

Astaxanthin Supplementation Augments the Benefits of CrossFit Workouts on Semaphorin 3C and Other Adipokines in Males with Obesity

 , ,
, ,  , , ,
, , ,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
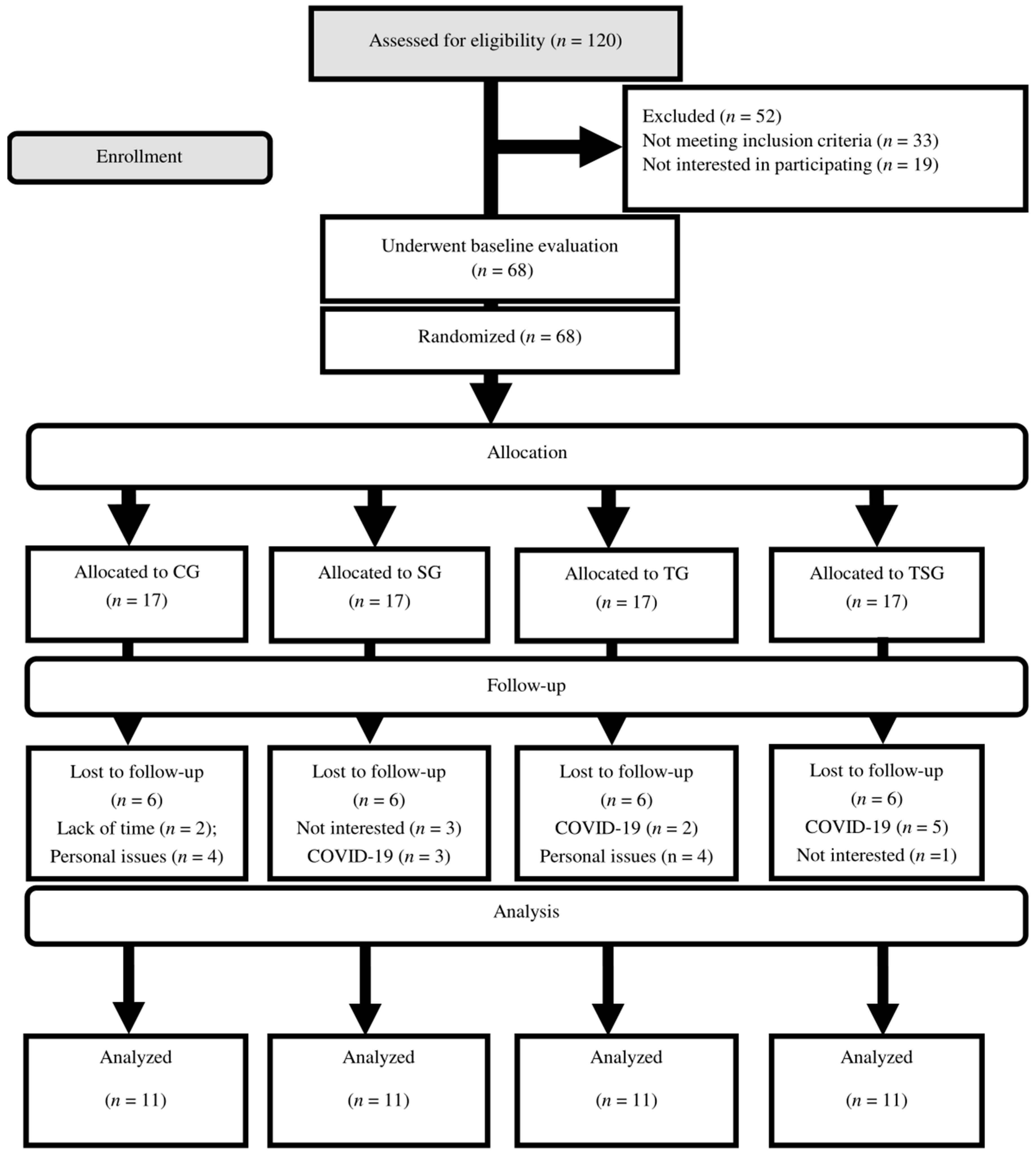
2. Methods
2.1. Experimental Design
2.2. Training Protocols
2.3. Astaxanthin Supplementation Protocol
2.4. Nutrient Intake and Dietary Analysis
2.5. Blood Markers
2.6. Statistical Analysis
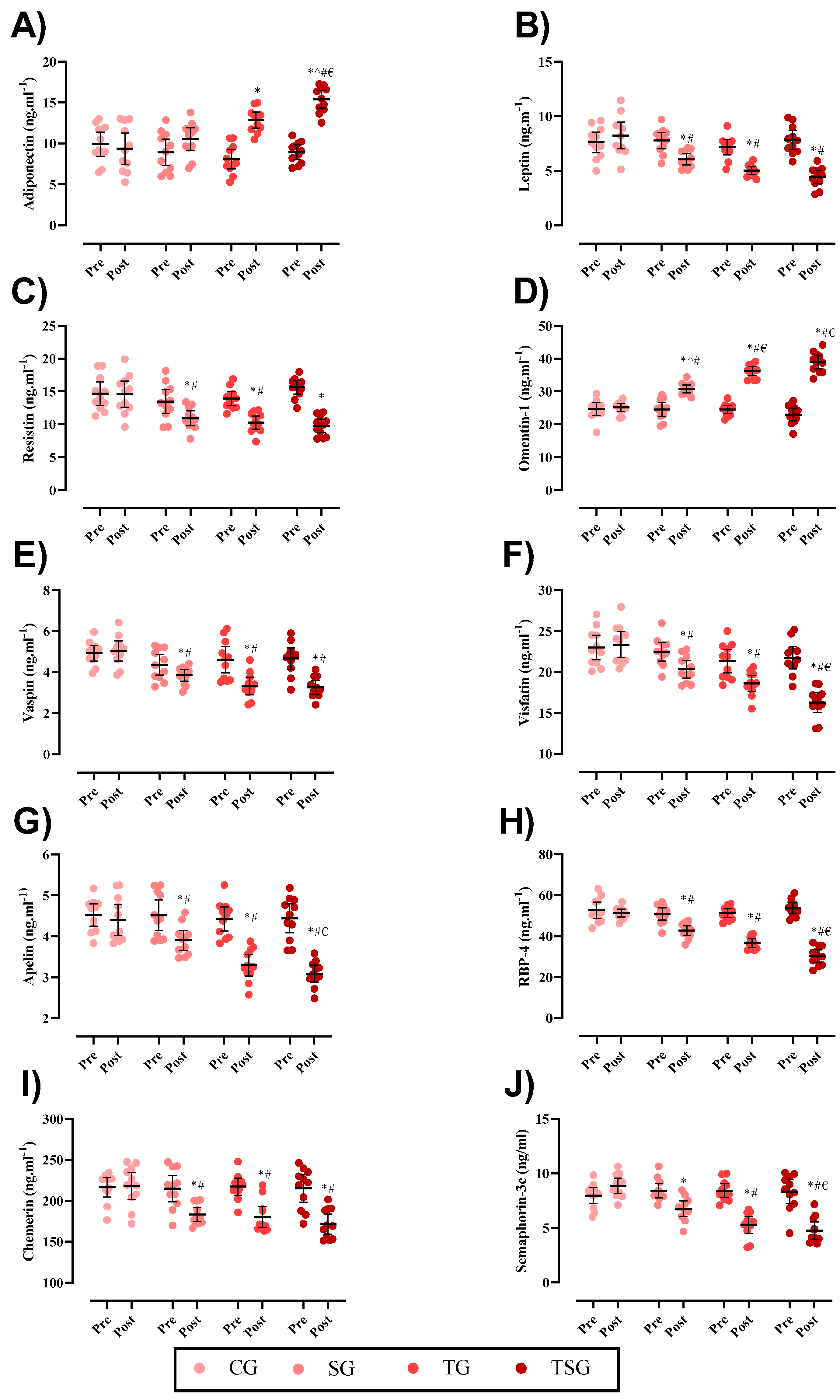
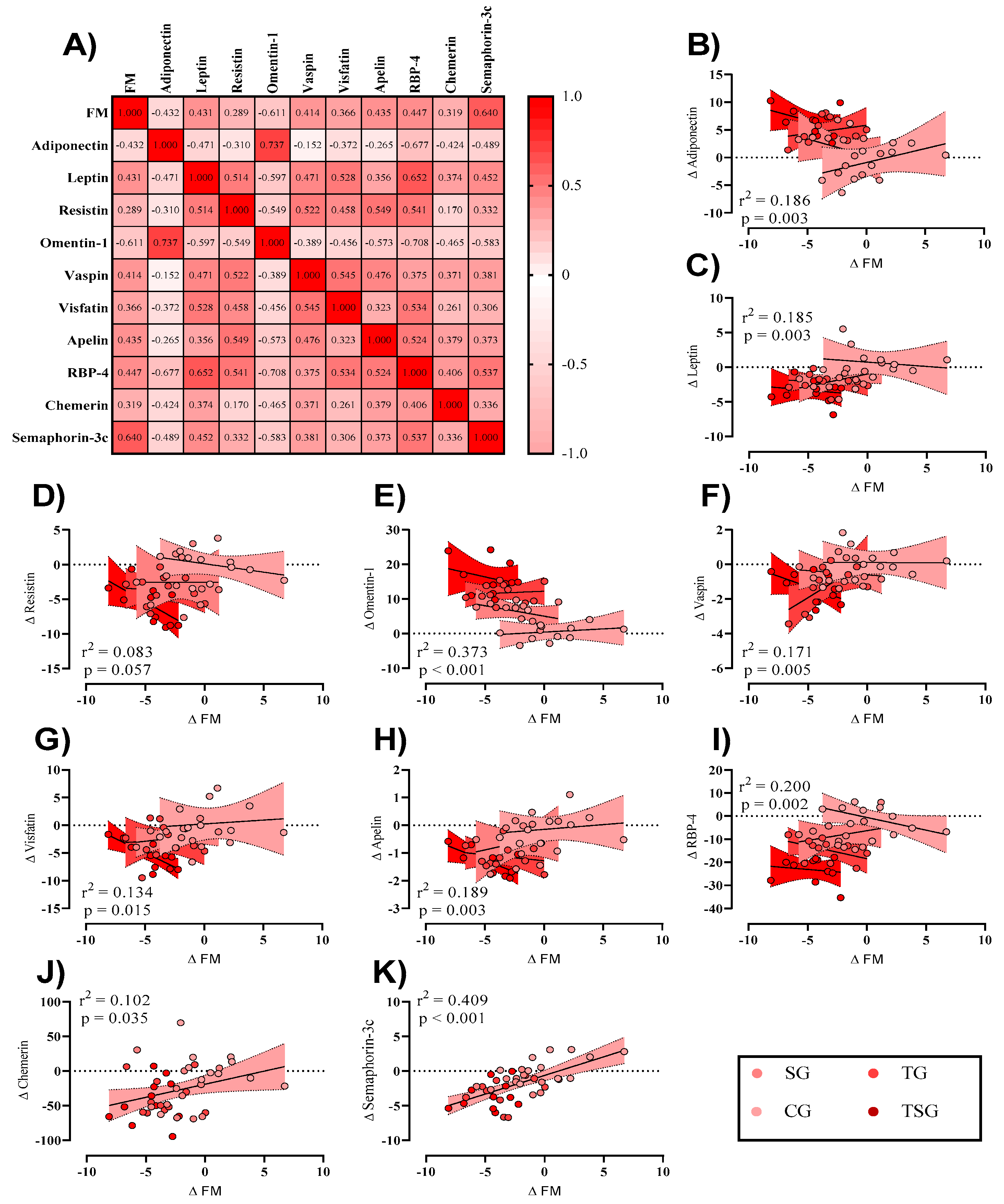
3. Results
3.1. Compliance, Adverse Events, and Nutrient Intakes
3.2. Adipokines
3.3. Weight and BMI
4. Discussion
Study Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taylor, E.B. The complex role of adipokines in obesity, inflammation, and autoimmunity. Clin. Sci. 2021, 135, 731–752. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, A.; Clinchamps, M.; Pereira, B.; Courteix, D.; Lesourd, B.; Chapier, R.; Obert, P.; Vinet, A.; Walther, G.; Chaplais, E.; et al. Dietary fibres and the management of obesity and metabolic syndrome: The RESOLVE study. Nutrients 2020, 12, 2911. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, K.; Shibata, R.; Murohara, T.; Ouchi, N. Role of anti-inflammatory adipokines in obesity-related diseases. Trends Endocrinol. Metab. 2014, 25, 348–355. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Leal, V.; Mafra, D. Adipokines in obesity. Clin. Chim. Acta 2013, 419, 87–94. [Google Scholar] [CrossRef]
- Eskandari, M.; Moghadam, B.H.; Bagheri, R.; Ashtary-Larky, D.; Eskandari, E.; Nordvall, M.; Dutheil, F.; Wong, A. Effects of interval jump rope exercise combined with dark chocolate supplementation on inflammatory adipokine, cytokine concentrations, and body composition in obese adolescent boys. Nutrients 2020, 12, 3011. [Google Scholar] [CrossRef]
- Abdi, A.; Mehrabani, J.; Nordvall, M.; Wong, A.; Fallah, A.; Bagheri, R. Effects of concurrent training on irisin and fibronectin type-III domain containing 5 (FNDC5) expression in visceral adipose tissue in type-2 diabetic rats. Arch. Physiol. Biochem. 2022, 128, 651–656. [Google Scholar] [CrossRef]
- Khan, M.; Joseph, F. Adipose tissue and adipokines: The association with and application of adipokines in obesity. Scientifica 2014, 2014, 328592. [Google Scholar] [CrossRef]
- González Izquierdo, A.; Crujeiras Martínez, A.B.; Casanueva Freijo, F.; Couselo Carreira, M. Leptin, obesity, and leptin resistance: Where are we 25 years later? Nutrients 2019, 11, 2704. [Google Scholar] [CrossRef]
- Ashtary-Larky, D.; Kashkooli, S.; Bagheri, R.; Lamuchi-Deli, N.; Alipour, M.; Mombaini, D.; Baker, J.S.; Ahmadi, A.R.; Wong, A. The effect of exercise training on serum concentrations of chemerin in patients with metabolic diseases: A systematic review and meta-analysis. Arch. Physiol. Biochem. 2021, 129, 1–10. [Google Scholar] [CrossRef]
- Glassman, G. CrossFit Training: Level 1 Training Guide; CrossFit, Inc.: Santa Cruz, CA, USA, 2019. [Google Scholar]
- Claudino, J.G.; Gabbett, T.J.; Bourgeois, F.; Souza, H.d.S.; Miranda, R.C.; Mezêncio, B.; Soncin, R.; Filho, C.A.C.; Bottaro, M.; Hernandez, A.J.; et al. CrossFit overview: Systematic review and meta-analysis. Sports Med. Open 2018, 4, 1–14. [Google Scholar] [CrossRef]
- Gianzina, E.A.; Kassotaki, O.A. The benefits and risks of the high-intensity CrossFit training. Sport Sci. Health 2019, 15, 21–33. [Google Scholar] [CrossRef]
- Farkhondeh, T.; Llorens, S.; Pourbagher-Shahri, A.M.; Ashrafizadeh, M.; Talebi, M.; Shakibaei, M.; Samarghandian, S. An overview of the role of adipokines in cardiometabolic diseases. Molecules 2020, 25, 5218. [Google Scholar] [CrossRef]
- Hosseini, M.; Bagheri, R.; Nikkar, H.; Baker, J.S.; Jaime, S.J.; Mosayebi, Z.; Zouraghi, M.R.; Wong, A. The effect of interval training on adipokine plasmatic levels in rats with induced myocardial infarction. Arch. Physiol. Biochem. 2022, 128, 1249–1253. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.H.; Park, H.S.; Kim, K.-S.; Choi, W.H.; Ahn, C.W.; Kim, B.T.; Kim, S.M.; Lee, S.Y.; Ahn, S.M.; Kim, Y.K.; et al. Effect of weight loss on some serum cytokines in human obesity: Increase in IL-10 after weight loss. J. Nutr. Biochem. 2008, 19, 371–375. [Google Scholar] [CrossRef]
- Bagheri, R.; Rashidlamir, A.; Ashtary-Larky, D.; Wong, A.; Alipour, M.; Motevalli, M.S.; Chebbi, A.; Laher, I.; Zouhal, H. Does green tea extract enhance the anti-inflammatory effects of exercise on fat loss? Br. J. Clin. Pharmacol. 2020, 86, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, R.; Rashidlamir, A.; Ashtary-Larky, D.; Wong, A.; Grubbs, B.; Motevalli, M.S.; Baker, J.S.; Laher, I.; Zouhal, H. Effects of green tea extract supplementation and endurance training on irisin, pro-inflammatory cytokines, and adiponectin concentrations in overweight middle-aged men. Eur. J. Appl. Physiol. 2020, 120, 915–923. [Google Scholar] [CrossRef] [PubMed]
- Düzova, H.; Güllü, E.; Çiçek, G.; Köksal, B.; Kayhan, B.; Güllü, A.; Şahin, İ. The effect of exercise induced weight loss on myokines and adipokines in overweight sedentary females steps aerobics vs. jogging walking exercises. J. Sports Med. Phys. Fit. 2016, 58, 295–308. [Google Scholar]
- Ouerghi, N.; Ben Fradj, M.K.; Bezrati, I.; Feki, M.; Kaabachi, N.; Bouassida, A. Effect of high-intensity interval training on plasma omentin-1 concentration in overweight/obese and normal-weight youth. Obes. Facts 2017, 10, 323–331. [Google Scholar] [CrossRef]
- Urbanová, M.; Dostálová, I.; Trachta, P.; Drápalová, J.; Kaválková, P.; Haluzíková, D.; Matoulek, M.; Lacinová, Z.; Mráz, M.; Kasalický, M.; et al. Serum concentrations and subcutaneous adipose tissue mRNA expression of omentin in morbid obesity and type 2 diabetes mellitus: The effect of very-low-calorie diet, physical activity and laparoscopic sleeve gastrectomy. Physiol. Res. 2014, 63, 207–218. [Google Scholar] [CrossRef]
- Faramarzi, M.; Banitalebi, E.; Nori, S.; Farzin, S.; Taghavian, Z. Effects of rhythmic aerobic exercise plus core stability training on serum omentin, chemerin and vaspin levels and insulin resistance of overweight women. J. Sports Med. Phys. Fit. 2015, 56, 476–482. [Google Scholar]
- Taherkhani, S.; Suzuki, K.; Castell, L. A short overview of changes in inflammatory cytokines and oxidative stress in response to physical activity and antioxidant supplementation. Antioxidants 2020, 9, 886. [Google Scholar] [CrossRef]
- Aoi, W.; Naito, Y.; Takanami, Y.; Ishii, T.; Kawai, Y.; Akagiri, S.; Kato, Y.; Osawa, T.; Yoshikawa, T. Astaxanthin improves muscle lipid metabolism in exercise via inhibitory effect of oxidative CPT I modification. Biochem. Biophys. Res. Commun. 2008, 366, 892–897. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Tang, N.; Kord-Varkaneh, H.; Low, T.Y.; Tan, S.C.; Wu, X.; Zhu, Y. The effects of astaxanthin supplementation on obesity, blood pressure, CRP, glycemic biomarkers, and lipid profile: A meta-analysis of randomized controlled trials. Pharmacol. Res. 2020, 161, 105113. [Google Scholar] [CrossRef] [PubMed]
- Fakhri, S.; Abbaszadeh, F.; Dargahi, L.; Jorjani, M. Astaxanthin: A mechanistic review on its biological activities and health benefits. Pharmacol. Res. 2018, 136, 1–20. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, H. Inhibitory effect of astaxanthin on oxidative stress-induced mitochondrial dysfunction—A mini-review. Nutrients 2018, 10, 1137. [Google Scholar] [CrossRef] [PubMed]
- Nishida, Y.; Nawaz, A.; Kado, T.; Takikawa, A.; Igarashi, Y.; Onogi, Y.; Wada, T.; Sasaoka, T.; Yamamoto, S.; Sasahara, M.; et al. Astaxanthin stimulates mitochondrial biogenesis in insulin resistant muscle via activation of AMPK pathway. J. Cachexia Sarcopenia Muscle 2020, 11, 241–258. [Google Scholar] [CrossRef]
- Murai, T.; Kawasumi, K.; Tominaga, K.; Okada, Y.; Kobayashi, M.; Arai, T. Effects of astaxanthin supplementation in healthy and obese dogs. Vet. Med. Res. Rep. 2019, 10, 29. [Google Scholar] [CrossRef]
- Oharomari, L.K.; Ikemoto, M.J.; Hwang, D.J.; Koizumi, H.; Soya, H. Benefits of Exercise and Astaxanthin Supplementation: Are There Additive or Synergistic Effects? Antioxidants 2021, 10, 870. [Google Scholar] [CrossRef]
- Saeidi, A.; Nouri-Habashi, A.; Razi, O.; Ataeinosrat, A.; Rahmani, H.; Mollabashi, S.S.; Bagherzadeh-Rahmani, B.; Aghdam, S.M.; Khalajzadeh, L.; Al Kiyumi, M.H.; et al. Astaxanthin Supplemented with High-Intensity Functional Training Decreases Adipokines Levels and Cardiovascular Risk Factors in Men with Obesity. Nutrients 2023, 15, 286. [Google Scholar] [CrossRef]
- Moghadam, B.H.; Bagheri, R.; Roozbeh, B.; Ashtary-Larky, D.; Gaeini, A.A.; Dutheil, F.; Wong, A. Impact of saffron (Crocus sativus Linn) supplementation and resistance training on markers implicated in depression and happiness levels in untrained young males. Physiol. Behav. 2021, 233, 113352. [Google Scholar] [CrossRef]
- Thomas, S.; Reading, J.; Shephard, R.J. Revision of the physical activity readiness questionnaire (PAR-Q). Can. J. Sport Sci. 1992, 17, 338–345. [Google Scholar] [PubMed]
- Nathanson, V. Revising the declaration of Helsinki. BMJ 2013, 346, f2837. [Google Scholar] [CrossRef] [PubMed]
- Glassman, G. A theoretical template for crossfit’s programming. CrossFit J. 2003, 6, 1–5. [Google Scholar]
- Heinrich, K.M.; Patel, P.M.; O’Neal, J.L.; Heinrich, B.S. High-intensity compared to moderate-intensity training for exercise initiation, enjoyment, adherence, and intentions: An intervention study. BMC Public Health 2014, 14, 1–6. [Google Scholar] [CrossRef]
- Choi, H.D.; Kim, J.H.; Chang, M.J.; Kyu-Youn, Y.; Shin, W.G. Effects of astaxanthin on oxidative stress in overweight and obese adults. Phytother. Res. 2011, 25, 1813–1818. [Google Scholar] [CrossRef]
- Annibalini, G.; Lucertini, F.; Agostini, D.; Vallorani, L.; Gioacchini, A.; Barbieri, E.; Guescini, M.; Casadei, L.; Passalia, A.; Del Sal, M.; et al. Concurrent aerobic and resistance training has anti-inflammatory effects and increases both plasma and leukocyte levels of IGF-1 in late middle-aged type 2 diabetic patients. Oxidative Med. Cell. Longev. 2017, 2017, 3937842. [Google Scholar] [CrossRef]
- Ratner, B. The correlation coefficient: Its values range between +1/−1, or do they? J. Target. Meas. Anal. Mark. 2009, 17, 139–142. [Google Scholar] [CrossRef]
- Ashtary-Larky, D.; Lamuchi-Deli, N.; Kashkooli, S.; Mombaini, D.; Alipour, M.; Khodadadi, F.; Bagheri, R.; Dutheil, F.; Wong, A. The effects of exercise training on serum concentrations of chemerin in individuals with overweight and obesity: A systematic review, meta-analysis, and meta-regression of 43 clinical trials. Arch. Physiol. Biochem. 2021, 129, 1012–1027. [Google Scholar] [CrossRef]
- Saeidi, A.; Haghighi, M.M.; Kolahdouzi, S.; Daraei, A.; Ben Abderrahmane, A.; Essop, M.F.; Laher, I.; Hackney, A.C.; Zouhal, H. The effects of physical activity on adipokines in individuals with overweight/obesity across the lifespan: A narrative review. Obes. Rev. 2021, 22, e13090. [Google Scholar] [CrossRef]
- Højbjerre, L.; Rosenzweig, M.; Dela, F.; Bruun, J.M.; Stallknecht, B.; Verkauskiene, R.; Beltrand, J.; Claris, O.; Chevenne, D.; Deghmoun, S.; et al. Acute exercise increases adipose tissue interstitial adiponectin concentration in healthy overweight and lean subjects. Eur. J. Endocrinol. 2007, 157, 613–623. [Google Scholar] [CrossRef]
- Saunders, T.J.; Palombella, A.; McGuire, K.A.; Janiszewski, P.M.; Després, J.-P.; Ross, R. Acute exercise increases adiponectin levels in abdominally obese men. J. Nutr. Metab. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Varady, K.A.; Bhutani, S.; Church, E.C.; Phillips, S.A. Adipokine responses to acute resistance exercise in trained and untrained men. Med. Sci. Sports Exerc. 2010, 42, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Bouassida, A.; Lakhdar, N.; Benaissa, N.; Mejri, S.; Zaouali, M.; Zbidi, A.; Tabka, Z. Adiponectin responses to acute moderate and heavy exercises in overweight middle aged subjects. J. Sports Med. Phys. Fit. 2010, 50, 330–335. [Google Scholar]
- Jamurtas, A.Z.; Theocharis, V.; Koukoulis, G.; Stakias, N.; Fatouros, I.G.; Kouretas, D.; Koutedakis, Y. The effects of acute exercise on serum adiponectin and resistin levels and their relation to insulin sensitivity in overweight males. Eur. J. Appl. Physiol. 2006, 97, 122–126. [Google Scholar] [CrossRef]
- Yoshida, H.; Yanai, H.; Ito, K.; Tomono, Y.; Koikeda, T.; Tsukahara, H.; Tada, N. Administration of natural astaxanthin increases serum HDL-cholesterol and adiponectin in subjects with mild hyperlipidemia. Atherosclerosis 2010, 209, 520–523. [Google Scholar] [CrossRef]
- Legakis, I.N.; Mantzouridis, T.; Saramantis, A.; Lakka-Papadodima, E. Rapid decrease of leptin in middle-aged sedentary individuals after 20 minutes of vigorous exercise with early recovery after the termination of the test. J. Endocrinol. Investig. 2004, 27, 117–120. [Google Scholar] [CrossRef]
- Cruz, I.S.; Rosa, G.; Valle, V.; Mello, D.B.D.; Fortes, M.; Dantas, E.H. Acute effects of concurrent training on serum leptin and cortisol in overweight young adults. Rev. Bras. Med. Esporte 2012, 18, 81–86. [Google Scholar] [CrossRef]
- Racette, S.B.; Coppack, S.W.; Landt, M.; Klein, S. Leptin production during moderate-intensity aerobic exercise. J. Clin. Endocrinol. Metab. 1997, 82, 2275–2277. [Google Scholar] [CrossRef]
- Weltman, A.; Pritzlaff, C.J.; Wideman, L.; Considine, R.V.; Fryburg, D.A.; Gutgesell, M.E.; Hartman, M.L.; Veldhuis, J.D. Intensity of acute exercise does not affect serum leptin concentrations in young men. Med. Sci. Sports Exerc. 2000, 32, 1556–1561. [Google Scholar] [CrossRef]
- Kumar, A.; Dhaliwal, N.; Dhaliwal, J.; Dharavath, R.N.; Chopra, K. Astaxanthin attenuates oxidative stress and inflammatory responses in complete Freund-adjuvant-induced arthritis in rats. Pharmacol. Rep. 2020, 72, 104–114. [Google Scholar] [CrossRef]
- Feng, Y.; Chu, A.; Luo, Q.; Wu, M.; Shi, X.; Chen, Y. The protective effect of astaxanthin on cognitive function via inhibition of oxidative stress and inflammation in the brains of chronic T2DM rats. Front. Pharmacol. 2018, 9, 748. [Google Scholar] [CrossRef] [PubMed]
- Kohandel, Z.; Farkhondeh, T.; Aschner, M.; Samarghandian, S. Nrf2 a molecular therapeutic target for Astaxanthin. Biomed. Pharmacother. 2021, 137, 111374. [Google Scholar] [CrossRef] [PubMed]
- Hussein, G.; Sankawa, U.; Goto, H.; Matsumoto, A.K.; Watanabe, H. Astaxanthin, a carotenoid with potential in human health and nutrition. J. Nat. Prod. 2006, 69, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Boshra, S.A. Astaxanthin Attenuates Adiponectin, Calprotectin, miRNA222 and miRNA378 in Obesity induced by High-Fat Diet in Rats. Curr. Pharm. Biotechnol. 2022, 23, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Vardar, S.A.; Karaca, A.; Güldiken, S.; Palabıyık, O.; Süt, N.; Demir, A.M. High-intensity interval training acutely alters plasma adipokine levels in young overweight/obese women. Arch. Physiol. Biochem. 2018, 124, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Falcão-Pires, I.; Castro-Chaves, P.; Miranda-Silva, D.; Lourenço, A.P.; Leite-Moreira, A.F. Physiological, pathological and potential therapeutic roles of adipokines. Drug Discov. Today 2012, 17, 880–889. [Google Scholar] [CrossRef]
- Racil, G.; Zouhal, H.; Elmontassar, W.; Ben Abderrahmane, A.; De Sousa, M.V.; Chamari, K.; Amri, M.; Coquart, J.B. Plyometric exercise combined with high-intensity interval training improves metabolic abnormalities in young obese females more so than interval training alone. Appl. Physiol. Nutr. Metab. 2016, 41, 103–109. [Google Scholar] [CrossRef]
- Madsen, S.M.; Thorup, A.C.; Bjerre, M.; Jeppesen, P.B. Does 8 weeks of strenuous bicycle exercise improve diabetes-related inflammatory cytokines and free fatty acids in type 2 diabetes patients and individuals at high-risk of metabolic syndrome? Arch. Physiol. Biochem. 2015, 121, 129–138. [Google Scholar] [CrossRef]
- Verheggen, R.J.H.M.; Poelkens, F.; Roerink, S.H.P.P.; Ramakers, R.E.F.S.; Catoire, M.; Hermus, A.R.M.M.; Thijssen, D.H.J.; Hopman, M.T.E. Exercise Improves Insulin Sensitivity in the Absence of Changes in Cytokines. Med. Sci. Sports Exerc. 2016, 48, 2378–2386. [Google Scholar] [CrossRef]
- Feito, Y.; Hoffstetter, W.; Serafini, P.; Mangine, G. Changes in body composition, bone metabolism, strength, and skill-specific performance resulting from 16-weeks of HIFT. PLoS ONE 2018, 13, e0198324. [Google Scholar] [CrossRef]
- Zouhal, H.; Zare-Kookandeh, N.; Haghighi, M.M.; Daraei, A.; de Sousa, M.; Soltani, M.; Ben Abderrahman, A.; Tijani, J.M.; Hackney, A.C.; Laher, I.; et al. Physical activity and adipokine levels in individuals with type 2 diabetes: A literature review and practical applications. Rev. Endocr. Metab. Disord. 2021, 22, 987–1011. [Google Scholar] [CrossRef] [PubMed]
- Nikseresht, M.; Hafezi Ahmadi, M.R.; Hedayati, M. Detraining-induced alterations in adipokines and cardiometabolic risk factors after nonlinear periodized resistance and aerobic interval training in obese men. Appl. Physiol. Nutr. Metab. 2016, 41, 1018–1025. [Google Scholar] [CrossRef]
- AminiLari, Z.; Fararouei, M.; Amanat, S.; Sinaei, E.; Dianatinasab, S.; AminiLari, M.; Daneshi, N.; Dianatinasab, M. The effect of 12 weeks aerobic, resistance, and combined exercises on omentin-1 levels and insulin resistance among type 2 diabetic middle-aged women. Diabetes Metab. J. 2017, 41, 205. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Tian, Y.; Valenzuela, P.L.; Huang, C.; Zhao, J.; Hong, P.; He, Z.; Yin, S.; Lucia, A. Myokine/adipokine response to “aerobic” exercise: Is it just a matter of exercise load? Front. Physiol. 2019, 10, 691. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Watanabe-Kominato, K.; Takahashi, Y.; Kojima, M.; Watanabe, R. Adipose tissue-derived omentin-1 function and regulation. Compr. Physiol. 2011, 7, 765–781. [Google Scholar]
- Moreno-Navarrete, J.M.; Catalán, V.; Ortega, F.; Gómez-Ambrosi, J.; Ricart, W.; Frühbeck, G.; Fernández-Real, J.M. Circulating omentin concentration increases after weight loss. Nutr. Metab. 2010, 7, 27. [Google Scholar] [CrossRef]
- Ouerghi, N.; Ben Fradj, M.K.; Duclos, M.; Bouassida, A.; Feki, M.; Weiss, K.; Knechtle, B. Effects of High-Intensity Interval Training on Selected Adipokines and Cardiometabolic Risk Markers in Normal-Weight and Overweight/Obese Young Males: A Pre-Post Test Trial. Biology 2022, 11, 853. [Google Scholar] [CrossRef]
- Kazemi, A.; Rahmati, M.; Dabaghzadeh, R.; Raisi, S.; Aghamolaie, S. The effect of high volume high intensity interval training on serum visfatin and vaspin, insulin resistance, lipid profile and body composition of overweight men. Daneshvar Med. 2015, 23, 325. [Google Scholar]
- Nam, J.S.; Ahn, C.W.; Park, H.J.; Kim, Y.S. Semaphorin 3 C is a novel adipokine representing exercise-induced improvements of metabolism in metabolically healthy obese young males. Sci. Rep. 2020, 10, 10005. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| CG | SG | TG | TSG | |||||
|---|---|---|---|---|---|---|---|---|
| Pre | Post | Pre | Post | Pre | Post | Pre | Post | |
| Energy (kcal/day) | 2260 ± 47 | 2269 ± 56 | 2278 ± 101 | 2149 ± 100 | 2269 ± 117 | 2141 ± 117 | 2273 ± 157 | 2129 ± 126 |
| CHO (g/day) | 281 ± 31.4 | 283 ± 33.3 | 279.4 ± 27.1 | 261 ± 27.5 | 289 ± 48.6 | 261 ± 39.2 | 288 ± 38.6 | 259 ± 29.1 |
| Fat (g/day) | 82.2 ± 11.0 | 81 ± 9.8 | 86.5 ± 10.7 | 75 ± 11.2 | 83.4 ± 12.4 | 73.1 ± 11.2 | 80.8 ± 13.87 | 70.2 ± 11.3 |
| Protein (g/day) | 104 ± 12.0 | 106 ± 11.3 | 101 ± 13.5 | 93 ± 12.6 | 103 ± 14.8 | 94 ± 11.7 | 102 ± 14.5 | 90 ± 13.5 |
| Body Weight (kg) | 95.3 ± 1.8 | 92.1 ± 2.1 | 94.2 ± 2.6 | 90.1 ± 2.3 a | 94.3 ± 0.9 | 90.1 ± 2.3 a,b | 95.1 ± 1.9 | 88.2 ± 2.3 a,b,ab |
| BMI (kg/m2) | 34.1 ± 2.5 | 33.7 ± 1.4 | 33.2 ± 1.4 | 32.4 ± 1.6 a,b | 33.5 ± 1.7 | 32.1 ± 1.5 a,b | 33.8 ± 1.2 | 31.8 ± 0.6 a,b,ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Supriya, R.; Shishvan, S.R.; Kefayati, M.; Abednatanzi, H.; Razi, O.; Bagheri, R.; Escobar, K.A.; Pashaei, Z.; Saeidi, A.; Shahrbanian, S.; et al. Astaxanthin Supplementation Augments the Benefits of CrossFit Workouts on Semaphorin 3C and Other Adipokines in Males with Obesity. Nutrients 2023, 15, 4803. https://doi.org/10.3390/nu15224803
Supriya R, Shishvan SR, Kefayati M, Abednatanzi H, Razi O, Bagheri R, Escobar KA, Pashaei Z, Saeidi A, Shahrbanian S, et al. Astaxanthin Supplementation Augments the Benefits of CrossFit Workouts on Semaphorin 3C and Other Adipokines in Males with Obesity. Nutrients. 2023; 15(22):4803. https://doi.org/10.3390/nu15224803
Chicago/Turabian StyleSupriya, Rashmi, Sevda Rahbari Shishvan, Movahed Kefayati, Hossein Abednatanzi, Omid Razi, Reza Bagheri, Kurt A. Escobar, Zhaleh Pashaei, Ayoub Saeidi, Shahnaz Shahrbanian, and et al. 2023. "Astaxanthin Supplementation Augments the Benefits of CrossFit Workouts on Semaphorin 3C and Other Adipokines in Males with Obesity" Nutrients 15, no. 22: 4803. https://doi.org/10.3390/nu15224803
APA StyleSupriya, R., Shishvan, S. R., Kefayati, M., Abednatanzi, H., Razi, O., Bagheri, R., Escobar, K. A., Pashaei, Z., Saeidi, A., Shahrbanian, S., Bagchi, S., Sengupta, P., Al Kiyumi, M. H., Heinrich, K. M., & Zouhal, H. (2023). Astaxanthin Supplementation Augments the Benefits of CrossFit Workouts on Semaphorin 3C and Other Adipokines in Males with Obesity. Nutrients, 15(22), 4803. https://doi.org/10.3390/nu15224803