
Sex-Specific Response of the Brain Free Oxylipin Profile to Soluble Epoxide Hydrolase Inhibition

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
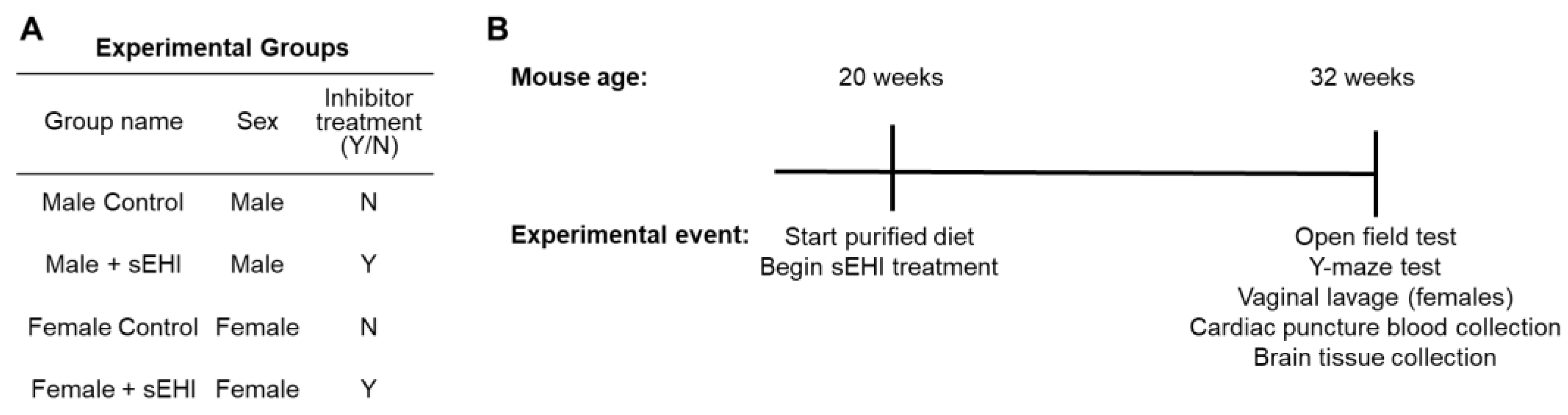
2.1. Animals
2.2. Soluble Epoxide Hydrolase Inhibitor (sEHI)
2.3. Assessment of Estrus Cycle Phase
2.4. Tissue Collection
2.5. Serum Analyses
2.6. Mouse Behavioral and Cognition Testing
2.6.1. Open Field Test
2.6.2. Y-Maze
2.7. Analysis of the Brain Free Oxylipin Profile
2.7.1. Free Oxylipin Extraction from Brain
2.7.2. UHPLC-MS/MS Analysis of Free Oxylipins
2.7.3. Data Analysis
2.8. Statistical Analysis
3. Results
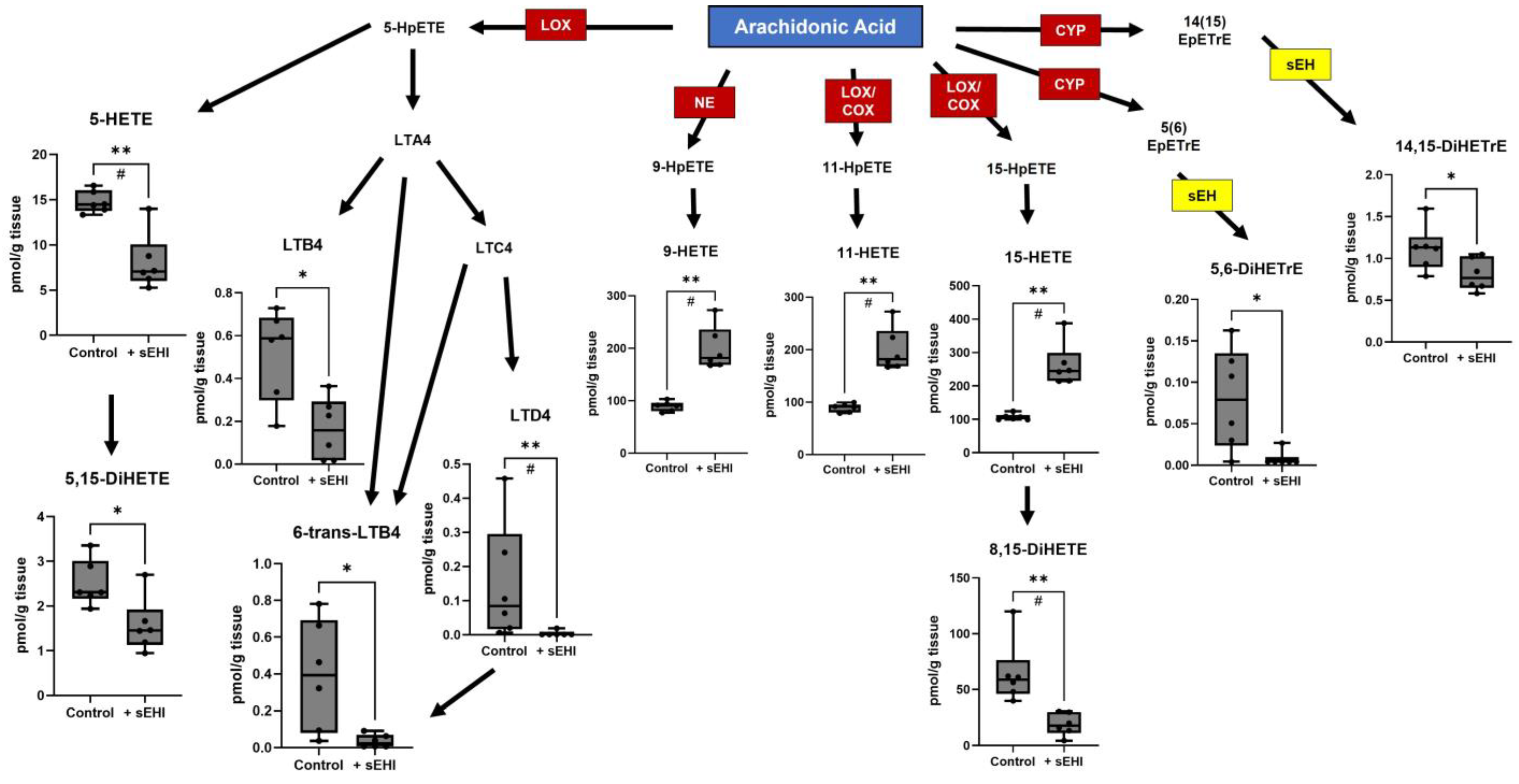
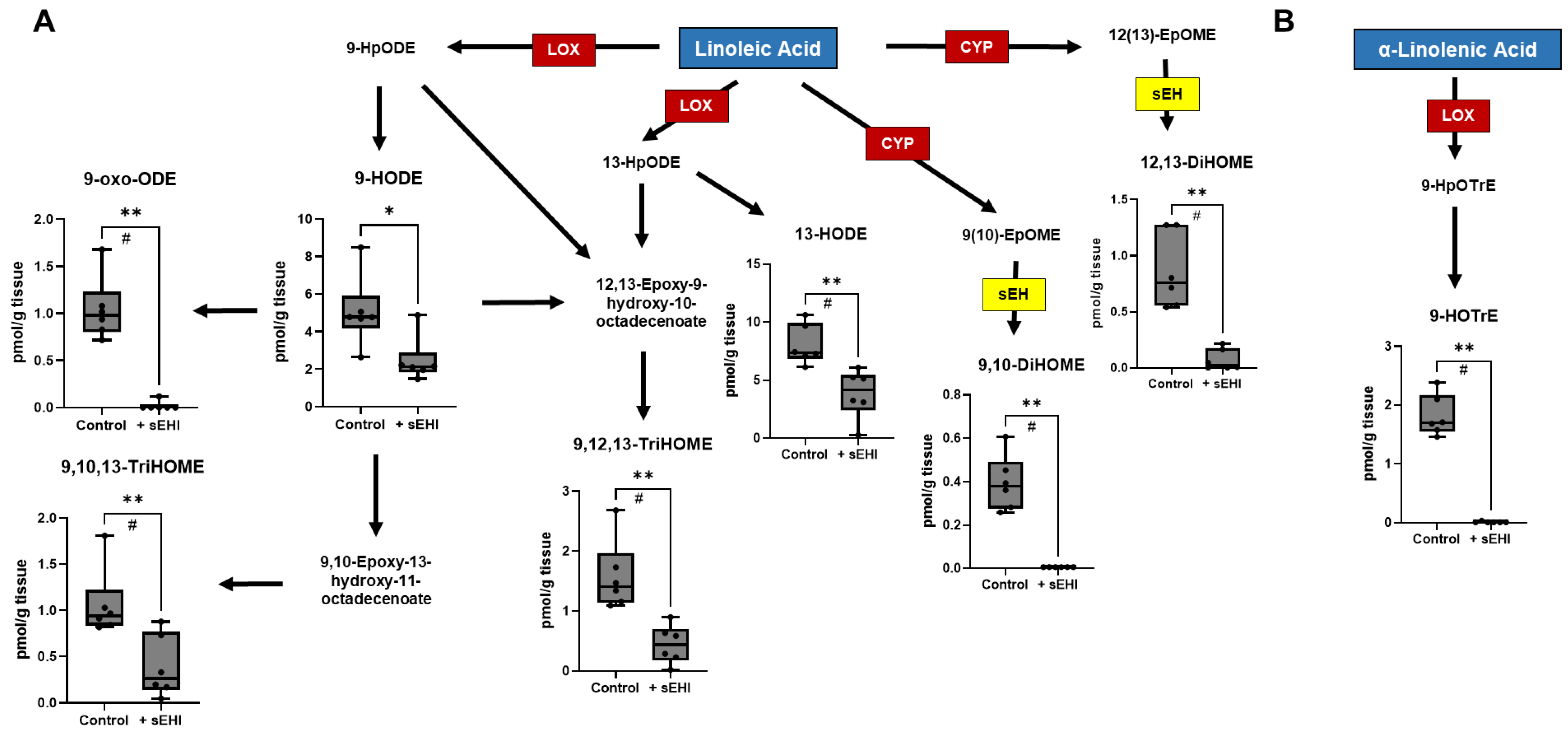
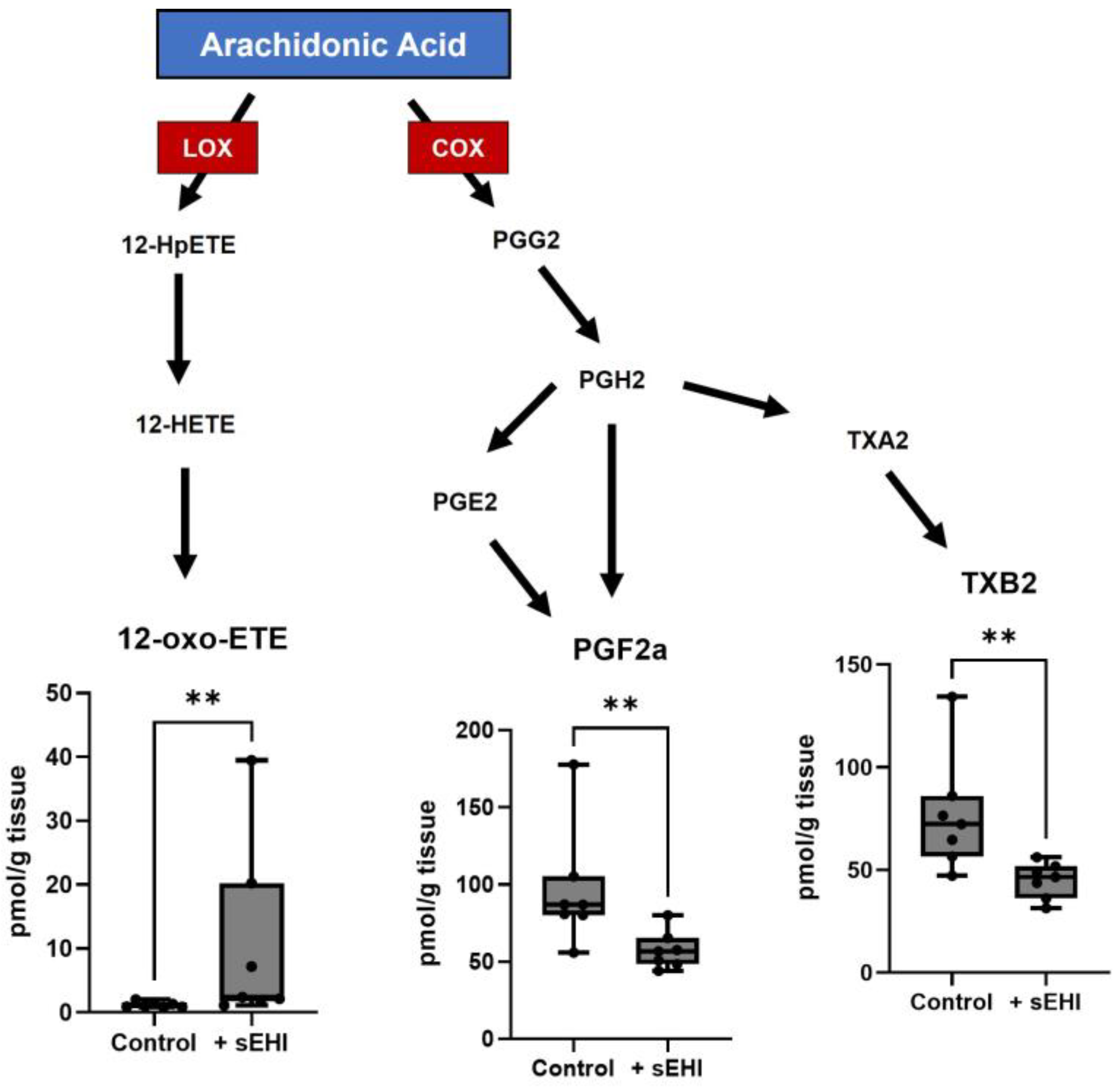
3.1. Effects of sEHI on the Brain Oxylipin Content in Males
3.2. Effects of sEHI on the Brain Oxylipin Content in Females
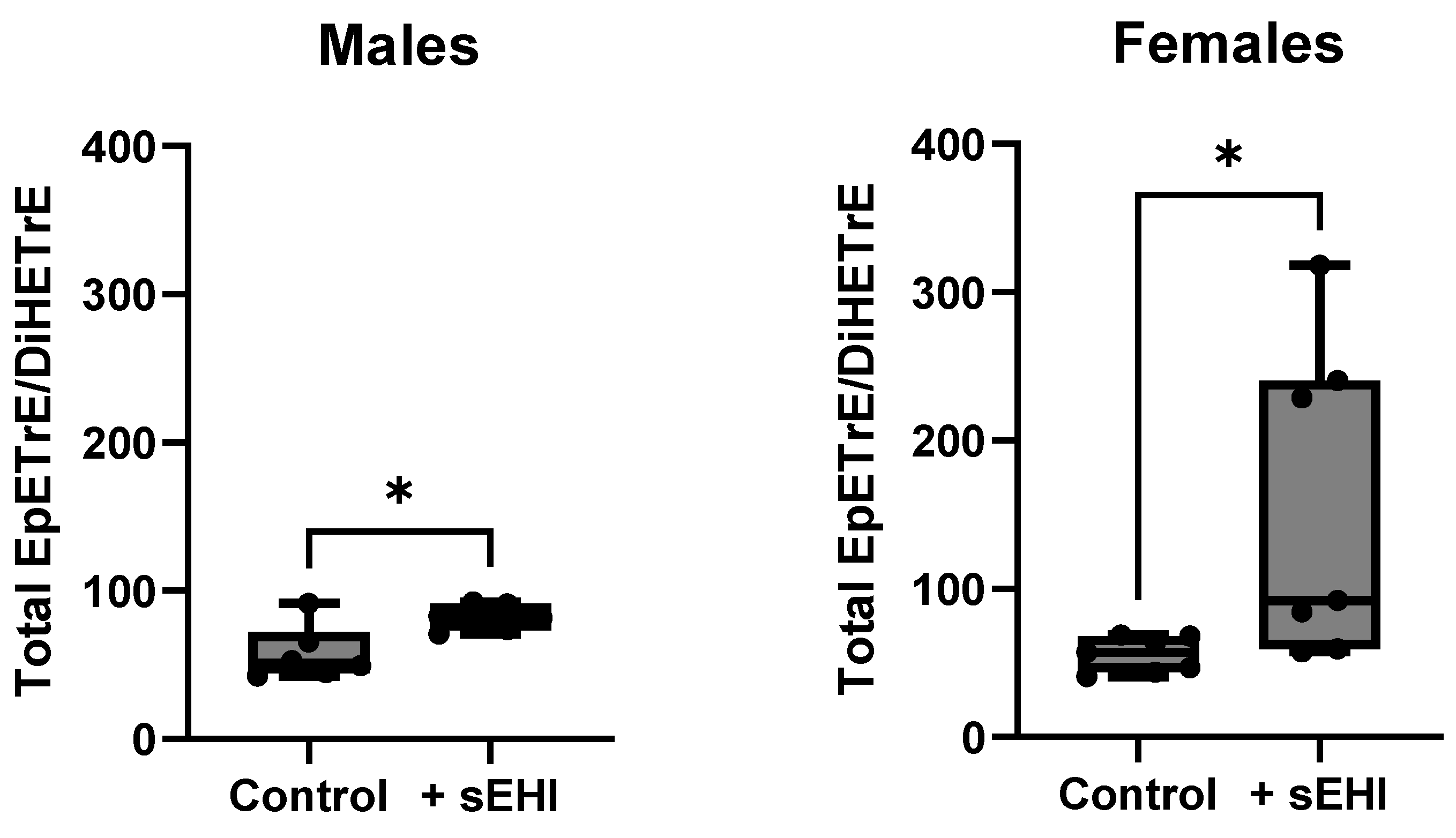
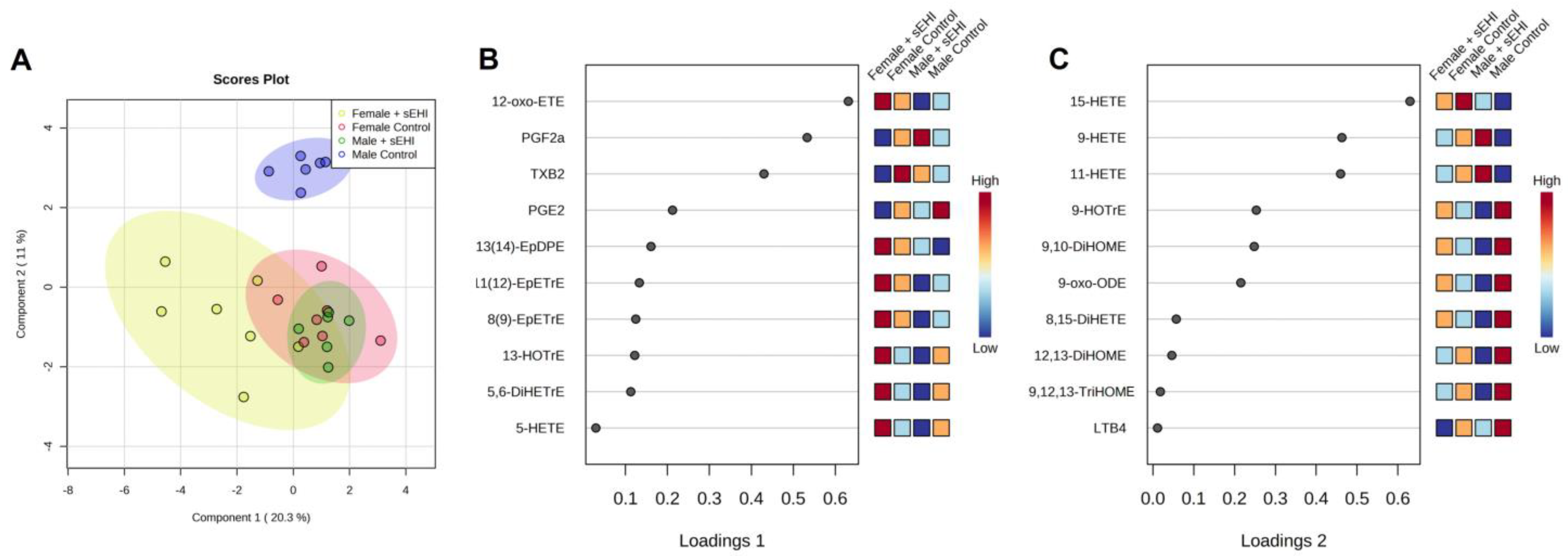
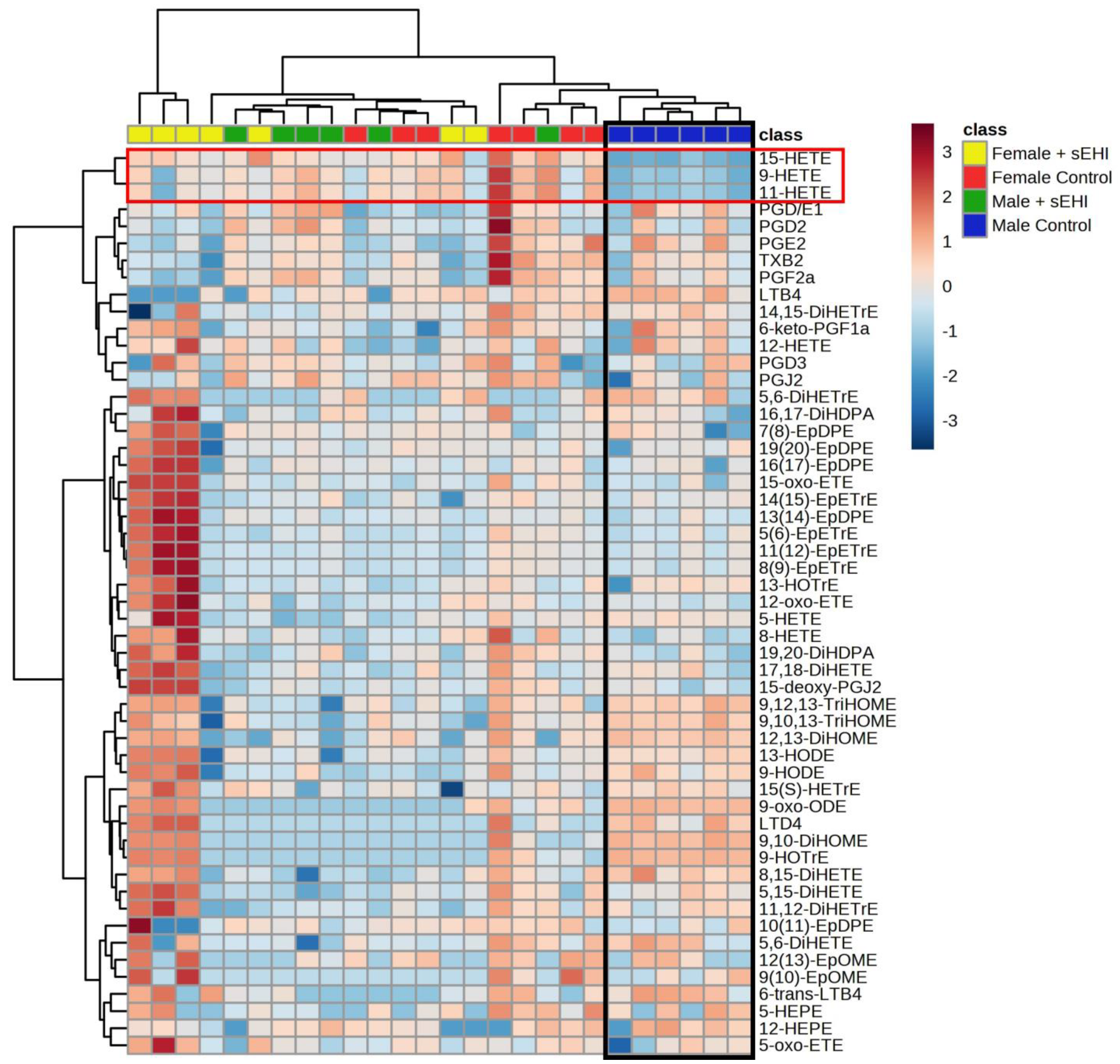
3.3. Comparison of the Effect of sEHI on the Brain Oxylipin Profile in Males and Females
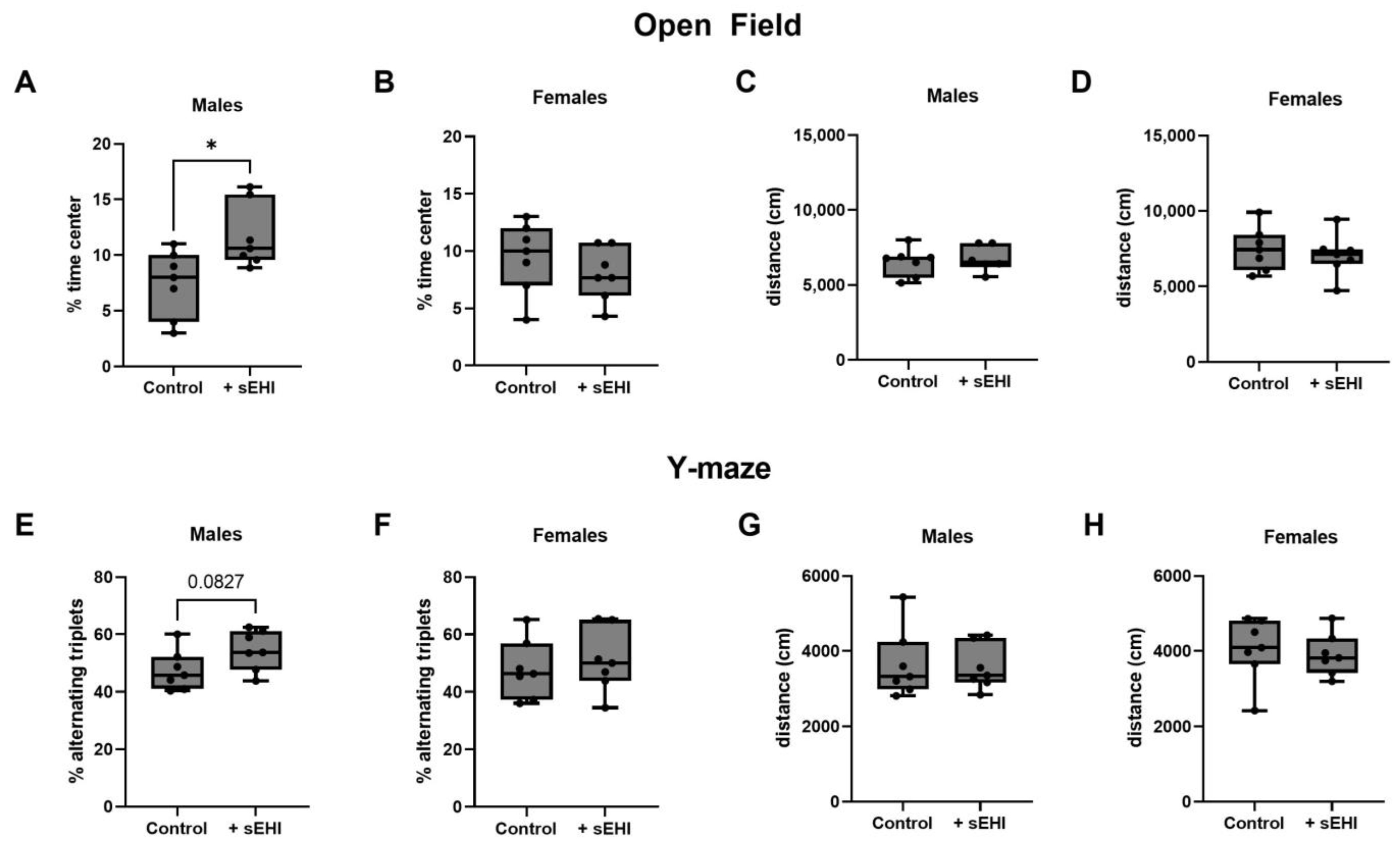
3.4. sEHI Effects on Behavior and Cognition in Male and Female Mice
4. Discussion
4.1. Implications of Oxylipins Altered by sEHI in Males
4.2. Implications of Oxylipins Altered by sEHI in Females
4.3. Sex Differences in the Response of the Brain Oxylipin Profile to sEHI
4.4. Sex Differences in the Cognitive and Behavioral Outcomes with sEHI
4.5. Potential Mechanisms of sEHI Effects on Brain Oxylipins
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gabbs, M.; Leng, S.; Devassy, J.G.; Monirujjaman, M.; Aukema, H.M. Advances in Our Understanding of Oxylipins Derived from Dietary PUFAs. Adv. Nutr. 2015, 6, 513–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauls, S.D.; Du, Y.; Clair, L.; Winter, T.; Aukema, H.M.; Taylor, C.G.; Zahradka, P. Impact of Age, Menopause, and Obesity on Oxylipins Linked to Vascular Health. Arterioscler. Thromb. Vasc. Biol. 2021, 41, 883–897. [Google Scholar] [CrossRef] [PubMed]
- Nayeem, M.A. Role of Oxylipins in Cardiovascular Diseases. Acta Pharmacol. Sin. 2018, 39, 1142–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caligiuri, S.P.B.; Aukema, H.M.; Ravandi, A.; Lavallée, R.; Guzman, R.; Pierce, G.N. Specific Plasma Oxylipins Increase the Odds of Cardiovascular and Cerebrovascular Events in Patients with Peripheral Artery Disease. Can. J. Physiol. Pharmacol. 2017, 95, 961–968. [Google Scholar] [CrossRef] [Green Version]
- Caligiuri, S.P.B.; Parikh, M.; Stamenkovic, A.; Pierce, G.N.; Aukema, H.M. Dietary Modulation of Oxylipins in Cardiovascular Disease and Aging. Am. J. Physiol. Heart Circ. Physiol. 2017, 313, H903–H918. [Google Scholar] [CrossRef] [Green Version]
- Tourdot, B.; Ahmed, I.; Holinstat, M. The Emerging Role of Oxylipins in Thrombosis and Diabetes. Front. Pharmacol. 2014, 4, 176. [Google Scholar] [CrossRef]
- Gleim, S.; Stitham, J.; Tang, W.H.; Martin, K.A.; Hwa, J. An Eicosanoid-Centric View of Atherothrombotic Risk Factors. Cell. Mol. Life Sci. 2012, 69, 3361–3380. [Google Scholar] [CrossRef] [Green Version]
- Shinto, L.H.; Raber, J.; Mishra, A.; Roese, N.; Silbert, L.C. A Review of Oxylipins in Alzheimer’s Disease and Related Dementias (ADRD): Potential Therapeutic Targets for the Modulation of Vascular Tone and Inflammation. Metabolites 2022, 12, 826. [Google Scholar] [CrossRef]
- Borkowski, K.; Pedersen, T.L.; Seyfried, N.T.; Lah, J.J.; Levey, A.I.; Hales, C.M.; Dammer, E.B.; Blach, C.; Louie, G.; Kaddurah-Daouk, R.; et al. Association of Plasma and CSF Cytochrome P450, Soluble Epoxide Hydrolase, and Ethanolamide Metabolism with Alzheimer’s Disease. Alzheimers Res. Ther. 2021, 13, 149. [Google Scholar] [CrossRef]
- Shinto, L.; Lahna, D.; Murchison, C.F.; Dodge, H.; Hagen, K.; David, J.; Kaye, J.; Quinn, J.F.; Wall, R.; Silbert, L.C. Oxidized Products of Omega-6 and Omega-3 Long Chain Fatty Acids Are Associated with Increased White Matter Hyperintensity and Poorer Executive Function Performance in a Cohort of Cognitively Normal Hypertensive Older Adults. J. Alzheimers Dis. 2020, 74, 65–77. [Google Scholar] [CrossRef]
- Devassy, J.G.; Leng, S.; Gabbs, M.; Monirujjaman, M.; Aukema, H.M. Omega-3 Polyunsaturated Fatty Acids and Oxylipins in Neuroinflammation and Management of Alzheimer Disease. Adv. Nutr. 2016, 7, 905–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tassoni, D.; Kaur, G.; Weisinger, R.S.; Sinclair, A.J. The Role of Eicosanoids in the Brain. Asia Pac. J. Clin. Nutr. 2008, 17 (Suppl. 1), 220–228. [Google Scholar]
- Wagner, K.M.; McReynolds, C.B.; Schmidt, W.K.; Hammock, B.D. Soluble Epoxide Hydrolase as a Therapeutic Target for Pain, Inflammatory and Neurodegenerative Diseases. Pharmacol. Ther. 2017, 180, 62–76. [Google Scholar] [CrossRef] [PubMed]
- Enayetallah, A.E.; French, R.A.; Thibodeau, M.S.; Grant, D.F. Distribution of Soluble Epoxide Hydrolase and of Cytochrome P450 2C8, 2C9, and 2J2 in Human Tissues. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2004, 52, 447–454. [Google Scholar] [CrossRef] [Green Version]
- Sura, P.; Sura, R.; Enayetallah, A.E.; Grant, D.F. Distribution and Expression of Soluble Epoxide Hydrolase in Human Brain. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2008, 56, 551–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, K.; Jain, S.; Paliwal, S.; Paliwal, S.; Sharma, S. A Clinical Perspective of Soluble Epoxide Hydrolase Inhibitors in Metabolic and Related Cardiovascular Diseases. Curr. Mol. Pharmacol. 2022, 15, 763–778. [Google Scholar] [CrossRef]
- Imig, J.D.; Cervenka, L.; Neckar, J. Epoxylipids and Soluble Epoxide Hydrolase in Heart Diseases. Biochem. Pharmacol. 2022, 195, 114866. [Google Scholar] [CrossRef] [PubMed]
- Wagner, K.M.; Yang, J.; Morisseau, C.; Hammock, B.D. Soluble Epoxide Hydrolase Deletion Limits High-Fat Diet-Induced Inflammation. Front. Pharmacol. 2021, 12, 778470. [Google Scholar] [CrossRef]
- Imig, J.D. Epigenetic Soluble Epoxide Hydrolase Regulation Causes Endothelial Dysfunction. Acta Physiol. Oxf. Engl. 2019, 225, e13203. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Wang, C.; Zhu, Y.; Ai, D. Soluble Epoxide Hydrolase: A Potential Target for Metabolic Diseases. J. Diabetes 2016, 8, 305–313. [Google Scholar] [CrossRef] [Green Version]
- Shan, J.; Hashimoto, K. Soluble Epoxide Hydrolase as a Therapeutic Target for Neuropsychiatric Disorders. Int. J. Mol. Sci. 2022, 23, 4951. [Google Scholar] [CrossRef]
- Ren, Q. Soluble Epoxide Hydrolase Inhibitor: A Novel Potential Therapeutic or Prophylactic Drug for Psychiatric Disorders. Front. Pharmacol. 2019, 10, 420. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K. Role of Soluble Epoxide Hydrolase in Metabolism of PUFAs in Psychiatric and Neurological Disorders. Front. Pharmacol. 2019, 10, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borsini, A. The Role of Soluble Epoxide Hydrolase and Its Inhibitors in Depression. Brain Behav. Immun.—Health 2021, 16, 100325. [Google Scholar] [CrossRef] [PubMed]
- Pu, Y.; Yang, J.; Chang, L.; Qu, Y.; Wang, S.; Zhang, K.; Xiong, Z.; Zhang, J.; Tan, Y.; Wang, X.; et al. Maternal Glyphosate Exposure Causes Autism-like Behaviors in Offspring through Increased Expression of Soluble Epoxide Hydrolase. Proc. Natl. Acad. Sci. USA 2020, 117, 11753–11759. [Google Scholar] [CrossRef]
- Shen, Y.; Peng, W.; Chen, Q.; Hammock, B.D.; Liu, J.; Li, D.; Yang, J.; Ding, J.; Wang, X. Anti-Inflammatory Treatment with a Soluble Epoxide Hydrolase Inhibitor Attenuates Seizures and Epilepsy-Associated Depression in the LiCl-Pilocarpine Post-Status Epilepticus Rat Model. Brain. Behav. Immun. 2019, 81, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Swardfager, W.; Hennebelle, M.; Yu, D.; Hammock, B.D.; Levitt, A.J.; Hashimoto, K.; Taha, A.Y. Metabolic/Inflammatory/Vascular Comorbidity in Psychiatric Disorders; Soluble Epoxide Hydrolase (SEH) as a Possible New Target. Neurosci. Biobehav. Rev. 2018, 87, 56–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, D.; Liang, N.; Zebarth, J.; Shen, Q.; Ozzoude, M.; Goubran, M.; Rabin, J.S.; Ramirez, J.; Scott, C.J.M.; Gao, F.; et al. Soluble Epoxide Hydrolase Derived Linoleic Acid Oxylipins, Small Vessel Disease Markers, and Neurodegeneration in Stroke. J. Am. Heart Assoc. 2023, 12, e026901. [Google Scholar] [CrossRef]
- Hasumi, K.; Suzuki, E. Impact of SMTP Targeting Plasminogen and Soluble Epoxide Hydrolase on Thrombolysis, Inflammation, and Ischemic Stroke. Int. J. Mol. Sci. 2021, 22, 954. [Google Scholar] [CrossRef]
- Yeh, C.-F.; Chuang, T.-Y.; Hung, Y.-W.; Lan, M.-Y.; Tsai, C.-H.; Huang, H.-X.; Lin, Y.-Y. Soluble Epoxide Hydrolase Inhibition Enhances Anti-Inflammatory and Antioxidative Processes, Modulates Microglia Polarization, and Promotes Recovery after Ischemic Stroke. Neuropsychiatr. Dis. Treat. 2019, 15, 2927–2941. [Google Scholar] [CrossRef] [Green Version]
- Yeh, C.-F.; Chuang, T.-Y.; Hung, Y.-W.; Lan, M.-Y.; Tsai, C.-H.; Huang, H.-X.; Lin, Y.-Y. Inhibition of Soluble Epoxide Hydrolase Regulates Monocyte/Macrophage Polarization and Improves Neurological Outcome in a Rat Model of Ischemic Stroke. Neuroreport 2019, 30, 567–572. [Google Scholar] [CrossRef]
- Wu, C.-H.; Shyue, S.-K.; Hung, T.-H.; Wen, S.; Lin, C.-C.; Chang, C.-F.; Chen, S.-F. Genetic Deletion or Pharmacological Inhibition of Soluble Epoxide Hydrolase Reduces Brain Damage and Attenuates Neuroinflammation after Intracerebral Hemorrhage. J. NeuroInflamm. 2017, 14, 230. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Dahl, M.; Grande, P.; Tybjaerg-Hansen, A.; Nordestgaard, B.G. Genetically Reduced Soluble Epoxide Hydrolase Activity and Risk of Stroke and Other Cardiovascular Disease. Stroke 2010, 41, 27–33. [Google Scholar] [CrossRef]
- Pallàs, M.; Vázquez, S.; Sanfeliu, C.; Galdeano, C.; Griñán-Ferré, C. Soluble Epoxide Hydrolase Inhibition to Face Neuroinflammation in Parkinson’s Disease: A New Therapeutic Strategy. Biomolecules 2020, 10, 703. [Google Scholar] [CrossRef]
- Jarne-Ferrer, J.; Griñán-Ferré, C.; Bellver-Sanchis, A.; Vázquez, S.; Muñoz-Torrero, D.; Pallàs, M. A Combined Chronic Low-Dose Soluble Epoxide Hydrolase and Acetylcholinesterase Pharmacological Inhibition Promotes Memory Reinstatement in Alzheimer’s Disease Mice Models. Pharmaceuticals 2022, 15, 908. [Google Scholar] [CrossRef] [PubMed]
- Griñán-Ferré, C.; Codony, S.; Pujol, E.; Yang, J.; Leiva, R.; Escolano, C.; Puigoriol-Illamola, D.; Companys-Alemany, J.; Corpas, R.; Sanfeliu, C.; et al. Pharmacological Inhibition of Soluble Epoxide Hydrolase as a New Therapy for Alzheimer’s Disease. Neurother. J. Am. Soc. Exp. Neurother. 2020, 17, 1825–1835. [Google Scholar] [CrossRef]
- Ghosh, A.; Comerota, M.M.; Wan, D.; Chen, F.; Propson, N.E.; Hwang, S.H.; Hammock, B.D.; Zheng, H. An Epoxide Hydrolase Inhibitor Reduces Neuroinflammation in a Mouse Model of Alzheimer’s Disease. Sci. Transl. Med. 2020, 12, eabb1206. [Google Scholar] [CrossRef]
- Nelson, J.W.; Young, J.M.; Borkar, R.N.; Woltjer, R.L.; Quinn, J.F.; Silbert, L.C.; Grafe, M.R.; Alkayed, N.J. Role of Soluble Epoxide Hydrolase in Age-Related Vascular Cognitive Decline. Prostaglandins Other Lipid Mediat. 2014, 113–115, 30–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.-T.; Lee, K.-I.; Chen, C.-H.; Lee, T.-S. Genetic Deletion of Soluble Epoxide Hydrolase Delays the Progression of Alzheimer’s Disease. J. Neuroinflamm. 2019, 16, 267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, D.; Hennebelle, M.; Sahlas, D.J.; Ramirez, J.; Gao, F.; Masellis, M.; Cogo-Moreira, H.; Swartz, R.H.; Herrmann, N.; Chan, P.C.; et al. Soluble Epoxide Hydrolase-Derived Linoleic Acid Oxylipins in Serum Are Associated with Periventricular White Matter Hyperintensities and Vascular Cognitive Impairment. Transl. Stroke Res. 2019, 10, 522–533. [Google Scholar] [CrossRef] [Green Version]
- Kuo, Y.-M.; Lee, Y.-H. Epoxyeicosatrienoic Acids and Soluble Epoxide Hydrolase in Physiology and Diseases of the Central Nervous System. Chin. J. Physiol. 2022, 65, 1–11. [Google Scholar] [CrossRef]
- Tian, Y.; Yuan, X.; Wang, Y.; Wu, Q.; Fang, Y.; Zhu, Z.; Song, G.; Xu, L.; Wang, W.; Xie, M. Soluble Epoxide Hydrolase Inhibitor Attenuates BBB Disruption and Neuroinflammation after Intracerebral Hemorrhage in Mice. Neurochem. Int. 2021, 150, 105197. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhao, Y.; Fan, Z.; Chen, Q.; Chen, J.; Sun, Y.; Jiang, X.; Xiao, Q. Soluble Epoxide Hydrolase Inhibitor Protects against Blood-Brain Barrier Dysfunction in a Mouse Model of Type 2 Diabetes via the AMPK/HO-1 Pathway. Biochem. Biophys. Res. Commun. 2020, 524, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.-M.; Hsu, P.-C.; Hung, C.-C.; Hu, Y.-Y.; Huang, Y.-J.; Gan, Y.-L.; Lin, C.-H.; Shie, F.-S.; Chang, W.-K.; Kao, L.-S.; et al. Soluble Epoxide Hydrolase Inhibition Attenuates Excitotoxicity Involving 14,15-Epoxyeicosatrienoic Acid-Mediated Astrocytic Survival and Plasticity to Preserve Glutamate Homeostasis. Mol. Neurobiol. 2019, 56, 8451–8474. [Google Scholar] [CrossRef] [PubMed]
- Hung, T.-H.; Shyue, S.-K.; Wu, C.-H.; Chen, C.-C.; Lin, C.-C.; Chang, C.-F.; Chen, S.-F. Deletion or Inhibition of Soluble Epoxide Hydrolase Protects against Brain Damage and Reduces Microglia-Mediated Neuroinflammation in Traumatic Brain Injury. Oncotarget 2017, 8, 103236–103260. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.-F.; Yen, H.-J.; Huang, C.-C.; Lee, Y.-C.; Wu, S.-Z.; Lee, T.-S.; Lin, H.-C. Soluble Epoxide Hydrolase Inhibitor Enhances Synaptic Neurotransmission and Plasticity in Mouse Prefrontal Cortex. J. Biomed. Sci. 2015, 22, 94. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Davis, C.M.; Edin, M.L.; Lee, C.R.; Zeldin, D.C.; Alkayed, N.J. Role of Endothelial Soluble Epoxide Hydrolase in Cerebrovascular Function and Ischemic Injury. PloS ONE 2013, 8, e61244. [Google Scholar] [CrossRef]
- Pinot, F.; Grant, D.F.; Spearow, J.L.; Parker, A.G.; Hammock, B.D. Differential Regulation of Soluble Epoxide Hydrolase by Clofibrate and Sexual Hormones in the Liver and Kidneys of Mice. Biochem. Pharmacol. 1995, 50, 501–508. [Google Scholar] [CrossRef]
- Davis, C.M.; Fairbanks, S.L.; Alkayed, N.J. Mechanism of the Sex Difference in Endothelial Dysfunction after Stroke. Transl. Stroke Res. 2013, 4, 381–389. [Google Scholar] [CrossRef]
- Zhang, W.; Iliff, J.J.; Campbell, C.J.; Wang, R.K.; Hurn, P.D.; Alkayed, N.J. Role of Soluble Epoxide Hydrolase in the Sex-Specific Vascular Response to Cerebral Ischemia. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2009, 29, 1475–1481. [Google Scholar] [CrossRef] [Green Version]
- Gupta, N.C.; Davis, C.M.; Nelson, J.W.; Young, J.M.; Alkayed, N.J. Soluble Epoxide Hydrolase: Sex Differences and Role in Endothelial Cell Survival. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1936–1942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fairbanks, S.L.; Young, J.M.; Nelson, J.W.; Davis, C.M.; Koerner, I.P.; Alkayed, N.J. Mechanism of the Sex Difference in Neuronal Ischemic Cell Death. Neuroscience 2012, 219, 183–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koerner, I.P.; Zhang, W.; Cheng, J.; Parker, S.; Hurn, P.D.; Alkayed, N.J. Soluble Epoxide Hydrolase: Regulation by Estrogen and Role in the Inflammatory Response to Cerebral Ischemia. Front. Biosci. J. Virtual Libr. 2008, 13, 2833–2841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuthikattu, S.; Milenkovic, D.; Norman, J.E.; Rutledge, J.; Villablanca, A. The Brain’s Microvascular Response to High Glycemia and to the Inhibition of Soluble Epoxide Hydrolase Is Sexually Dimorphic. Nutrients 2022, 14, 3451. [Google Scholar] [CrossRef]
- The ARRIVE Guidelines 2.0. Available online: https://arriveguidelines.org/arrive-guidelines (accessed on 29 August 2022).
- Nuthikattu, S.; Milenkovic, D.; Norman, J.E.; Rutledge, J.; Villablanca, A. Inhibition of Soluble Epoxide Hydrolase Is Protective against the Multiomic Effects of a High Glycemic Diet on Brain Microvascular Inflammation and Cognitive Dysfunction. Nutrients 2021, 13, 3913. [Google Scholar] [CrossRef]
- Nuthikattu, S.; Milenkovic, D.; Norman, J.E.; Rutledge, J.; Villablanca, A. High Glycemia and Soluble Epoxide Hydrolase in Females: Differential Multiomics in Murine Brain Microvasculature. Int. J. Mol. Sci. 2022, 23, 13044. [Google Scholar] [CrossRef] [PubMed]
- Poli, G.; Corda, E.; Martino, P.A.; Dall’ara, P.; Bareggi, S.R.; Bondiolotti, G.; Iulini, B.; Mazza, M.; Casalone, C.; Hwang, S.H.; et al. Therapeutic Activity of Inhibition of the Soluble Epoxide Hydrolase in a Mouse Model of Scrapie. Life Sci. 2013, 92, 1145–1150. [Google Scholar] [CrossRef] [Green Version]
- Roche, C.; Besnier, M.; Cassel, R.; Harouki, N.; Coquerel, D.; Guerrot, D.; Nicol, L.; Loizon, E.; Remy-Jouet, I.; Morisseau, C.; et al. Soluble Epoxide Hydrolase Inhibition Improves Coronary Endothelial Function and Prevents the Development of Cardiac Alterations in Obese Insulin-Resistant Mice. Am. J. Physiol. Heart Circ. Physiol. 2015, 308, H1020–H1029. [Google Scholar] [CrossRef] [Green Version]
- Gui, Y.-J.; Yang, T.; Liu, Q.; Liao, C.-X.; Chen, J.-Y.; Wang, Y.-T.; Hu, J.-H.; Xu, D.-Y. Soluble Epoxide Hydrolase Inhibitors, t-AUCB, Regulated MicroRNA-1 and Its Target Genes in Myocardial Infarction Mice. Oncotarget 2017, 8, 94635–94649. [Google Scholar] [CrossRef] [Green Version]
- Hermansky, S.J.; Neptun, D.A.; Loughran, K.A.; Leung, H.W. Effects of Polyethylene Glycol 400 (PEG 400) Following 13 Weeks of Gavage Treatment in Fischer-344 Rats. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 1995, 33, 139–149. [Google Scholar] [CrossRef] [PubMed]
- McLean, A.C.; Valenzuela, N.; Fai, S.; Bennett, S.A.L. Performing Vaginal Lavage, Crystal Violet Staining, and Vaginal Cytological Evaluation for Mouse Estrous Cycle Staging Identification. J. Vis. Exp. JoVE 2012, 67, e4389. [Google Scholar] [CrossRef] [Green Version]
- Kraeuter, A.-K.; Guest, P.C.; Sarnyai, Z. The Open Field Test for Measuring Locomotor Activity and Anxiety-Like Behavior. Methods Mol. Biol. Clifton NJ 2019, 1916, 99–103. [Google Scholar] [CrossRef]
- Hölter, S.M.; Garrett, L.; Einicke, J.; Sperling, B.; Dirscherl, P.; Zimprich, A.; Fuchs, H.; Gailus-Durner, V.; Hrabě de Angelis, M.; Wurst, W. Assessing Cognition in Mice. Curr. Protoc. Mouse Biol. 2015, 5, 331–358. [Google Scholar] [CrossRef] [PubMed]
- Norman, J.E.; Nuthikattu, S.; Milenkovic, D.; Rutledge, J.C.; Villablanca, A.C. A High Sucrose Diet Modifies Brain Oxylipins in a Sex-Dependent Manner. Prostaglandins Leukot. Essent. Fatty Acids 2022, 186, 102506. [Google Scholar] [CrossRef] [PubMed]
- MetaboAnalyst. Available online: https://www.metaboanalyst.ca/MetaboAnalyst/ModuleView.xhtml (accessed on 25 August 2021).
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the Gap between Raw Spectra and Functional Insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef] [PubMed]
- Lê Cao, K.-A.; Boitard, S.; Besse, P. Sparse PLS Discriminant Analysis: Biologically Relevant Feature Selection and Graphical Displays for Multiclass Problems. BMC Bioinform. 2011, 12, 253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Zhu, Y.-M.; Li, Y.-N.; Li, P.-Y.; Wang, D.; Liu, Y.; Qu, Y.-Y.; Zhu, D.-L.; Zhu, Y.-L. The 15-LO-1/15-HETE System Promotes Angiogenesis by Upregulating VEGF in Ischemic Brains. Neurol. Res. 2017, 39, 795–802. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Liu, Y.; Chen, L.; Li, P.; Qu, Y.; Zhu, Y.; Zhu, Y. Key Role of 15-LO/15-HETE in Angiogenesis and Functional Recovery in Later Stages of Post-Stroke Mice. Sci. Rep. 2017, 7, 46698. [Google Scholar] [CrossRef] [Green Version]
- Tajima, Y.; Ishikawa, M.; Maekawa, K.; Murayama, M.; Senoo, Y.; Nishimaki-Mogami, T.; Nakanishi, H.; Ikeda, K.; Arita, M.; Taguchi, R.; et al. Lipidomic Analysis of Brain Tissues and Plasma in a Mouse Model Expressing Mutated Human Amyloid Precursor Protein/Tau for Alzheimer’s Disease. Lipids Health Dis. 2013, 12, 68. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; Wang, S.; Li, W.; Liu, Z.; Tang, J.; Tang, X. Activation of Peroxisome Proliferator-Activated Receptor-γ by a 12/15-Lipoxygenase Product of Arachidonic Acid: A Possible Neuroprotective Effect in the Brain after Experimental Intracerebral Hemorrhage. J. Neurosurg. 2017, 127, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Farias, S.E.; Heidenreich, K.A.; Wohlauer, M.V.; Murphy, R.C.; Moore, E.E. Lipid Mediators in Cerebral Spinal Fluid of Traumatic Brain Injured Patients. J. Trauma 2011, 71, 1211–1218. [Google Scholar] [CrossRef] [PubMed]
- Attaluri, S.; Upadhya, R.; Kodali, M.; Madhu, L.N.; Upadhya, D.; Shuai, B.; Shetty, A.K. Brain-Specific Increase in Leukotriene Signaling Accompanies Chronic Neuroinflammation and Cognitive Impairment in a Model of Gulf War Illness. Front. Immunol. 2022, 13, 853000. [Google Scholar] [CrossRef] [PubMed]
- Do, K.V.; Hjorth, E.; Wang, Y.; Jun, B.; Kautzmann, M.-A.I.; Ohshima, M.; Eriksdotter, M.; Schultzberg, M.; Bazan, N.G. Cerebrospinal Fluid Profile of Lipid Mediators in Alzheimer’s Disease. Cell. Mol. Neurobiol. 2023, 43, 797–811. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, Y.; Zhang, S.; Li, C.; Zhang, L. Modulation of Neuroinflammation by Cysteinyl Leukotriene 1 and 2 Receptors: Implications for Cerebral Ischemia and Neurodegenerative Diseases. Neurobiol. Aging 2020, 87, 1–10. [Google Scholar] [CrossRef]
- Wong, P.T.; McGeer, P.L.; McGeer, E.G. Decreased Prostaglandin Synthesis in Postmortem Cerebral Cortex from Patients with Alzheimer’s Disease. Neurochem. Int. 1992, 21, 197–202. [Google Scholar] [CrossRef]
- Iwamoto, N.; Kobayashi, K.; Kosaka, K. The Formation of Prostaglandins in the Postmortem Cerebral Cortex of Alzheimer-Type Dementia Patients. J. Neurol. 1989, 236, 80–84. [Google Scholar] [CrossRef]
- Liu, L.; Feng, A.; Du, C.; Qi, C. Relationship between the Changes in Thromboxane B2, 6-Keto-Prostaglandin Fla, and Blood Glucose Levels and Progressive Ischemic Stroke. Ann. Palliat. Med. 2021, 10, 5373–5379. [Google Scholar] [CrossRef]
- Wang, P.; Wang, W.; Hu, Y.; Li, Y. Prolonged Soluble Epoxide Hydrolase Reactivity in Brain Endothelial Cells Is Associated with Long Cognitive Deficits in Sepsis. Mol. Neurobiol. 2020, 57, 2846–2855. [Google Scholar] [CrossRef] [PubMed]
- Matin, N.; Fisher, C.; Lansdell, T.A.; Hammock, B.D.; Yang, J.; Jackson, W.F.; Dorrance, A.M. Soluble Epoxide Hydrolase Inhibition Improves Cognitive Function and Parenchymal Artery Dilation in a Hypertensive Model of Chronic Cerebral Hypoperfusion. Microcirculation 2021, 28, e12653. [Google Scholar] [CrossRef]
- Wu, J.; Fan, Z.; Zhao, Y.; Chen, Q.; Xiao, Q. Inhibition of Soluble Epoxide Hydrolase (SEH) Protects Hippocampal Neurons and Reduces Cognitive Decline in Type 2 Diabetic Mice. Eur. J. Neurosci. 2021, 53, 2532–2540. [Google Scholar] [CrossRef]
- Minaz, N.; Razdan, R.; Hammock, B.D.; Goswami, S.K. An Inhibitor of Soluble Epoxide Hydrolase Ameliorates Diabetes-Induced Learning and Memory Impairment in Rats. Prostaglandins Other Lipid Mediat. 2018, 136, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Chen, Y.; Yao, E.; Liu, X. Soluble Epoxide Hydrolase Inhibition Alleviated Cognitive Impairments via NRG1/ErbB4 Signaling after Chronic Cerebral Hypoperfusion Induced by Bilateral Carotid Artery Stenosis in Mice. Brain Res. 2018, 1699, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Pardeshi, R.; Bolshette, N.; Gadhave, K.; Arfeen, M.; Ahmed, S.; Jamwal, R.; Hammock, B.D.; Lahkar, M.; Goswami, S.K. Docosahexaenoic Acid Increases the Potency of Soluble Epoxide Hydrolase Inhibitor in Alleviating Streptozotocin-Induced Alzheimer’s Disease-Like Complications of Diabetes. Front. Pharmacol. 2019, 10, 288. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Li, N.; Pang, W.; Zhang, X.; Hammock, B.D.; Ai, D.; Zhu, Y. Opposite Effects of Gene Deficiency and Pharmacological Inhibition of Soluble Epoxide Hydrolase on Cardiac Fibrosis. PLoS ONE 2014, 9, e94092. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Males | Females | |||
|---|---|---|---|---|
| Control | + sEHI | Control | + sEHI | |
| Bodyweight (g) | 36.19 ± 6.183 | 36.46 ± 5.384 | 29.20 ± 4.940 | 27.75 ± 2.559 |
| Insulin (pg/mL) | 230.5 ± 173.6 | 285.4 ± 142.6 | 208.2 ± 55.18 | 332.4 ± 170.4 |
| Glucose (mg/dL) | 416.1 ± 108.6 | 432.7 ± 128.5 | 282.9 ± 77.08 | 319.4 ± 117.9 |
| Total Cholesterol (mg/dL) | 154.9 ± 33.75 | 139.4 ± 14.35 | 89.74 ± 17.29 | 98.81 ± 13.56 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Norman, J.E.; Nuthikattu, S.; Milenkovic, D.; Rutledge, J.C.; Villablanca, A.C. Sex-Specific Response of the Brain Free Oxylipin Profile to Soluble Epoxide Hydrolase Inhibition. Nutrients 2023, 15, 1214. https://doi.org/10.3390/nu15051214
Norman JE, Nuthikattu S, Milenkovic D, Rutledge JC, Villablanca AC. Sex-Specific Response of the Brain Free Oxylipin Profile to Soluble Epoxide Hydrolase Inhibition. Nutrients. 2023; 15(5):1214. https://doi.org/10.3390/nu15051214
Chicago/Turabian StyleNorman, Jennifer E., Saivageethi Nuthikattu, Dragan Milenkovic, John C. Rutledge, and Amparo C. Villablanca. 2023. "Sex-Specific Response of the Brain Free Oxylipin Profile to Soluble Epoxide Hydrolase Inhibition" Nutrients 15, no. 5: 1214. https://doi.org/10.3390/nu15051214
APA StyleNorman, J. E., Nuthikattu, S., Milenkovic, D., Rutledge, J. C., & Villablanca, A. C. (2023). Sex-Specific Response of the Brain Free Oxylipin Profile to Soluble Epoxide Hydrolase Inhibition. Nutrients, 15(5), 1214. https://doi.org/10.3390/nu15051214