
Fermented Lettuce Extract Containing Nitric Oxide Metabolites Attenuates Inflammatory Parameters in Model Mice and in Human Fibroblast-Like Synoviocytes

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of FLE
2.2. Animal Care
2.3. Generation of CIA Mice and Experimental Intervention
2.4. Clinical Assessment of Arthritis
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. IHC
2.7. Histochemical Analysis
2.8. Micro-Computed Tomography (CT)
2.9. In Vivo Safety Evaluation
2.10. Cell Culture
2.11. Cell Viability Assay
2.12. Cell Proliferation Assay
2.13. Reverse-Transcriptase PCR (RT-PCR)
2.14. Western Blotting
2.15. Cell Migration Assay
2.16. Detection of Autophagosome Formation
2.17. Statistical Analysis
3. Results
3.1. FLE Treatment Reduced RA Severity in CIA Mice
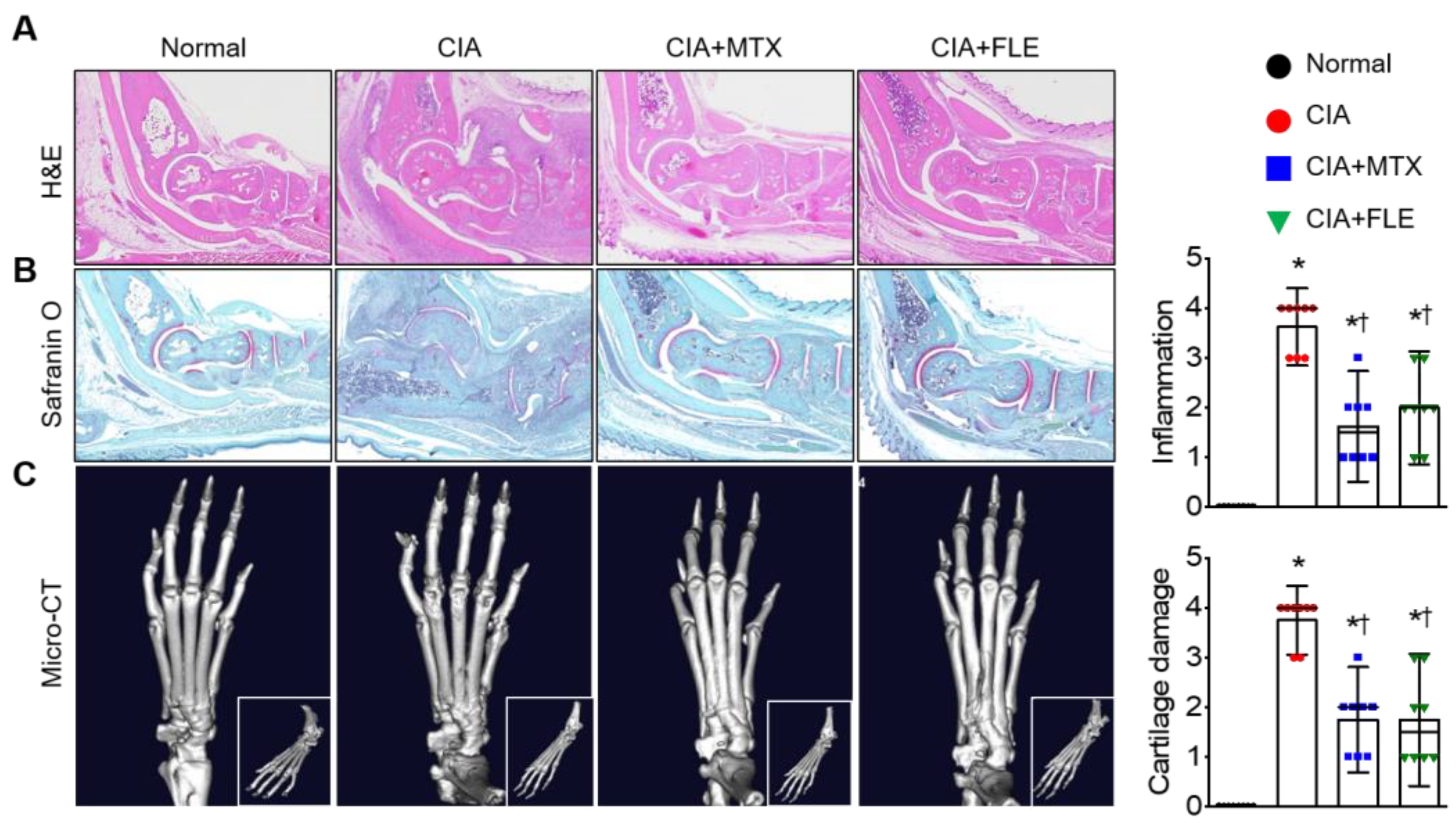
3.2. Administering FLE to CIA Mice Alleviated Histological Signs of RA
3.3. Safety Assessment of FLE in Mice
3.4. FLE Attenuated TGF-β/Smad Mediated Migration in MH7A Cells
3.5. FLE Induces Autophagosome Formation via Inhibiting Degradation in MH7A Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smolen, J.S.; Aletaha, D.; McInnes, I.B. Rheumatoid arthritis. Lancet 2016, 388, 2023–2038. [Google Scholar] [CrossRef]
- Xie, C.; Jiang, J.; Liu, J.; Yuan, G.; Zhao, Z. Triptolide suppresses human synoviocyte MH7A cells mobility and maintains redox balance by inhibiting autophagy. Biomed Pharm. 2019, 115, 108911. [Google Scholar] [CrossRef] [PubMed]
- Woś, I.; Tabarkiewicz, J. Effect of interleukin-6, -17, -21, -22, and -23 and STAT3 on signal transduction pathways and their inhibition in autoimmune arthritis. Immunol. Res. 2021, 69, 26–42. [Google Scholar] [CrossRef] [PubMed]
- Bartok, B.; Firestein, G.S. Fibroblast-like synoviocytes: Key effector cells in rheumatoid arthritis. Immunol. Rev. 2010, 233, 233–255. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo-Gil, E.; Galindo-Izquierdo, M. Role of transforming growth factor-beta (TGF) beta in the physiopathology of rheumatoid arthritis. Reum. Clin. 2014, 10, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Zhao, J.; Lou, A.; Huang, Q.; OuYang, Q.; Zhu, J.; Fan, M.; He, Y.; Ren, H.; Yang, M. Transforming growth factor β1 promotes fibroblast-like synoviocytes migration and invasion via TGF-β1/Smad signaling in rheumatoid arthritis. Mol. Cell. Biochem. 2019, 459, 141–150. [Google Scholar] [CrossRef]
- Walton, K.L.; Johnson, K.E.; Harrison, C.A. Targeting TGF-β mediated SMAD signaling for the prevention of fibrosis. Front. Pharmacol. 2017, 8, 461. [Google Scholar] [CrossRef] [Green Version]
- Ardoin, S.P.; Pisetsky, D.S. The role of cell death in the pathogenesis of autoimmune disease: HMGB1 and microparticles as intercellular mediators of inflammation. Mod. Rheumatol. 2008, 18, 319–326. [Google Scholar] [CrossRef]
- Zhao, J.; Jiang, P.; Guo, S.; Schrodi, S.J.; He, D. Apoptosis, Autophagy, NETosis, Necroptosis, and Pyroptosis Mediated Programmed Cell Death as Targets for Innovative Therapy in Rheumatoid Arthritis. Front. Immunol. 2021, 12, 809806. [Google Scholar] [CrossRef]
- Korb, A.; Pavenstadt, H.; Pap, T. Cell death in rheumatoid arthritis. Apoptosis 2009, 14, 447–454. [Google Scholar] [CrossRef]
- Parzych, K.R.; Klionsky, D.J. An overview of autophagy: Morphology, mechanism, and regulation. Antioxid. Redox Signal. 2014, 20, 460–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dikic, I.; Elazar, Z. Mechanism and medical implications of mammalian autophagy. Nat. Rev. Mol. Cell. Biol. 2018, 19, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Mizushima, N. LC3- and p62-based biochemical methods for the analysis of autophagy progression in mammalian cells. Methods 2015, 75, 13–18. [Google Scholar] [CrossRef]
- Tanida, I.; Ueno, T.; Kominami, E. LC3 and Autophagy. Methods Mol. Biol. 2008, 445, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Lamark, T.; Svenning, S.; Johansen, T. Regulation of selective autophagy: The p62/SQSTM1 paradigm. Essays Biochem. 2017, 61, 609–624. [Google Scholar] [CrossRef]
- Yang, X.; Gil, M.I.; Yang, Q.; Tomás-Barberán, F.A. Bioactive compounds in lettuce: Highlighting the benefits to human health and impacts of preharvest and postharvest practices. Compr. Rev. Food Sci. Food Saf. 2022, 21, 4–45. [Google Scholar] [CrossRef]
- Sayyah, M.; Hadidi, N.; Kamalinejad, M. Analgesic and anti-inflammatory activity of Lactuca sativa seed extract in rats. J. Ethnopharmacol. 2004, 92, 325–329. [Google Scholar] [CrossRef]
- Nicolle, C.; Cardinault, N.; Gueux, E.; Jaffrelo, L.; Rock, E.; Mazur, A.; Amouroux, P.; Rémésy, C. Health effect of vegetable-based diet: Lettuce consumption improves cholesterol metabolism and antioxidant status in the rat. Clin. Nutr. 2004, 23, 605–614. [Google Scholar] [CrossRef]
- Okada, Y.; Okada, M. Protective effects of plant seed extracts against amyloid β-induced neurotoxicity in cultured hippocampal neurons. J. Pharm. Bioallied Sci. 2013, 5, 141. [Google Scholar] [CrossRef]
- Koyama, M.; Ogasawara, Y.; Endou, K.; Akano, H.; Nakajima, T.; Aoyama, T.; Nakamura, K. Fermentation-induced changes in the concentrations of organic acids, amino acids, sugars, and minerals and superoxide dismutase-like activity in tomato vinegar. Int. J. Food Prop. 2017, 20, 888–898. [Google Scholar] [CrossRef] [Green Version]
- Han, A.L.; Lee, H.K.; Chon, H.S.; Pae, H.O.; Kim, M.S.; Shin, Y.I.; Kim, S. Evaluation of the Effectiveness of Fermented Soybean-Lettuce Powder for Improving Menopausal Symptoms. Nutrients 2022, 14, 2878. [Google Scholar] [CrossRef] [PubMed]
- Baik, J.S.; Min, J.H.; Ju, S.M.; Ahn, J.H.; Ko, S.H.; Chon, H.S.; Kim, M.S.; Shin, Y.I. Effects of Fermented Garlic Extract Containing Nitric Oxide Metabolites on Blood Flow in Healthy Participants: A Randomized Controlled Trial. Nutrients 2022, 14, 5238. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.Y.; Kim, E.; Zhang, M.; Lee, Y.-S.; Ji, B.; Lee, S.-H.; Cheong, Y.E.; Yun, S.-I.; Kim, Y.-S.; Kim, K.H.; et al. Antidiabetic Effect of Noodles Containing Fermented Lettuce Extracts. Metabolites 2021, 11, 520. [Google Scholar] [CrossRef] [PubMed]
- Holmdahl, R.; Jansson, L.; Larsson, E.; Rubin, K.; Klareskog, L. Homologous type II collagen induces chronic and progressive arthritis in mice. Arthritis Rheum. Off. J. Am. Coll. Rheumatol. 1986, 29, 106–113. [Google Scholar] [CrossRef]
- Wilder, R.L. Hormones and autoimmunity: Animal models of arthritis. Baillieres Clin. Rheumatol. 1996, 10, 259–271. [Google Scholar] [CrossRef]
- Lee, Y.-K.; Choi, K.-H.; Kwak, H.-S.; Chang, Y.H. The preventive effects of nanopowdered red ginseng on collagen-induced arthritic mice. Int. J. Food Sci. Nutr. 2018, 69, 308–317. [Google Scholar] [CrossRef]
- Brand, D.D.; Latham, K.A.; Rosloniec, E.F. Collagen-induced arthritis. Nat. Protoc. 2007, 2, 1269–1275. [Google Scholar] [CrossRef]
- Alqahtani, M.S.; Kazi, M.; Alsenaidy, M.A.; Ahmad, M.Z. Advances in oral drug delivery. Front. Pharmacol. 2021, 12, 618411. [Google Scholar] [CrossRef]
- Kim, E.-K.; Lee, S.H.; Lee, S.-Y.; Kim, J.-K.; Jhun, J.Y.; Na, H.S.; Kim, S.-Y.; Choi, J.Y.; Yang, C.W.; Park, S.-H.; et al. Metformin ameliorates experimental-obesity-associated autoimmune arthritis by inducing FGF21 expression and brown adipocyte differentiation. Exp. Mol. Med. 2018, 50, e432. [Google Scholar] [CrossRef] [Green Version]
- Ji, M.; Ryu, H.J.; Baek, H.-M.; Shin, D.M.; Hong, J.H. Dynamic synovial fibroblasts are modulated by NBCn1 as a potential target in rheumatoid arthritis. Exp. Mol. Med. 2022, 54, 503–517. [Google Scholar] [CrossRef]
- Ryu, J.H.; Kim, Y.; Kim, M.J.; Park, J.; Kim, J.W.; Park, H.S.; Kim, Y.S.; Shin, H.K.; Shin, Y.-I. Membrane-Free Stem Cell Extract Enhances Blood–Brain Barrier Integrity by Suppressing NF-κB-Mediated Activation of NLRP3 Inflammasome in Mice with Ischemic Stroke. Life 2022, 12, 503. [Google Scholar] [CrossRef] [PubMed]
- Riss, T.L.; Moravec, R.A.; Niles, A.L.; Duellman, S.; Benink, H.A.; Worzella, T.J.; Minor, L. Cell Viability Assays. In Assay Guidance Manual; Markossian, S., Grossman, A., Brimacombe, K., Arkin, M., Auld, D., Austin, C., Baell, J., Chung, T.D.Y., Coussens, N.P., Dahlin, J.L., et al., Eds.; Eli Lilly & Company and the National Center for Advancing Translational Sciences: Bethesda, MD, USA, 2004. [Google Scholar]
- Luo, C.; Lu, Z.; Chen, Y.; Chen, X.; Liu, N.; Chen, J.; Dong, S. MicroRNA-640 promotes cell proliferation and adhesion in glioblastoma by targeting Slit guidance ligand 1. Oncol. Lett. 2021, 21, 161. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Kim, Y.; Kim, Y.M.; Jung, J.; Kim, T.; Lee, S.-Y.; Shin, Y.-I.; Ryu, J.H. Anti-oxidant and anti-inflammatory effects of aquatic exercise in allergic airway inflammation in mice. Front. Physiol. 2019, 10, 1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pijuan, J.; Barceló, C.; Moreno, D.F.; Maiques, O.; Sisó, P.; Marti, R.M.; Macià, A.; Panosa, A. In vitro Cell Migration, Invasion, and Adhesion Assays: From Cell Imaging to Data Analysis. Front. Cell Dev. Biol. 2019, 7, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, L.L.-Y.; Shen, D.; Wilkinson, A.R.; Patton, W.; Lai, N.; Chan, E.; Kuksin, D.; Lin, B.; Qiu, J. A novel image-based cytometry method for autophagy detection in living cells. Autophagy 2012, 8, 1371–1382. [Google Scholar] [CrossRef] [Green Version]
- Lefèvre, S.; Knedla, A.; Tennie, C.; Kampmann, A.; Wunrau, C.; Dinser, R.; Korb, A.; Schnäker, E.-M.; Tarner, I.H.; Robbins, P.D. Synovial fibroblasts spread rheumatoid arthritis to unaffected joints. Nat. Med. 2009, 15, 1414–1420. [Google Scholar] [CrossRef] [Green Version]
- Bullock, J.; Rizvi, S.A.A.; Saleh, A.M.; Ahmed, S.S.; Do, D.P.; Ansari, R.A.; Ahmed, J. Rheumatoid Arthritis: A Brief Overview of the Treatment. Med. Princ. Pr. 2018, 27, 501–507. [Google Scholar] [CrossRef]
- Burmester, G.R.; Pope, J.E. Novel treatment strategies in rheumatoid arthritis. Lancet 2017, 389, 2338–2348. [Google Scholar] [CrossRef]
- Demoruelle, M.K.; Deane, K.D. Treatment strategies in early rheumatoid arthritis and prevention of rheumatoid arthritis. Curr. Rheumatol. Rep. 2012, 14, 472–480. [Google Scholar] [CrossRef]
- Lin, Y.J.; Anzaghe, M.; Schülke, S. Update on the Pathomechanism, Diagnosis, and Treatment Options for Rheumatoid Arthritis. Cells 2020, 9, 880. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Kim, Y.R.; Min, Y.; Zhao, Y.; Do, K.; Son, Y.O. Natural Plant Extracts and Compounds for Rheumatoid Arthritis Therapy. Medicina 2021, 57, 266. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Z.; Zhang, S.N. Herbal compounds for rheumatoid arthritis: Literatures review and cheminformatics prediction. Phytother. Res. 2020, 34, 51–66. [Google Scholar] [CrossRef] [PubMed]
- Deligiannidou, G.-E.; Gougoula, V.; Bezirtzoglou, E.; Kontogiorgis, C.; Constantinides, T.K. The Role of Natural Products in Rheumatoid Arthritis: Current Knowledge of Basic In Vitro and In Vivo Research. Antioxidants 2021, 10, 599. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Gangwar, V.; Garg, A.; Lomore, K.; Korla, K.; Bhat, S.S.; Rao, R.P.; Rafiq, M.; Kumawath, R.; Uddagiri, B.V.; Kareenhalli, V.V. Immunomodulatory Effects of a Concoction of Natural Bioactive Compounds-Mechanistic Insights. Biomedicines 2021, 9, 1522. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, T.G.; Mahajan, K.; Dhiman, S. Medicinal plants used against various inflammatory biomarkers for the management of rheumatoid arthritis. J. Pharm. Pharm. 2020, 72, 1306–1327. [Google Scholar] [CrossRef] [PubMed]
- Schuster, R.; Rockel, J.S.; Kapoor, M.; Hinz, B. The inflammatory speech of fibroblasts. Immunol. Rev. 2021, 302, 126–146. [Google Scholar] [CrossRef]
- Ma, J.-D.; Jing, J.; Wang, J.-W.; Yan, T.; Li, Q.-H.; Mo, Y.-Q.; Zheng, D.-H.; Gao, J.-L.; Nguyen, K.-A.; Dai, L. A novel function of artesunate on inhibiting migration and invasion of fibroblast-like synoviocytes from rheumatoid arthritis patients. Arthritis Res. Ther. 2019, 21, 1–14. [Google Scholar] [CrossRef] [Green Version]
- You, S.; Koh, J.H.; Leng, L.; Kim, W.U.; Bucala, R. The Tumor-Like Phenotype of Rheumatoid Synovium: Molecular Profiling and Prospects for Precision Medicine. Arthritis Rheumatol. 2018, 70, 637–652. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.J.; Adamopoulos, I.E. Autophagy and autoimmunity. Clin. Immunol. 2017, 176, 55–62. [Google Scholar] [CrossRef] [Green Version]
- Shin, Y.-J.; Han, S.-H.; Kim, D.-S.; Lee, G.-H.; Yoo, W.-H.; Kang, Y.-M.; Choi, J.-Y.; Lee, Y.C.; Park, S.J.; Jeong, S.-K.; et al. Autophagy induction and CHOP under-expression promotes survival of fibroblasts from rheumatoid arthritis patients under endoplasmic reticulum stress. Arthritis Res. Ther. 2010, 12, R19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabeya, Y.; Mizushima, N.; Ueno, T.; Yamamoto, A.; Kirisako, T.; Noda, T.; Kominami, E.; Ohsumi, Y.; Yoshimori, T. LC3, a mammalian homologue of yeast Apg8p, is localized in autophagosome membranes after processing. EMBO J. 2000, 19, 5720–5728. [Google Scholar] [CrossRef] [PubMed]
- Yeo, J.; Lee, Y.M.; Lee, J.; Park, D.; Kim, K.; Kim, J.; Park, J.; Kim, W.J. Nitric oxide-scavenging nanogel for treating rheumatoid arthritis. Nano Lett. 2019, 19, 6716–6724. [Google Scholar] [CrossRef] [PubMed]
- Paul-Clark, M.J.; Gilroy, D.W.; Willis, D.; Willoughby, D.A.; Tomlinson, A. Nitric oxide synthase inhibitors have opposite effects on acute inflammation depending on their route of administration. J. Immunol. 2001, 166, 1169–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chenevier-Gobeaux, C.; Morin-Robinet, S.; Lemarechal, H.; Poiraudeau, S.; Ekindjian, J.-C.O.; Borderie, D. Effects of pro-and anti-inflammatory cytokines and nitric oxide donors on hyaluronic acid synthesis by synovial cells from patients with rheumatoid arthritis. Clin. Sci. 2004, 107, 291–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomaa, A.A.; Elshenawy, M.M.; Afifi, N.A.; Mohammed, E.A.; Thabit, R.H. Dual effect of nitric oxide donor on adjuvant arthritis. Int. Immunopharmacol. 2009, 9, 439–447. [Google Scholar] [CrossRef]
- Kato, M.; Ospelt, C.; Gay, R.E.; Gay, S.; Klein, K. Dual role of autophagy in stress-induced cell death in rheumatoid arthritis synovial fibroblasts. Arthritis Rheumatol. 2014, 66, 40–48. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fermentation Period (Day) | 0 | 2 | 21 |
|---|---|---|---|
| Nitrite (μg/mL) | N.D * | 3.34 (3.28–3.41) ** | 1127.41 (1113.32–1141.50) ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.; Ryu, J.H.; Kim, B.-Y.; Chun, H.S.; Kim, M.S.; Shin, Y.-I. Fermented Lettuce Extract Containing Nitric Oxide Metabolites Attenuates Inflammatory Parameters in Model Mice and in Human Fibroblast-Like Synoviocytes. Nutrients 2023, 15, 1106. https://doi.org/10.3390/nu15051106
Park J, Ryu JH, Kim B-Y, Chun HS, Kim MS, Shin Y-I. Fermented Lettuce Extract Containing Nitric Oxide Metabolites Attenuates Inflammatory Parameters in Model Mice and in Human Fibroblast-Like Synoviocytes. Nutrients. 2023; 15(5):1106. https://doi.org/10.3390/nu15051106
Chicago/Turabian StylePark, Jisu, Ji Hyeon Ryu, Bo-Young Kim, Hyun Soo Chun, Min Sun Kim, and Yong-Il Shin. 2023. "Fermented Lettuce Extract Containing Nitric Oxide Metabolites Attenuates Inflammatory Parameters in Model Mice and in Human Fibroblast-Like Synoviocytes" Nutrients 15, no. 5: 1106. https://doi.org/10.3390/nu15051106
APA StylePark, J., Ryu, J. H., Kim, B. -Y., Chun, H. S., Kim, M. S., & Shin, Y. -I. (2023). Fermented Lettuce Extract Containing Nitric Oxide Metabolites Attenuates Inflammatory Parameters in Model Mice and in Human Fibroblast-Like Synoviocytes. Nutrients, 15(5), 1106. https://doi.org/10.3390/nu15051106