabic (8.7 g/kg) during 2 weeks | Decrease B cell proportion in peripheral blood and decrease mitogen responses involving B cell function in GALT | Field et al. [27] |
| Chicory (1% of diet) or Inulin (0–3% of diet) during 4 weeks | No effect on IgA, IgM, or IgG concentration in the serum | Grieshop et al. [28] Verlinden et al. [29] | ||
| FOS (1 g/kg) or MOS (1 g/kg) or FOS + MOS (1 g/kg) during 2 weeks | Increase IgA concentration in small intestine | Swanson et al. [30] | ||
| Preclinical study Rats | Cellulose and lactilose (5% of diet) during 3 weeks | Increase IgA+ B cells and IgA secretion in GALT | Kudoh et al. [31] | |
| Cellulose, Konjak manna, pectin and chitosan (5 g/100 g of diet) during 2 weeks | Increase IgA, IgM, and IgG concentration in serum, increase IgA+, IgM+, and IgG+ B cells and decrease IgE concentration in MLN and spleen | Lim et al. [32] | ||
| Glucomannan (5% of the diet) during 3 weeks; low or high-fiber diet during 24 weeks | Increase IgA, IgM, and IgG concentration in MLN, spleen and serum | Yamada et al. [33] Zusman et al. [34] | ||
| Raftilose (100 g/kg) during 4 weeks | Increase IgA concentration in caecum | Roller et al. [35] | ||
| Preclinical study Mice | FOS (2.5; 3; 5; 7.5; 10% of diet) during 2 weeks | No effect on IgA, IgM, or IgG concentration, on B cells and nor IL-10 secretion | Delgado et al. [36] | |
| Increase IgA secretion in dose-dependant way | Hosono et al. [42] | |||
| Increase B cells in PPs what ever the dose | Manhart et al. [40] | |||
| Cellulose, FOS, Inulin (100 g/kg) during 6 weeks | Increase fecal IgA secretion | Buddington et al. [37] | ||
| LBP (0.1 mL/10 g of body weight) during 2 weeks | Increase IgA concentration in the colon | Wei Zhu et al. [38] | ||
| Clinical trial Adults | XOS (8 g/day) during 3 weeks | No effect on blood B cells frequency but decrease IL-10 production | Childs et al. [41] |
2.2. Effects of Probiotic Supplementation on B Cells Immunomodulation (Table 2)
2.2.1. Lactobacillus rhamnosus LGG
2.2.2. Lactobacillus acidophilus
2.2.3. Bacillus polyfermenticus (Bispan)
2.2.4. Limosilactobacillus reuteri
2.2.5. Tetragenococcus halophilus
2.2.6. Bifidobacteria
2.2.7. Mixture of Probiotics
- -
- Prebiotic supplementation can increase the IgA and IgG humoral response.
- -
- The cellular response and, in particular, regulatory B cells, have not been observed.
2.3. Effects of Synbiotic Supplementation on B Cells Immunomodulation (Table 2)
- -
- Because of the low number of studies in the literature, we cannot draw a conclusion on the effect of synbiotics on the B cell response (Table 2).
- -
- More studies are needed to obtain more information about synbiotics’ mechanisms on the immune B response.
| Type of Supplementation | Models | Type of Probiotics or Synbiotics | Results | Refences |
|---|---|---|---|---|
| Probiotics | Preclinical study mice | LGG (107 CFU/10 μL) during 2 weeks | Increase CD40, CD80 and MHC-II expression on B cells and IgG and IgM but decrease IgA secretion | Shi et al. [43] |
| L. reuteri (108 CFU/100 µL) during 2 weeks | Increase in PP Pre-GC and GC-like B cells, B220+ B cells, expansion of TGFb+ B cells and IgA germline transcript | Liu et al. [42] | ||
| T. halophilus (1% of the diet) during 2 weeks | Increase genes of B cell activation, IL-22 and IL-10 induction and IgA and IgG production in B cells, IgA level in feces and serum but no effect on IgG and IgM levels in serum | Kumasawa et al. [51] | ||
| Bifidobacteria (108 CFU/0.5 mL) during 6 weeks | Increase total IgG | El Hadad et al. [53] | ||
| Preclinical study calves | Normosil (30 mL/animal), Vetosporin Zh (20 mL/animal), Gumi-malysh (30 mL/animal) during 3 weeks | Increase number of B cell and IgA and IgG levels in blood and decrease IgM level in serum | Andreeva et al. [55] | |
| Preclinical study Piglet | LGG (1010 CFU/5 mL) during 2 weeks | Decrease B cells in lamina propria but increase mature B cells in PPs, IgA leveles in feces and lamina propria | Jin et al. [44] | |
| LGG, L. acidophilus and B. longum | Increase IgA+ and IgM+ B cells | Kalita et al. [56] | ||
| Clinical trial Adults | L. acidophilus, L. lactis, L. casei, B. longum, B. bifidum and B. infantis (3010 CFU) during 12 weeks | No effect on salivary IgA nor total B cells | Ibrahim et al. [54] | |
| LGG (450 Bn LGG/ 50 mL) 2 h after ingestion | Increase genes of B cell activation (CD22, CD19, CD21, CD79a(IGa) CD79B(IGb), FCGR2B) | Bornholdt et al. [45] | ||
| Bacillus polyfermenticus (Bispan) (108 CFU/day) during 8 weeks | No effect on IgA, IgG and IgM levels in serum but increase total IgG | Kim et al. [48] | ||
| Clinical trial Infants | L. acidophilus (309 CFU/1–2 mL) during the first 6 months of life | No effect on total B cells | Taylor et al. [47] | |
| Synbiotics | Preclinical study Chickens | Inulin (1.76 mg), Bi2 tos (0.528 mg), Lactobacillus (1000 CFU) During 3 weeks | Increase proportion of Bu1 cells in ceacal tonsil but no effect on Bu1+ cell density in cortex and medulla | Madej et al. [57] |
| Preclinical Piglets | FOS Raftilose P95 (3 g/day), Lactobacillus (109 CFU/g) during 3 weeks | Increase total B cells in blood | Herich et al. [58] | |
| Clinical trial Adults | XOS (8 g/day), B. lactis (109 CFU/day) during 3 weeks | Decrease total B cells but no effect on salivary and fecal IgA concentration | Childs et al. [41] |
3. Immunomodulation of B Lymphocytes by Prebiotics, Probiotics and Synbiotics Supplementation in Pathological Contexts
3.1. Vaccination
3.1.1. Vaccination with Prebiotics
3.1.2. Vaccination with Probiotics
- -
- In a murine model, prebiotics supplementation improved vaccine-specific humoral immunity linked to proper B cell memory development, but this is not currently conclusive in humans.
- -
- Both in pre-clinical and clinical studies, probiotic intake activated B cell response and improved vaccine-specific humoral immunity.
3.2. Pathological Context
3.2.1. Colorectal Cancer
3.2.2. Diabetes
3.2.3. Allergies
Prevention of Allergies
Treatment of Allergies
- -
- B cell response associated with prebiotic intake in the context of allergy was more investigated in prevention than treatment and has shown promising results, especially in the modulation of the humoral response.
- -
- Once again, the humoral response mainly described with a decrease in IgE and the B cellular response needs to be further investigated.
- -
- The use of probiotics has shown major clinical interest in the context of SIT to induce tolerance associated with an increase in Breg rate.
3.2.4. Endotoxemia
3.2.5. Virus Infection
COVID-19
Rotavirus
3.2.6. Autoimmunity
3.2.7. Intestinal Inflammation
3.2.8. Graft-Versus-Host Disease
- -
- However, for other pathologies, more studies are necessary to conclude on the beneficial effect of prebiotics or probiotics.
- -
- Furthermore, several studies (both preclinical and clinical) are needed to understand B-mediated mechanisms and evaluate their effects, not only investigating B cell Ig secretion but also cellular immune response (cytokine secretion, Breg differentiation, etc.).
| Pathology | Models | Type of Supplementation | Results | References |
|---|---|---|---|---|
| Vaccination | Preclinical study—Mice | Prebiotics scGOS/lcFOS/2′-FL during 6 weeks—Influenza infection | Increase vaccine specific response with higher IgG1 and IgG2a levels, higher activated B cells (CD27+ and CD138+ B cells) | Xiao et al. [62]; van den Elsen et al. [61] |
| 2′-FL (0.25–5% w/w) during 6 weeks—Influenza infection | Increase vaccine specific response wih higher specific IgG1 and IgG2a in a dose-dependent manner and higher CD27 expression on splenic B-cells | Xiao et al. [63] | ||
| FOS (5% of diet) during 8 weeks—Salmonella infection | Increase vaccine specific IgA but no effect on total IgA and IgG | Benyacoub et al. [64] | ||
| Preclinical study—Piglets | Probiotics LGG and B. animalis lactis (105 CFU) during 5 weeks—Rotavirus infection | Increase vaccine specific response with higher specific IgA and IgA+ B cells and less specific IgG | Kandasamy et al. [70] | |
| LGG and B. lactis (105 CFU) 2 doses before vaccination—Rotavirus infection | Increase vaccine specific response with higher specific IgA in ileum and duodenum | Chattha et al. [71] | ||
| Clinical trial—Adults | Prebiotics FOS (6 g/day) during 2.5 months—Influenza and Pneumococcal infections | Increase vaccine specific response with higher specific antibody titers | Langkamp-Henken et al. [65]; Bunout et al. [66] | |
| Clinical trial—Infants | Prebiotics scGOS, lcFOS and pectin derived acidic alogosaccharides (8 g/L) during 12 months—Hepatitis B, Tetanus and Poliomyelitis | No effect on the vaccine specific response against anti-hepatitis B virus, on anti-tetanus, nor the poliomyelitis | Salvini et al. [67]; Stam et al. [68]; van den Berg et al. [69] | |
| Probiotics LGG (30 mL) during the first 5 months of age—Rotavirus infection | Increase IgM sASC against rotavirus but no effect on IgA IgM, and IgG sASC response | Isolauri et al. [72] | ||
| Virus infection | Preclinical study—Piglets | Probiotics LAB (103, 104, 105, 106 CFU) 1 dose at at 3, 5, 7, 9, 11 days of age—Rotavirus infection | No effect on specific IgA, IgG and IgM in serum but increase the developpment of B cell in gut | Zhang et al. [99] |
| Probiotics EcN (105 CFU) during 5 weeks—Rotavirus infection | Increase total IgA and IgA+ B cells in small intestine but decrease specific IgA and IgG antibody titers | Kandasami et al. [70] | ||
| Preclinical study—Mice | Prebiotics scGOS/lcFOS (0.8 g/100 g of body weight) during 2 weeks—Rotavirus infection | Decrease IgG1, IgG2a/b/c and total IgA | Azagra-Boronat et al. [97] | |
| Probiotics B. bifidum (7.58 CFU/mL) and B. infantis (7.58 CFU/mL) during 6 weeks—Rotavirus infection | Increase specific IgA response but decrease total and IgA+, IgG+ B cells | Qiao et al. [100] | ||
| Preclinical study—Rats | B. breve (458 CFU/100 g of body weight), scGOS/lcFOS (0.8 g/100 g of body weight) during 3 weeks—Rotavirus infection | Probiotics, prebiotics and synbiotics decrease specific IgA, IgG and IgM in the serum but prebiotics increase specific IgA, IgG and IgM in intestinal wash | Rigo-Adrover et al. [101] Rigo-Adrover et al. [102] | |
| Synbiotics scGOS/lcFOS (0.8 g/100 g of body weight) and Lactobifidus (0.92 g/100 g body weight) during 2 weeks—Rotavirus infection | Decrease the rate of specific IgA, IgG1, IgG2a in the serum | Morales-Ferré et al. [98] | ||
| Clinical trial—Adults | Synbiotics Bifidobacterium, Lactobacillus, Enterrococcus and Bacillus (0.5 g/capsule) during 2 weeks—COVID-19 infection | Increase total B lymphocytes | Li et al. [96] | |
| Allergy | Preclinical study—Mice | Prevention Prebiotics 4% GOS/Inulin during 5 weeks—Food allergy | Increase CD9+ and CD25+ Breg | Selle et al. [17] |
| Prevention Prebiotics scGOS/lcFOS (3% of the diet) during 2 weeks—Allergic asthma | No effect on IgE, IgG1, IgG2a and on the immune cell count | Hogenkamp et al. [77] | ||
| Prevention Probiotics B. bifidum, L. casei, E. coli (0.2% of the diet) during 8 weeks—Food allergy | Decrease specific and total IgE, specific IgA and IgG1 | Kim et al. [79] | ||
| Prevention Synbiotics B. breve, GOS and FOS (2% of the diet) during 8 weeks—Food allergy | Increase specific IgA and IgG2a but no effect on specific IgE and IgG1 | Schouten et al. [81] | ||
| Treatment Prebiotics 2′-FL and 6′SL (1 mg/200 µL) during 2 weeks—Food allergy | Increase specific IgG2a but no effect on specific and total IgE nor specific IgG1 | Castillo-Courtade et al. [82] | ||
| Treatment Prebiotics GOS and FOS (10 mg/kg) or Probiotics L. acidophilus (7.5 billion CFU), LGG (8.75 billion CFU), and B. lactis (8.75 billion CFU) during 1 week—Allergic asthma | Decrease specific and total IgE and IgG1 | Whu et al. [85] | ||
| Treatments Probiotcis heat-killed LcS (0.05% of the diet) for 2 injections; B. infantis (5010 CFU/mL) during 2 weeks; C. butyricum (50 mg/mL) during 1 week—Allergic asthma, Food allergy | Decrease specific IgE and IgG1 and total IgE | Matsuzaki et al. [84]; Liu et al. [86]; Zeng et al. [87] | ||
| Treatment Synbiotics scGOS, lcFOS (1% of the diet), and B. breve (209 CFU/g) during 3 weeks—Food allergy | No effect on specific IgE | van Esch et al. [91] | ||
| AIT Treatment Probiotics C. butyricum and LGG (109 CFU/500 µL) and SIT (OVA) during 2 weeks—Food allergy | Increase the frequency of IL-10-producing OVA-specific B cell | Shi et al. [93] | ||
| Clinical trial—Infants | Prevention Prebiotics GOS and FOS (8 g/L) during 6 months—Food allergy | Decrease total IgE, IgG1, IgG2 and IgG3 in serum | van Hoffen et al. [78] | |
| Prevention Probiotics LGG (509 CFU), B. breve (108 CFU) and P. shermanii (209 CFU) during 1 month before delivery to the mother and the first 6 months of age—Allergies | Increase total IgA, IgE | Marschan et al. [80] | ||
| Treatment Probiotics L. casei and B. lactis during 12 months; Probiotics LGG (349 CFU) during 3 months—Food allergy and atopic dermatitis | No effect on mature B cell; decrease IgA and IgM+ B cells but increase memory B cells | Hol et al. [88]; Nermes et al. [90] | ||
| Treatment Synbiotics B. breve (139 CFU/g), scGOS, and lcFOS (0.8 g/100 mL) during 3 months—Atopic dermatitis | No effect on IgG1/4 concentration | van der Aa et al. [92] | ||
| Clinical trial—Adults | Treatment Probiotics L. paracasei (398 CFU/g), L. acidophilus (294 CFU/g) and B. lactis (594 CFU/g) during 2 months—Atopic dermatitis | No effect on total B cell nor IgE concentration | Roessler et al. [89] | |
| AIT Treatment Probiotics C. butyricum and SIT (Dermatophagoides pteronyssinus (Der p) during 12 months—Allergic rhinitis | Increase frequency of Breg and Derp1 specific B10 cells | Xu et al. [94] | ||
| Diabetes | Preclinical study—Rats | Prebiotics Inulin (4.8% w/w) during 2 weeks | Increase proportion on B cells in PP and IgA+ B cells in jejunum | Stillie et al. [75] |
| Endotoxemia | Preclinical study—Mice | Prebiotics FOS (10% of the diet) during 2 weeks | Increase B lymphocytes | Manhart et al. [95] |
| Autoimmunity | Probiotics B. fragilis (508 CFU/mouse) during 4 weeks | Increase CD1d expression on B cell and decrease CD86 | Li et al. [103] | |
| Intestinal inflammation | Probiotics Limosilactobacillus reuteri (108 CFU/100 µL) during 2 weeks | Increase in PP Pre-GC and GC-like B cells, B220+ B cells, expansion of TGFb+ B cells and IgA germline transcript | Liu et al. [42] | |
| Graft—versus—host—disease | Probiotics L. lactis during 5 days | Increase IL-10 B cells with higher level of CD86 and MHC-II and increase transitionnal B cells | Mercadante et al. [104] |
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Davani-Davari, D.; Negahdaripour, M.; Karimzadeh, I.; Seifan, M.; Mohkam, M.; Masoumi, S.J.; Berenjian, A.; Ghasemi, Y. Prebiotics: Definition, Types, Sources, Mechanisms, and Clinical Applications. Foods 2019, 8, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rastall, R.A.; Maitin, V. Prebiotics and Synbiotics: Towards the next Generation. Curr. Opin. Biotechnol. 2002, 13, 490–496. [Google Scholar] [CrossRef]
- Pujari, R.; Banerjee, G. Impact of Prebiotics on Immune Response: From the Bench to the Clinic. Immunol. Cell Biol. 2021, 99, 255–273. [Google Scholar] [CrossRef] [PubMed]
- Bermudez-Brito, M.; Plaza-Díaz, J.; Muñoz-Quezada, S.; Gómez-Llorente, C.; Gil, A. Probiotic Mechanisms of Action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef]
- Brosseau, C.; Selle, A.; Palmer, D.J.; Prescott, S.L.; Barbarot, S.; Bodinier, M. Prebiotics: Mechanisms and Preventive Effects in Allergy. Nutrients 2019, 11, 1841. [Google Scholar] [CrossRef] [Green Version]
- Schley, P.D.; Field, C.J. The Immune-Enhancing Effects of Dietary Fibres and Prebiotics. Br. J. Nutr. 2002, 87 (Suppl. S2), S221–S230. [Google Scholar] [CrossRef] [PubMed]
- LeBien, T.W. Fates of Human B-Cell Precursors. Blood 2000, 96, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Bemark, M. Translating Transitions—How to Decipher Peripheral Human B Cell Development. J. Biomed. Res. 2015, 29, 264–284. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, H.W.; Cavacini, L. Structure and Function of Immunoglobulins. J. Allergy Clin. Immunol. 2010, 125, S41–S52. [Google Scholar] [CrossRef] [Green Version]
- Keyt, B.A.; Baliga, R.; Sinclair, A.M.; Carroll, S.F.; Peterson, M.S. Structure, Function, and Therapeutic Use of IgM Antibodies. Antibodies 2020, 9, 53. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Jialal, I. Biochemistry, Immunoglobulin A. In StatPearls; StatPearls Publishing: Treasure, FL, USA, 2022. [Google Scholar]
- Sutton, B.J.; Davies, A.M.; Bax, H.J.; Karagiannis, S.N. IgE Antibodies: From Structure to Function and Clinical Translation. Antibodies 2019, 8, 19. [Google Scholar] [CrossRef] [Green Version]
- Rosser, E.C.; Mauri, C. The Emerging Field of Regulatory B Cell Immunometabolism. Cell Metab. 2021, 33, 1088–1097. [Google Scholar] [CrossRef] [PubMed]
- Vossenkämper, A.; Blair, P.A.; Safinia, N.; Fraser, L.D.; Das, L.; Sanders, T.J.; Stagg, A.J.; Sanderson, J.D.; Taylor, K.; Chang, F.; et al. A Role for Gut-Associated Lymphoid Tissue in Shaping the Human B Cell Repertoire. J. Exp. Med. 2013, 210, 1665–1674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, J.E.; Sinkora, M. The Enigma of the Lower Gut-Associated Lymphoid Tissue (GALT). J. Leukoc. Biol. 2013, 94, 259–270. [Google Scholar] [CrossRef]
- Brosseau, C.; Selle, A.; Duval, A.; Misme-Aucouturier, B.; Chesneau, M.; Brouard, S.; Cherbuy, C.; Cariou, V.; Bouchaud, G.; Mincham, K.T.; et al. Prebiotic Supplementation During Pregnancy Modifies the Gut Microbiota and Increases Metabolites in Amniotic Fluid, Driving a Tolerogenic Environment In Utero. Front. Immunol. 2021, 12, 712614. [Google Scholar] [CrossRef] [PubMed]
- Selle, A.; Brosseau, C.; Dijk, W.; Duval, A.; Bouchaud, G.; Rousseaux, A.; Bruneau, A.; Cherbuy, C.; Mariadassou, M.; Cariou, V.; et al. Prebiotic Supplementation During Gestation Induces a Tolerogenic Environment and a Protective Microbiota in Offspring Mitigating Food Allergy. Front. Immunol. 2021, 12, 745535. [Google Scholar] [CrossRef]
- Berrocoso, J.D.; Kida, R.; Singh, A.K.; Kim, Y.S.; Jha, R. Effect of in Ovo Injection of Raffinose on Growth Performance and Gut Health Parameters of Broiler Chicken. Poult. Sci. 2017, 96, 1573–1580. [Google Scholar] [CrossRef]
- Rousseaux, A.; Brosseau, C.; Le Gall, S.; Piloquet, H.; Barbarot, S.; Bodinier, M. Human Milk Oligosaccharides: Their Effects on the Host and Their Potential as Therapeutic Agents. Front. Immunol. 2021, 12, 680911. [Google Scholar] [CrossRef]
- Li, M.; Bai, Y.; Zhou, J.; Huang, W.; Yan, J.; Tao, J.; Fan, Q.; Liu, Y.; Mei, D.; Yan, Q.; et al. Core Fucosylation of Maternal Milk N-Glycan Evokes B Cell Activation by Selectively Promoting the l-Fucose Metabolism of Gut Bifidobacterium spp. and Lactobacillus spp. mBio 2019, 10, e00128-19. [Google Scholar] [CrossRef] [Green Version]
- Adogony, V.; Respondek, F.; Biourge, V.; Rudeaux, F.; Delaval, J.; Bind, J.-L.; Salmon, H. Effects of Dietary ScFOS on Immunoglobulins in Colostrums and Milk of Bitches. J. Anim. Physiol. Anim. Nutr. 2007, 91, 169–174. [Google Scholar] [CrossRef]
- Alizadeh, A.; Akbari, P.; Difilippo, E.; Schols, H.A.; Ulfman, L.H.; Schoterman, M.H.C.; Garssen, J.; Fink-Gremmels, J.; Braber, S. The Piglet as a Model for Studying Dietary Components in Infant Diets: Effects of Galacto-Oligosaccharides on Intestinal Functions. Br. J. Nutr. 2016, 115, 605–618. [Google Scholar] [CrossRef] [Green Version]
- Janardhana, V.; Broadway, M.M.; Bruce, M.P.; Lowenthal, J.W.; Geier, M.S.; Hughes, R.J.; Bean, A.G.D. Prebiotics Modulate Immune Responses in the Gut-Associated Lymphoid Tissue of Chickens. J. Nutr. 2009, 139, 1404–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, Y.; Nosaka, S.; Suzuki, M.; Nagafuchi, S.; Takahashi, T.; Yajima, T.; Takenouchi-Ohkubo, N.; Iwase, T.; Moro, I. Dietary Fructooligosaccharides Up-Regulate Immunoglobulin A Response and Polymeric Immunoglobulin Receptor Expression in Intestines of Infant Mice. Clin. Exp. Immunol. 2004, 137, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Raes, M.; Scholtens, P.A.M.J.; Alliet, P.; Hensen, K.; Jongen, H.; Boehm, G.; Vandenplas, Y.; Rummens, J.L. Exploration of Basal Immune Parameters in Healthy Infants Receiving an Infant Milk Formula Supplemented with Prebiotics. Pediatr. Allergy Immunol. 2010, 21, e377–e385. [Google Scholar] [CrossRef] [PubMed]
- Hjelm, F.; Carlsson, F.; Getahun, A.; Heyman, B. Antibody-Mediated Regulation of the Immune Response. Scand. J. Immunol. 2006, 64, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Field, C.J.; McBurney, M.I.; Massimino, S.; Hayek, M.G.; Sunvold, G.D. The Fermentable Fiber Content of the Diet Alters the Function and Composition of Canine Gut Associated Lymphoid Tissue. Vet. Immunol. Immunopathol. 1999, 72, 325–341. [Google Scholar] [CrossRef]
- Grieshop, C.M.; Flickinger, E.A.; Bruce, K.J.; Patil, A.R.; Czarnecki-Maulden, G.L.; Fahey, G.C. Gastrointestinal and Immunological Responses of Senior Dogs to Chicory and Mannan-Oligosaccharides. Arch. Anim. Nutr. 2004, 58, 483–493. [Google Scholar] [CrossRef]
- Verlinden, A.; Hesta, M.; Hermans, J.M.; Janssens, G.P.J. The Effects of Inulin Supplementation of Diets with or without Hydrolysed Protein Sources on Digestibility, Faecal Characteristics, Haematology and Immunoglobulins in Dogs. Br. J. Nutr. 2006, 96, 936–944. [Google Scholar] [CrossRef] [Green Version]
- Swanson, K.S.; Grieshop, C.M.; Flickinger, E.A.; Bauer, L.L.; Healy, H.-P.; Dawson, K.A.; Merchen, N.R.; Fahey, G.C. Supplemental Fructooligosaccharides and Mannanoligosaccharides Influence Immune Function, Ileal and Total Tract Nutrient Digestibilities, Microbial Populations and Concentrations of Protein Catabolites in the Large Bowel of Dogs. J. Nutr. 2002, 132, 980–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudoh, K.; Shimizu, J.; Wada, M.; Takita, T.; Kanke, Y.; Innami, S. Effect of Indigestible Saccharides on B Lymphocyte Response of Intestinal Mucosa and Cecal Fermentation in Rats. J. Nutr. Sci. Vitaminol. 1998, 44, 103–112. [Google Scholar] [CrossRef]
- Lim, B.O.; Yamada, K.; Nonaka, M.; Kuramoto, Y.; Hung, P.; Sugano, M. Dietary Fibers Modulate Indices of Intestinal Immune Function in Rats. J. Nutr. 1997, 127, 663–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, K.; Tokunaga, Y.; Ikeda, A.; Ohkura, K.; Mamiya, S.; Kaku, S.; Sugano, M.; Tachibana, H. Dietary Effect of Guar Gum and Its Partially Hydrolyzed Product on the Lipid Metabolism and Immune Function of Sprague Dawley Rats. Biosci. Biotechnol. Biochem. 1999, 63, 2163–2167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zusman, I.; Gurevich, P.; Benhur, H.; Berman, V.; Sandler, B.; Tendler, Y.; Madar, Z. The Immune Response of Rat Spleen to Dietary Fibers and to Low Doses of Carcinogen: Morphometric and Immunohistochemical Studies. Oncol. Rep. 1998, 5, 1577–1581. [Google Scholar] [CrossRef]
- Roller, M.; Rechkemmer, G.; Watzl, B. Prebiotic Inulin Enriched with Oligofructose in Combination with the Probiotics Lactobacillus Rhamnosus and Bifidobacterium Lactis Modulates Intestinal Immune Functions in Rats. J. Nutr. 2004, 134, 153–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado, G.T.C.; Thomé, R.; Gabriel, D.L.; Tamashiro, W.M.S.C.; Pastore, G.M. Yacon (Smallanthus Sonchifolius)-Derived Fructooligosaccharides Improves the Immune Parameters in the Mouse. Nutr. Res. 2012, 32, 884–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buddington, K.K.; Donahoo, J.B.; Buddington, R.K. Dietary Oligofructose and Inulin Protect Mice from Enteric and Systemic Pathogens and Tumor Inducers. J. Nutr. 2002, 132, 472–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, W.; Zhou, S.; Liu, J.; McLean, R.J.C.; Chu, W. Prebiotic, Immuno-Stimulating and Gut Microbiota-Modulating Effects of Lycium Barbarum Polysaccharide. Biomed. Pharmacother. Biomed. Pharmacother. 2020, 121, 109591. [Google Scholar] [CrossRef]
- Hosono, A.; Ozawa, A.; Kato, R.; Ohnishi, Y.; Nakanishi, Y.; Kimura, T.; Nakamura, R. Dietary Fructooligosaccharides Induce Immunoregulation of Intestinal IgA Secretion by Murine Peyer’s Patch Cells. Biosci. Biotechnol. Biochem. 2003, 67, 758–764. [Google Scholar] [CrossRef] [Green Version]
- Manhart, N.; Spittler, A.; Bergmeister, H.; Mittlböck, M.; Roth, E. Influence of Fructooligosaccharides on Peyer’s Patch Lymphocyte Numbers in Healthy and Endotoxemic Mice. Nutrion 2003, 19, 657–660. [Google Scholar] [CrossRef]
- Childs, C.E.; Röytiö, H.; Alhoniemi, E.; Fekete, A.A.; Forssten, S.D.; Hudjec, N.; Lim, Y.N.; Steger, C.J.; Yaqoob, P.; Tuohy, K.M.; et al. Xylo-Oligosaccharides Alone or in Synbiotic Combination with Bifidobacterium Animalis Subsp. Lactis Induce Bifidogenesis and Modulate Markers of Immune Function in Healthy Adults: A Double-Blind, Placebo-Controlled, Randomised, Factorial Cross-over Study. Br. J. Nutr. 2014, 111, 1945–1956. [Google Scholar] [CrossRef]
- Liu, H.-Y.; Giraud, A.; Seignez, C.; Ahl, D.; Guo, F.; Sedin, J.; Walden, T.; Oh, J.-H.; van Pijkeren, J.P.; Holm, L.; et al. Distinct B Cell Subsets in Peyer’s Patches Convey Probiotic Effects by Limosilactobacillus Reuteri. Microbiome 2021, 9, 198. [Google Scholar] [CrossRef]
- Shi, C.-W.; Zeng, Y.; Yang, G.-L.; Jiang, Y.-L.; Yang, W.-T.; Chen, Y.-Q.; Wang, J.-Y.; Wang, J.-Z.; Kang, Y.-H.; Huang, H.-B.; et al. Effect of Lactobacillus Rhamnosus on the Development of B Cells in Gut-Associated Lymphoid Tissue of BALB/c Mice. J. Cell. Mol. Med. 2020, 24, 8883–8886. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.-B.; Cao, X.; Shi, C.-W.; Feng, B.; Huang, H.-B.; Jiang, Y.-L.; Wang, J.-Z.; Yang, G.-L.; Yang, W.-T.; Wang, C.-F. Lactobacillus Rhamnosus GG Promotes Early B Lineage Development and IgA Production in the Lamina Propria in Piglets. J. Immunol. 2021, 207, 2179–2191. [Google Scholar] [CrossRef] [PubMed]
- Bornholdt, J.; Broholm, C.; Chen, Y.; Rago, A.; Sloth, S.; Hendel, J.; Melsæther, C.; Müller, C.V.; Juul Nielsen, M.; Strickertsson, J.; et al. Personalized B Cell Response to the Lactobacillus Rhamnosus GG Probiotic in Healthy Human Subjects: A Randomized Trial. Gut Microbes 2020, 12, 1854639. [Google Scholar] [CrossRef]
- Mikulic, J.; Longet, S.; Favre, L.; Benyacoub, J.; Corthesy, B. Secretory IgA in Complex with Lactobacillus Rhamnosus Potentiates Mucosal Dendritic Cell-Mediated Treg Cell Differentiation via TLR Regulatory Proteins, RALDH2 and Secretion of IL-10 and TGF-β. Cell. Mol. Immunol. 2017, 14, 546–556. [Google Scholar] [CrossRef] [Green Version]
- Taylor, A.; Hale, J.; Wiltschut, J.; Lehmann, H.; Dunstan, J.A.; Prescott, S.L. Evaluation of the Effects of Probiotic Supplementation from the Neonatal Period on Innate Immune Development in Infancy. Clin. Exp. Allergy 2006, 36, 1218–1226. [Google Scholar] [CrossRef]
- Kim, H.-S.; Park, H.; Cho, I.-Y.; Paik, H.-D.; Park, E. Dietary Supplementation of Probiotic Bacillus polyfermenticus, Bispan Strain, Modulates Natural Killer Cell and T Cell Subset Populations and Immunoglobulin G Levels in Human Subjects. J. Med. Food 2006, 9, 321–327. [Google Scholar] [CrossRef]
- Piccioni, A.; Franza, L.; Vaccaro, V.; Saviano, A.; Zanza, C.; Candelli, M.; Covino, M.; Franceschi, F.; Ojetti, V. Microbiota and Probiotics: The Role of Limosilactobacillus Reuteri in Diverticulitis. Medicina 2021, 57, 802. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Nakamoto, M.; Shuto, E.; Hata, A.; Aki, N.; Shikama, Y.; Bando, Y.; Ichihara, T.; Minamigawa, T.; Kuwamura, Y.; et al. Associations between Intake of Dietary Fermented Soy Food and Concentrations of Inflammatory Markers: A Cross-Sectional Study in Japanese Workers. J. Med. Investig. 2018, 65, 74–80. [Google Scholar] [CrossRef] [Green Version]
- Kumazawa, T.; Nishimura, A.; Asai, N.; Adachi, T. Isolation of Immune-Regulatory Tetragenococcus Halophilus from Miso. PloS ONE 2018, 13, e0208821. [Google Scholar] [CrossRef]
- Bahmani, S.; Azarpira, N.; Moazamian, E. Anti-Colon Cancer Activity of Bifidobacterium Metabolites on Colon Cancer Cell Line SW742. Turk. J. Gastroenterol. 2019, 30, 835–842. [Google Scholar] [CrossRef] [PubMed]
- El Hadad, S.; Zakareya, A.; Al-Hejin, A.; Aldahlawi, A.; Alharbi, M. Sustaining Exposure to High Concentrations of Bifidobacteria Inhibits Gene Expression of Mouse’s Mucosal Immunity. Heliyon 2019, 5, e02866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, N.S.; Ooi, F.K.; Chen, C.K.; Muhamad, A.S. Effects of Probiotics Supplementation and Circuit Training on Immune Responses among Sedentary Young Males. J. Sport. Med. Phys. Fit. 2018, 58, 1102–1109. [Google Scholar] [CrossRef] [PubMed]
- Andreeva, A.V.; Khakimova, A.Z.; Ivanov, A.I.; Nikolaeva, O.N.; Altynbekov, O.M. Immunomodulatory Effect of the Combined Use of Vetosporin Zh Probiotic and Gumi-Malysh Biologically Active Additive. Vet. World 2021, 14, 1915–1921. [Google Scholar] [CrossRef]
- Kalita, A.; Talukdar, M.; Sarma, K.; Kalita, P.C.; Barman, N.N.; Roychoudhury, P.; Kalita, G.; Choudhary, O.P.; Doley, P.J.; Debroy, S.; et al. Lymphocyte Subsets in the Small Intestine of Piglets Fed with Probiotic and Zinc: A Qualitative and Quantitative Micro-Anatomical Study. Folia Morphol. 2022, 81, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Madej, J.P.; Bednarczyk, M. Effect of in Ovo-Delivered Prebiotics and Synbiotics on the Morphology and Specific Immune Cell Composition in the Gut-Associated Lymphoid Tissue. Poult. Sci. 2016, 95, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Herich, R.; Révajová, V.; Levkut, M.; Bomba, A.; Nemcová, R.; Guba, P.; Gancarciková, S. The Effect of Lactobacillus Paracasei and Raftilose P95 Upon the Non-Specific Immune Response of Piglets. Food Agric. Immunol. 2002, 14, 171–179. [Google Scholar] [CrossRef]
- Ajetunmobi, O.M.; Whyte, B.; Chalmers, J.; Tappin, D.M.; Wolfson, L.; Fleming, M.; MacDonald, A.; Wood, R.; Stockton, D.L. Glasgow Centre for Population Health Breastfeeding Project Steering Group Breastfeeding Is Associated with Reduced Childhood Hospitalization: Evidence from a Scottish Birth Cohort (1997–2009). J. Pediatr. 2015, 166, 620–625.e4. [Google Scholar] [CrossRef] [Green Version]
- Lyons, K.E.; Ryan, C.A.; Dempsey, E.M.; Ross, R.P.; Stanton, C. Breast Milk, a Source of Beneficial Microbes and Associated Benefits for Infant Health. Nutrients 2020, 12, 1039. [Google Scholar] [CrossRef] [Green Version]
- van den Elsen, L.W.J.; Tims, S.; Jones, A.M.; Stewart, A.; Stahl, B.; Garssen, J.; Knol, J.; Forbes-Blom, E.E.; Va not Land, B. Prebiotic Oligosaccharides in Early Life Alter Gut Microbiome Development in Male Mice While Supporting Influenza Vaccination Responses. Benef. Microbes 2019, 10, 279–291. [Google Scholar] [CrossRef]
- Xiao, L.; Engen, P.A.; Leusink-Muis, T.; van Ark, I.; Stahl, B.; Overbeek, S.A.; Garssen, J.; Naqib, A.; Green, S.J.; Keshavarzian, A.; et al. The Combination of 2′-Fucosyllactose with Short-Chain Galacto-Oligosaccharides and Long-Chain Fructo-Oligosaccharides That Enhance Influenza Vaccine Responses Is Associated with Mucosal Immune Regulation in Mice. J. Nutr. 2019, 149, 856–869. [Google Scholar] [CrossRef] [Green Version]
- Xiao, L.; Leusink-Muis, T.; Kettelarij, N.; van Ark, I.; Blijenberg, B.; Hesen, N.A.; Stahl, B.; Overbeek, S.A.; Garssen, J.; Folkerts, G.; et al. Human Milk Oligosaccharide 2′-Fucosyllactose Improves Innate and Adaptive Immunity in an Influenza-Specific Murine Vaccination Model. Front. Immunol. 2018, 9, 452. [Google Scholar] [CrossRef] [Green Version]
- Benyacoub, J.; Rochat, F.; Saudan, K.-Y.; Rochat, I.; Antille, N.; Cherbut, C.; von der Weid, T.; Schiffrin, E.J.; Blum, S. Feeding a Diet Containing a Fructooligosaccharide Mix Can Enhance Salmonella Vaccine Efficacy in Mice. J. Nutr. 2008, 138, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Langkamp-Henken, B.; Wood, S.M.; Herlinger-Garcia, K.A.; Thomas, D.J.; Stechmiller, J.K.; Bender, B.S.; Gardner, E.M.; DeMichele, S.J.; Schaller, J.P.; Murasko, D.M. Nutritional Formula Improved Immune Profiles of Seniors Living in Nursing Homes. J. Am. Geriatr. Soc. 2006, 54, 1861–1870. [Google Scholar] [CrossRef]
- Bunout, D.; Hirsch, S.; Pía de la Maza, M.; Muñoz, C.; Haschke, F.; Steenhout, P.; Klassen, P.; Barrera, G.; Gattas, V.; Petermann, M. Effects of Prebiotics on the Immune Response to Vaccination in the Elderly. J. Parenter. Enter. Nutr. 2002, 26, 372–376. [Google Scholar] [CrossRef]
- Salvini, F.; Riva, E.; Salvatici, E.; Boehm, G.; Jelinek, J.; Banderali, G.; Giovannini, M. A Specific Prebiotic Mixture Added to Starting Infant Formula Has Long-Lasting Bifidogenic Effects. J. Nutr. 2011, 141, 1335–1339. [Google Scholar] [CrossRef] [Green Version]
- Stam, J.; van Stuijvenberg, M.; Garssen, J.; Knipping, K.; Sauer, P.J.J. A Mixture of Three Prebiotics Does Not Affect Vaccine Specific Antibody Responses in Healthy Term Infants in the First Year of Life. Vaccine 2011, 29, 7766–7772. [Google Scholar] [CrossRef]
- van den Berg, J.P.; Westerbeek, E.A.M.; van der Klis, F.R.M.; Berbers, G.A.M.; Lafeber, H.N.; van Elburg, R.M. Neutral and Acidic Oligosaccharides Supplementation Does Not Increase the Vaccine Antibody Response in Preterm Infants in a Randomized Clinical Trial. PLoS ONE 2013, 8, e70904. [Google Scholar] [CrossRef]
- Kandasamy, S.; Chattha, K.S.; Vlasova, A.N.; Rajashekara, G.; Saif, L.J. Lactobacilli and Bifidobacteria Enhance Mucosal B Cell Responses and Differentially Modulate Systemic Antibody Responses to an Oral Human Rotavirus Vaccine in a Neonatal Gnotobiotic Pig Disease Model. Gut Microbes 2014, 5, 639–651. [Google Scholar] [CrossRef]
- Chattha, K.S.; Vlasova, A.N.; Kandasamy, S.; Esseili, M.A.; Siegismund, C.; Rajashekara, G.; Saif, L.J. Probiotics and Colostrum/Milk Differentially Affect Neonatal Humoral Immune Responses to Oral Rotavirus Vaccine. Vaccine 2013, 31, 1916–1923. [Google Scholar] [CrossRef]
- Isolauri, E.; Joensuu, J.; Suomalainen, H.; Luomala, M.; Vesikari, T. Improved Immunogenicity of Oral D x RRV Reassortant Rotavirus Vaccine by Lactobacillus Casei GG. Vaccine 1995, 13, 310–312. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; He, Y.; Li, H.; Yu, D.; Na, L.; Sun, T.; Zhang, D.; Shi, X.; Xia, Y.; Jiang, T.; et al. Effects of Prebiotics on Immunologic Indicators and Intestinal Microbiota Structure in Perioperative Colorectal Cancer Patients. Nutrion 2019, 61, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Staff, C.; Magnusson, C.G.M.; Hojjat-Farsangi, M.; Mosolits, S.; Liljefors, M.; Frödin, J.-E.; Wahrén, B.; Mellstedt, H.; Ullenhag, G.J. Induction of IgM, IgA and IgE Antibodies in Colorectal Cancer Patients Vaccinated with a Recombinant CEA Protein. J. Clin. Immunol. 2012, 32, 855–865. [Google Scholar] [CrossRef]
- Stillie, R.; Bell, R.C.; Field, C.J. Diabetes-Prone BioBreeding Rats Do Not Have a Normal Immune Response When Weaned to a Diet Containing Fermentable Fibre. Br. J. Nutr. 2005, 93, 645–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Larsen, V.; Ierodiakonou, D.; Jarrold, K.; Cunha, S.; Chivinge, J.; Robinson, Z.; Geoghegan, N.; Ruparelia, A.; Devani, P.; Trivella, M.; et al. Diet during Pregnancy and Infancy and Risk of Allergic or Autoimmune Disease: A Systematic Review and Meta-Analysis. PLoS Med. 2018, 15, e1002507. [Google Scholar] [CrossRef] [PubMed]
- Hogenkamp, A.; Thijssen, S.; van Vlies, N.; Garssen, J. Supplementing Pregnant Mice with a Specific Mixture of Nondigestible Oligosaccharides Reduces Symptoms of Allergic Asthma in Male Offspring. J. Nutr. 2015, 145, 640–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Hoffen, E.; Ruiter, B.; Faber, J.; M’Rabet, L.; Knol, E.F.; Stahl, B.; Arslanoglu, S.; Moro, G.; Boehm, G.; Garssen, J. A Specific Mixture of Short-Chain Galacto-Oligosaccharides and Long-Chain Fructo-Oligosaccharides Induces a Beneficial Immunoglobulin Profile in Infants at High Risk for Allergy. Allergy 2009, 64, 484–487. [Google Scholar] [CrossRef]
- Kim, H.; Kwack, K.; Kim, D.-Y.; Ji, G.E. Oral Probiotic Bacterial Administration Suppressed Allergic Responses in an Ovalbumin-Induced Allergy Mouse Model. FEMS Immunol. Med. Microbiol. 2005, 45, 259–267. [Google Scholar] [CrossRef]
- Marschan, E.; Kuitunen, M.; Kukkonen, K.; Poussa, T.; Sarnesto, A.; Haahtela, T.; Korpela, R.; Savilahti, E.; Vaarala, O. Probiotics in Infancy Induce Protective Immune Profiles That Are Characteristic for Chronic Low-Grade Inflammation. Clin. Exp. Allergy 2008, 38, 611–618. [Google Scholar] [CrossRef]
- Schouten, B.; van Esch, B.C.A.M.; Hofman, G.A.; van Doorn, S.A.C.M.; Knol, J.; Nauta, A.J.; Garssen, J.; Willemsen, L.E.M.; Knippels, L.M.J. Cow Milk Allergy Symptoms Are Reduced in Mice Fed Dietary Synbiotics during Oral Sensitization with Whey. J. Nutr. 2009, 139, 1398–1403. [Google Scholar] [CrossRef] [Green Version]
- Castillo-Courtade, L.; Han, S.; Lee, S.; Mian, F.M.; Buck, R.; Forsythe, P. Attenuation of Food Allergy Symptoms Following Treatment with Human Milk Oligosaccharides in a Mouse Model. Allergy 2015, 70, 1091–1102. [Google Scholar] [CrossRef] [PubMed]
- Dargahi, N.; Johnson, J.; Donkor, O.; Vasiljevic, T.; Apostolopoulos, V. Immunomodulatory Effects of Probiotics: Can They Be Used to Treat Allergies and Autoimmune Diseases? Maturitas 2019, 119, 25–38. [Google Scholar] [CrossRef]
- Matsuzaki, T.; Chin, J. Modulating Immune Responses with Probiotic Bacteria. Immunol. Cell Biol. 2000, 78, 67–73. [Google Scholar] [CrossRef]
- Wu, Z.; Mehrabi Nasab, E.; Arora, P.; Athari, S.S. Study Effect of Probiotics and Prebiotics on Treatment of OVA-LPS-Induced of Allergic Asthma Inflammation and Pneumonia by Regulating the TLR4/NF-KB Signaling Pathway. J. Transl. Med. 2022, 20, 130. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.-Y.; Yang, Z.-Y.; Dai, W.-K.; Huang, J.-Q.; Li, Y.-H.; Zhang, J.; Qiu, C.-Z.; Wei, C.; Zhou, Q.; Sun, X.; et al. Protective Effect of Bifidobacterium Infantis CGMCC313-2 on Ovalbumin-Induced Airway Asthma and β-Lactoglobulin-Induced Intestinal Food Allergy Mouse Models. World J. Gastroenterol. 2017, 23, 2149–2158. [Google Scholar] [CrossRef]
- Zeng, X.-H.; Yang, G.; Liu, J.-Q.; Geng, X.-R.; Cheng, B.-H.; Liu, Z.-Q.; Yang, P.-C. Nasal Instillation of Probiotic Extracts Inhibits Experimental Allergic Rhinitis. Immunotherapy 2019, 11, 1315–1323. [Google Scholar] [CrossRef]
- Hol, J.; van Leer, E.H.G.; Elink Schuurman, B.E.E.; de Ruiter, L.F.; Samsom, J.N.; Hop, W.; Neijens, H.J.; de Jongste, J.C.; Nieuwenhuis, E.E.S. Cow’s Milk Allergy Modified by Elimination and Lactobacilli study group The Acquisition of Tolerance toward Cow’s Milk through Probiotic Supplementation: A Randomized, Controlled Trial. J. Allergy Clin. Immunol. 2008, 121, 1448–1454. [Google Scholar] [CrossRef]
- Roessler, A.; Friedrich, U.; Vogelsang, H.; Bauer, A.; Kaatz, M.; Hipler, U.C.; Schmidt, I.; Jahreis, G. The Immune System in Healthy Adults and Patients with Atopic Dermatitis Seems to Be Affected Differently by a Probiotic Intervention. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2008, 38, 93–102. [Google Scholar] [CrossRef]
- Nermes, M.; Kantele, J.M.; Atosuo, T.J.; Salminen, S.; Isolauri, E. Interaction of Orally Administered Lactobacillus Rhamnosus GG with Skin and Gut Microbiota and Humoral Immunity in Infants with Atopic Dermatitis. Clin. Exp. Allergy 2011, 41, 370–377. [Google Scholar] [CrossRef]
- van Esch, B.C.A.M.; Abbring, S.; Diks, M.A.P.; Dingjan, G.M.; Harthoorn, L.F.; Vos, A.P.; Garssen, J. Post-Sensitization Administration of Non-Digestible Oligosaccharides and Bifidobacterium Breve M-16V Reduces Allergic Symptoms in Mice. Immun. Inflamm. Dis. 2016, 4, 155–165. [Google Scholar] [CrossRef]
- van der Aa, L.B.; Lutter, R.; Heymans, H.S.A.; Smids, B.S.; Dekker, T.; van Aalderen, W.M.C.; Sillevis Smitt, J.H.; Knippels, L.M.J.; Garssen, J.; Nauta, A.J.; et al. No Detectable Beneficial Systemic Immunomodulatory Effects of a Specific Synbiotic Mixture in Infants with Atopic Dermatitis. Clin. Exp. Allergy 2012, 42, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Xu, L.-Z.; Peng, K.; Wu, W.; Wu, R.; Liu, Z.-Q.; Yang, G.; Geng, X.-R.; Liu, J.; Liu, Z.-G.; et al. Specific Immunotherapy in Combination with Clostridium Butyricum Inhibits Allergic Inflammation in the Mouse Intestine. Sci. Rep. 2015, 5, 17651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.-Z.; Yang, L.-T.; Qiu, S.-Q.; Yang, G.; Luo, X.-Q.; Miao, B.-P.; Geng, X.-R.; Liu, Z.-Q.; Liu, J.; Wen, Z.; et al. Combination of Specific Allergen and Probiotics Induces Specific Regulatory B Cells and Enhances Specific Immunotherapy Effect on Allergic Rhinitis. Oncotarget 2016, 7, 54360–54369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manhart, N.; Vierlinger, K.; Habel, O.; Bergmeister, L.H.; Götzinger, P.; Sautner, T.; Spittler, A.; Boltz-Nitulescu, G.; Marian, B.; Roth, E. Lipopolysaccharide Causes Atrophy of Peyer’s Patches and an Increased Expression of CD28 and B7 Costimulatory Ligands. Shock 2000, 14, 478–483. [Google Scholar] [CrossRef]
- Li, Q.; Cheng, F.; Xu, Q.; Su, Y.; Cai, X.; Zeng, F.; Zhang, Y. The Role of Probiotics in Coronavirus Disease-19 Infection in Wuhan: A Retrospective Study of 311 Severe Patients. Int. Immunopharmacol. 2021, 95, 107531. [Google Scholar] [CrossRef] [PubMed]
- Azagra-Boronat, I.; Massot-Cladera, M.; Knipping, K.; Va not Land, B.; Stahl, B.; Garssen, J.; Rodríguez-Lagunas, M.J.; Franch, À.; Castell, M.; Pérez-Cano, F.J. Supplementation With 2′-FL and ScGOS/LcFOS Ameliorates Rotavirus-Induced Diarrhea in Suckling Rats. Front. Cell. Infect. Microbiol. 2018, 8, 372. [Google Scholar] [CrossRef]
- Morales-Ferré, C.; Azagra-Boronat, I.; Massot-Cladera, M.; Tims, S.; Knipping, K.; Garssen, J.; Knol, J.; Franch, À.; Castell, M.; Pérez-Cano, F.J.; et al. Preventive Effect of a Postbiotic and Prebiotic Mixture in a Rat Model of Early Life Rotavirus Induced-Diarrhea. Nutrients 2022, 14, 1163. [Google Scholar] [CrossRef]
- Zhang, W.; Azevedo, M.S.P.; Gonzalez, A.M.; Saif, L.J.; Van Nguyen, T.; Wen, K.; Yousef, A.E.; Yuan, L. Influence of Probiotic Lactobacilli Colonization on Neonatal B Cell Responses in a Gnotobiotic Pig Model of Human Rotavirus Infection and Disease. Vet. Immunol. Immunopathol. 2008, 122, 175–181. [Google Scholar] [CrossRef] [Green Version]
- Qiao, H.; Duffy, L.C.; Griffiths, E.; Dryja, D.; Leavens, A.; Rossman, J.; Rich, G.; Riepenhoff-Talty, M.; Locniskar, M. Immune Responses in Rhesus Rotavirus-Challenged BALB/c Mice Treated with Bifidobacteria and Prebiotic Supplements. Pediatr. Res. 2002, 51, 750–755. [Google Scholar] [CrossRef]
- Rigo-Adrover, M.; Saldaña-Ruíz, S.; van Limpt, K.; Knipping, K.; Garssen, J.; Knol, J.; Franch, A.; Castell, M.; Pérez-Cano, F.J. A Combination of ScGOS/LcFOS with Bifidobacterium Breve M-16V Protects Suckling Rats from Rotavirus Gastroenteritis. Eur. J. Nutr. 2017, 56, 1657–1670. [Google Scholar] [CrossRef]
- Rigo-Adrover, M.D.M.; van Limpt, K.; Knipping, K.; Garssen, J.; Knol, J.; Costabile, A.; Franch, À.; Castell, M.; Pérez-Cano, F.J. Preventive Effect of a Synbiotic Combination of Galacto- and Fructooligosaccharides Mixture with Bifidobacterium Breve M-16V in a Model of Multiple Rotavirus Infections. Front. Immunol. 2018, 9, 1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Pan, Y.; Xia, X.; Liang, J.; Liu, F.; Dou, H.; Hou, Y. Bacteroides Fragilis Alleviates the Symptoms of Lupus Nephritis via Regulating CD1d and CD86 Expressions in B Cells. Eur. J. Pharmacol. 2020, 884, 173421. [Google Scholar] [CrossRef] [PubMed]
- Mercadante, A.C.T.; Perobelli, S.M.; Alves, A.P.G.; Gonçalves-Silva, T.; Mello, W.; Gomes-Santos, A.C.; Miyoshi, A.; Azevedo, V.; Faria, A.M.C.; Bonomo, A. Oral Combined Therapy with Probiotics and Alloantigen Induces B Cell-Dependent Long-Lasting Specific Tolerance. J. Immunol. 2014, 192, 1928–1937. [Google Scholar] [CrossRef] [Green Version]
- Kleerebezem, M.; Binda, S.; Bron, P.A.; Gross, G.; Hill, C.; van Hylckama Vlieg, J.E.; Lebeer, S.; Satokari, R.; Ouwehand, A.C. Understanding Mode of Action Can Drive the Translational Pipeline towards More Reliable Health Benefits for Probiotics. Curr. Opin. Biotechnol. 2019, 56, 55–60. [Google Scholar] [CrossRef]
- Shen, N.T.; Maw, A.; Tmanova, L.L.; Pino, A.; Ancy, K.; Crawford, C.V.; Simon, M.S.; Evans, A.T. Timely Use of Probiotics in Hospitalized Adults Prevents Clostridium Difficile Infection: A Systematic Review with Meta-Regression Analysis. Gastroenterology 2017, 152, 1889–1900.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zmora, N.; Zilberman-Schapira, G.; Suez, J.; Mor, U.; Dori-Bachash, M.; Bashiardes, S.; Kotler, E.; Zur, M.; Regev-Lehavi, D.; Brik, R.B.-Z.; et al. Personalized Gut Mucosal Colonization Resistance to Empiric Probiotics Is Associated with Unique Host and Microbiome Features. Cell 2018, 174, 1388–1405.e21. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rousseaux, A.; Brosseau, C.; Bodinier, M. Immunomodulation of B Lymphocytes by Prebiotics, Probiotics and Synbiotics: Application in Pathologies. Nutrients 2023, 15, 269. https://doi.org/10.3390/nu15020269
Rousseaux A, Brosseau C, Bodinier M. Immunomodulation of B Lymphocytes by Prebiotics, Probiotics and Synbiotics: Application in Pathologies. Nutrients. 2023; 15(2):269. https://doi.org/10.3390/nu15020269
Chicago/Turabian StyleRousseaux, Anaïs, Carole Brosseau, and Marie Bodinier. 2023. "Immunomodulation of B Lymphocytes by Prebiotics, Probiotics and Synbiotics: Application in Pathologies" Nutrients 15, no. 2: 269. https://doi.org/10.3390/nu15020269
APA StyleRousseaux, A., Brosseau, C., & Bodinier, M. (2023). Immunomodulation of B Lymphocytes by Prebiotics, Probiotics and Synbiotics: Application in Pathologies. Nutrients, 15(2), 269. https://doi.org/10.3390/nu15020269