
α-Mangostin Inhibits the Activation of Myofibroblasts via Downregulation of Linc-ROR-Mediated TGFB1/Smad Signaling

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Collection, Primary Culture and Reagents
2.2. Cell Proliferation and Survival Assay
2.3. Collagen Gel Contraction Assay
2.4. Transwell Migration Assays
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. RNA-Sequencing
2.7. Real-Time Quantitative Polymerase Chain Reaction (Qrt-PCR)
2.8. Western Blot Analysis
2.9. Lentiviral-Mediated Silencing and Overexpression of Lncrna-ROR
2.10. Statistical Analysis
3. Results
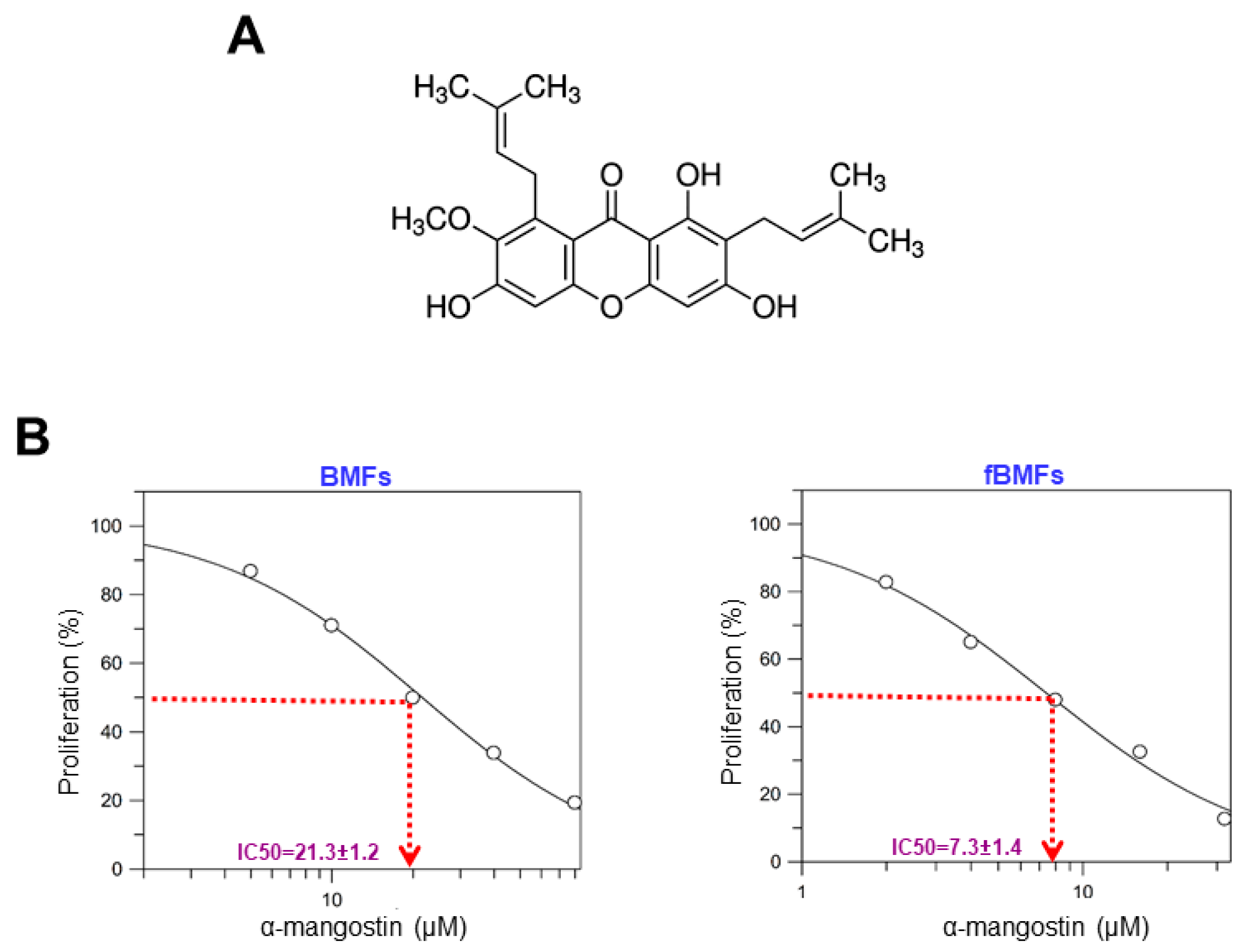
3.1. α-Mangostin Reduces the Cell Viability of Fbmfs and Has Minimal Effect on Normal Oral Cells
3.2. α-Mangostin Suppresses the Myofibroblast Activation of fBMFs
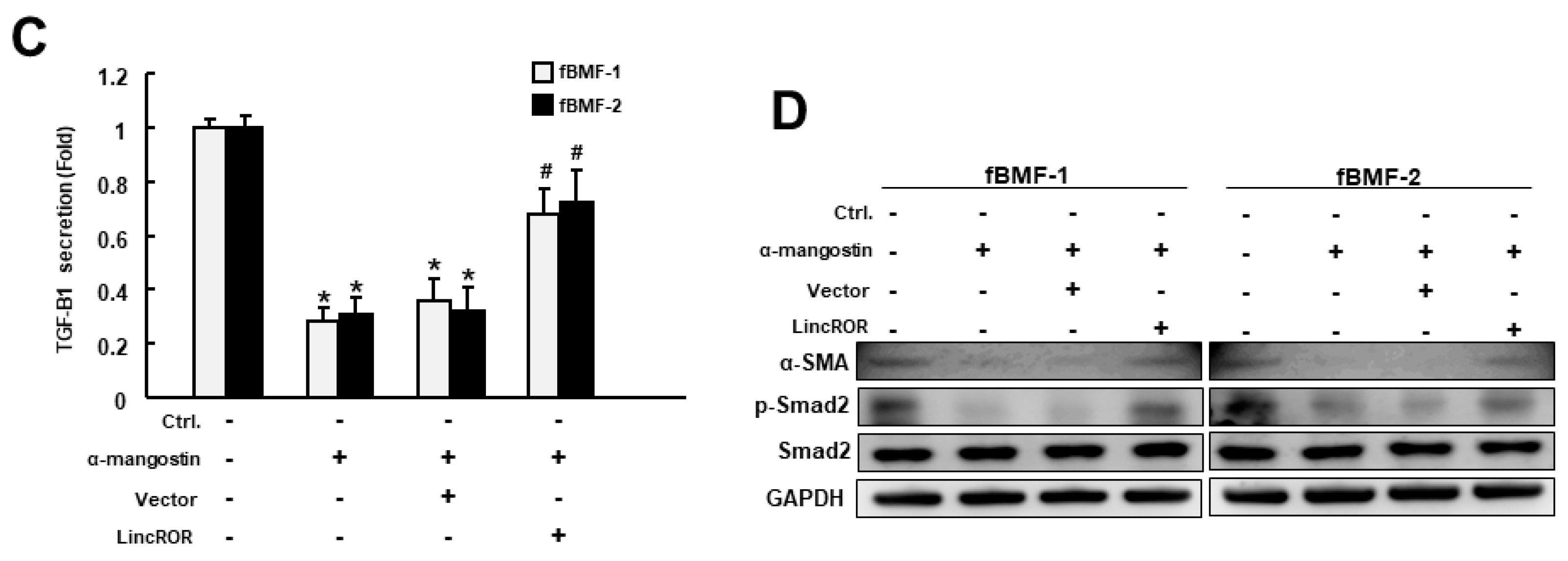
3.3. Incubation of α-Mangostin Downregulates the Expression of TGF-β1 Signaling, Myofibroblast Marker, and LincROR
3.4. The Inhibitory Property of α-Mangostin on Myofibroblast Activities and TGF-β Signaling Is Mediated by LincROR
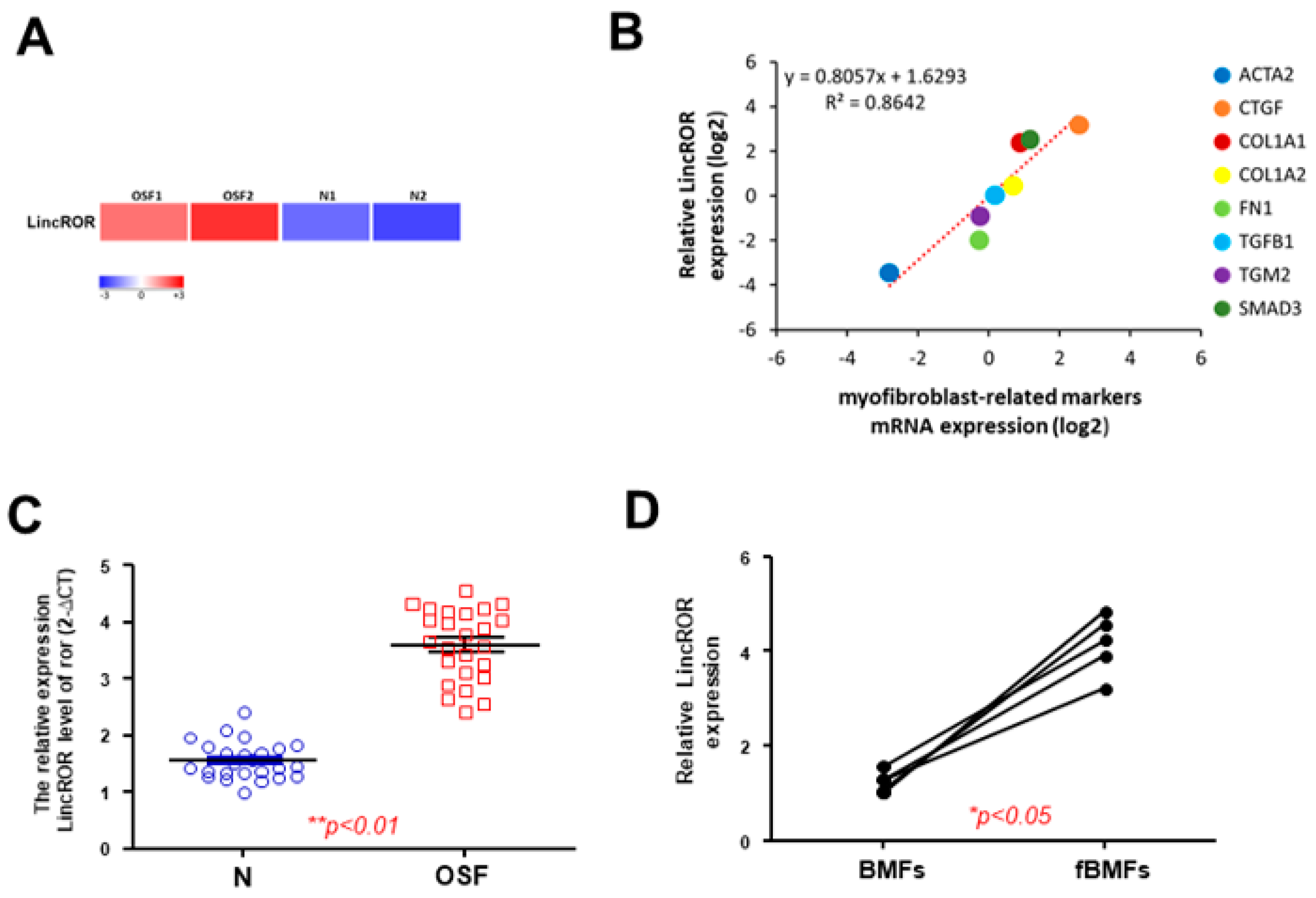
3.5. LincROR Is Aberrantly Overexpressed in OSF Specimens
3.6. Silencign of LincROR Inhibits Myofibroblast Activation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Iocca, O.; Sollecito, T.P.; Alawi, F.; Weinstein, G.S.; Newman, J.G.; De Virgilio, A.; Di Maio, P.; Spriano, G.; Pardiñas López, S.; Shanti, R.M. Potentially malignant disorders of the oral cavity and oral dysplasia: A systematic review and meta-analysis of malignant transformation rate by subtype. Head Neck 2020, 42, 539–555. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-Y.; Tail, Y.-H.; Wang, W.-C.; Chen, C.-Y.; Kao, Y.-H.; Chen, Y.-K.; Chen, C.-H. Malignant transformation in 5071 southern Taiwanese patients with potentially malignant oral mucosal disorders. BMC Oral Health 2014, 14, 99. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.-J.; Chang, M.-L.; Chiang, C.-P.; Hahn, L.-J.; Hsieh, L.-L.; Chen, C.-J. Interaction of collagen-related genes and susceptibility to betel quid-induced oral submucous fibrosis. Cancer Epidemiol. Biomark. Prev. 2002, 11, 646–653. [Google Scholar]
- Elamin, F.; Steingrimsdottir, H.; Wanakulasuriya, S.; Johnson, N.; Tavassoli, M. Prevalence of human papillomavirus infection in premalignant and malignant lesions of the oral cavity in U.K. subjects: A novel method of detection. Oral Oncol. 1998, 34, 191–197. [Google Scholar] [CrossRef]
- Lee, C.-H.; Ko, Y.-C.; Huang, H.-L.; Chao, Y.-Y.; Tsai, C.-C.; Shieh, T.-Y.; Lin, L.-M. The precancer risk of betel quid chewing, tobacco use and alcohol consumption in oral leukoplakia and oral submucous fibrosis in southern Taiwan. Br. J. Cancer 2003, 88, 366–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.C.; Ma, R.H.; Shieh, T.Y. Deficiency in collagen and fibronectin phagocytosis by human buccal mucosa fibroblasts in vitro as a possible mechanism for oral submucous fibrosis. J. Oral Pathol. Med. 1999, 28, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Shieh, D.-H.; Chiang, L.-C.; Shieh, T.-Y. Augmented mRNA expression of tissue inhibitor of metalloproteinase-1 in buccal mucosal fibroblasts by arecoline and safrole as a possible pathogenesis for oral submucous fibrosis. Oral Oncol. 2003, 39, 728–735. [Google Scholar] [CrossRef]
- Chang, Y.-C.; Yang, S.-F.; Tai, K.-W.; Chou, M.-Y.; Hsieh, Y.-S. Increased tissue inhibitor of metalloproteinase-1 expression and inhibition of gelatinase A activity in buccal mucosal fibroblasts by arecoline as possible mechanisms for oral submucous fibrosis. Oral Oncol. 2002, 38, 195–200. [Google Scholar] [CrossRef]
- Khan, I.; Kumar, N.; Pant, I.; Narra, S.; Kondaiah, P. Activation of TGF-β Pathway by Areca Nut Constituents: A Possible Cause of Oral Submucous Fibrosis. PLoS ONE 2012, 7, e51806. [Google Scholar] [CrossRef] [Green Version]
- Pant, I.; Kumar, N.; Khan, I.; Rao, S.G.; Kondaiah, P. Role of Areca Nut Induced TGF-β and Epithelial-Mesenchymal Interaction in the Pathogenesis of Oral Submucous Fibrosis. PLoS ONE 2015, 10, e0129252. [Google Scholar] [CrossRef]
- Hinz, B.; Phan, S.H.; Thannickal, V.J.; Galli, A.; Bochaton-Piallat, M.-L.; Gabbiani, G. The Myofibroblast: One Function, Multiple Origins. Am. J. Pathol. 2007, 170, 1807–1816. [Google Scholar] [CrossRef] [PubMed]
- Iwaisako, K.; Jiang, C.; Zhang, M.; Cong, M.; Moore-Morris, T.J.; Park, T.J.; Liu, X.; Xu, J.; Wang, P.; Paik, Y.-H.; et al. Origin of myofibroblasts in the fibrotic liver in mice. Proc. Natl. Acad. Sci. USA 2014, 111, E3297–E3305. [Google Scholar] [CrossRef] [Green Version]
- LeBleu, V.S.; Taduri, G.; O’Connell, J.; Teng, Y.; Cooke, V.G.; Woda, C.; Sugimoto, H.; Kalluri, R. Origin and function of myofibroblasts in kidney fibrosis. Nat. Med. 2013, 19, 1047–1053. [Google Scholar] [CrossRef] [PubMed]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.; Nieto, M.A. Epithelial-Mesenchymal Transitions in Development and Disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.-W.; Lu, M.-Y.; Chiu, Y.-W.; Liao, Y.-W.; Huang, Y.-F.; Ju Chueh, P.; Hsieh, P.-L.; Yu, C.-C. Hinokitiol ablates myofibroblast activation in precancerous oral submucous fibrosis by targeting Snail. Environ. Toxicol. 2018, 33, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.Y.; Hsia, S.M.; Hsieh, P.L.; Liao, Y.W.; Peng, C.Y.; Wu, C.Z.; Lin, K.C.; Tsai, L.L.; Yu, C. Slug mediates myofibroblastic differentiation to promote fibrogenesis in buccal mucosa. J. Cell. Physiol. 2019, 234, 6721–6730. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.-Y.; Liao, Y.-W.; Lu, M.-Y.; Yang, C.-M.; Hsieh, P.-L.; Yu, C.-C. Positive Feedback Loop of SNAIL-IL-6 Mediates Myofibroblastic Differentiation Activity in Precancerous Oral Submucous Fibrosis. Cancers 2020, 12, 1611. [Google Scholar] [CrossRef]
- Lee, P.-H.; Chu, P.-M.; Hsieh, P.-L.; Yang, H.-W.; Chueh, P.J.; Huang, Y.-F.; Liao, Y.-W.; Yu, C.-C. Glabridin inhibits the activation of myofibroblasts in human fibrotic buccal mucosal fibroblasts through TGF-β/smad signaling. Environ. Toxicol. 2018, 33, 248–255. [Google Scholar] [CrossRef]
- Chen, P.-Y.; Ho, D.C.; Liao, Y.-W.; Hsieh, P.-L.; Lu, K.-H.; Tsai, L.-L.; Su, S.-H.; Yu, C.-C. Honokiol inhibits arecoline-induced oral fibrogenesis through transforming growth factor-β/Smad2/3 signaling inhibition. J. Formos. Med. Assoc. 2021, 120, 1988–1993. [Google Scholar] [CrossRef]
- Gu, S.; Jin, L.; Zhang, F.; Sarnow, P.; Kay, M.A. Biological basis for restriction of microRNA targets to the 3′ untranslated region in mammalian mRNAs. Nat. Struct. Mol. Biol. 2009, 16, 144–150. [Google Scholar] [CrossRef] [Green Version]
- Kopp, F.; Mendell, J.T. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA Hypothesis: The Rosetta Stone of a Hidden RNA Language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, P.-L.; Chen, S.-H.; Huang, Y.-F.; Lu, M.-Y.; Yu, C.-C. The functional roles of microRNAs in the pathogenesis of oral submucous fibrosis. J. Dent. Sci. 2022, 17, 683–687. [Google Scholar] [CrossRef]
- Yu, C.-C.; Liao, Y.-W.; Hsieh, P.-L.; Chang, Y.-C. Targeting lncRNA H19/miR-29b/COL1A1 Axis Impedes Myofibroblast Activities of Precancerous Oral Submucous Fibrosis. Int. J. Mol. Sci. 2021, 22, 2216. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.Y.; Yu, C.C.; Liao, Y.W.; Hsieh, P.L.; Lu, M.Y.; Lin, K.C.; Wu, C.Z.; Tsai, L.L. LncRNA LINC00974 activates TGF-β/Smad signaling to promote oral fibrogenesis. J. Oral Pathol. Med. 2019, 48, 151–158. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Hsieh, P.-L.; Liao, Y.-W.; Peng, C.-Y.; Yu, C.-C.; Lu, M.-Y. Arctigenin Reduces Myofibroblast Activities in Oral Submucous Fibrosis by LINC00974 Inhibition. Int. J. Mol. Sci. 2019, 20, 1328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ransohoff, J.D.; Wei, Y.; Khavari, P.A. The functions and unique features of long intergenic non-coding RNA. Nat. Rev. Mol. Cell Biol. 2018, 19, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Kwak, H.-H.; Kim, I.-R.; Kim, H.-J.; Park, B.-S.; Yu, S.-B. α-Mangostin Induces Apoptosis and Cell Cycle Arrest in Oral Squamous Cell Carcinoma Cell. Evid.-Based Complement. Altern. Med. 2016, 2016, 5352412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tangsuksan, P.; Chuerduangphui, J.; Takahashi Yupanqui, C.; Srichana, T.; Hitakomate, E.; Pientong, C.; Ekalaksananan, T.; Nittayananta, W. Mucoadhesive film containing α-mangostin shows potential role in oral cancer treatment. BMC Oral Health 2021, 21, 512. [Google Scholar] [CrossRef]
- Li, R.S.; Xu, G.-H.; Cao, J.; Liu, B.; Xie, H.-F.; Ishii, Y.; Zhang, C.-F. Alpha-Mangostin Ameliorates Bleomycin-Induced Pulmonary Fibrosis in Mice Partly through Activating Adenosine 5′-Monophosphate-Activated Protein Kinase. Front. Pharmacol. 2019, 10, 1305. [Google Scholar] [CrossRef]
- Soetikno, V.; Murwantara, A.; Andini, P.; Charlie, F.; Lazarus, G.; Louisa, M.; Arozal, W. Alpha-Mangostin Improves Cardiac Hypertrophy and Fibrosis and Associated Biochemical Parameters in High-Fat/High-Glucose Diet and Low-Dose Streptozotocin Injection-Induced Type 2 Diabetic Rats. J. Exp. Pharmacol. 2020, 12, 27–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lestari, N.; Louisa, M.; Soetikno, V.; Suwana, A.G.; Ramadhan, P.A.; Akmal, T.; Arozal, W. Alpha Mangostin Inhibits the Proliferation and Activation of Acetaldehyde Induced Hepatic Stellate Cells through TGF-β and ERK 1/2 Pathways. J. Toxicol. 2018, 2018, 5360496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sari, N.; Katanasaka, Y.; Sugiyama, Y.; Miyazaki, Y.; Sunagawa, Y.; Funamoto, M.; Shimizu, K.; Shimizu, S.; Hasegawa, K.; Morimoto, T. Alpha Mangostin Derived from Garcinia magostana Linn Ameliorates Cardiomyocyte Hypertrophy and Fibroblast Phenotypes in Vitro. Biol. Pharm. Bull. 2021, 44, 1465–1472. [Google Scholar] [CrossRef] [PubMed]
- Arunkumar, G.; Deva Magendhra Rao, A.K.; Manikandan, M.; Arun, K.; Vinothkumar, V.; Revathidevi, S.; Rajkumar, K.S.; Rajaraman, R.; Munirajan, A.K. Expression profiling of long non-coding RNA identifies linc-RoR as a prognostic biomarker in oral cancer. Tumor Biol. 2017, 39, 1010428317698366. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.-W.; Tsai, L.-L.; Lee, Y.-H.; Hsieh, P.-L.; Yu, C.-C.; Lu, M.-Y. miR-21 promotes the fibrotic properties in oral mucosa through targeting PDCD4. J. Dent. Sci. 2022, 17, 677–682. [Google Scholar] [CrossRef] [PubMed]
- Bell, E.; Ivarsson, B.; Merrill, C. Production of a tissue-like structure by contraction of collagen lattices by human fibroblasts of different proliferative potential in vitro. Proc. Natl. Acad. Sci. USA 1979, 76, 1274–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinz, B.; Celetta, G.; Tomasek, J.J.; Gabbiani, G.; Chaponnier, C. Alpha-Smooth Muscle Actin Expression Upregulates Fibroblast Contractile Activity. Mol. Biol. Cell 2001, 12, 2730–2741. [Google Scholar] [CrossRef] [Green Version]
- Kuo, M.Y.; Chen, H.M.; Hahn, L.J.; Hsieh, C.C.; Chiang, C.-P. Collagen Biosynthesis in Human Oral Submucous Fibrosis Fibroblast Cultures. J. Dent. Res. 1995, 74, 1783–1788. [Google Scholar] [CrossRef]
- Yiemwattana, I.; Kaomongkolgit, R. Alpha-mangostin suppresses IL-6 and IL-8 expression in P. gingivalis LPS-stimulated human gingival fibroblasts. Odontology 2015, 103, 348–355. [Google Scholar] [CrossRef]
- Shih, Y.-H.; Wang, T.-H.; Shieh, T.-M.; Tseng, Y.-H. Oral Submucous Fibrosis: A Review on Etiopathogenesis, Diagnosis, and Therapy. Int. J. Mol. Sci. 2019, 20, 2940. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Fang, Z.; Ling, Y.; Luo, W. LncRNA-H19 acts as a ceRNA to regulate HE4 expression by sponging miR-140 in human umbilical vein endothelial cells under hyperglycemia with or without α-Mangostin. Biomed. Pharmacother. 2019, 118, 109256. [Google Scholar] [CrossRef] [PubMed]
- Loewer, S.; Cabili, M.N.; Guttman, M.; Loh, Y.-H.; Thomas, K.; Park, I.H.; Garber, M.; Curran, M.; Onder, T.; Agarwal, S.; et al. Large intergenic non-coding RNA-RoR modulates reprogramming of human induced pluripotent stem cells. Nat. Genet. 2010, 42, 1113–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Xu, Z.; Jiang, J.; Xu, C.; Kang, J.; Xiao, L.; Wu, M.; Xiong, J.; Guo, X.; Liu, H. Endogenous miRNA Sponge lincRNA-RoR Regulates Oct4, Nanog, and Sox2 in Human Embryonic Stem Cell Self-Renewal. Dev. Cell 2013, 25, 69–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.Y.; Peng, C.-Y.; Lee, S.-S.; Chou, M.-Y.; Yu, C.-C.; Chang, Y.-C. Acquisition cancer stemness, mesenchymal transdifferentiation, and chemoresistance properties by chronic exposure of oral epithelial cells to arecoline. Oncotarget 2016, 7, 84072–84081. [Google Scholar] [CrossRef]
- Lee, Y.-H.; Liao, Y.-W.; Lu, M.-Y.; Hsieh, P.-L.; Yu, C.-C. LINC00084/miR-204/ZEB1 Axis Mediates Myofibroblastic Differentiation Activity in Fibrotic Buccal Mucosa Fibroblasts: Therapeutic Target for Oral Submucous Fibrosis. J. Pers. Med. 2021, 11, 707. [Google Scholar] [CrossRef]
- Yu, C.-H.; Fang, C.-Y.; Yu, C.-C.; Hsieh, P.-L.; Liao, Y.-W.; Tsai, L.-L.; Chu, P.-M. LINC00312/YBX1 Axis Regulates Myofibroblast Activities in Oral Submucous Fibrosis. Int. J. Mol. Sci. 2020, 21, 2979. [Google Scholar] [CrossRef] [Green Version]
- Hou, P.; Zhao, Y.; Li, Z.; Yao, R.; Ma, M.; Gao, Y.; Zhao, L.; Zhang, Y.; Huang, B.; Lu, J. LincRNA-ROR induces epithelial-to-mesenchymal transition and contributes to breast cancer tumorigenesis and metastasis. Cell Death Dis. 2014, 5, e1287. [Google Scholar] [CrossRef] [Green Version]
- Zhan, H.-X.; Wang, Y.; Li, C.; Xu, J.-W.; Zhou, B.; Zhu, J.-K.; Han, H.-F.; Wang, L.; Wang, Y.-S.; Hu, S.-Y. LincRNA-ROR promotes invasion, metastasis and tumor growth in pancreatic cancer through activating ZEB1 pathway. Cancer Lett. 2016, 374, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Tsai, C.H.; Lai, Y.L.; Yu, C.C.; Chi, W.Y.; Li, J.J.; Chang, W.W. Arecoline-induced myofibroblast transdifferentiation from human buccal mucosal fibroblasts is mediated by ZEB 1. J. Cell. Mol. Med. 2014, 18, 698–708. [Google Scholar] [CrossRef]
- Liao, Y.-W.; Yu, C.-C.; Hsieh, P.-L.; Chang, Y.-C. miR-200b ameliorates myofibroblast transdifferentiation in precancerous oral submucous fibrosis through targeting ZEB 2. J. Cell. Mol. Med. 2018, 22, 4130–4138. [Google Scholar] [CrossRef]
- Lee, Y.-H.; Yang, L.-C.; Hu, F.-W.; Peng, C.-Y.; Yu, C.-H.; Yu, C.-C. Elevation of Twist expression by arecoline contributes to the pathogenesis of oral submucous fibrosis. J. Formos. Med. Assoc. 2016, 115, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wu, W.; Sun, Q.; Ye, L.; Zhou, D.; Wang, W. linc-ROR facilitates hepatocellular carcinoma resistance to doxorubicin by regulating TWIST1-mediated epithelial-mesenchymal transition. Mol. Med. Rep. 2020, 23, 340. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Wang, Y.; Wang, D.; Zhou, H.; Zhang, H.; Li, L. miR-145-5p attenuates hypertrophic scar via reducing Smad2/Smad3 expression. Biochem. Biophys. Res. Commun. 2019, 521, 1042–1048. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.-Y.; Chen, S.-H.; Huang, C.-C.; Liao, Y.-W.; Chao, S.-C.; Yu, C.-C. Fucoidan-Mediated Inhibition of Fibrotic Properties in Oral Submucous Fibrosis via the MEG3/miR-181a/Egr1 Axis. Pharmaceuticals 2022, 15, 833. [Google Scholar] [CrossRef]
- Brockhausen, J.; Tay, S.S.; Grzelak, C.A.; Bertolino, P.; Bowen, D.G.; D’Avigdor, W.M.; Teoh, N.; Pok, S.; Shackel, N.; Gamble, J.R.; et al. miR-181a mediates TGF-β-induced hepatocyte EMT and is dysregulated in cirrhosis and hepatocellular cancer. Liver Int. 2015, 35, 240–253. [Google Scholar] [CrossRef]
- Xiao, Z.; Reddy, D.P.K.; Xue, C.; Liu, X.; Chen, X.; Li, J.; Ling, X.; Zheng, S. Profiling of miR-205/P4HA3 following Angiotensin II-Induced Atrial Fibrosis: Implications for Atrial Fibrillation. Front. Cardiovasc. Med. 2021, 8, 609300. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.-H.; Hsieh, P.-L.; Chao, S.-C.; Liao, Y.-W.; Liu, C.-M.; Yu, C.-C. α-Mangostin Inhibits the Activation of Myofibroblasts via Downregulation of Linc-ROR-Mediated TGFB1/Smad Signaling. Nutrients 2023, 15, 1321. https://doi.org/10.3390/nu15061321
Lee Y-H, Hsieh P-L, Chao S-C, Liao Y-W, Liu C-M, Yu C-C. α-Mangostin Inhibits the Activation of Myofibroblasts via Downregulation of Linc-ROR-Mediated TGFB1/Smad Signaling. Nutrients. 2023; 15(6):1321. https://doi.org/10.3390/nu15061321
Chicago/Turabian StyleLee, Yu-Hsien, Pei-Ling Hsieh, Shih-Chi Chao, Yi-Wen Liao, Chia-Ming Liu, and Cheng-Chia Yu. 2023. "α-Mangostin Inhibits the Activation of Myofibroblasts via Downregulation of Linc-ROR-Mediated TGFB1/Smad Signaling" Nutrients 15, no. 6: 1321. https://doi.org/10.3390/nu15061321
APA StyleLee, Y. -H., Hsieh, P. -L., Chao, S. -C., Liao, Y. -W., Liu, C. -M., & Yu, C. -C. (2023). α-Mangostin Inhibits the Activation of Myofibroblasts via Downregulation of Linc-ROR-Mediated TGFB1/Smad Signaling. Nutrients, 15(6), 1321. https://doi.org/10.3390/nu15061321