The Association between Caffeine Intake and the Colonic Mucosa-Associated Gut Microbiota in Humans—A Preliminary Investigation

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
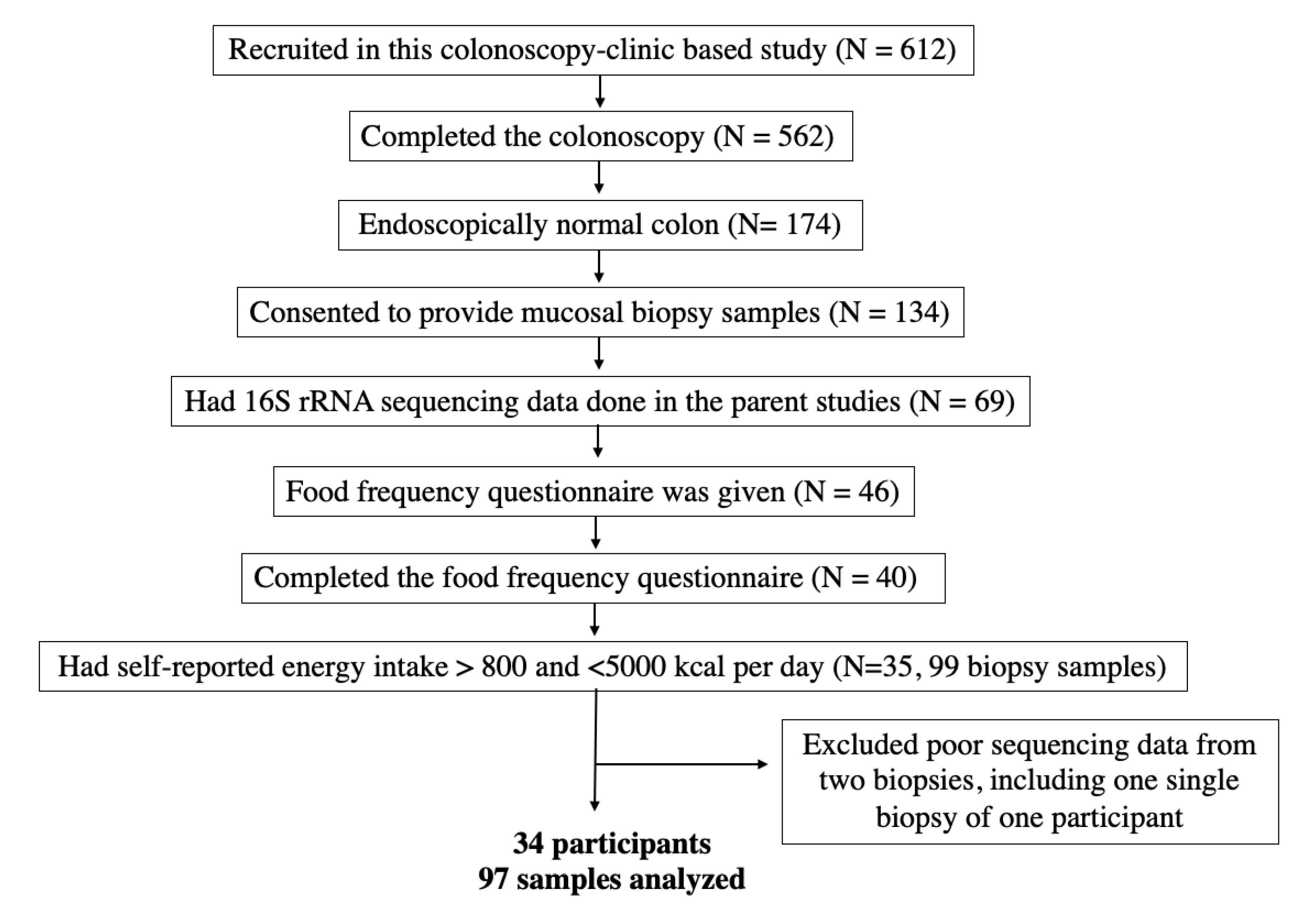
2.1. Study Participants
2.2. Data Collection
2.3. Colonoscopy and Biopsy Requirement
2.4. Microbial DNA Extraction and 16S rRNA Gene Sequencing
2.5. Bioinformatics and Taxonomic Assignment
2.6. Statistical Analysis
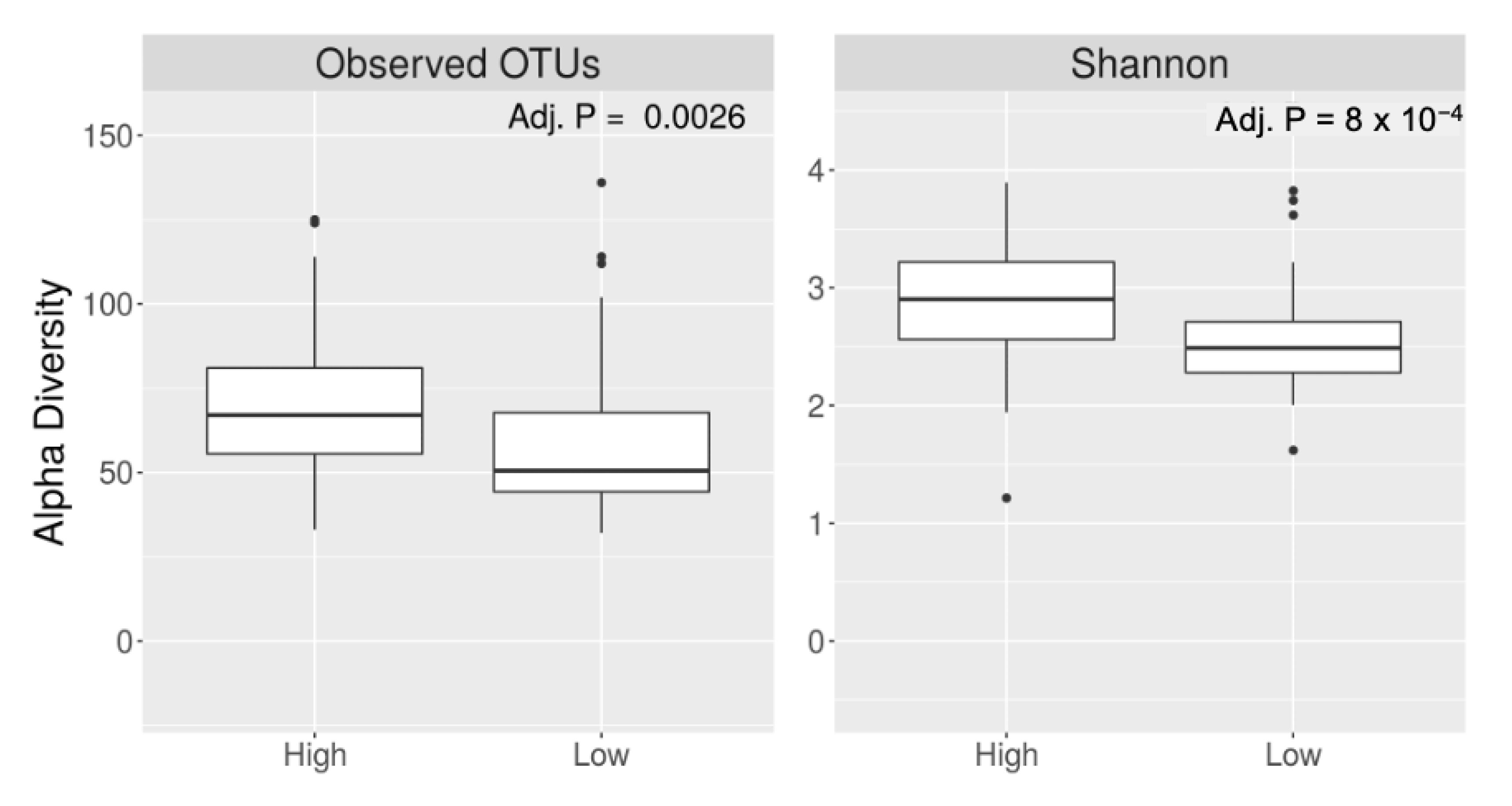
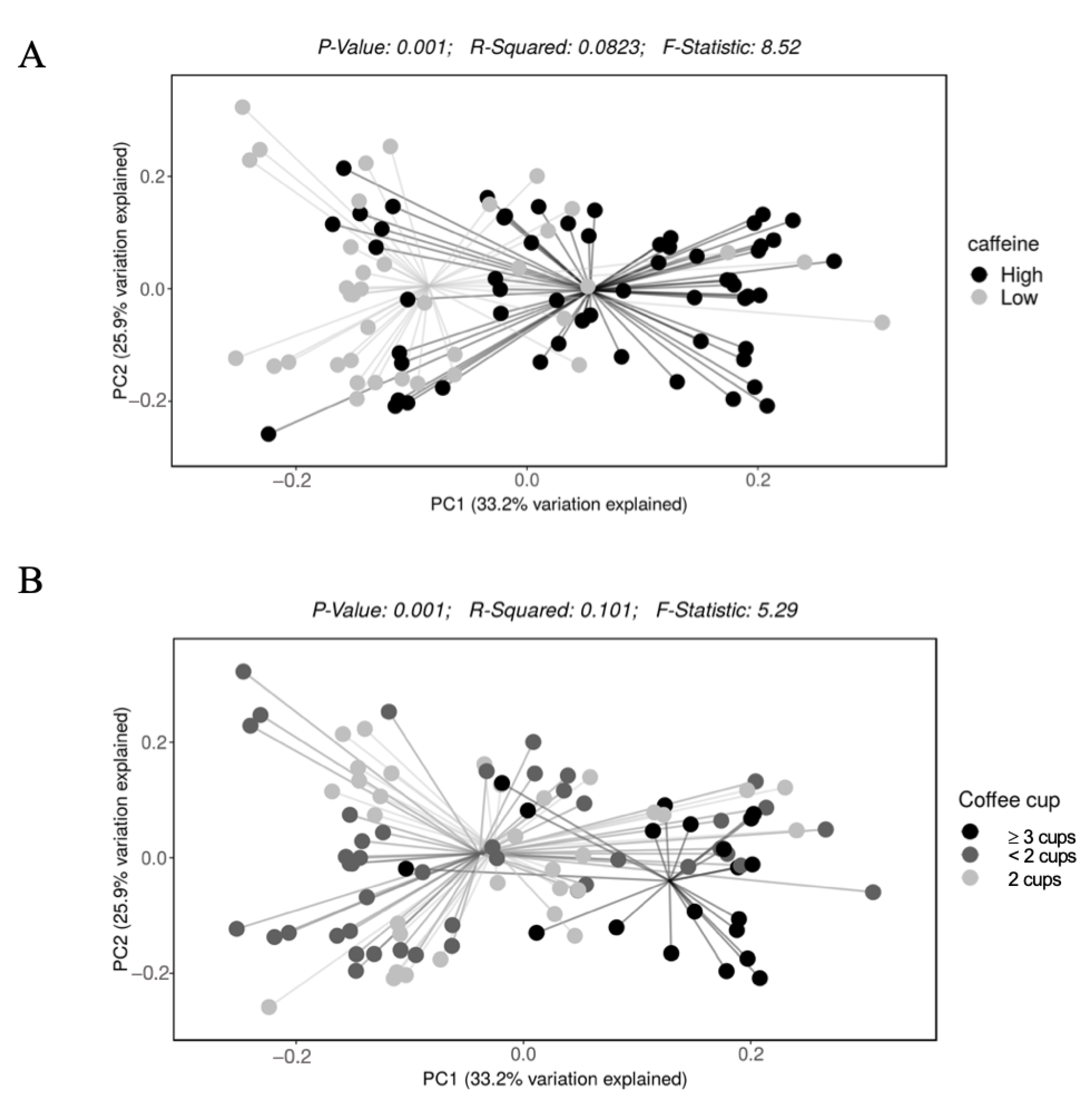
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grosso, G.; Godos, J.; Galvano, F.; Giovannucci, E.L. Coffee, Caffeine, and Health Outcomes: An Umbrella Review. Annu. Rev. Nutr. 2017, 37, 131–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.; Zhang, D.; Jiang, W. Coffee and caffeine intake and incidence of type 2 diabetes mellitus: A meta-analysis of prospective studies. Eur. J. Nutr. 2014, 53, 25–38. [Google Scholar] [CrossRef]
- Almofarreh, A.; Sheerah, H.A.; Arafa, A.; Ahamed, S.S.; Alzeer, O.; Al-Hunaishi, W.; Mhimed, M.M.; Al-Hazmi, A.; Lim, S.H. Beverage Consumption and Ulcerative Colitis: A Case-Control Study from Saudi Arabia. Int. J. Environ. Res. Public Health 2022, 19, 2287. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.A.; Low, D.; Kamba, A.; Llado, V.; Mizoguchi, E. Oral caffeine administration ameliorates acute colitis by suppressing chitinase 3-like 1 expression in intestinal epithelial cells. J. Gastroenterol. 2014, 49, 1206–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barre, T.; Fontaine, H.; Ramier, C.; Di Beo, V.; Pol, S.; Carrieri, P.; Marcellin, F.; Cagnot, C.; Dorival, C.; Zucman-Rossi, J.; et al. Elevated coffee consumption is associated with a lower risk of elevated liver fibrosis biomarkers in patients treated for chronic hepatitis B (ANRS CO22 Hepather cohort). Clin. Nutr. 2022, 41, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Shan, L.; Wang, F.; Zhai, D.; Meng, X.; Liu, J.; Lv, X. Caffeine in liver diseases: Pharmacology and toxicology. Front. Pharmacol. 2022, 13, 1030173. [Google Scholar] [CrossRef]
- Schmit, S.L.; Rennert, H.S.; Rennert, G.; Gruber, S.B. Coffee Consumption and the Risk of Colorectal Cancer. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2016, 25, 634–639. [Google Scholar] [CrossRef] [Green Version]
- Fiani, B.; Zhu, L.; Musch, B.L.; Briceno, S.; Andel, R.; Sadeq, N.; Ansari, A.Z. The Neurophysiology of Caffeine as a Central Nervous System Stimulant and the Resultant Effects on Cognitive Function. Cureus 2021, 13, e15032. [Google Scholar] [CrossRef]
- Nishitsuji, K.; Watanabe, S.; Xiao, J.; Nagatomo, R.; Ogawa, H.; Tsunematsu, T.; Umemoto, H.; Morimoto, Y.; Akatsu, H.; Inoue, K.; et al. Effect of coffee or coffee components on gut microbiome and short-chain fatty acids in a mouse model of metabolic syndrome. Sci. Rep. 2018, 8, 16173. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, S.; Salazar, N.; Ruiz-Saavedra, S.; Gomez-Martin, M.; de Los Reyes-Gavilan, C.G.; Gueimonde, M. Long-Term Coffee Consumption is Associated with Fecal Microbial Composition in Humans. Nutrients 2020, 12, 1287. [Google Scholar] [CrossRef]
- Jaquet, M.; Rochat, I.; Moulin, J.; Cavin, C.; Bibiloni, R. Impact of coffee consumption on the gut microbiota: A human volunteer study. Int. J. Food Microbiol. 2009, 130, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Heckman, M.A.; Weil, J.; Gonzalez de Mejia, E. Caffeine (1, 3, 7-trimethylxanthine) in foods: A comprehensive review on consumption, functionality, safety, and regulatory matters. J. Food Sci. 2010, 75, R77–R87. [Google Scholar] [CrossRef] [PubMed]
- Xu, A.A.; Kennedy, L.K.; Hoffman, K.; White, D.L.; Kanwal, F.; El-Serag, H.B.; Petrosino, J.F.; Jiao, L. Dietary Fatty Acid Intake and the Colonic Gut Microbiota in Humans. Nutrients 2022, 14, 2722. [Google Scholar] [CrossRef] [PubMed]
- Subar, A.F.; Thompson, F.E.; Kipnis, V.; Midthune, D.; Hurwitz, P.; McNutt, S.; McIntosh, A.; Rosenfeld, S. Comparative validation of the Block, Willett, and National Cancer Institute food frequency questionnaires: The Eating at America’s Table Study. Am. J. Epidemiol. 2001, 154, 1089–1099. [Google Scholar] [CrossRef] [Green Version]
- Reedy, J.; Lerman, J.L.; Krebs-Smith, S.M.; Kirkpatrick, S.I.; Pannucci, T.E.; Wilson, M.M.; Subar, A.F.; Kahle, L.L.; Tooze, J.A. Evaluation of the Healthy Eating Index-2015. J. Acad. Nutr. Diet. 2018, 118, 1622–1633. [Google Scholar] [CrossRef]
- The Human Microbiome Project Consortium. A framework for human microbiome research. Nature 2012, 486, 215–221. [Google Scholar] [CrossRef] [Green Version]
- The Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Barone, J.J.; Roberts, H.R. Caffeine consumption. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 1996, 34, 119–129. [Google Scholar] [CrossRef]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. UniFrac: An effective distance metric for microbial community comparison. ISME J. 2011, 5, 169–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurwara, S.; Ajami, N.J.; Jang, A.; Hessel, F.C.; Chen, L.; Plew, S.; Wang, Z.; Graham, D.Y.; Hair, C.; White, D.L.; et al. Dietary Nutrients Involved in One-Carbon Metabolism and Colonic Mucosa-Associated Gut Microbiome in Individuals with an Endoscopically Normal Colon. Nutrients 2019, 11, 613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Yekutieli, D. Quantitative trait Loci analysis using the false discovery rate. Genetics 2005, 171, 783–790. [Google Scholar] [CrossRef] [Green Version]
- Zhernakova, A.; Kurilshikov, A.; Bonder, M.J.; Tigchelaar, E.F.; Schirmer, M.; Vatanen, T.; Mujagic, Z.; Vila, A.V.; Falony, G.; Vieira-Silva, S.; et al. Population-based metagenomics analysis reveals markers for gut microbiome composition and diversity. Science 2016, 352, 565–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, J.; Sinha, R.; Pei, Z.; Dominianni, C.; Wu, J.; Shi, J.; Goedert, J.J.; Hayes, R.B.; Yang, L. Human gut microbiome and risk for colorectal cancer. J. Natl. Cancer Inst. 2013, 105, 1907–1911. [Google Scholar] [CrossRef] [Green Version]
- Kostic, A.D.; Xavier, R.J.; Gevers, D. The microbiome in inflammatory bowel disease: Current status and the future ahead. Gastroenterology 2014, 146, 1489–1499. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Xie, Q.; Kong, P.; Liu, L.; Sun, S.; Xiong, B.; Huang, B.; Yan, L.; Sheng, J.; Xiang, H. Polyphenol- and Caffeine-Rich Postfermented Pu-erh Tea Improves Diet-Induced Metabolic Syndrome by Remodeling Intestinal Homeostasis in Mice. Infect. Immun. 2018, 86, e00601-17. [Google Scholar] [CrossRef] [Green Version]
- Rios-Covian, D.; Gueimonde, M.; Duncan, S.H.; Flint, H.J.; de los Reyes-Gavilan, C.G. Enhanced butyrate formation by cross-feeding between Faecalibacterium prausnitzii and Bifidobacterium adolescentis. FEMS Microbiol. Lett. 2015, 362, fnv176. [Google Scholar] [CrossRef] [Green Version]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Backhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermudez-Humaran, L.G.; Gratadoux, J.J.; Blugeon, S.; Bridonneau, C.; Furet, J.P.; Corthier, G.; et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl. Acad. Sci. USA 2008, 105, 16731–16736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palleja, A.; Kashani, A.; Allin, K.H.; Nielsen, T.; Zhang, C.; Li, Y.; Brach, T.; Liang, S.; Feng, Q.; Jorgensen, N.B.; et al. Roux-en-Y gastric bypass surgery of morbidly obese patients induces swift and persistent changes of the individual gut microbiota. Genome Med. 2016, 8, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alipour, M.; Zaidi, D.; Valcheva, R.; Jovel, J.; Martinez, I.; Sergi, C.; Walter, J.; Mason, A.L.; Wong, G.K.; Dieleman, L.A.; et al. Mucosal Barrier Depletion and Loss of Bacterial Diversity are Primary Abnormalities in Paediatric Ulcerative Colitis. J. Crohn’s Colitis 2016, 10, 462–471. [Google Scholar] [CrossRef]
- Milani, C.; Ticinesi, A.; Gerritsen, J.; Nouvenne, A.; Lugli, G.A.; Mancabelli, L.; Turroni, F.; Duranti, S.; Mangifesta, M.; Viappiani, A.; et al. Gut microbiota composition and Clostridium difficile infection in hospitalized elderly individuals: A metagenomic study. Sci. Rep. 2016, 6, 25945. [Google Scholar] [CrossRef] [Green Version]
- Saulnier, D.M.; Riehle, K.; Mistretta, T.A.; Diaz, M.A.; Mandal, D.; Raza, S.; Weidler, E.M.; Qin, X.; Coarfa, C.; Milosavljevic, A.; et al. Gastrointestinal microbiome signatures of pediatric patients with irritable bowel syndrome. Gastroenterology 2011, 141, 1782–1791. [Google Scholar] [CrossRef] [Green Version]
- Woting, A.; Pfeiffer, N.; Loh, G.; Klaus, S.; Blaut, M. Clostridium ramosum promotes high-fat diet-induced obesity in gnotobiotic mouse models. mBio 2014, 5, e01530-14. [Google Scholar] [CrossRef] [Green Version]
- Smith-Brown, P.; Morrison, M.; Krause, L.; Davies, P.S. Dairy and plant based food intakes are associated with altered faecal microbiota in 2 to 3 year old Australian children. Sci. Rep. 2016, 6, 32385. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; DiBaise, J.K.; Zuccolo, A.; Kudrna, D.; Braidotti, M.; Yu, Y.; Parameswaran, P.; Crowell, M.D.; Wing, R.; Rittmann, B.E.; et al. Human gut microbiota in obesity and after gastric bypass. Proc. Natl. Acad. Sci. USA 2009, 106, 2365–2370. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Luo, X.; Mao, X.; Tao, Y.; Ran, X.; Zhao, H.; Xiong, J.; Li, L. Gut microbiome analysis of type 2 diabetic patients from the Chinese minority ethnic groups the Uygurs and Kazaks. PLoS ONE 2017, 12, e0172774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Wang, J.; Zhou, Y.; Han, H.; Liu, W.; Li, D.; Li, F.; Cao, D.; Lei, Q. Integrated omics analysis reveals differences in gut microbiota and gut-host metabolite profiles between obese and lean chickens. Poult. Sci. 2022, 101, 102165. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Liu, H.; Wu, Y.; Hu, N.; Lei, M.; Zhang, Y.; Wang, S. Intervention with the crude polysaccharides of Physalis pubescens L. mitigates colitis by preventing oxidative damage, aberrant immune responses, and dysbacteriosis. J. Food Sci. 2020, 85, 2596–2607. [Google Scholar] [CrossRef] [PubMed]
- Dinh, D.M.; Volpe, G.E.; Duffalo, C.; Bhalchandra, S.; Tai, A.K.; Kane, A.V.; Wanke, C.A.; Ward, H.D. Intestinal microbiota, microbial translocation, and systemic inflammation in chronic HIV infection. J. Infect. Dis. 2015, 211, 19–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zorkina, Y.A.; Syunyakov, T.S.; Abramova, O.V.; Yunes, R.A.; Averina, O.V.; Kovtun, A.S.; Angelova, I.Y.; Khobta, E.B.; Susloparova, D.A.; Pavlichenko, A.V.; et al. Effects of diet on the gut microbiome in patients with depression. Zhurnal Nevrol. I Psikhiatrii Im. S.S. Korsakova 2022, 122, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.; Sun, Y.; Pan, D.; Sang, L.X.; Sun, M.J.; Li, Y.L.; Chang, B. Distinctive gut microbial dysbiosis between chronic alcoholic fatty liver disease and metabolic-associated fatty liver disease in mice. Exp. Ther. Med. 2021, 21, 418. [Google Scholar] [CrossRef]
- Chen, W.; Liu, F.; Ling, Z.; Tong, X.; Xiang, C. Human intestinal lumen and mucosa-associated microbiota in patients with colorectal cancer. PLoS ONE 2012, 7, e39743. [Google Scholar] [CrossRef]
- Kaakoush, N.O. Insights into the Role of Erysipelotrichaceae in the Human Host. Front. Cell. Infect. Microbiol. 2015, 5, 84. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Ma, Y.; Fang, C.; Peng, Z.; Gao, F.; Moll, J.M.; Qin, S.; Yu, Q.; Hou, Y.; Kristiansen, K.; et al. Genomic and microbial factors affect the prognosis of anti-pd-1 immunotherapy in nasopharyngeal carcinoma. Front. Oncol. 2022, 12, 953884. [Google Scholar] [CrossRef]
- Holzhausen, E.A.; Malecki, K.C.; Sethi, A.K.; Gangnon, R.; Cadmus-Bertram, L.; Deblois, C.L.; Suen, G.; Safdar, N.; Peppard, P.E. Assessing the relationship between physical activity and the gut microbiome in a large, population-based sample of Wisconsin adults. PLoS ONE 2022, 17, e0276684. [Google Scholar] [CrossRef]
- Kim, Y.; Je, Y. Moderate coffee consumption is inversely associated with the metabolic syndrome in the Korean adult population. Br. J. Nutr. 2018, 120, 1279–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordestgaard, A.T.; Thomsen, M.; Nordestgaard, B.G. Coffee intake and risk of obesity, metabolic syndrome and type 2 diabetes: A Mendelian randomization study. Int. J. Epidemiol. 2015, 44, 551–565. [Google Scholar] [CrossRef] [PubMed]
- Shang, F.; Li, X.; Jiang, X. Coffee consumption and risk of the metabolic syndrome: A meta-analysis. Diabetes Metab. 2016, 42, 80–87. [Google Scholar] [CrossRef]
- de Melo Pereira, G.V.; de Carvalho Neto, D.P.; Magalhaes Junior, A.I.; do Prado, F.G.; Pagnoncelli, M.G.B.; Karp, S.G.; Soccol, C.R. Chemical composition and health properties of coffee and coffee by-products. Adv. Food Nutr. Res. 2020, 91, 65–96. [Google Scholar] [CrossRef] [PubMed]
- Mills, C.E.; Tzounis, X.; Oruna-Concha, M.J.; Mottram, D.S.; Gibson, G.R.; Spencer, J.P. In vitro colonic metabolism of coffee and chlorogenic acid results in selective changes in human faecal microbiota growth. Br. J. Nutr. 2015, 113, 1220–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansour, A.; Mohajeri-Tehrani, M.R.; Karimi, S.; Sanginabadi, M.; Poustchi, H.; Enayati, S.; Asgarbeik, S.; Nasrollahzadeh, J.; Hekmatdoost, A. Short term effects of coffee components consumption on gut microbiota in patients with non-alcoholic fatty liver and diabetes: A pilot randomized placebo-controlled, clinical trial. EXCLI J. 2020, 19, 241–250. [Google Scholar] [CrossRef]
- Dong, L.N.; Wang, J.P.; Liu, P.; Yang, Y.F.; Feng, J.; Han, Y. Faecal and mucosal microbiota in patients with functional gastrointestinal disorders: Correlation with toll-like receptor 2/toll-like receptor 4 expression. World J. Gastroenterol. 2017, 23, 6665–6673. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Characteristics Mean ± Standard Deviation or n (%) | Low Caffeine < Median (n = 17) | High Caffeine ≥ Median (n = 17) | p Value |
|---|---|---|---|
| Caffeine (mg) | 39.2 ± 6.4 | 138.9 ± 13.9 | <0.0001 |
| Age (years) | 61.7 ± 1.3 | 62.2 ± 1.5 | 0.78 |
| Men, n (%) | 17 (100%) | 16 (94%) | 1.00 |
| Racial Group | |||
| Non-Hispanic white, n (%) | 14 (82.4%) | 14 (82.4%) | 1.00 |
| Body mass index (kg/m2) | 32.6 ± 1.5 | 35.2 ± 1.6 | 0.25 |
| Smoking status, n (%) | 0.55 | ||
| Never smokers | 7 (41.2%) | 6 (35.3%) | |
| Former smokers | 8 (47.1%) | 6 (35.3%) | |
| Current smokers | 2 (11.8%) | 5 (29.4%) | |
| Alcohol Status, n (%) | 0.68 | ||
| Never drinkers | 4 (23.5%) | 5 (29.4%) | |
| Former drinkers | 4 (23.5%) | 6 (35.3%) | |
| Current drinker | 9 (53%) | 6 (35.3%) | |
| Hypertension, yes, n (%) | 12 (70.6%) | 13 (76.5%) | 1.00 |
| Diabetes, yes, n (%) | 8 (47.1%) | 9 (52.9%) | 1.00 |
| Daily total calorie intake (kcal) | 2039 (±557) | 1748 (±813) | 0.23 |
| Total carbohydrate (grams/1000 kcal/day) | 116 (±23.5) | 111 (±17.5) | 0.53 |
| Total protein (grams/1000 kcal/day) | 36.4 (±5.78) | 40.0 (±8.37) | 0.16 |
| Total fat (grams/1000 kcal/day) | 41.0 (±8.78) | 43.4 (±4.96) | 0.33 |
| Vitamin B2 (mg/1000 kcal/day) | 0.89 (±0.20) | 1.28 (± 0.30) | 0.0001 |
| Vitamin B6 (mg/1000 kcal/day) | 0.82 (±0.16) | 1.02 (± 0.34) | 0.02 |
| Vitamin B12 (mcg/1000 kcal/day) | 2.20 (±0.23) | 2.76 (± 0.98) | 0.06 |
| HEI score 1 | 60.5 (±2.0) | 61.4 (±2.4) | 0.77 |
| Lower Caffeine | Higher Caffeine | <2 Cups Coffee | 2 Cups Coffee | ≥3 Cups Coffee | |||
|---|---|---|---|---|---|---|---|
| Bacterial Genus | Relative Abundance (%) | q Value | Relative Abundance (%) | q Value | |||
| Erysipelatoclostridium | 3.14 | 0.10 | <0.0001 | 1.42 | 1.79 | 0.19 | 0.22 |
| Faecalibacterium | 4.29 | 9.54 | 0.0003 | 5.66 | 5.26 | 15.16 | <0.0001 |
| Lachnospiraceae (ASV0006) | 4.88 | 1.58 | 0.0007 | 3.46 | 2.56 | 2.03 | 0.33 |
| Alistipes | 0.57 | 1.32 | 0.01 | 0.46 | 1.31 | 1.84 | <0.0001 |
| Subdoligranulum | 0.10 | 0.76 | <0.0001 | 0.15 | 1.13 | 0.29 | <0.0001 |
| Sutterella | 1.96 | 1.59 | 0.89 | 1.54 | 2.21 | 1.41 | 0.02 |
| Prevotella | 1.39 | 3.41 | 0.03 | 2.40 | 3.64 | 1.50 | 0.16 |
| Genera | IRR (95% CI) 1 | IRR (95% CI) 2 | IRR (95% CI) 3 |
|---|---|---|---|
| Caffeine | |||
| Alistipes | 3.17 (1.16–8.66) | 3.05 (1.10–8.48) | 1.40 (0.32–6.19) |
| Faecalibacterium | 5.56 (2.84–10.9) | 5.28 (2.68–10.4) | 2.10 (0.85–5.20) |
| Erysipelatoclostridium | 0.07 (0.02–0.25) | 0.07 (0.02–0.25) | 0.02 (0.003–0.17) |
| Subdoligranulum | 0.85(0.49–1.48) | 0.86 (0.52–1.42) | 1.17 (0.57–2.42) |
| Coffee | |||
| Alistipes | 2.86 (1.37–5.97) | 2.84 (1.38–5.84) | 2.20 (1.00–4.89) |
| Faecalibacterium | 2.39 (1.53–3.73) | 2.35 (1.50–3.68) | 1.49 (0.87–2.56) |
| Erysipelatoclostridium | 0.26 (0.08–0.84) | 0.24 (0.07–0.84) | 0.31(0.07–1.36) |
| Subdoligranulum | 1.08 (0.74–1.57) | 0.97 (0.66–1.42) | 1.10 (0.73–1.66) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, A.; Hoffman, K.; Xu, A.A.; Gurwara, S.; White, D.L.; Kanwal, F.; Jang, A.; El-Serag, H.B.; Petrosino, J.F.; Jiao, L. The Association between Caffeine Intake and the Colonic Mucosa-Associated Gut Microbiota in Humans—A Preliminary Investigation. Nutrients 2023, 15, 1747. https://doi.org/10.3390/nu15071747
Dai A, Hoffman K, Xu AA, Gurwara S, White DL, Kanwal F, Jang A, El-Serag HB, Petrosino JF, Jiao L. The Association between Caffeine Intake and the Colonic Mucosa-Associated Gut Microbiota in Humans—A Preliminary Investigation. Nutrients. 2023; 15(7):1747. https://doi.org/10.3390/nu15071747
Chicago/Turabian StyleDai, Annie, Kristi Hoffman, Anthony A. Xu, Shawn Gurwara, Donna L. White, Fasiha Kanwal, Albert Jang, Hashem B. El-Serag, Joseph F. Petrosino, and Li Jiao. 2023. "The Association between Caffeine Intake and the Colonic Mucosa-Associated Gut Microbiota in Humans—A Preliminary Investigation" Nutrients 15, no. 7: 1747. https://doi.org/10.3390/nu15071747
APA StyleDai, A., Hoffman, K., Xu, A. A., Gurwara, S., White, D. L., Kanwal, F., Jang, A., El-Serag, H. B., Petrosino, J. F., & Jiao, L. (2023). The Association between Caffeine Intake and the Colonic Mucosa-Associated Gut Microbiota in Humans—A Preliminary Investigation. Nutrients, 15(7), 1747. https://doi.org/10.3390/nu15071747