
Relationship of Low Vitamin B6 Status with Sarcopenia, Frailty, and Mortality: A Narrative Review
 ,
,
Abstract
:1. Introduction
2. Epidemiological Studies
2.1. Method of Literature Survey
2.2. Relatioship of B6 Status with Sarcopenia
2.3. Relationship of B6 Status with Frailty
2.4. Relationship of B6 Status with Mortality
2.5. Relationship of Other Vitamin Statuses with Sarcopenia, Frailty, and Mortality
2.6. B6 Status and Homocysteine
2.7. Intervention Studies on the Effects of Supplemental B6 Treatment
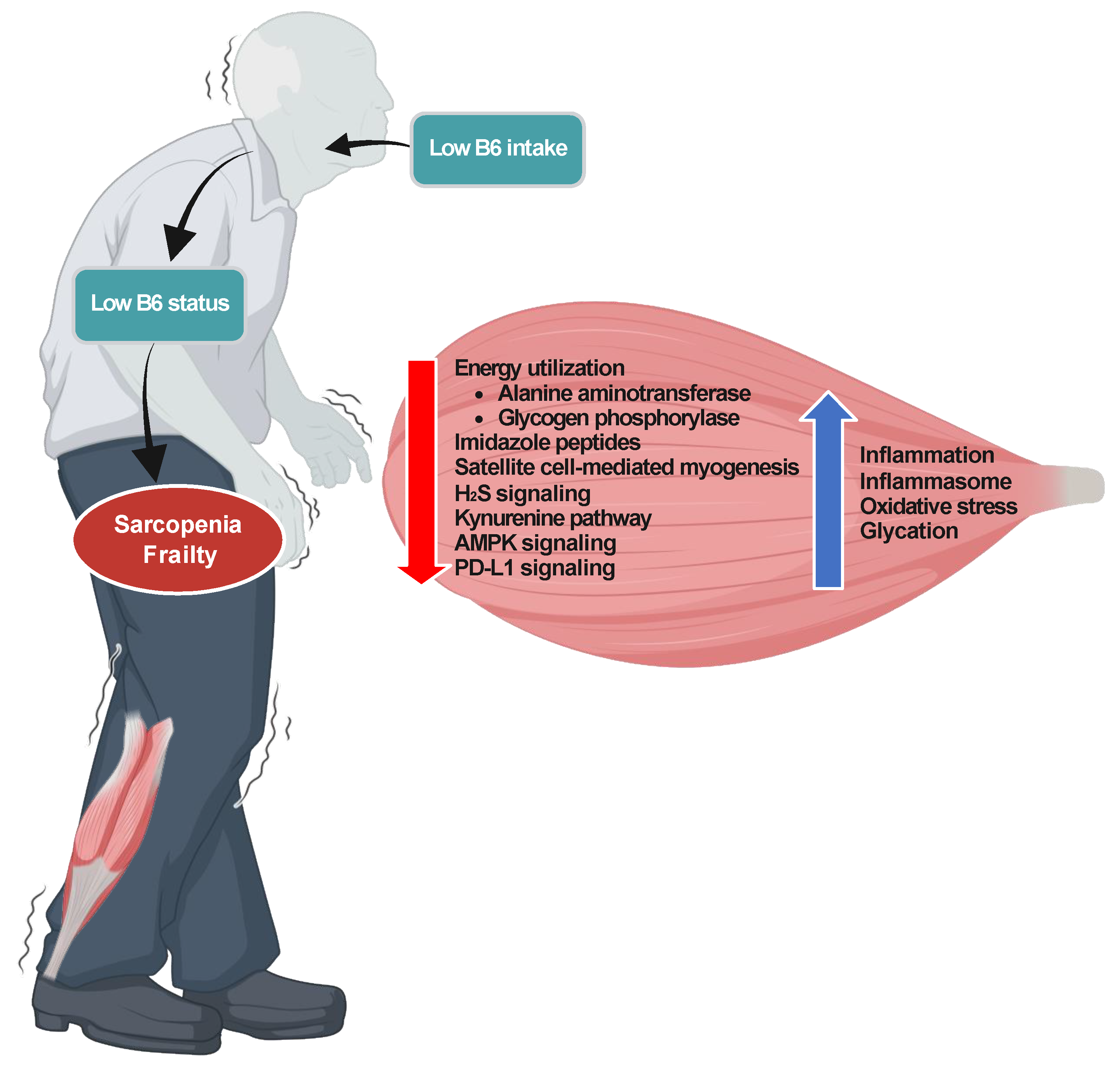
3. Potential Mechanisms Underlying the Association of Low B6 Status with Sarcopenia, Frailty, and All-Cause Mortality
3.1. Energy Utilization
3.2. Growth of Skeletal Muscles
3.3. Imidazole Peptides
3.4. Satellite Cells
3.5. Inflammasome and Inflammation
3.6. PD-L1
3.7. Musculoskeletal Aging with Low B6 Status and Other Age-Related Disorders
4. Outlook for Future Research
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| B6 | vitamin B6 |
| PLP | pyridoxal 5′-phosphate |
| PN | pyridoxine |
| PL | pyridoxal |
| PA | pyridoxic acid |
| HK | 3-hydroxykynurenine |
| XA | xanthurenic acid |
| OR | odds ratio |
| HR | hazard ratio |
| CI | confidence interval |
| 25(OH)D | 25-hydroxyvitamin D |
| T2DM | type 2 diabetic mellitus |
| COPD | chronic obstructive pulmonary disease |
| B1 | vitamin B1 |
| B2 | vitamin B2 |
| B12 | vitamin B12 |
| CVD | cardiovascular disease |
| CHD | coronary heart disease |
| NHANES | National Health and Nutrition Examination Survey |
| PAr index | (pyridoxic acid)/(pyridoxal + pyridoxal 5′-phosphate) index |
| SD | standard deviation |
| IL-6 | interleukin-6 |
| hs-CRP | high sensitive C-reactive protein |
| RTR | renal transplant recipient |
| A | vitamin A |
| C | vitamin C |
| E | vitamin E |
| RDA | recommended dietary allowance |
| ALT | alanine aminotransferase |
| BCAA | branched chain amino acids |
| IGF1 | insulin-like growth factor-1 |
| Nrf2 | NF-E2-related factor 2 |
| HSP | heat shock protein |
| AKT | Akt serine/threonine kinase (also called protein kinase B) |
| mTOR | mammalian target of rapamycin |
| 4EBP1 | eukaryotic initiation factor 4E binding protein 1 |
| S6K | p70 S6K kinase |
| L-carnosine | β-alanyl-L-histidine |
| L-anserine | β-alanyl-3-methylhistidine |
| MAPK | mitogen-activated protein kinase |
| NLRP3 | nucleotide-binding oligomerization domain-like receptor family, pyrin domain-containing 3 |
| NF-κB | nuclear factor-kappa B |
| LPS | lipopolysaccharide |
| AMPK | AMP-activated protein kinase |
| Sirt1 | NAD+-dependent histone deacetylase (also called Sirtuin 1) |
| PD-L1 | programmed death-ligand 1 |
| PD-1 | programmed cell death protein 1 |
References
- Zhang, P.; Suda, T.; Suidasari, S.; Kumrungsee, T.; Yanaka, N.; Kato, N. Chapter 15—Novel preventive mechanisms of vitamin B6 against inflammation, inflammasome, and chronic diseases. In Molecular Nutrition, Vitamins; Vinood, B.P., Ed.; Academic Press: Cambride, MA, USA, 2020; pp. 283–299. [Google Scholar] [CrossRef]
- Kumrungsee, T.; Zhang, P.; Yanaka, N.; Suda, T.; Kato, N. Emerging cardioprotective mechanisms of vitamin B6: A narrative review. Eur. J. Nutr. 2022, 61, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Troesch, B.; Hoeft, B.; McBurney, M.; Eggersdorfer, M.; Weber, P. Dietary surveys indicate vitamin intakes below recommendations are common in representative Western countries. Br. J. Nutr. 2012, 108, 692–698. [Google Scholar] [CrossRef]
- Ho, C.-L.; Quay, T.A.W.; Devlin, A.M.; Lamers, Y. Prevalence and Predictors of Low Vitamin B6 Status in Healthy Young Adult Women in Metro Vancouver. Nutrients 2016, 8, 538. [Google Scholar] [CrossRef] [PubMed]
- The Department of Economic and Social Affairs of the United Nations. World Social Report 2023: Leaving No One behind in an Aging World; United Nations: New York, NY, USA, 2023. [Google Scholar]
- Chang, K.-V.; Hsu, T.-H.; Wu, W.-T.; Huang, W.-T.; Han, D.-S. Association Between Sarcopenia and Cognitive Impairment: A Systematic Review and Meta-Analysis. Review. J. Am. Med. Dir. Assoc. 2016, 17, 1164.e7–1164.e15. [Google Scholar] [CrossRef] [PubMed]
- Gielen, E.; Dupont, J.; Dejaeger, M.; Laurent, M.R. Sarcopenia, osteoporosis and frailty. Metabolism 2023, 145, 155638. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Shiraishi, R.; Nakayama, Y.; Taira, Y. Can Nutrition Contribute to a Reduction in Sarcopenia, Frailty, and Comorbidities in a Super-Aged Society? Nutrients 2023, 15, 2991. [Google Scholar] [CrossRef] [PubMed]
- Welch, A. Micronutrient malnutrition across the life course, sarcopenia and frailty. Proc. Nutr. Soc. 2021, 80, 279–282. [Google Scholar] [CrossRef]
- Struijk, E.A.; Lana, A.; Guallar-Castillon, P.; Rodríguez-Artalejo, F.; Lopez-Garcia, E. Intake of B vitamins and impairment in physical function in older adults. Clin. Nutr. 2018, 37, 1271–1278. [Google Scholar] [CrossRef]
- Grootswagers, P.; Mensink, M.; Berendsen, A.A.M.; Deen, C.P.J.; Kema, I.P.; Bakker, S.J.L.; Santoro, A.; Franceschi, C.; Meunier, N.; Malpuech-Brugère, C.; et al. Vitamin B-6 intake is related to physical performance in European older adults: Results of the New Dietary Strategies Addressing the Specific Needs of the Elderly Population for Healthy Aging in Europe (NU-AGE) study. Am. J. Clin. Nutr. 2021, 113, 781–789. [Google Scholar] [CrossRef]
- Park, S.-J.; Park, J.; Won, C.W.; Lee, H.-J. The Inverse Association of Sarcopenia and Protein-Source Food and Vegetable Intakes in the Korean Elderly: The Korean Frailty and Aging Cohort Study. Nutrients 2022, 14, 1375. [Google Scholar] [CrossRef]
- Magalhães, N.V.; Waitzberg, D.L.; Lopes, N.C.; Vicedomini, A.C.C.; Prudêncio, A.P.A.; Jacob-Filho, W.; Busse, A.L.; Ferdinando, D.; Alves, T.P.; Pereira, R.M.R.; et al. High Prevalence of Energy and Nutrients Inadequacy among Brazilian Older Adults. Nutrients 2023, 15, 3246. [Google Scholar] [CrossRef] [PubMed]
- Bartali, B.; Semba, R.D.; Frongillo, E.A.; Varadhan, R.; Ricks, M.O.; Blaum, C.S.; Ferrucci, L.; Guralnik, J.M.; Fried, L.P. Low Micronutrient Levels as a Predictor of Incident Disability in Older Women. Arch. Intern. Med. 2006, 166, 2335–2340. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Karagounis, L.G.; Ng, T.P.; Carre, C.; Narang, V.; Wong, G.; Tan, C.T.Y.; Nyunt, M.S.; Gao, Q.; Abel, B.; et al. Systemic and Metabolic Signature of Sarcopenia in Community-Dwelling Older Adults. J. Gerontol. A Biol. Sci. Med. Sci. 2020, 75, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, F.; Hashimoto, Y.; Kaji, A.; Sakai, R.; Kawate, Y.; Okamura, T.; Kondo, Y.; Fukuda, T.; Kitagawa, N.; Okada, H.; et al. Vitamin Intake and Loss of Muscle Mass in Older People with Type 2 Diabetes: A Prospective Study of the KAMOGAWA-DM Cohort. Nutrients 2021, 13, 2335. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.; Son, K.; Lim, H. Association between Skeletal Muscle Mass-to-Visceral Fat Ratio and Dietary and Cardiometabolic Health Risk Factors among Korean Women with Obesity. Nutrients 2023, 15, 1574. [Google Scholar] [CrossRef] [PubMed]
- Balboa-Castillo, T.; Struijk, E.A.; Lopez-Garcia, E.; Banegas, J.R.; Rodríguez-Artalejo, F.; Guallar-Castillon, P. Low vitamin intake is associated with risk of frailty in older adults. Age Ageing 2018, 47, 872–879. [Google Scholar] [CrossRef] [PubMed]
- Semba, R.D.; Bartali, B.; Zhou, J.; Blaum, C.; Ko, C.-W.; Fried, L.P. Low serum micronutrient concentrations predict frailty among older women living in the community. J. Gerontol. A Biol. Sci. Med. Sci. 2006, 61, 594–599. [Google Scholar] [CrossRef]
- Cheng, X.; Hu, Y.; Ruan, Z.; Zang, G.; Chen, X.; Qiu, Z. Association between B-vitamins intake and frailty among patients with chronic obstructive pulmonary disease. Aging Clin. Exp. Res. 2023, 35, 793–801. [Google Scholar] [CrossRef]
- Koole, J.L.; Bours, M.J.L.; Geijsen, A.J.M.R.; Gigic, B.; Ulvik, A.; Kok, D.E.; Brezina, S.; Ose, J.; Baierl, A.; Böhm, J.; et al. Circulating B-vitamin biomarkers and B-vitamin supplement use in relation to quality of life in patients with colorectal cancer: Results from the FOCUS consortium. Am. J. Clin. Nutr. 2021, 113, 1468–1481. [Google Scholar] [CrossRef]
- Cui, R.; Iso, H.; Date, C.; Kikuchi, S.; Tamakoshi, A. Dietary Folate and Vitamin B6 and B12 Intake in Relation to Mortality From Cardiovascular Diseases. Japan Collaborative Cohort Study. Stroke 2010, 41, 1285–1289. [Google Scholar]
- Zhao, L.-G.; Shu, X.-O.; Li, H.-L.; Gao, J.; Han, L.-H.; Wang, J.; Fang, J.; Gao, Y.-T.; Zheng, W.; Xiang, Y.-B. Prospective cohort studies of dietary vitamin B6 intake and risk of cause-specific mortality. Clin. Nutr. 2019, 38, 1180–1187. [Google Scholar] [CrossRef] [PubMed]
- Bo, Y.; Xu, H.; Zhang, H.; Zhang, J.; Wan, Z.; Zhao, X.; Yu, Z. Intakes of Folate, Vitamin B6, and Vitamin B12 in Relation to All-Cause and Cause-Specific Mortality: A National Population-Based Cohort. Nutrients 2022, 14, 2253. [Google Scholar] [CrossRef] [PubMed]
- Huang, V.-C.; Lee, M.-S.; Wahlqvist, M.L. Prediction of all-cause mortality by B group vitamin status in the elderly. Clin. Nutr. 2012, 31, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Liu, Y.; Wang, Y.; Ma, Y.; Bai, J.; Yu, C. Association of Serum Vitamin B6 with All-Cause and Cause-Specific Mortality in a Prospective Study. Nutrients 2021, 13, 2977. [Google Scholar] [CrossRef] [PubMed]
- Schorgg, P.; Karavasiloglou, N.; Beyer, A.; Cantwell, M.; Danquah, I.; Gojda, J.; Rohrmann, S.; Cassidy, A.; Barnighausen, T.; Cahova, M.; et al. Increased vitamin B6 turnover is associated with greater mortality risk in the general US population: A prospective biomarker study. Clin. Nutr. 2022, 41, 1343–1356. [Google Scholar] [CrossRef] [PubMed]
- Dugué, P.-A.; Hodge, A.M.; Ulvik, A.; Ueland, P.M.; Midttun, Ø.; Rinaldi, S.; MacInnis, R.J.; Sherly, X.; Li, S.X.; Meyer, K.; et al. Association of Markers of Inflammation, the Kynurenine Pathway and B Vitamins with Age and Mortality, and a Signature of Inflammaging. J. Gerontol. A Biol. Sci. Med. Sci. 2022, 77, 826–836. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Gammon, M.D.; Wetmur, J.G.; Bradshaw, P.T.; Teitelbaum, S.L.; Neugut, A.I.; Santella, R.M.; Jia Chen, J. B-Vitamin Intake, One-Carbon Metabolism, and Survival in a Population-Based Study of Women with Breast Cancer. Cancer Epidemiol. Biomarkers Prev. 2008, 17, 2109–2116. [Google Scholar] [CrossRef]
- Lochhead, P.; Nishihara, R.; Qian, Z.R.; Mima, K.; Cao, Y.; Sukawa, Y.; Kim, S.A.; Inamura, K.; Zhang, X.; Wu, K.; et al. Postdiagnostic intake of one-carbon nutrients and alcohol in relation to colorectal cancer survival. Am. J. Clin. Nutr. 2015, 102, 1134–1141. [Google Scholar] [CrossRef]
- Ricci, C.; Freisling, H.; Leitzmann, M.F.; Taljaard-Krugell, C.; Jacobs, I.; Kruger, H.S.; Smuts, C.M.; Pieters, M. Diet and sedentary behaviour in relation to cancer survival. A report from the national health and nutrition examination survey linked to the U.S. mortality registry. Clin. Nutr. 2020, 39, 3489–3496. [Google Scholar] [CrossRef]
- Xu, H.-L.; Gong, T.-T.; Liu, F.-H.; Wei, Y.-F.; Chen, H.-Y.; Yan, S.; Zhao, Y.-H.; Gao, S.; Jiao, Y.-S.; Wu, Q.-J. Pre-diagnosis Dietary One-Carbon Metabolism Micronutrients Consumption and Ovarian Cancer Survival: A Prospective Cohort Study. Front. Nutr. 2022, 9, 873249. [Google Scholar] [CrossRef]
- He, T.; Xiao, H.; Wusiman, M.; Yishake, D.; Luo, Y.; Liu, X.; Liu, Z.; Zhu, H. Dietary intake of one-carbon metabolism-related nutrients and hepatocellular carcinoma survival in the Guangdong Liver Cancer Cohort. Food Funct. 2022, 13, 8081–8090. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Peng, H.; Xia, C.; Zhou, Y.; Shen, L.; Cheng, X.; Yang, C.; Yang, Y.; Long, L. Association of B vitamins intake and total homocysteine levels With All-cause and Cause-Specific Mortality in Central obesity. Nutrition 2023, 116, 112189. [Google Scholar] [CrossRef] [PubMed]
- Je, Y.; Lee, J.E.; Ma, J.; Zhang, X.; Cho, E.; Rosner, B.; Selhub, J.; Fuchs, C.S.; Meyerhardt, J.; Giovannucci, E. Prediagnostic plasma vitamin B6 (pyridoxal 5′-phosphate) and survival in patients with colorectal cancer. Cancer Causes Control 2013, 24, 71929. [Google Scholar] [CrossRef] [PubMed]
- Muller, D.C.; Johansson, M.; Zaridze, D.; Moukeria, A.; Janout, V.; Holcatova, I.; Navratilova, M.; Mates, D.; Midttun, Ø.; Ueland, P.M.; et al. Circulating Concentrations of Vitamin B6 and Kidney Cancer Prognosis: A Prospective Case-Cohort Study. PLoS ONE 2015, 10, e0140677. [Google Scholar] [CrossRef] [PubMed]
- Ulvik, A.; Pedersen, E.R.; Svingen, G.F.; McCann, A.; Midttun, Ø.; Nygård, O.; Ueland, P.M. Vitamin B-6 catabolism and long-term mortality risk in patients with coronary artery disease. Am. J. Clin. Nutr. 2016, 103, 1417–1425. [Google Scholar] [CrossRef] [PubMed]
- Minovic, I.; van der Veen, A.; van Faassen, M.; Riphagen, I.J.; van den Berg, E.; van der Ley, C.; Gomes-Neto, A.W.; Geleijnse, J.M.; Eggersdorfer, M.; Navis, G.J.; et al. Functional vitamin B-6 status and long-term mortality in renal transplant recipients. Am. J. Clin. Nutr. 2017, 106, 1366–1374. [Google Scholar] [CrossRef] [PubMed]
- Pusceddu, I.; Herrmann, W.; Kleber, M.E.; Scharnagl, H.; Hoffmann, M.M.; Winklhofer-Roob, B.M.; März, W.; Herrmann, M. Subclinical inflammation, telomere shortening, homocysteine, vitamin B6, and mortality: The Ludwigshafen Risk and Cardiovascular Health Study. Eur. J. Nutr. 2020, 59, 1399–1411. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Li, Y.; Lang, X.; Zhang, Y. Associations of Serum Vitamin B6 Status and Catabolism With All-Cause Mortality in Patients With T2DM. J. Clin. Endocrinol. Metab. 2022, 107, 2822–2832. [Google Scholar] [CrossRef]
- Holowatyj, A.N.; Ose, J.; Gigic, B.; Lin, T.; Ulvik, A.; Geijsen, A.J.; Brezina, S.; Kiblawi, R.; van Roekel, E.H.; Baierl, A.; et al. Higher vitamin B6 status is associated with improved survival among patients with stage I–III colorectal cancer. Am. J. Clin. Nutr. 2022, 116, 303–313. [Google Scholar] [CrossRef]
- Cui, Q.; Zhu, X.; Guan, G.; Hui, R.; Zhu, L.; Wang, J.; Zhao, J. Associations of vitamin B6 turnover rate with the risk of cardiovascular and all-cause mortality in hypertensive adults. Nutr. Metab. Cardiovasc. Dis. 2023, 33, 1225–1234. [Google Scholar] [CrossRef]
- Ebbing, M.; Bønaa, K.H.; Nygard, O.; Arnesen, E.; Ueland, P.M.; Nordrehaug, J.E.; Rasmussen, K.; Njølstad, I.; Helga Refsum, H.; Nilsen, D.W.; et al. Cancer Incidence and Mortality After Treatment With Folic Acid and Vitamin B12. JAMA 2009, 302, 2119–2126. [Google Scholar] [CrossRef] [PubMed]
- Albert, C.M.; Cook, N.R.; Gaziano, J.M.; Zaharris, E.; MacFadyen, J.; Danielson, E.; Buring, J.E.; HoAnn, E.; Manson, H.E. Effect of Folic Acid and B Vitamins on Risk of Cardiovascular Events and Total Mortality Among Women at High Risk for Cardiovascular Disease. A Randomized Trial. JAMA 2008, 299, 2027–2036. [Google Scholar] [CrossRef] [PubMed]
- Kataria, N.; Yadav, P.; Kumar, R.; Kumar, N.; Singh, M.; Kant, R.; Kalyani, V. Effect of Vitamin B6, B9, and B12 Supplementation on Homocysteine Level and Cardiovascular Outcomes in Stroke Patients: A Meta-Analysis of Randomized Controlled Trials. Cureus 2021, 13, e14958. [Google Scholar] [CrossRef] [PubMed]
- Brin, M.; Thiele, V.F. Relationships between Vitamin B6-vitamer Content and the Activities of Two Transaminase Enzymes in Rat Tissues at Varying Intake Levels of Vitamin B6. J. Nutr. 1967, 93, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Vespasiani-Gentilucci, U.; De Vincentis, A.; Ferrucci, L.; Bandinelli, S.; Incalzi, R.A.; Picardi, A. Low alanine aminotransferase levels in the elderly population: Frailty, disability, sarcopenia, and reduced survival. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2018, 73, 925–930. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Que, S.; Xu, J.; Peng, T. Alanine aminotransferase-old biomarker and new concept: A review. Int. J. Med. Sci. 2014, 11, 925–935. [Google Scholar] [CrossRef] [PubMed]
- Ndrepepa, G.; Kastrati, A. Alanine aminotransferase—A marker of cardiovascular risk at high and low activity levels. J. Lab. Precis. Med. 2019, 4, 29. [Google Scholar] [CrossRef]
- Izumi, R.; Suzuki, N.; Kato, K.; Warita, H.; Tateyama, M.; Nakashima, I.; Itoyama, Y. A Case of McArdle Disease: Efficacy of Vitamin B6 on Fatigability and Impaired Glycogenolysis. Intern. Med. 2010, 49, 1623–1625. [Google Scholar] [CrossRef]
- Migocka-Patrzałek, M.; Elias, M. Muscle Glycogen Phosphorylase and Its Functional Partners in Health and Disease. Cells 2021, 10, 883. [Google Scholar] [CrossRef]
- Sampson, D.A.; Young, L.A.; Kretsch, M.J. Marginal intake of vitamin B-6: Effect of protein synthesis in liver, kidney and muscle of rat. Nutr. Res. 1988, 8, 309–319. [Google Scholar] [CrossRef]
- Suidasari, S.; Uragami, S.; Yanaka, N.; Kato, N. Dietary vitamin B6 modulates the gene expression of myokines, Nrf2-related factors, myogenin and HSP60 in the skeletal muscle of rats. Exp. Ther. Med. 2017, 14, 3239–3246. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kosek, D.J.; Petrella, J.K.; Cross, J.M.; Bamman, M.M. Resting and load-induced levels of myogenic gene transcripts differ between older adults with demonstrable sarcopenia and young men and women. J. Appl. Physiol. 2005, 99, 2149–2158. [Google Scholar] [CrossRef] [PubMed]
- Rathor, R.; Suryakumar, G.; Singh, S.N.; Kumar, B. Heat Shock Protein 60 (HSP60): Role in Skeletal Muscle Diseases and Novel Prospects for Therapy. In Heat Shock Protein 60 in Human Diseases and Disorders Heat Shock Proteins; Asea, A., Kaur, P., Eds.; Springer: Cham, Switzerland, 2019; Volume 18. [Google Scholar] [CrossRef]
- Huang, D.-D.; Fan, S.-D.; Chen, X.-Y.; Yan, X.-L.; Zhang, X.-Z.; Ma, B.-W.; Yu, D.-Y.; Xiao, W.-Y.; Zhuang, C.-L.; Yu, Z. Nrf2 deficiency exacerbates frailty and sarcopenia by impairing skeletal muscle mitochondrial biogenesis and dynamics in an age-dependent manner. Exp. Gerontol. 2019, 1119, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.Y.; Wu, Z.Y.; Zhu, Y.L.; Liu, L.; Li, F.C. Effects of dietary vitamin B6 on the skeletal muscle protein metabolism of growing rabbits. Anim. Prod. Sci. 2016, 57, 2007–2015. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, X.; Ma, Y.; Yang, Y.; Ge, S. The effects of vitamin B6 on the nutritional support of BCAAs-enriched amino acids formula in rats with partial gastrectomy. Clin. Nutr. 2023, 42, 954–961. [Google Scholar] [CrossRef] [PubMed]
- Suidasari, S.; Stautemas, J.; Uragami, S.; Yanaka, N.; Derave, W.; Kato, N. Carnosine Content in Skeletal Muscle Is Dependent on Vitamin B6 Status in Rats. Front. Nutr. 2016, 2, 39. [Google Scholar] [CrossRef]
- Kumrungsee, T.; Nirmagustina, D.E.; Arima, T.; Onishi, K.; Sato, K.; Kato, N.; Yanaka, N. Novel metabolic disturbances in marginal vitamin B6-deficient rat heart. J. Nutr. Biochem. 2019, 65, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Artioli, G.G.; Sale, C.; Jones, R.L. Carnosine in health and disease. Eur. J. Sport. Sci. 2019, 19, 30–39. [Google Scholar] [CrossRef]
- Chaleckis, R.; Murakami, I.; Takada, J.; Kondoh, H.; Yanagida, M. Individual variability in human blood metabolites identifies age-related differences. Proc. Natl. Acad. Sci. USA 2016, 113, 4252–4259. [Google Scholar] [CrossRef]
- Michele, D.E. Mechanisms of skeletal muscle repair and regeneration in health and disease. FEBS J. 2022, 289, 6460–6462. [Google Scholar] [CrossRef]
- Komaru, T.; Yanaka, N.; Kumrungsee, T. Satellite Cells Exhibit Decreased Numbers and Impaired Functions on Single Myofibers Isolated from Vitamin B6-Deficient Mice. Nutrients 2021, 13, 4531. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, Y.; Sevak, J.K.; Kumar, S.; Kumar, N.; Gopinath, S.D. Metabolomic analysis of primary human skeletal muscle cells during myogenic progression. Sci. Rep. 2020, 10, 11824. [Google Scholar] [CrossRef]
- Palin, M.-F.; Lapointe, J.; Gariepy, C.; Beaudry, D.; Kalbe, C. Characterisation of intracellular molecular mechanisms modulated by carnosine in porcine myoblasts under basal and oxidative stress conditions. PLoS ONE 2020, 15, e0239496. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Shen, W.; Liu, T.; Mosenthin, R.; Bao, Y.; Chen, P.; Hao, W.; Zhao, L.; Zhang, J.; Ji, C.; et al. Improved Satellite Cell Proliferation Induced by L-Carnosine Benefits Muscle Growth of Pigs in Part through Activation of the Akt/mTOR/S6K Signaling Pathway. Agriculture 2022, 12, 988. [Google Scholar] [CrossRef]
- Nagai, A.; Ida, M.; Izumo, T.; Nakai, M.; Honda, H.; Shimizu, K. L-Anserine Increases Muscle Differentiation and Muscle Contractility in Human Skeletal Muscle Cells. J. Agric. Food Chem. 2023, 71, 8952–8958. [Google Scholar] [CrossRef] [PubMed]
- Onódi, Z.; Szabó, P.L.; Kucsera, D.; Pokreisz, P.; Dostal, C.; Hilber, K.; Oudit, G.Y.; Podesser, B.K.; Ferdinandy, P.; Varga, Z.V.; et al. Inflammasome Activity in the Skeletal Muscle and Heart of Rodent Models for Duchenne Muscular Dystrophy. Int. J. Mol. Sci. 2023, 24, 8497. [Google Scholar] [CrossRef] [PubMed]
- Górecki, D.C. P2X7 purinoceptor as a therapeutic target in muscular dystrophies. Curr. Opin. Pharmacol. 2019, 47, 40–45. [Google Scholar] [CrossRef]
- Mourkioti, F.; Rosenthal, N. NF-κB signaling in skeletal muscle: Prospects for intervention in muscle diseases. J. Mol. Med. 2008, 86, 747–759. [Google Scholar] [CrossRef]
- Yanaka, N.; Koyama, T.; Komatsu, S.; Nakamura, E.; Kanda, M.; Kato, N. Vitamin B6 suppresses NF-κB activation in LPS-stimulated mouse macrophages. Int. J. Mol. Med. 2005, 16, 1071–1075. [Google Scholar] [CrossRef]
- Zhang, P.; Tsuchiya, K.; Kinoshita, T.; Kushiyama, H.; Suidasari, S.; Hatakeyama, M.; Imura, H.; Kato, N.; Suda, T. Vitamin B6 prevents IL-1 beta protein production by inhibiting NLRP3 inflammasome activation. J. Biol. Chem. 2016, 291, 24517–24527. [Google Scholar] [CrossRef]
- Castelblanco, M.; Lugrin, J.; Ehirchiou, D.; Nasi, S.; Ishii, I.; So, A.; Martinon, F.; Busso, N. Hydrogen sulfide inhibits NLRP3 inflammasome activation and reduces cytokine production both in vitro and in a mouse model of inflammation. J. Biol. Chem. 2018, 293, 2546–2557. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros, J.; Rivas, D.; Duque, G. The Role of the Kynurenine Pathway in the Pathophysiology of Frailty, Sarcopenia, and Osteoporosis. Nutrients 2023, 15, 3132. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Xie, R.; He, H.; Xie, Q.; Zhao, X.; Kang, G.; Cheng, C.; Yin, W.; Cong, J.; Li, J.; et al. Kynurenic acid ameliorates NLRP3 inflammasome activation by blocking calcium mobilization via GPR35. Front. Immunol. 2022, 13, 1019365. [Google Scholar] [CrossRef] [PubMed]
- Aryana, I.G.P.S.; Hapsari, A.A.A.R.; Kuswardhani, R.A.T. Myokine Regulation as Marker of Sarcopenia in Elderly. Mol. Cell Biomed. Sci. 2018, 2, 38–47. [Google Scholar] [CrossRef]
- Cordero, M.D.; Williams, M.R.; Ryffel, B. AMP-Activated Protein Kinase Regulation of the NLRP3 Inflammasome during Aging. Trends Endocrinol. Metab. 2018, 29, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Shan, M.-R.; Zhou, S.-N.; Fu, C.-N.; Song, J.-W.; Wang, X.-Q.; Bai, W.-W.; Li, P.; Song, P.; Zhu, M.-L.; Ma, Z.-M.; et al. Vitamin B6 inhibits macrophage activation to prevent lipopolysaccharide-induced acute pneumonia in mice. J. Cell Mol. Med. 2020, 24, 3139–3148. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.-Y.; Chen, J.-L.; Xiao, M.-H.; Sun, Y.; Zhao, Y.-X.; Pu, D.; Lv, A.-K.; Wang, M.-L.; Zhou, J.; Zhu, S.-Y.; et al. The effect of exercise, resveratrol or their combination on Sarcopenia in aged rats via regulation of AMPK/Sirt1 pathway. Exp. Gerontol. 2017, 98, 177–183. [Google Scholar] [CrossRef]
- Zeng, Z.; Liang, J.; Wu, L.; Zhang, H.; Lv, J.; Chen, N. Exercise-Induced Autophagy Suppresses Sarcopenia Through Akt/mTOR and Akt/FoxO3a Signal Pathways and AMPK-Mediated Mitochondrial Quality Control. Front. Physiol. 2020, 11, 583478. [Google Scholar] [CrossRef]
- Yuan, J.; Li, J.; Shang, M.; Fu, Y.; Wang, T. Identification of vitamin B6 as a PD-L1 suppressor and an adjuvant for cancer immunotherapy. Biochem. Biophys. Res. Commun. 2021, 561, 187–194. [Google Scholar] [CrossRef]
- Kiriyama, Y.; Nochi, H. Regulation of PD-L1 Expression by Nuclear Receptors. Int. J. Mol. Sci. 2023, 24, 9891. [Google Scholar] [CrossRef]
- Onorati, A.; Havas, A.P.; Lin, B.; Rajagopal, J.; Sen, P.; Peter, D.; Adams, P.D.; Dou, Z. Upregulation of PD-L1 in Senescence and Aging. Mol. Cell Biol. 2022, 42, e0017122. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.W.; Johmura, Y.; Suzuki, N.; Omori, S.; Migita, T.; Yamaguchi, K.; Hatakeyama, S.; Yamazaki, S.; Shimizu, E.; Imoto, S.; et al. Blocking PD-L1–PD-1 improves senescence surveillance and ageing phenotypes. Nature 2022, 611, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Weng, X.; Xiao, W.; Xu, X.; Chen, Y.; Chen, P. Pharmacological and Genetic Inhibition of PD-1 Demonstrate an Important Role of PD-1 in Ischemia-Induced Skeletal Muscle Inflammation, Oxidative Stress, and Angiogenesis. Front. Immunol. 2021, 12, 586429. [Google Scholar] [CrossRef] [PubMed]
- Khalid, N.M.; Haron, H.; Shahar, S.; Fenech, M. Current Evidence on the Association of Micronutrient Malnutrition with Mild Cognitive Impairment, Frailty, and Cognitive Frailty among Older Adults: A Scoping Review. Int. J. Environ. Res. Public. Health 2022, 19, 15722. [Google Scholar] [CrossRef] [PubMed]
- Welan, R. Effect of Vitamin B6 on Osteoporosis Fracture. J. Bone Metab. 2023, 30, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Sato, R.; Vatic, M.; da Fonseca, G.W.W.; von Haehling, S. Sarcopenia and Frailty in Heart Failure: Is There a Biomarker Signature? Curr. Heart Fail. Rep. 2022, 19, 400–411. [Google Scholar] [CrossRef]
- Baumgartner, R.N. Body composition in healthy aging. Ann. N. Y. Acad. Sci. 2000, 904, 437–448. [Google Scholar] [CrossRef]
- Eitmann, S.; Matrai, P.; Hegyi, P.; Balasko, M.; Eross, B.; Dorogia, K.; Petervari, E. Obesity paradox in older sarcopenic adults—A delay in aging: A systematic review and meta-analysis. Ageing Res. Rev. 2024, 93, 102164. [Google Scholar] [CrossRef]
- Aasheim, E.T.; Hofsø, D.; Hjelmesæth, J.; Birkeland, K.I.; Bøhmer, T. Vitamin status in morbidly obese patients: A cross-sectional study. Am. J. Clin. Nutr. 2008, 87, 362–369. [Google Scholar] [CrossRef]
- Fu, Y.; Zhu, Z.; Huang, Z.; He, R.; Zhang, Y.; Li, Y.; Tan, W.; Rong, S. Association between Vitamin B and Obesity in Middle-Aged and Older Chinese Adults. Nutrients 2023, 15, 483. [Google Scholar] [CrossRef]


{kind=link}
| References, Publication Year (Recruiting Area) | Assessment of B6 Status | Study Design | Study Population | Key Findings |
|---|---|---|---|---|
| General population | ||||
| Struijk et al. [10] 2018 (Spain) | B6 intake | Cohort (median follow-up of 3.5 years) | n = 1630 participants (≥60 years, male 51–53%) | (+) The OR for highest vs. lowest tertile of B6 intake was 0.66 (95% CI, 0.44–0.99; p-trend < 0.05). No association was found between B12 and folate intakes and physical function. |
| Grootswagers et al. 2021 [11] (Italy, UK, Netherland, Poland, France) | B6 intake | Cross-sectional | 1249 healthy older adults (65–79 years, male, 36–50%) | (+) Positive associations between B6 intake and chair rise test performance in the full population (p < 0.05) and between B6 intake and handgrip strength in participants with low physical activity were observed (p < 0.05). Homocysteine concentrations did not mediate these associations. |
| Park et al. [12] 2022 (Korea) | B6 intake | Cohort (up to 10 years) | 801 participants (70–84 years, 48% male) | (+) The highest quartile of B6 intake was 0.45-fold lower for the risk of sarcopenia compared to the lowest quartile (HR = 0.45; 95% CI: 0.22–0.91; p-trend = 0.025). Intakes of retinol, D, K, B1, B2, niacin, C, B12, and folate were not associated with sarcopenia. |
| Magalhães et al. [13] 2023 (Brasil) | B6 intake | Cross-sectional | 295 community-living older adults (>60 years, mean age 70 years, male 18%) | (+) Seventy eight % of the participants had insufficient intake of B6. Inadequancy of B6 (OR = 2.18; 95% CI: 1.03–4.64) was identified as risk factor for sarcopenia (p < 0.05). |
| Bartali et al. [14] 2006 (USA) | Serum PLP | Cohort (up to 3 years) | 643 women (≥65 years) | (+) Women in the lowest quartile of serum concentrations of PLP (HR = 1.31; 95% CI: 1.03–1.67), B12 (1.40; 1.12–1.74), and Se (1.38; 1.12–1.71) had higher risk of disability compared with women in the upper 3 quartiles (p < 0.05). Serum levels of retinol, 25(OH)D, folate, and Zn were not associated with disability. |
| Lu et al. [15] 2020 (Singapore) | Serum HK/XA ratio | Cross-sectional | 189 participants (mean age 73.2 years, male 37%) | (+) Compared with non-sarcopenic adults, sarcopenic adults had higher HK/XA ratio (OR = 1.198; 95% CI: 1.010–1.420; p = 0.038), indicating a deficiency of B6. Serum levels of retinol, 25(OH)D3, α-tocopherol, and total B12 did not differ. |
| Patients | ||||
| Takahashi et al. [16] 2021 (Japan) | B6 intake | Cohort (mean 13.7-month follow-up) | 197 patients with T2DM (mean age 72 years, 57% male) | (−) Intake of D was significantly related to the loss of muscle mass (OR = 0.93; 95% CI: 0.88–0.97; p = 0.003). However, intakes of A, B2, B6, B12, C, and E were not related. |
| Lim et al. [17] 2023 (Korea) | B6 intake | Cross-sectional | 59 women (19–65 years, BMI values of 25–32 kg/m2) | (+) Intakes of B6 and B12 intake (B6: β = 0.338, p = 0.012; B12: β = 0.281, p = 0.024) showed positive associations with the skeletal muscle mass-to-visceral fat area ratio. Intakes of other vitamins such as A, D, E, K, B1, B2, niacin, panthothenic acid, C, and folate showed no association. |
| References, Publication Year (Recruiting Area) | Assessment of B6 Status | Study Design | Study Population | Key Findings |
|---|---|---|---|---|
| General population | ||||
| Balboa-Castillo et al. [18] 2018 (Spain) | B6 intake | Cohort (up to 3.5 years) | 1643 community-dwelling individuals (≥65 years, 50% male) | (+) The ORs (95% Cl) of frailty for those in the lowest versus the highest tertile of vitamin intake were 2.80 (1.38–5.67), p-trend 0.004, for B6; 1.65 (0.93–2.95), p-trend 0.007, for C; 1.93 (0.99–3.83), p-trend 0.06, for E; and 2.34 (1.21–4.52), p-trend 0.01, for folates. Non-adherence to the RDAs of B6 was related to frailty (OR 2.23; 95% CI: 1.30–3.83; p < 0.01). |
| Semba et al. [19] 2006 (USA) | Serum PLP | Cohort (up to 3 years) | 766 female participants (≥65 years) | (−) Compared with women in the upper three quartiles, women in the lowest quartile of serum carotenoids (HR = 1.39; 95% CI: 1.01–1.92) and α-tocopherol (HR = 1.39; 95% CI: 1.02–1.92) had an increased risk of becoming frail (p = 0.04). Other nutrients such as retinol, PLP, B12, and folate were not significantly associated with the incidence of frailty. |
| Patients | ||||
| Cheng et al. [20] 2023 (USA) | B6 intake | Population-based cross-sectional | 1201 COPD patients (mean age 63.9 years, 62% male) | (+) The association between B6 intake and frailty risk (OR = 0.80; 95% CI: 0.66–0.95; p = 0.013) was significant. Intakes of B1, B2, niacin, folate, and B12 were not associated with frailty risk. |
| Koole et al. [21] 2021 (Netherland, Germany) | Serum PLP | Cohort (up to 6 months) | 1676 diagnosed stage I–III colorectal cancer patients (mean age 65.6 years, 64% male) | (+) B6 status was found to be associated with better physical functioning, better social functioning, and less fatigue (p < 0.05). Dose–response relationships were found between B6 status and quality of life outcomes. Plasma levels of B2, folate, cobalamin, and homocysteine were not related to frailty. |
| References, Publication Year (Recruiting Area) | Assessment of B6 Status | Study Design | Study Population | Key Findings |
|---|---|---|---|---|
| General population | ||||
| Cui et al. [22] 2010 (Japan) | B6 intake | Cohort (median 14-year follow up) | 58,730 participants (40–79 years, 39% male) | The HRs (95% CI) of the mortality from heart failure for the highest versus lowest quintiles were 0.39 (0.15–1.00) for B6 and 0.50 (0.27–0.94) for folate intakes in men (p-trend < 0.05), and the HR of coronary heart disease (CHD) was 0.57 (0.34–0.96) for folate intakes in women (p-trend < 0.05). No association was found between B12 intake and mortality from CHD. |
| Zhao et al. [23] 2019 (China) | B6 intake | Cohort (median 10.3 year follow-up for men and median 16.2-year follow-up for women) | 134,480 participants (40–74 years, 44% male) | (+) The HR for the highest vs. lowest quintiles of B6 for total, cardiovascular disease (CVD), stroke, and CHD mortality among men was 0.66–0.83 (p-trend < 0.01). Similar association was found in women. No significant association was observed between dietary B6 and cancer mortality among both men and women. |
| Bo et al. [24] 2022 (USA) | B6 intake | Two cohort (median 9.8-year follow-up) | 55,569 participants (mean age 49 years, 52% male) | (+) In men, the HRs (95% CI) for the highest versus lowest quintiles of folate and B6 were 0.77 (0.71–0.85) and 0.79 (0.71–0.86) for all-cause mortality, 0.59 (0.48–0.72) and 0.69 (0.56–0.85) for CVD mortality, and 0.68 (0.56–0.84) and 0.73 (0.60–0.90) for cancer mortality, respectively (p trend < 0.05). Among women, the HRs (95% CI) for the highest versus lowest quintiles of folate and B6 were 0.86 (0.78–0.95) and 0.88 (0.80–0.97) for all-cause mortality and 0.53 (0.41–0.69) and 0.56 (0.44–0.73) for CVD mortality, respectively (p trend < 0.05). No significant associations between dietary B12 and all-cause and cause-specific mortality were observed. |
| Huang et al. [25] 2012 (Taiwan) | Plasma PLP | Cohort (up to 8 years) | 1747 participants (≥65 years, 50% male) | (+) Relative to the lowest tertile of B1 or B6 intakes, the HRs (95% CI) for 3rd tertile were 0.74 (0.58–0.95) and 0.74 (0.57–0.97) (both p-trend < 0.05) for all-cause mortality, respectively. No significant associations were observed between niacin and B2 and all-cause mortality. |
| Yang et al. [26] 2021 (USA) | Serum PLP | Cohort (up to 5 years) | 12,190 participants (mean age 46.6 years, 49% male) | (+) The participants with higher serum PLP had lower risk of all-cause mortality (HR = 0.85) and lower risk of CVD mortality (HR = 0.81) for each unit increment in natural log-transformed PLP, respectively. A higher log-transformed PLP was not significantly associated with a lower risk for cancer-specific mortality. Compared with sufficient B6, deficient (HR = 1.37) and insufficient (HR = 1.19) B6 levels were significantly associated with a higher risk for all-cause mortality. |
| Schorgg et al. [27] 2022 (USA) | Serum PA/PLP ratio | Cohort (median 7.8-year follow-up) | 15,304 participants (20–85 years, 48% male) | (+) A positive association between the PA/PLP ratio and all-cause mortality was observed with the HR (5th vs. 1st quitile) of 1.45 (95% CI: 1.14–1.85; p-trend < 0.0001). There were no significant associations of B6 turnover with CVD or cancer-specific mortality. |
| Dugué et al. [28] 2022 (Australia) | Serum HK/XA ratio and PAr index | Cohort (median 14.3-year follow-up | 41,513 participants (27–88 years, 48% male) | (+) Associations with all-cause mortality were observed for several markers of B6-related factors, including IL-6, neopterin, quinolinic acid, kynurenine/tryptophan ratio, cystatin C, HK/XA, calprotectin, PAr index, serum amyloid A, TNF-α, anthranilic acid, 3-hydroxykynurenine, 3-hydroxyanthranilic acid, and 4-pyridoxic acid (p < 0.05). |
| Patients | ||||
| Xu et al. [29] 2008 (USA) | B6 intake | Cohort (mean 5.6-year follow-up) | 1508 women with breast cancer (mean age 59 years) | (−) Higher intakes of B1 were associated with reduced all-cause mortality (HR = 0.44; 95% CI: 0.24–0.81; p-trend = 0.01). However, intakes of B6, folate, B2, niacin, and B12 were not associated with all-cause mortality. |
| Lochhead et al. [30] 2015 (USA) | B6 intake | Cohort (median 14.9-year follow-up) | 1550 patients with stage I–III colorectal cancer (mean age 66 years, 31% male) | (−) No significant associations were observed between postdiagnostic intakes of folate, B6, B12, and methionine and colorectal cancer-specific and all-cause mortality (p-trend ≥ 0.13). |
| Ricci et al. [31] 2020 (South Africa, Germany, France) | B6 intake | Cohort (median 5.7-year follow-up) | 2371 cancer survivors (≥19 years, 42.4% male) | (+) The highest versus the lowest tertiles of intakes of B6, B1, folate, B12, niacin, and C were inversely associated with all-cause mortality (HR = 0.55–0.75; p-trend < 0.05). Intakes of B6, B1, B12, niacin, and C were inversely associated with cancer-specific mortality (HR = 0.39–0.63; p-trend < 0.05). |
| Xu et al. [32] 2022 (China) | B6 intake | Cohort (median 37.2-month follow-up) | 635 newly diagnosed ovarian cancer patients (18–79 years) | A reduced ovarian cancer-specific mortality with the highest compared with the lowest tertile of dietary B6 (HR = 0.52, 95% CI: 0.32–0.84, p-trend < 0.05) was found. No significant associations with ovarian cancer mortality were observed for intakes of B2, niacin, folate, and B12. A curvilinear association between B6 intake and ovarian cancer mortality was found (p for non-linearity < 0.05). |
| He et al. [33] 2022 (China) | B6 intake | Cohort (median 791-day follow-up) | 905 diagnosed hepatocellular carcinoma patients (mean 51.9 years) | (−) There was no significant association of intakes of B6, folate, B12, B2, or niacin with all-cause mortality and with hepatocellular carcinoma-specific mortality (p for non-linearity > 0.05). |
| Wang et al. [34] 2023 (USA) | B6 intake | Cohort (median 15.7-year follow-up) | 7718 obesity adults (mean age 50 years, 20–85 years, 37% male) | (+) Folate intake was independently associated with a decreased incidence of all-cause mortality (HR = 0.71; 95% CI: 0.58–0.87; p-trend < 0.01). Higher intakes of B6 and B12 were inversely correlated with CVD mortality (0.63, 0.40–0.98 and 0.44, 0.29–0.65, respectively, all p-trends < 0.001), but not associated with cancer mortality. B6 intake was not associated with homocysteine levels. |
| Je et al. [35] 2013 (Korea) | Plasma PLP | Cohort (up to 7 years) | 472 patients with colorectal cancer (40–84 years, 68% male) | (−) Compared with patients who had less than 45 nmol/L of plasma PLP, those who had 110 nmol/L had HRs of 0.85 (95% CI: 0.50–1.45) and 0.87 (0.56–1.35) for colorectal cancer-specific and overall mortality (p-trend > 0.2). |
| Muller et al. [36] 2015 (Czech) | Plasma PLP | Case cohort (median 2.6-year follow-up) | 630 renal cell carcinoma patients (62% male, ≥18 years) | (+) The HR of the cancer-speficic death mortality for renal cell carcinoma patients was significantly lower among those in the highest compared to the lowest fourth of PLP concentration (HR (4th vs. 1st quartile) = 0.33; 95% CI: 0.18–0.60; p-trend < 0.001). |
| Ulvik et al. [37] 2016 (USA) | Plasma PLP, Par | Cohort (up to 8.8 years) | 3749 patients with acute myocardial infarction (mean age 63 years, 73% male) | (+) PAr, a proposed marker of B6 catabolism, predicted all-cause mortality. PAr provided an HR per SD of 1.31 (95% CI: 1.21–1.41) in patients hospitalized for acute myocardial infarction in a significant manner. |
| Minovic et al. [38] 2017 (Netherlands) | Plasma PLP, HK:XA ratio (a marker of functional B6 status) | Cohort (median 5.3-year follow-up) | 678 renal transplant recipients (RTRs) and 297 health control (mean age 54 years for control, and 53 years for RTRs, 55% male) | (+) RTRs had a higher median HK/XA than healthy controls (p < 0.05). In RTRs, the HK/XA was inversely associated with plasma PLP (p < 0.001). A higher HK/XA was significantly associated with increased risk of all-cause mortality, cancer mortality, and infectious disease mortality in RTRs (p < 0.05), but not with CV mortality (p > 0.05). |
| Pusceddu et al. [39] 2020 (Germany) | Plasma PLP | Cohort (median 9.9-year follow-up) | 2968 patients with coronary artery disease (48–76 years, 70% male) | (+) Patients in the 4th quartile of PLP had an HR for all-cause mortality of 0.41 (95% CI: 0.33–0.49) and for CVD mortality of 0.40 (0.33–0.49), compared to those in the 1st quartile. All-cause and CVD mortality significantly increased with higher concentrations of homocysteine and lower PLP. |
| Zhang et al. [40] 2022 (China) | Serum PLP | Cohort (median 85-month follow-up) | 2574 patients with T2DM (mean age 62.3 years, 51% male) | (+) The HRs and 95% CIs from lowest to highest serum levels of PLP (<21.4, 21.4–35.8, 35.7–63.6, and >63.6 nmol/L) were 1.00 (reference), 0.93 (0.71–1.23), 0.85 (0.64–1.13), and 0.74 (0.55–0.99), respectively, for all-cause mortality (p-trend = 0.035). |
| Holowatyj et al. [41] 2022 (Germany, USA, Netherland, Austria) | Serum PLP | Cohort (median 3.2-year follow-up) | 2031 patients diagnosed with stage I–III colorectal cancer (>18 years, 64% male) | (+) After a median follow-up of 3.2 years for overall survival, higher preoperative B6 status was associated with 16–32% higher all-cause and disease-free survival, although there was no significant association with disease recurrence. |
| Cui et al. [42] 2023 (China) | Serum PLP | Cohort (mean 11-year follow-up) | 5434 participants with hypertension (mean age 58.5 years, 50% male) | (+) PLP was negatively associated with CVD mortality (HR [95% CI] 4th vs. 1st quartile: 0.66; 0.47–0.94; p-trend = 0.03). Similarly, a higher quartile of PLP was associated with a lower risk of all-cause mortality (0.67; 0.56–0.80; p-trend < 0.01). |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kato, N.; Kimoto, A.; Zhang, P.; Bumrungkit, C.; Karunaratne, S.; Yanaka, N.; Kumrungsee, T. Relationship of Low Vitamin B6 Status with Sarcopenia, Frailty, and Mortality: A Narrative Review. Nutrients 2024, 16, 177. https://doi.org/10.3390/nu16010177
Kato N, Kimoto A, Zhang P, Bumrungkit C, Karunaratne S, Yanaka N, Kumrungsee T. Relationship of Low Vitamin B6 Status with Sarcopenia, Frailty, and Mortality: A Narrative Review. Nutrients. 2024; 16(1):177. https://doi.org/10.3390/nu16010177
Chicago/Turabian StyleKato, Norihisa, Akiko Kimoto, Peipei Zhang, Chanikan Bumrungkit, Sajith Karunaratne, Noriyuki Yanaka, and Thanutchaporn Kumrungsee. 2024. "Relationship of Low Vitamin B6 Status with Sarcopenia, Frailty, and Mortality: A Narrative Review" Nutrients 16, no. 1: 177. https://doi.org/10.3390/nu16010177
APA StyleKato, N., Kimoto, A., Zhang, P., Bumrungkit, C., Karunaratne, S., Yanaka, N., & Kumrungsee, T. (2024). Relationship of Low Vitamin B6 Status with Sarcopenia, Frailty, and Mortality: A Narrative Review. Nutrients, 16(1), 177. https://doi.org/10.3390/nu16010177