
Gut-Microbiota, and Multiple Sclerosis: Background, Evidence, and Perspectives
 ,
,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
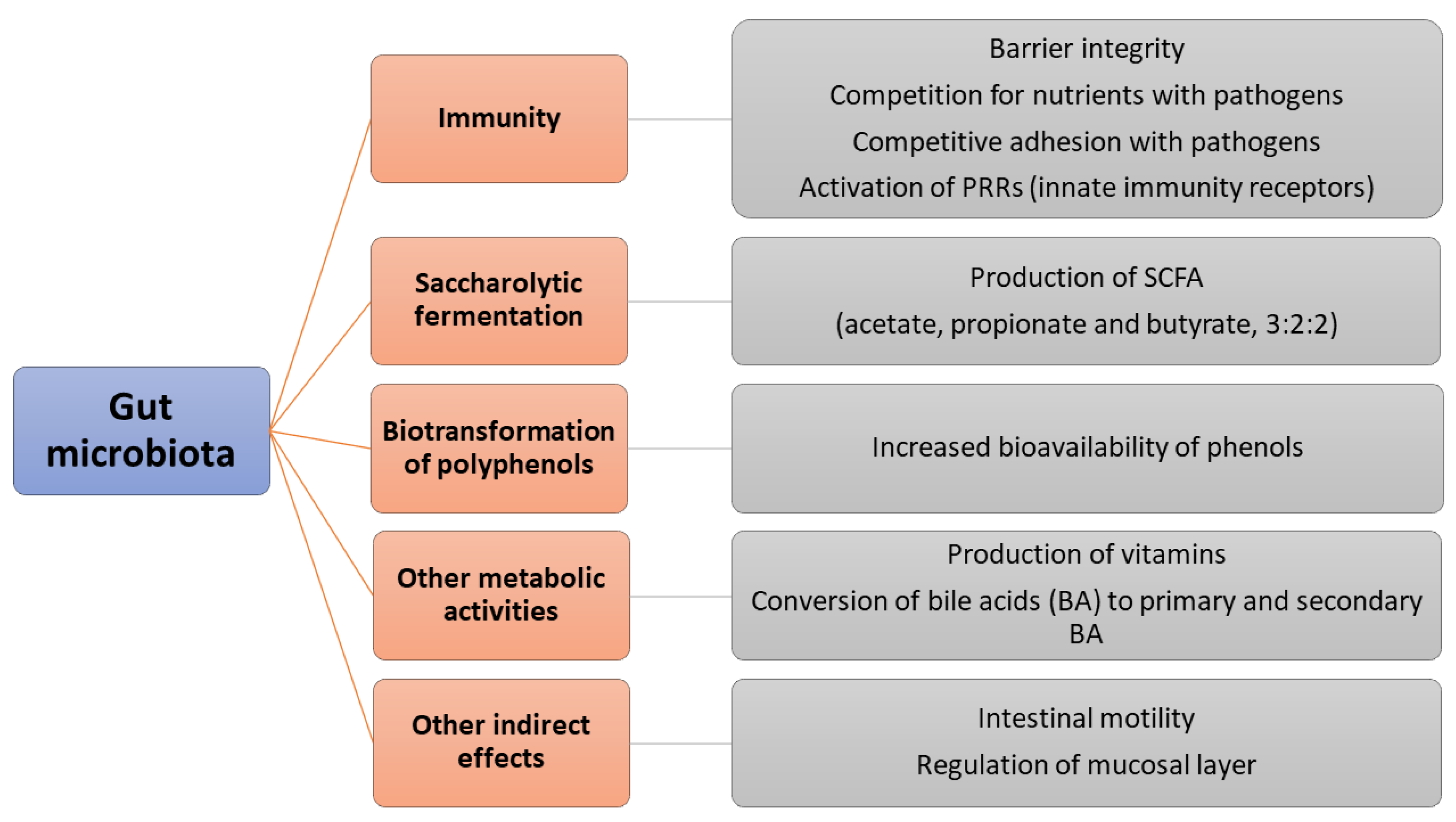
2. Gut Microbiota
- Enterotype 1: the dominant genus is Bacteroides, followed by Parabacteroides. These two genera show abundant expression of some enzymes, like galactosidase, hexosaminidase, and protease. Individuals with enterotype-1 can recover the most energy from the fermentation of carbohydrates and proteins; as a result, they are accustomed to a Western-style diet. This enterotype shows a higher production of riboflavin, pantothenic acid, and biotin (B2–B5–B8) compared to the others. Such a composition of the gut microbiota often correlates with a general inflammatory condition.
- Enterotype 2: the dominant genera are Prevotella and Desulfovibrio; they can produce high levels of thiamine (B1) and folic acid (B9). Therefore, this enterotype is the result of a purely vegetarian diet.
- Enterotype 3: the dominant genera are Ruminococcus and Akkermansia, which prefer simple sugars but can degrade mucin. These microorganisms can unbalance the normal functioning of the immune system [6].
3. Gut-Brain Axis: Role of Microbiota
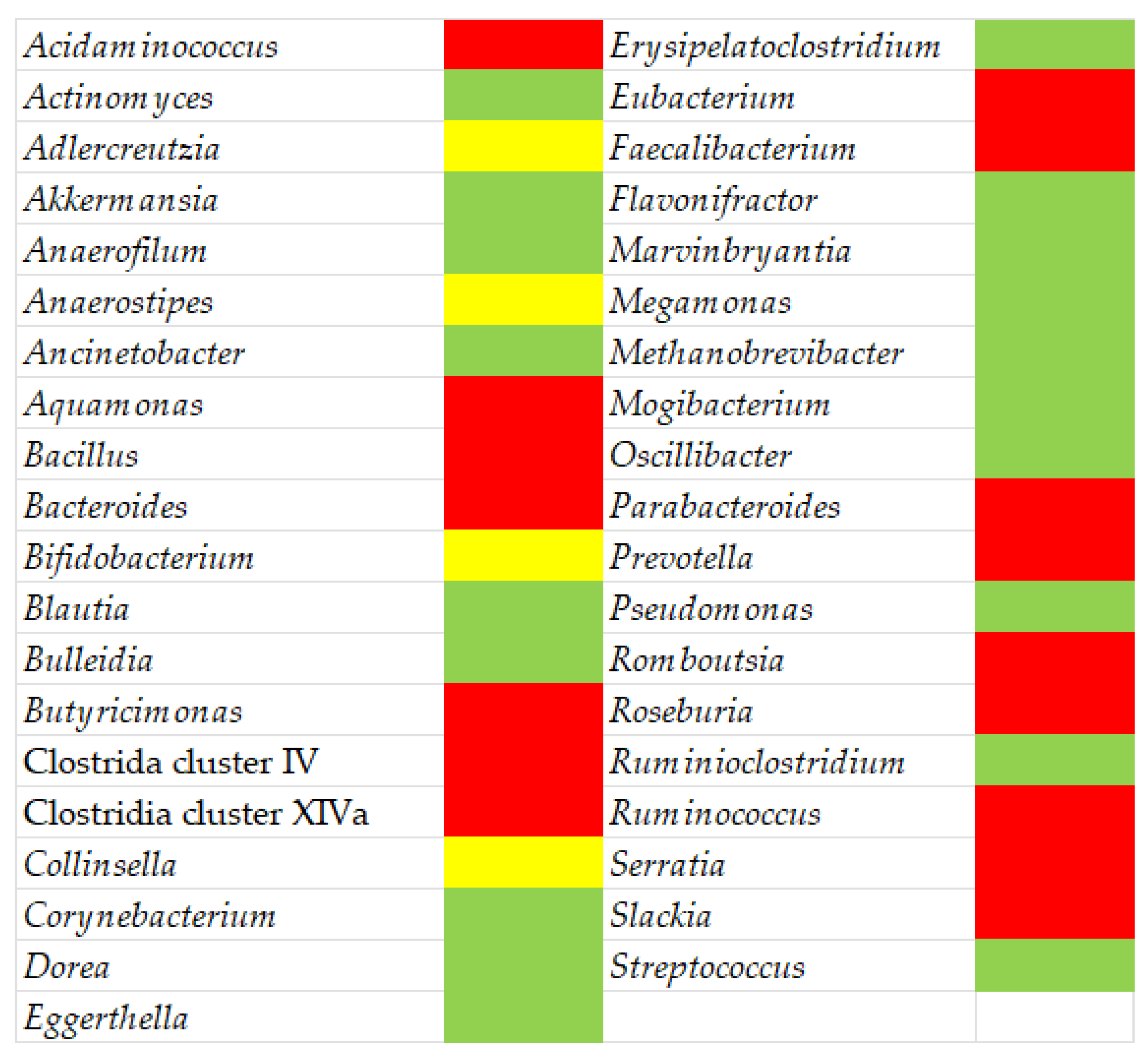
4. Multiple Sclerosis and Gut Microbiota
5. Dysbiosis in Multiple Sclerosis Patients and Therapeutic Approaches
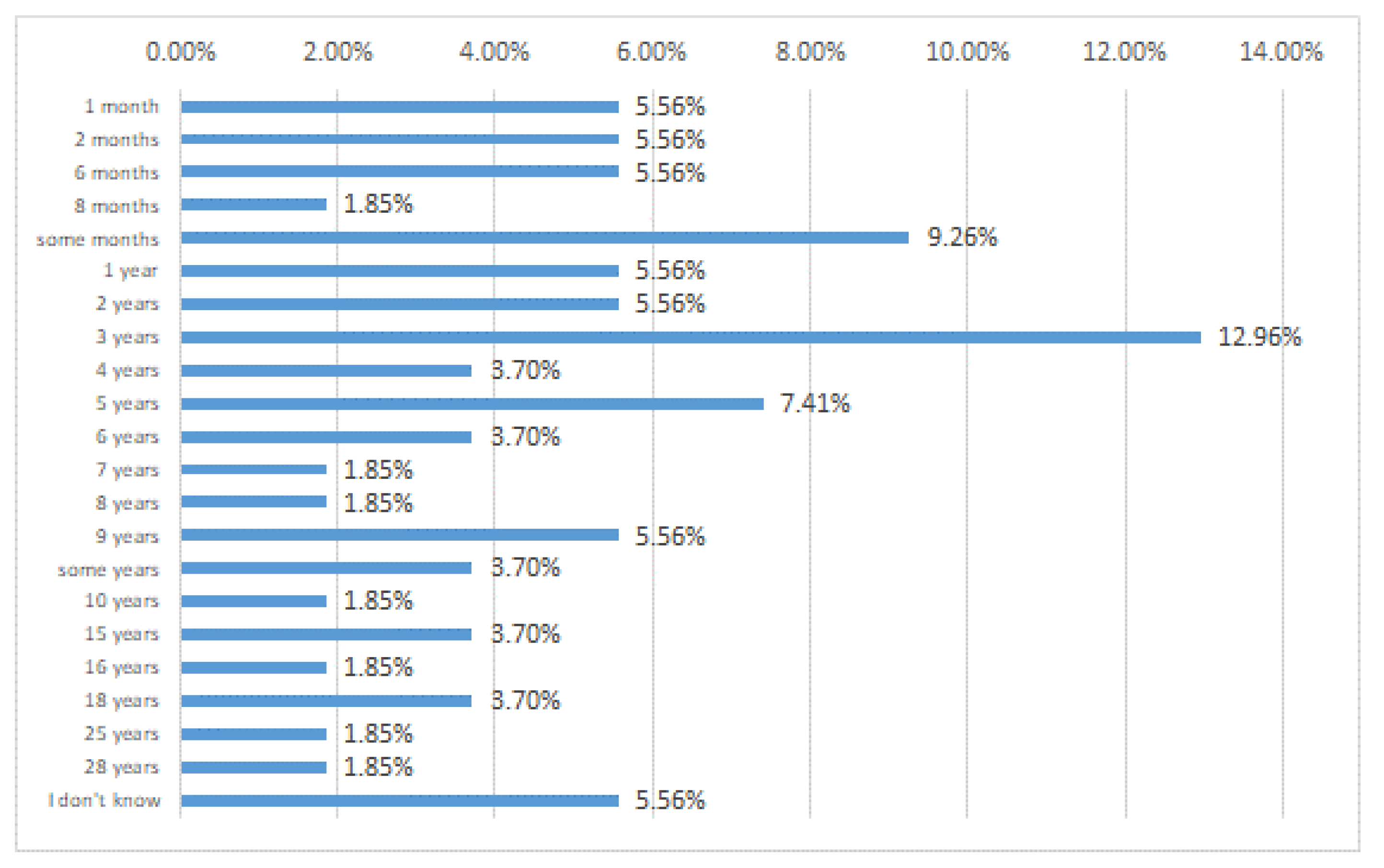
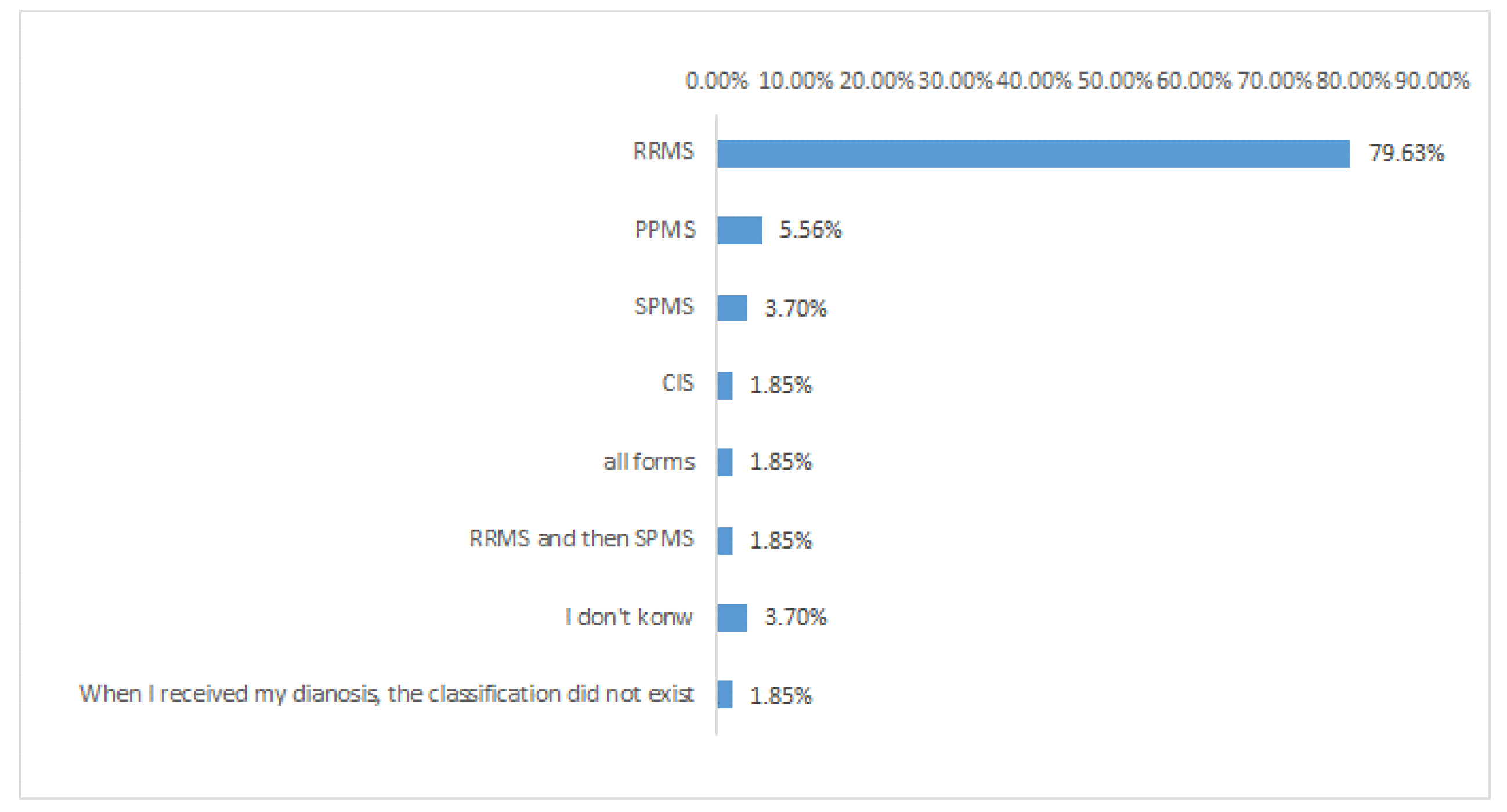
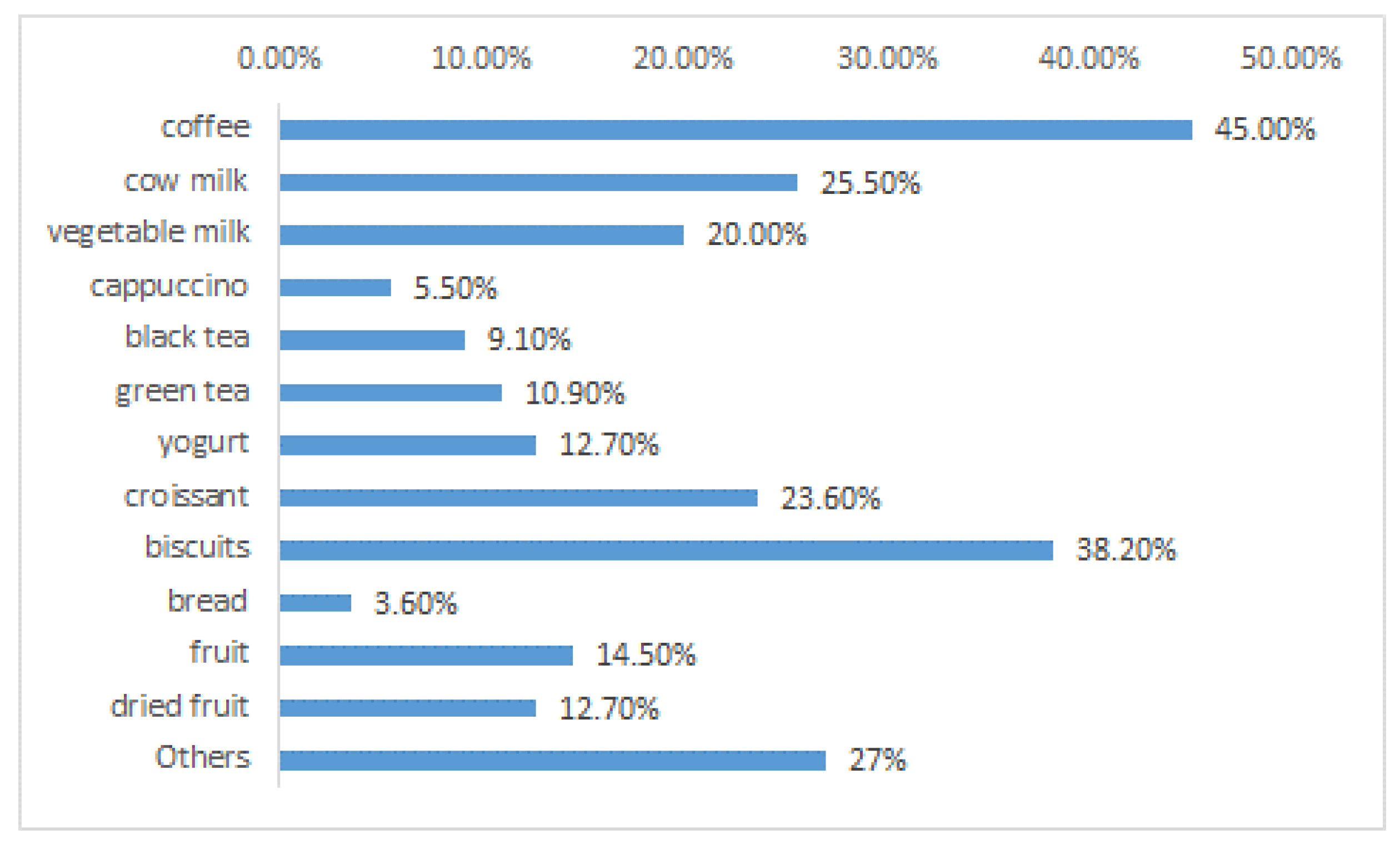
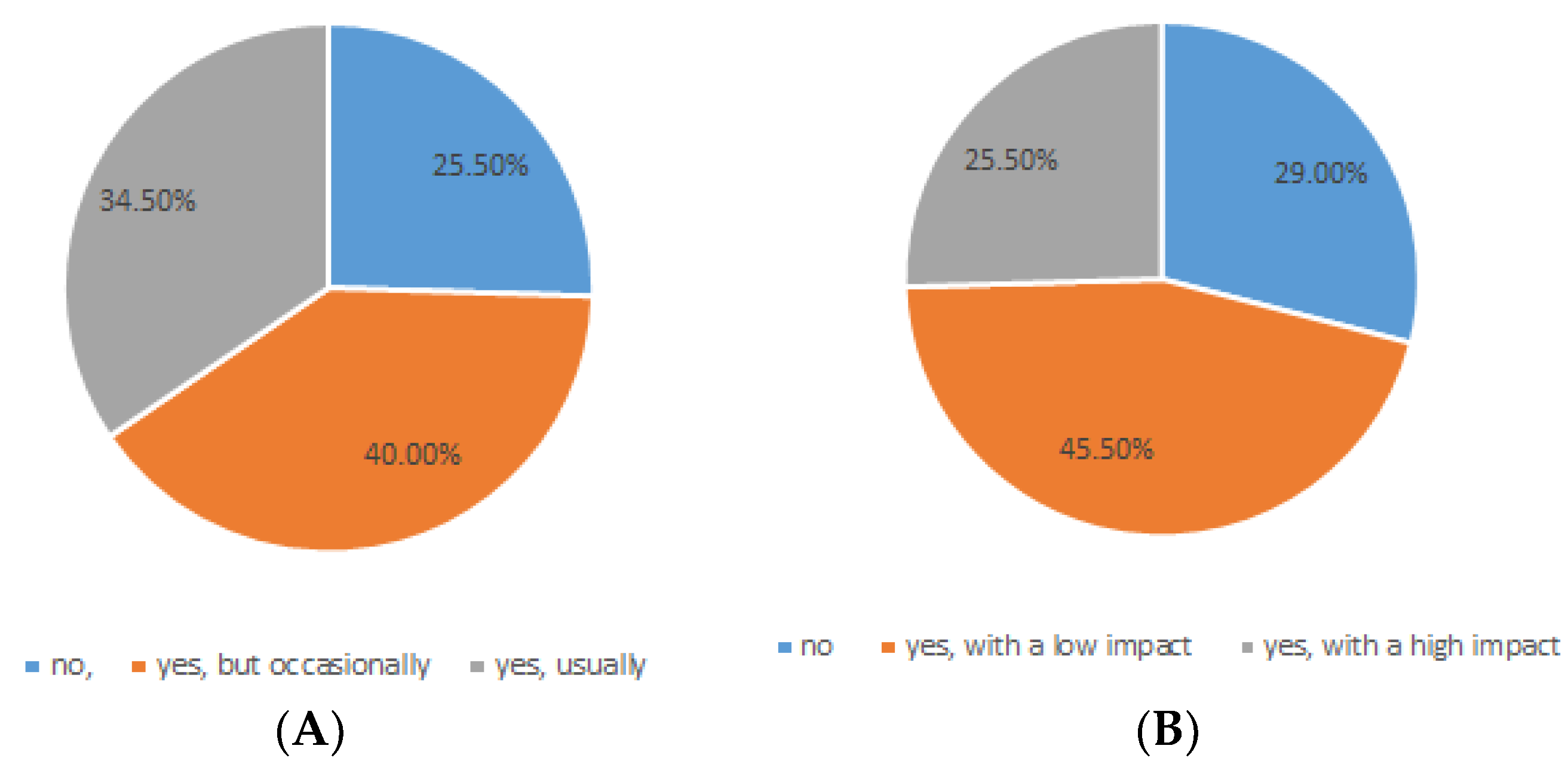
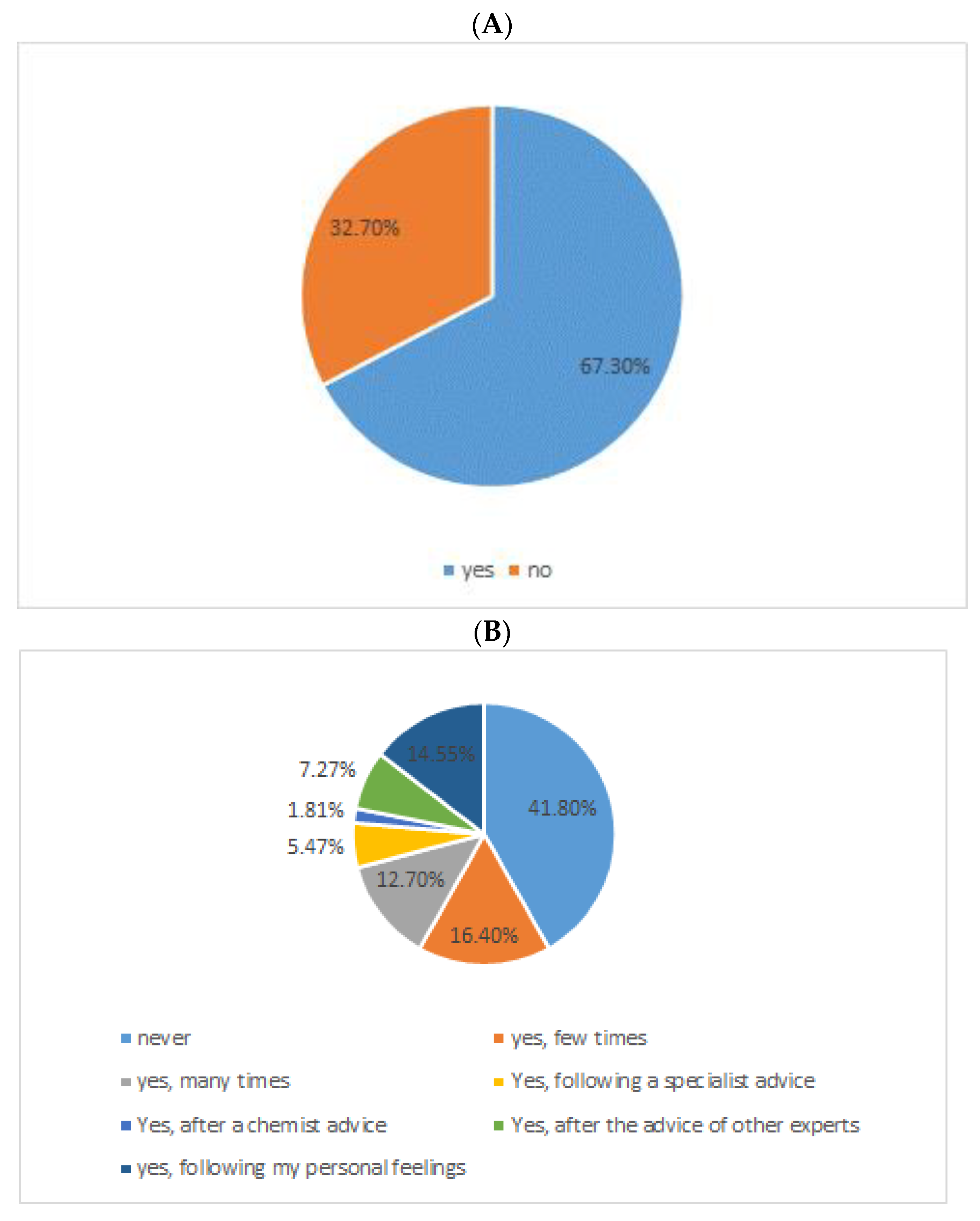
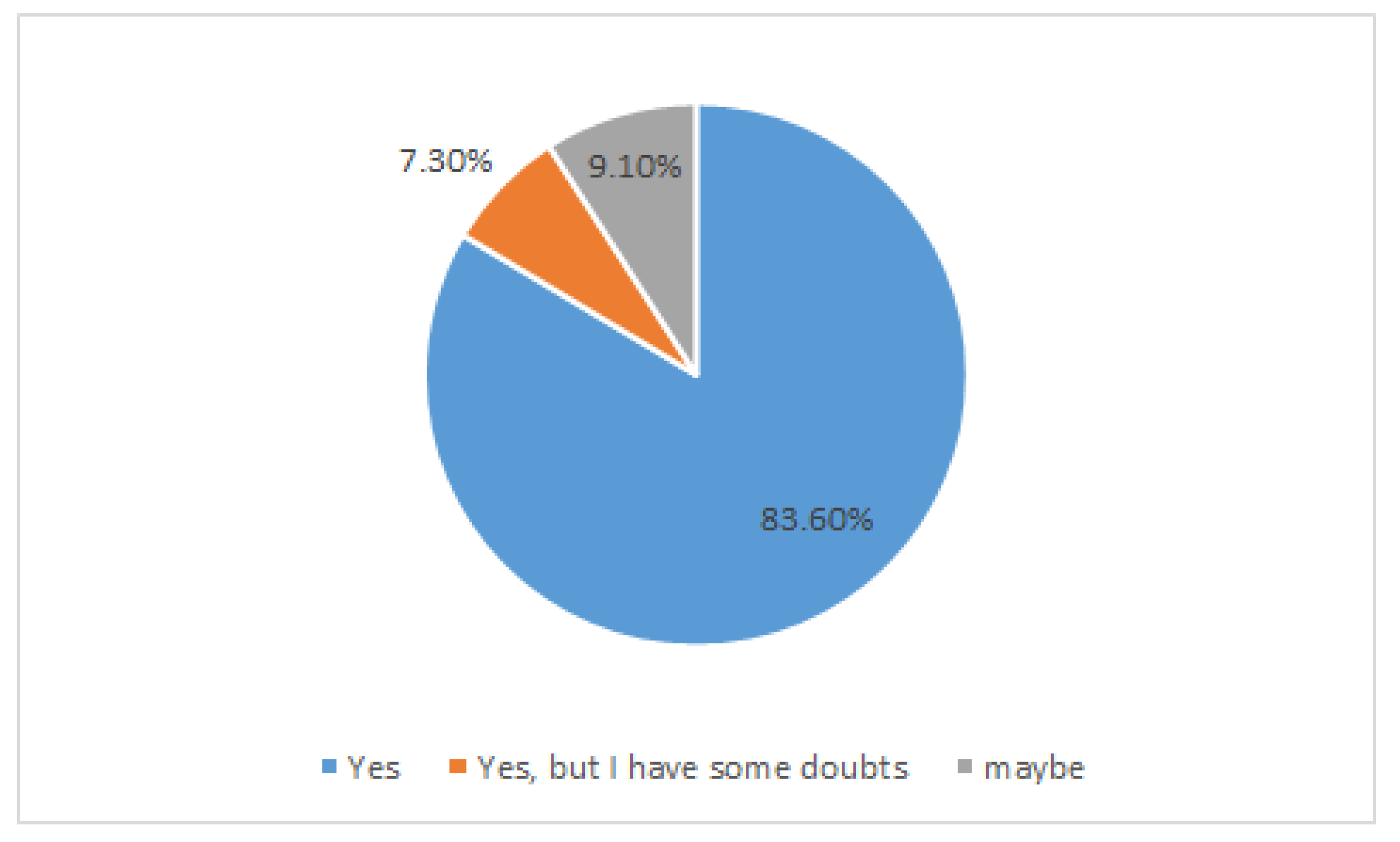
6. MS Patients, Abdominal Pains, and Probiotics: A Case Study
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Young, V.B. The role of the microbiome in human health and disease: An introduction for clinicians. BMJ 2017, 356, j831. [Google Scholar] [CrossRef] [PubMed]
- Forsythe, P.; Kunze, W.; Bienenstock, J. On communication between gut microbes and the brain. Curr. Opin. Gastroenterol. 2012, 28, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Kaskow, B.J.; Baecher-Allan, C. Effector T Cells in Multiple Sclerosis. Cold Spring Harb. Perspect. Med. 2018, 8, a029025. [Google Scholar] [CrossRef] [PubMed]
- Human Microbiome Project Consortium. Structure, function and diversity of the health human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef]
- Chu, H.; Khosraviindah, A.; Kusumawardhani, I.; Kwon, A.; Vasconcelos, A.; Cunha, L.; Mayer, A.; Shen, Y.; Wu, W.; Kambal, A.; et al. Gene-microbiota interactions contribute to the pathogenesis of inflammatory bowel disease. Science 2016, 352, 1116–1120. [Google Scholar] [CrossRef]
- Rizzetto, L.; Fava, F.; Tuohy, K.M.; Selmi, C. Connecting the immune system, systemic chronic inflammation and the gut microbiome: The role of sex. J. Autoimmun. 2018, 92, 12–34. [Google Scholar] [CrossRef]
- Mosele, J.I.; Macià, A.; Motilva, M.-J. Metabolic and Microbial Modulation of the Large Intestine Ecosystem by Non-Absorbed Diet Phenolic Compounds: A Review. Molecules 2015, 20, 17429–17468. [Google Scholar] [CrossRef]
- Fava, F.; Rizzetto, L.; Tuohy, K.M. Gut microbiota and health: Connecting actors across the metabolic system. 2019 Conference on ‘Targeted approaches to tackling current nutritional issues’ Symposium 2: Developing and using novel health and nutritional biomarkers. Proc. Nutr. Soc. 2019, 78, 177–188. [Google Scholar] [CrossRef]
- Bielik, V.; Kolisek, M. Bioaccessibility and Bioavailability of Minerals in Relation to a Healthy Gut Microbiome. Int. J. Mol. Sci. 2021, 22, 6803. [Google Scholar] [CrossRef]
- Rossi, M.; Amaretti, A.; Raimondi, S. Folate production by probiotic bacteria. Nutrients 2011, 3, 118–134. [Google Scholar] [CrossRef]
- Ostaff, M.J.; Stange, E.F.; Wehkamp, J. Antimicrobial peptides and gut microbiota in homeostasis and pathology. EMBO Mol. Med. 2013, 5, 1465–1483. [Google Scholar] [CrossRef]
- Holzer, P.; Farzi, A. Neuropeptides and the microbiota-gut-brain axis. Adv. Exp. Med. Biol. 2014, 817, 195–219. [Google Scholar]
- Cronin, O.; O’Sullivan, O.; Barton, W.; Cotter, P.D.; Molloy, M.G.; Shanahan, F. Gut microbiota: Implications for sports and exercise medicine. Br. J. Sports Med. 2017, 51, 700–701. [Google Scholar] [CrossRef]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef]
- Deaver, J.A.; Eum, S.Y.; Toborek, M. Circadian Disruption Changes Gut Microbiome Taxa and Functional Gene Composition. Front. Microbiol. 2018, 9, 737. [Google Scholar] [CrossRef]
- Gupta, V.K.; Paul, S.; Dutta, C. Geography, ethnicity or subsistence-specific variations in human microbiome composition and diversity. Front. Microbiol. 2017, 8, 1162. [Google Scholar] [CrossRef]
- Brooks, A.W.; Priya, S.; Blekhman, R.; Bordenstein, S.R. Gut microbiota diversity across ethnicities in the United States. PLoS Biol. 2018, 16, e2006842. [Google Scholar] [CrossRef]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef]
- Schnorr, S.L.; Candela, M.; Rampelli, S.; Centanni, M.; Consolandi, C.; Basaglia, G.; Turroni, S.; Biagi, E.; Peano, C.; Severgnini, M.; et al. Gut microbiome of the Hadza hunter-gatherers. Nat. Commun. 2014, 5, 3654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syromyatnikov, M.; Nesterova, E.; Gladkikh, M.; Smironova, Y.; Fryaznova, M.; Popov, V. Characteristics of the gut bacterial composition in people of different nationalities and religions. Microorganisms 2022, 10, 1866. [Google Scholar] [CrossRef] [PubMed]
- Ang, Q.Y.; Alba, D.L.; Upadhyay, V.; Bisanz, J.E.; Cai, J.; Lee, H.L.; Barajas, E.; Wei, G.; Noecker, C.; Patterson, A.D.; et al. The East Asian gut microbiome is distinct from colocalized White subjects and connected to metabolic health. eLife 2021, 10, e70349. [Google Scholar] [CrossRef] [PubMed]
- Takagi, T.; Inoue, R.; Oshima, A.; Sakazume, H.; Ogawa, K.; Tominaga, T.; Mihara, Y.; Sugaya, T.; Mizushima, K.; Uchiyama, K.; et al. Typing of the gut microbiota mommunity in Japanese subjects. Microorganisms 2022, 10, 664. [Google Scholar] [CrossRef]
- Chauhan, N.S.; Pandey, R.; Mondal, A.K.; Gupta, S.; Verma, M.K.; Jain, S.; Ahmed, V.; Patil, R.; Agarwal, D.; Girase, B.; et al. Western Indian Rural Gut Microbial Diversity in Extreme Prakriti Endo-Phenotypes Reveals Signature Microbes. Front. Microbiol. 2018, 9, 118. [Google Scholar] [CrossRef]
- Li, J.; Fu, R.; Yang, Y.; Horz, H.P.; Guan, Y.; Lu, Y.; Lou, H.; Tian, L.; Zheng, S.; Liu, H.; et al. A metagenomic approach to dissect the genetic composition of enterotypes in Han Chinese and two Muslim groups. Syst. Appl. Microbiol. 2018, 41, 1–12. [Google Scholar] [CrossRef]
- Ozkul, C.; Yalınay, M.; Karakan, T. Islamic fasting leads to an increased abundance of Akkermansia muciniphila and Bacteroides fragilis group: A preliminary study on intermittent fasting. Turk. J. Gastroenterol. 2019, 30, 1030–1035. [Google Scholar] [CrossRef]
- Karakan, T. Intermittent fasting and gut microbiota. Turk. J. Gastroenterol. 2019, 30, 1008. [Google Scholar] [CrossRef]
- Ozkul, C.; Yalinay, M.; Karakan, T. Structural changes in gut microbiome after Ramadan fasting: A pilot study. Benef. Microbes 2020, 11, 227–233. [Google Scholar] [CrossRef]
- Jia, W.; Zhen, J.; Liu, A.; Yuan, J.; Wu, X.; Zhao, P.; Zhao, L.; Li, X.; Liu, Q.; Huang, G.; et al. Long-term vegan meditationimproved human gut microbiota. Evid.-Based Complement. Altern. Med. 2020, 5, 9517897. [Google Scholar]
- Orlich, M.J.; Fraser, G.E. Vegetarian diets in the Adventist Health Study 2: A review of initial published findings. Am. J. Clin. Nutr. 2014, 100, 353S–358S. [Google Scholar] [CrossRef]
- Campaniello, D.; Corbo, M.R.; Sinigaglia, M.; Speranza, B.; Racioppo, A.; Altieri, C.; Bevilacqua, A. How diet and physical activity modulate gut microbiota: Evidence, and perspectives. Nutrients 2022, 14, 2456. [Google Scholar] [CrossRef]
- Ethnicity influences the gut microbiota of individuals sharing a geographical location: A cross-sectional study from a middle-income country. Sci. Rep. 2021, 11, 2618. [CrossRef]
- Sonnenburg, J.L.; Bäckhed, F. Diet-microbiota interactions as moderators of human metabolism. Nature 2016, 535, 56–64. [Google Scholar] [CrossRef]
- Statovci, D.; Aguilera, M.; MacSharry, J.; Melgar, S. The impact of western diet and nutrients on the microbiota and immune response at mucosal interfaces. Front. Immunol. 2017, 8, 838. [Google Scholar] [CrossRef]
- Wong, J.M.W. Gut microbiota and cardiometabolic outcomes: Influence of dietary patterns and their associated components. Am. J. Clin. Nutr. 2014, 100, 369–377. [Google Scholar] [CrossRef]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2016, 65, 1812–1821. [Google Scholar] [CrossRef]
- Wang, S.Z.; Yu, Y.J.; Adeli, K. Role of Gut Microbiota in Neuroendocrine Regulation of Carbohydrate and Lipid Metabolism via the Microbiota-Gut-Brain-Liver Axis. Microorganisms 2020, 8, 527. [Google Scholar] [CrossRef]
- Negi, S.; Das, D.K.; Pahari, S.; Nadeem, S.; Agrewala, J.N. Potential Role of Gut Microbiota in Induction and Regulation of Innate Immune Memory. Front. Immunol. 2019, 10, 2441. [Google Scholar] [CrossRef]
- Ahmed, H.; Leyrolle, Q.; Koistinen, V.; Kärkkäinen, O.; Layé, S.; Delzenne, N.; Hanhineva, K. Microbiota-derived metabolites as drivers of gut-brain communication. Gut Microbes 2022, 14, 2102878. [Google Scholar] [CrossRef]
- Haase, S.; Linker, R.A. Inflammation in multiple sclerosis. Therap. Adv. Neurol. Disord. 2021, 14, 17562864211007687. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Cai, X.; Ye, Y.; Wang, F.; Chen, F.; Zheng, C. The role of microbiota in infant health: From early life to adulthood. Front. Immunol. 2021, 12, 708472. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, X.; Zhao, C. Guillain-Barré Syndrome-Like Polyneuropathy Associated with Immune Checkpoint Inhibitors: A Systematic Review of 33 Cases. BioMed Res. Int. 2021, 2021, 9800488. [Google Scholar] [CrossRef] [PubMed]
- Maslowski, K.; Vieira, A.; Ng, A.; Kranich, J.; Sierro, F.; Yu, D.; Schilter, H.C.; Rolph, M.S.; Mackay, F.; Artis, D.; et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 2009, 461, 1282–1286. [Google Scholar] [CrossRef]
- Yoo, J.Y.; Groer, M.; Dutra, S.V.O.; Sarkar, A.; McSkimming, D.I. Gut Microbiota and Immune System Interactions. Microorganisms 2020, 8, 1587. [Google Scholar] [CrossRef]
- Compston, A.; Coles, A. Multiple sclerosis. Lancet 2008, 372, 1502–1517. [Google Scholar] [CrossRef]
- Riccio, P.; Rossano, R. Diet, Gut Microbiota, and Vitamins D + A in Multiple Sclerosis. Neurotherapeutics 2018, 15, 75–91. [Google Scholar] [CrossRef]
- Ghasemi, N.; Razavi, S.; Nikzad, E. Multiple sclerosis: Pathogenesis, symptoms, diagnoses, and cell based therapy. Cell J. 2017, 19, 1–10. [Google Scholar]
- Miller, D.H.; Hornabrook, R.W.; Dagger, J.; Fong, R. Class II HLA antigens in multiple sclerosis. J. Neurol. Neurosurg. Psychiatry 1989, 52, 575–577. [Google Scholar] [CrossRef]
- Sawcer, S.; Franklin, R.J.; Ban, M. Multiple sclerosis genetics. Lancet Neurol. 2014, 13, 700–709. [Google Scholar] [CrossRef]
- Küçükali, C.; Kürtüncü, M.; Çoban, A.; Çebi, M.; Tüzün, E. Epigenetics of multiple sclerosis: An update review. Neuromol. Med. 2015, 17, 83–96. [Google Scholar] [CrossRef]
- Huang, J.; Kockum, I.; Stridh, P. Trends in the environmental risks associated with earlier onset in multiple sclerosis. Mult. Scler. Relat. Disord. 2022, 68, 104250. [Google Scholar] [CrossRef]
- Hedstrom, A.K.; Katsoulis, M.; Hossjer, O.; Bomfim, I.L.; Oturai, A.; Sondergaard, H.B.; Sellebjerg, F.; Ullum, H.; Thorner, L.W.; Gustavsen, M.W.; et al. The interaction between smoking and HLA genes in multiple sclerosis: Replication and refinement. Eur. J. Epidemiol. 2017, 32, 909–919. [Google Scholar] [CrossRef] [Green Version]
- Odoardi, F.; Sie, C.; Streyl, K.; Ulaganathan, V.K.; Schlager, C.; Lodygin, D.; Heckelsmiller, K.; Nietfeld, W.; Ellwart, J.; Klinkert, W.E.; et al. T cells become licensed in the lung to enter the central nervous system. Nature 2012, 488, 675–679. [Google Scholar] [CrossRef]
- Ascherio, A.; Munger, K.L. Environmental risk factors for multiple sclerosis. Part II: Noninfectious factors. Ann. Neurol. 2007, 61, 504–513. [Google Scholar] [CrossRef]
- Cantorna, M.T.; Hayes, C.E.; DeLuca, H.F. 1,25-dihydroxyvitamin D3 reversibly blocks the progression of relapsing encephalomyelitis, a model of multiple sclerosis. Proc. Natl. Acad. Sci. USA 1996, 93, 7861–7864. [Google Scholar] [CrossRef]
- Cortese, M.; Riise, T.; Bjornevik, K.; Myhr, K.M. Multiple Sclerosis Conscript Service Database Study, G. Body size and physical exercise, and the risk of multiple sclerosis. Mult. Scler. 2018, 24, 270–278. [Google Scholar] [CrossRef]
- Hardy, D.; Chitnis, T.; Waubant, E.; Banwell, B. Preventing Multiple Sclerosis: The Pediatric Perspective. Front. Neurol. 2022, 13, 802380. [Google Scholar] [CrossRef]
- Januel, E.; Dessimond, B.; Colette, A.; Annesi-Maesano, I.; Stankoff, B. Fine Particulate Matter Related to Multiple Sclerosis Relapse in Young Patient. Front. Neurol. 2021, 12, 651084. [Google Scholar] [CrossRef]
- Lavery, A.M.; Collins, B.N.; Waldman, A.T.; Hart, C.N.; Bar-Or, A.; Marrie, R.A.; Arnold, D.; O’Mahony, J.; Banwell, B. The contribution of secondhand tobacco smoke exposure to pediatric multiple sclerosis risk. Mult. Scler. 2018, 25, 515–522. [Google Scholar] [CrossRef]
- Mirza, A.; Forbes, J.B.; Zhu, F.; Bernstein, C.N.; Van Domselaar, G.; Graham, M.; Waubant, E.; Tremlett, H. The multiple sclerosis gut microbiota: A systematic review. Mult. Scler. Relat. Disord. 2020, 37, 101427. [Google Scholar] [CrossRef] [PubMed]
- Boziki, M.K.; Kesidou, E.; Theotokis, P.; Mentis, A.A.; Karafoulidou, E.; Melnikov, M.; Sviridova, A.; Rogovski, V.; Boyko, A.; Grigoriadis, N. Microbiome in Multiple Sclerosis; Where Are We, What We Know and Do Not Know. Brain Sci. 2020, 10, 234. [Google Scholar] [CrossRef]
- Rea, K.; Dinan, T.G.; Cryan, J.F. The microbiome: A key regulator of stress and neuroinflammation. Neurobiol. Stress 2016, 4, 23–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daisuke, N.; Miyake, S. Gut dysbiosis and multiple sclerosis. Clin. Immunol. 2022, 235, 108380. [Google Scholar]
- Berer, K.; Krishnamoorthy, G. Microbial view of central nervous system autoimmunity. FEBS Lett. 2014, 588, 4207–4213. [Google Scholar] [CrossRef]
- Wekerle, H.; Hohlfeld, R. Gut Microbiota in Multiple Sclerosis: A Bioreactor Driving Brain Autoimmunity. In Translational Neuroimmunology in Multiple Sclerosis. From Disease Mechanisms to Clinical Applications; Elsevier: Amsterdam, The Netherlands, 2016; Volume 9, pp. 113–125. [Google Scholar]
- Miyahuchi, E.; Shimokawa, C.; Steimle, A.; Desai, M.S.; Ohno, H. The impact of gut microbiome on extra-intestinal autoimmune diseases. Nat. Rev. Immunol. 2023, 23, 9–23. [Google Scholar] [CrossRef]
- Shi, P.; Qu, H.; Nian, D.; Chen, Y.; Liu, X.; Li, Q.; Wang, C.; Ye, M.; Ma, B. Treatment of Guillain-Barré syndrome with Bifidobacterium infantis through regulation of T helper cells subsets. Int. Immunopharmacol. 2018, 61, 290–296. [Google Scholar] [CrossRef]
- Li, S.; Jin, T.; Zhang, H.L.; Yu, H.; Meng, F.; Quezada, H.C.; Zhu, J. Circulating Th17, Th22, and Th1 Cells Are Elevated in the Guillain-Barré Syndrome and Downregulated by IVIg Treatments. Mediat. Inflamm. 2014, 4, 1–10. [Google Scholar] [CrossRef]
- Etemadifar, M.; Roomizadeh, P.; Abtahi, S.H.; Sajjadi, S.; Abedini, A.; Golabbakhsh, A.; Fereidan-Esfahani, M.; Akbari, M. Linkage of multiple sclerosis and guillain-barre syndrome: A population-based survey in Isfahan, Iran. Autoimmune Dis. 2012, 2012, 232139. [Google Scholar] [CrossRef]
- Berer, K.; Gerdes, L.A.; Cekanaviciute, E.; Jiac, X.; Xiao, L.; Xia, Z.; Liud, C.; Klotze, L.; Staufferf, U.; Baranzinic, S.E.; et al. Gut microbiota from multiple sclerosis patients enables spontaneous autoimmune encephalomyelitis in mice. Proc. Natl. Acad. Sci. USA 2017, 114, 10719–10724. [Google Scholar] [CrossRef]
- Cekanaviciute, E.; Yoob, B.B.; Runiaa, T.F.; Debelius, J.W.; Singh, S.; Nelson, C.A.; Kanner, R.; Bencosme, Y.; Lee, Y.K.; Hauser, S.L.; et al. Gut bacteria from multiple sclerosis patients modulate human T cells and exacerbate symptoms in mouse models. Proc. Natl. Acad. Sci. USA 2017, 114, 10713–10718. [Google Scholar] [CrossRef]
- Calvo- Barreiro, L.; Eixarch, H.A.; Ponce-Alonso, M.; Castillo, M.; Lebrón-Galán, R.; Mestre, L.; Guaza, C.; Clemente, D.; Del Campo, R.; Montalban, X.; et al. A commercial probiotic induces tolerogenic and reduces pathogenic responses in experimental autoimmune encephalomyelitis. Cells 2020, 9, 906. [Google Scholar] [CrossRef]
- He, B.; Hoanh, T.K.; Tian, X.; Taylor, C.M.; Blanchard, E.; Luo, M.; Bhattacharjee, M.B.; Freeborn, J.; Park, S.; Couturier, J.; et al. Lactobacillus reuteri reduces the severity of experimental autoimmune encephalomyelitis in mice by modulating gut microbiota. Front. Immunol. 2019, 10, 385. [Google Scholar] [CrossRef]
- Kwon, H.K.; Kim, G.-C.; Kim, Y.; Hwang, W.; Jash, A.; Sahoo, A.; Kim, J.-E.; Nam, J.H.; Im, S.-H. Amelioration of experimental autoimmune encephalomyelitis by probiotic mixture is mediated by a shift in T helper cell immune response. Clin. Immunol. 2013, 146, 217–227. [Google Scholar] [CrossRef]
- Makkawi, S.; Camara- Lemarroy, C.; Metz, L. Fecal microbiota transplantation associated with 10 years of stability in a patient with SPMS. Neurol. Neuroimmunol. Neuroinflamm. 2018, 5, e459. [Google Scholar] [CrossRef]
- Borody, T.; Leis, S.; Campbell, J.; Margaux, T.; Anna, N. Fecal microbiota transplantation (FMT) in multiple sclerosis (MS). Am. J. Gastoenterol. 2011, 106, S352. [Google Scholar] [CrossRef]
- Chen, T.; Noto, D.; Hoshino, Y.; Mizuno, M.; Miyake, S. Butyrate suppresses demyelination and enhances remyelination. J. Neuroinflamm. 2019, 16, 165. [Google Scholar] [CrossRef]
- Paul, A.; Comabella, M.; Gandhi, R. Biomarkets in multiple sclerosis. Cold. Spring Harb. Perspect. Med. 2019, 9, a029058. [Google Scholar] [CrossRef]
- Lorefice, L.; Pitzalis, M.; Murgia, F.; Fenu, G.; Atzori, L.; Cocco, E. Omics approaches to understanding the efficacy and safety of disease-modifying treatments in multiple sclerosis. Front. Genet. 2023, 14, 1076421. [Google Scholar]
- Farah, M.; Haraty, H.; Salame, Z.; Fares, Y.; Ojcius, D.M.; Sadier, N.S. Salivary biomarkers for the diagnosis and monitoring of neurological disease. Biomed. J. 2018, 41, 63–87. [Google Scholar] [CrossRef]
- Lotan, I.; Ganelin-Cohen, E.; Tartakovsky, E.; Khasminsky, V.; Hellmann, M.A.; Steiner, I.; Ben-Zvi, I.; Livneh, A.; Golderman, S.; Kaplan, B. Saliva immunoglobulin free light chain analysis for monitoring disease activity and response to treatment in multiple sclerosis. Mult. Scler. Relat. Disord. 2020, 44, 102339. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.A.; Kang, C.-K.; Kim, E.-M.; Kim, A.R.; Kim, A. Proteomics of early detection of non-muscle invasive bladder cancer: Clinically useful urine protein biomarkers. Life 2022, 12, 395. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Altieri, C.; Speranza, B.; Corbo, M.R.; Sinigaglia, M.; Bevilacqua, A. Gut-Microbiota, and Multiple Sclerosis: Background, Evidence, and Perspectives. Nutrients 2023, 15, 942. https://doi.org/10.3390/nu15040942
Altieri C, Speranza B, Corbo MR, Sinigaglia M, Bevilacqua A. Gut-Microbiota, and Multiple Sclerosis: Background, Evidence, and Perspectives. Nutrients. 2023; 15(4):942. https://doi.org/10.3390/nu15040942
Chicago/Turabian StyleAltieri, Clelia, Barbara Speranza, Maria Rosaria Corbo, Milena Sinigaglia, and Antonio Bevilacqua. 2023. "Gut-Microbiota, and Multiple Sclerosis: Background, Evidence, and Perspectives" Nutrients 15, no. 4: 942. https://doi.org/10.3390/nu15040942
APA StyleAltieri, C., Speranza, B., Corbo, M. R., Sinigaglia, M., & Bevilacqua, A. (2023). Gut-Microbiota, and Multiple Sclerosis: Background, Evidence, and Perspectives. Nutrients, 15(4), 942. https://doi.org/10.3390/nu15040942