
A Combination of a Dopamine Receptor 2 Agonist and a Kappa Opioid Receptor Antagonist Synergistically Reduces Weight in Diet-Induced Obese Rodents


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Model and Diets
2.2. Treatments and Surgeries
2.2.1. Intracerebroventricular Treatment
2.2.2. Intraperitoneal Treatment
2.2.3. Weight Measurements
2.2.4. Nuclear Magnetic Resonance
2.2.5. Temperature Measurements and Thermal Imaging
2.2.6. Indirect Calorimetry
2.3. Statistical Analysis
3. Results
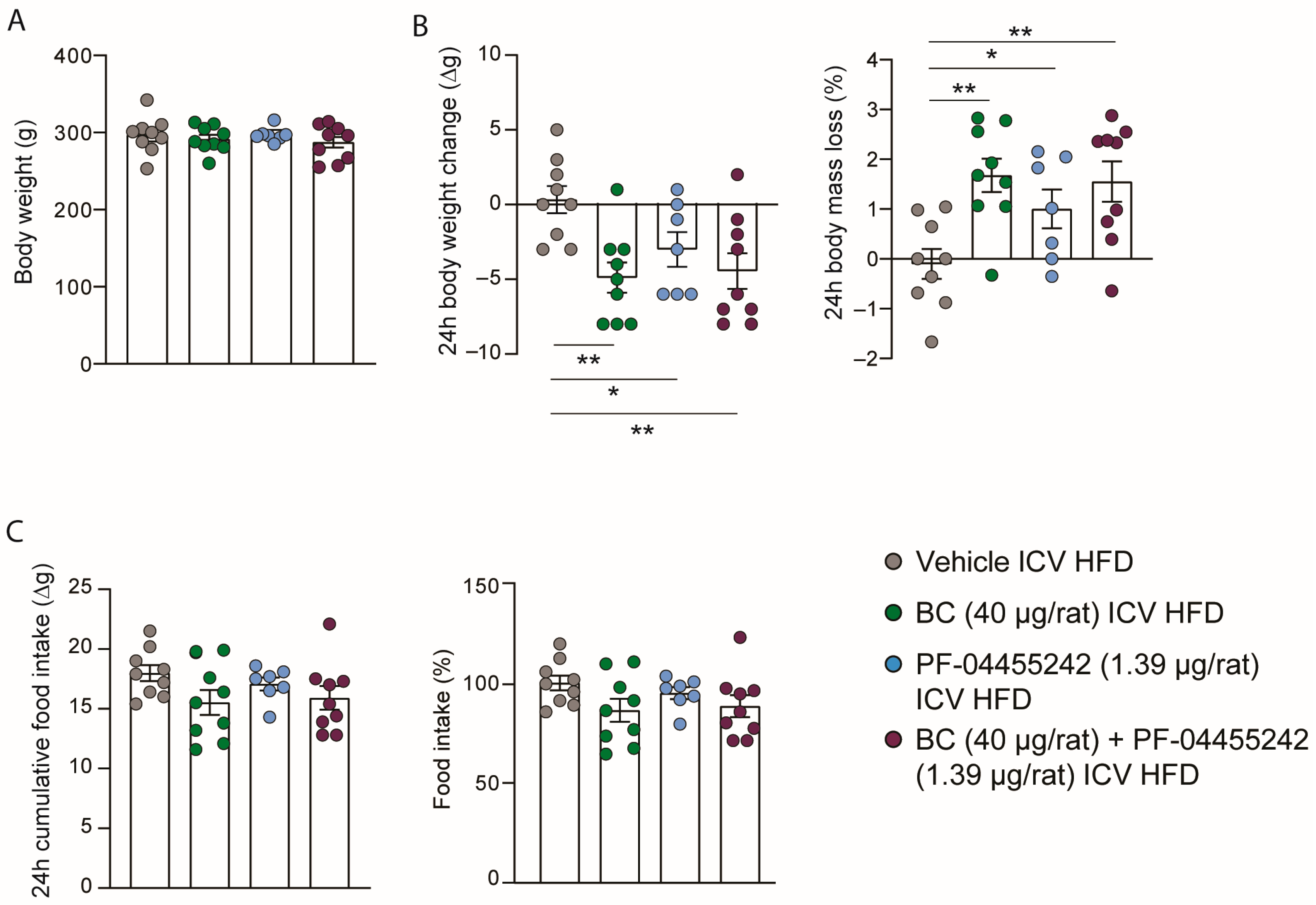
3.1. Central Administration of BC and PF-04455242 Decreases Body Weight in Combination and Individually
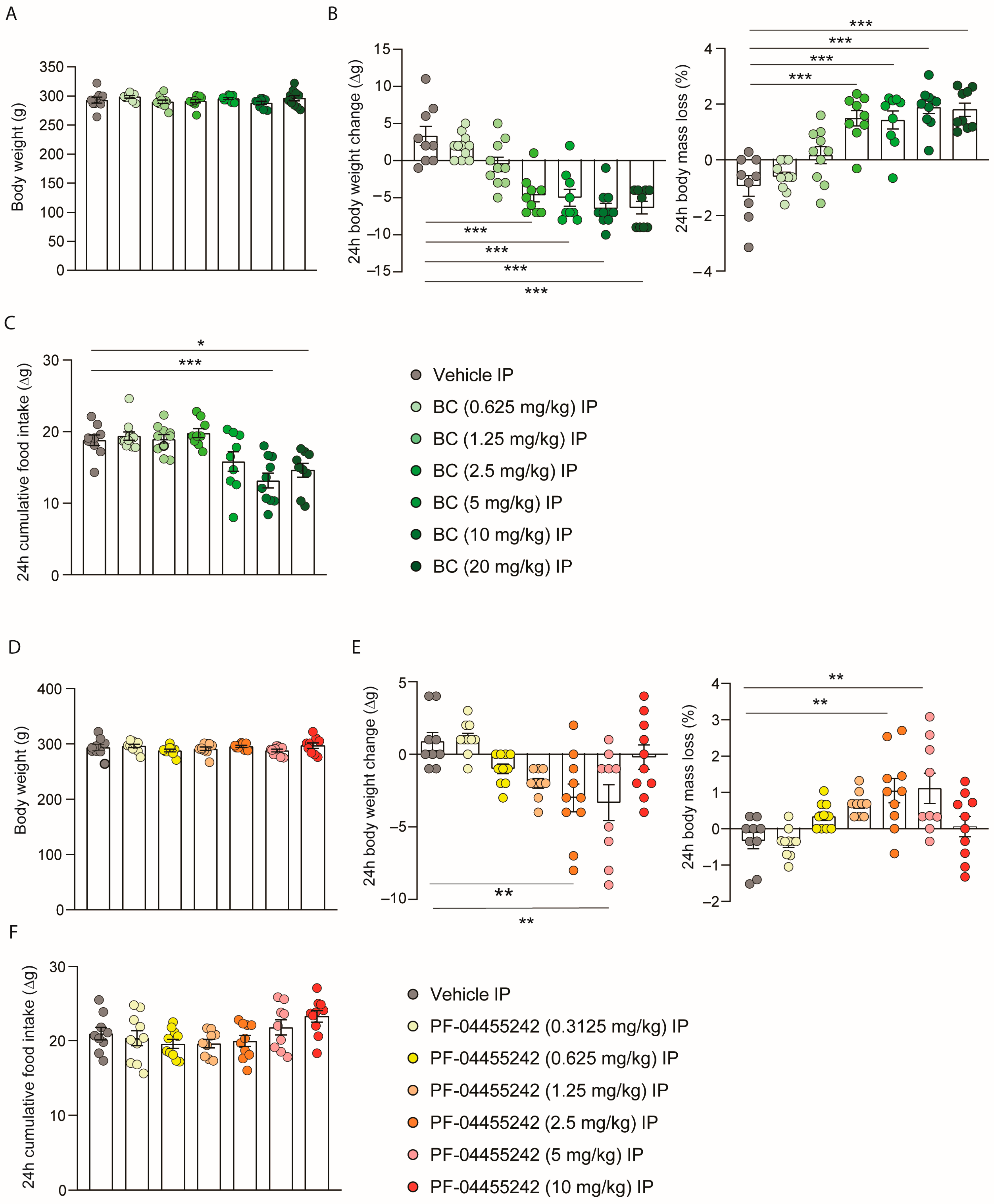
3.2. Peripheral Administration of BC and PF-04455242 Increases Body Weight Loss in a Dose-Dependent Manner
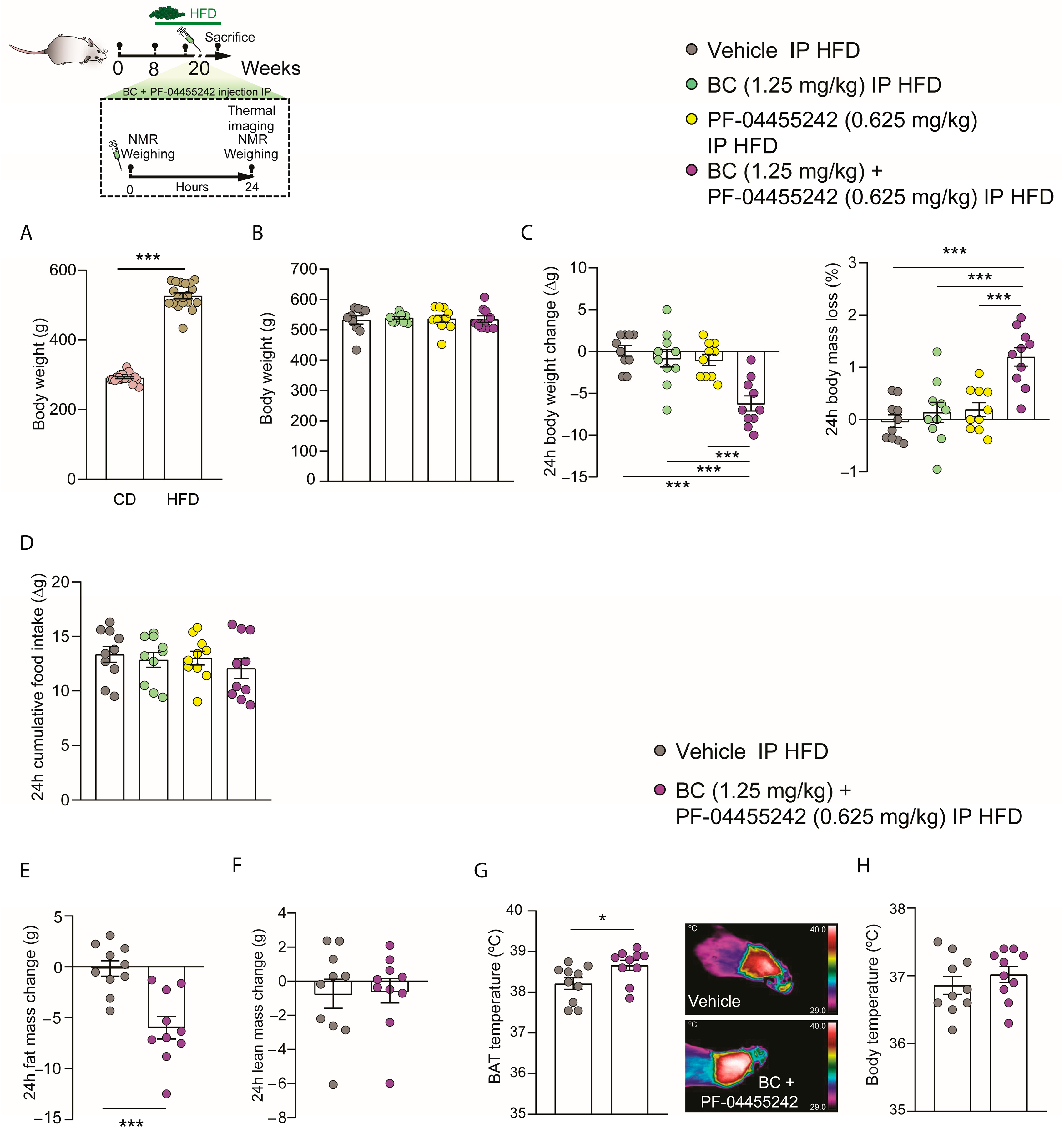
3.3. Acute Peripheral Combination of BC and PF-04455242 Causes Synergistic Increased Body Weight Loss in Diet-Induced Obese (DIO) Rats
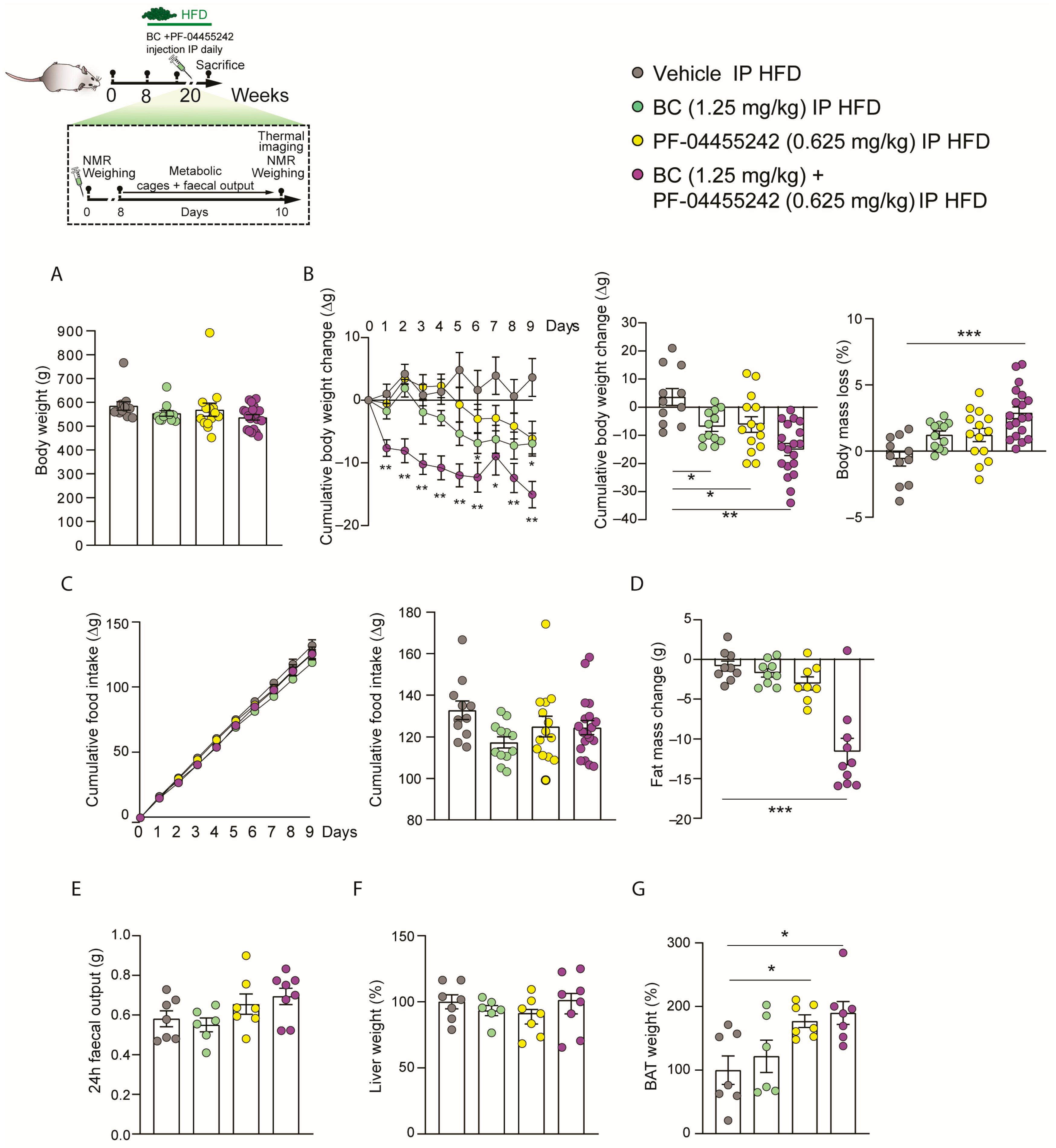
3.4. Chronic Peripheral Coadministration of BC and PF-04455242 Reduces Body Weight and Adiposity Independently of Food Intake in DIO Rats
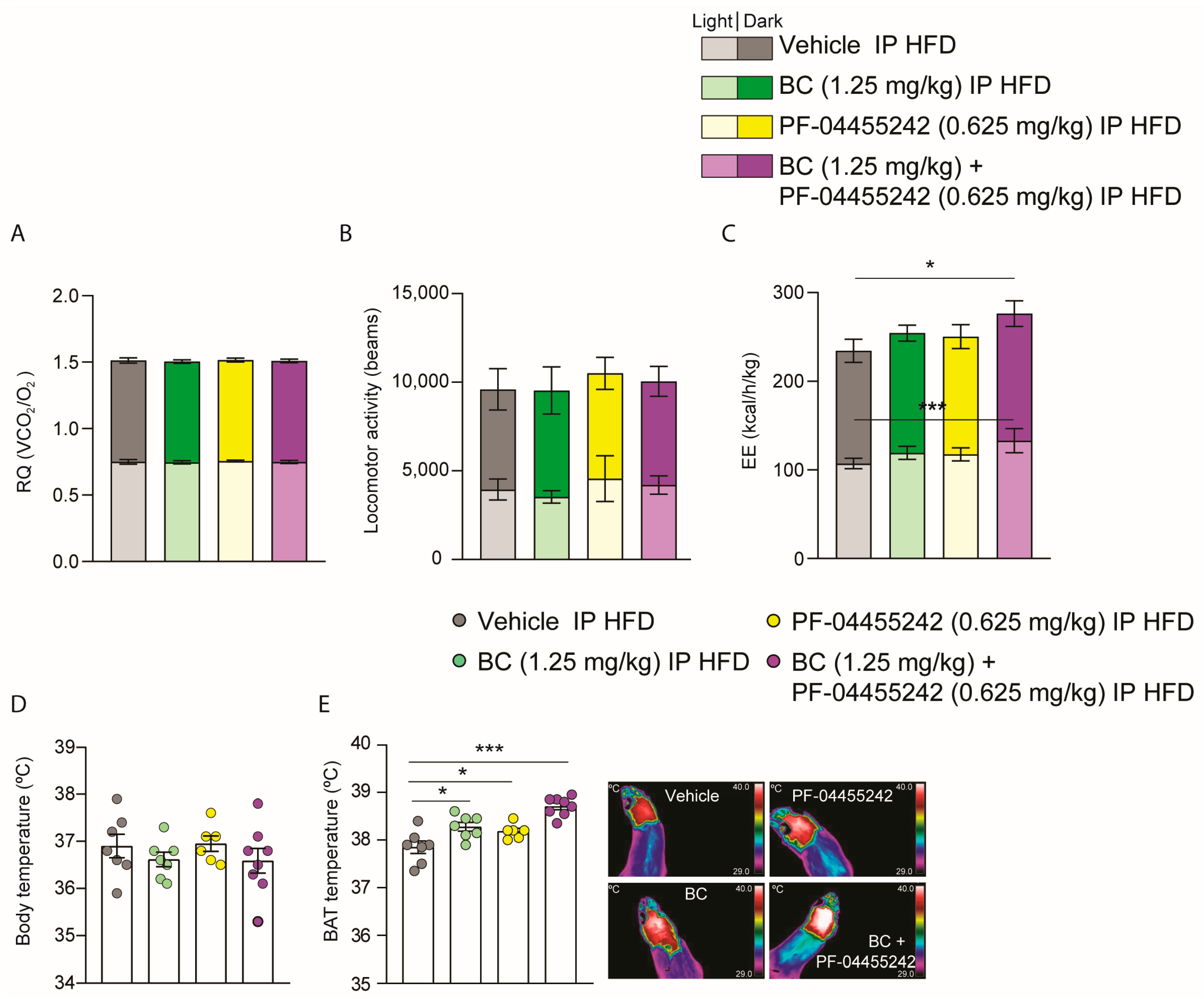
3.5. Chronic Peripheral Cotreatment with BC and PF-04455242 Induces Negative Energy Balance, Triggering Thermogenesis in DIO Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Obesity and Overweight. 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 1 April 2020).
- Kelly, T.; Yang, W.; Chen, C.S.; Reynolds, K.; He, J. Global burden of obesity in 2005 and projections to 2030. Int. J. Obes. 2008, 32, 1431–1437. [Google Scholar] [CrossRef]
- Heck, A.M.; Yanovski, J.A.; Calis, K.A. Orlistat, a new lipase inhibitor for the management of obesity. Pharmacotherapy 2000, 20, 270–279. [Google Scholar] [CrossRef]
- Idrees, Z.; Cancarevic, I.; Huang, L. FDA-Approved Pharmacotherapy for Weight Loss Over the Last Decade. Cureus 2022, 14, e29262. [Google Scholar] [CrossRef]
- Nogueiras, R.; Nauck, M.A.; Tschop, M.H. Gut hormone co-agonists for the treatment of obesity: From bench to bedside. Nat. Metab. 2023, 5, 933–944. [Google Scholar] [CrossRef]
- Salari, N.; Jafari, S.; Darvishi, N.; Valipour, E.; Mohammadi, M.; Mansouri, K.; Shohaimi, S. The best drug supplement for obesity treatment: A systematic review and network meta-analysis. Diabetol. Metab. Syndr. 2021, 13, 110. [Google Scholar] [CrossRef] [PubMed]
- Wadden, T.A.; Bailey, T.S.; Billings, L.K.; Davies, M.; Frias, J.P.; Koroleva, A.; Lingvay, I.; O’Neil, P.M.; Rubino, D.M.; Skovgaard, D.; et al. Effect of Subcutaneous Semaglutide vs. Placebo as an Adjunct to Intensive Behavioral Therapy on Body Weight in Adults with Overweight or Obesity: The STEP 3 Randomized Clinical Trial. JAMA 2021, 325, 1403–1413. [Google Scholar] [CrossRef] [PubMed]
- Jordan, J.; Astrup, A.; Engeli, S.; Narkiewicz, K.; Day, W.W.; Finer, N. Cardiovascular effects of phentermine and topiramate: A new drug combination for the treatment of obesity. J. Hypertens. 2014, 32, 1178–1188. [Google Scholar] [CrossRef] [PubMed]
- Makowski, C.T.; Gwinn, K.M.; Hurren, K.M. Naltrexone/bupropion: An investigational combination for weight loss and maintenance. Obes. Facts 2011, 4, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Folgueira, C.; Beiroa, D.; Porteiro, B.; Duquenne, M.; Puighermanal, E.; Fondevila, M.F.; Barja-Fernandez, S.; Gallego, R.; Hernandez-Bautista, R.; Castelao, C.; et al. Hypothalamic dopamine signaling regulates brown fat thermogenesis. Nat. Metab. 2019, 1, 811–829. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.J.; Volkow, N.D.; Logan, J.; Pappas, N.R.; Wong, C.T.; Zhu, W.; Netusil, N.; Fowler, J.S. Brain dopamine and obesity. Lancet 2001, 357, 354–357. [Google Scholar] [CrossRef] [PubMed]
- Pijl, H.; Ohashi, S.; Matsuda, M.; Miyazaki, Y.; Mahankali, A.; Kumar, V.; Pipek, R.; Iozzo, P.; Lancaster, J.L.; Cincotta, A.H.; et al. Bromocriptine: A novel approach to the treatment of type 2 diabetes. Diabetes Care 2000, 23, 1154–1161. [Google Scholar] [CrossRef]
- Holt, R.I.; Barnett, A.H.; Bailey, C.J. Bromocriptine: Old drug, new formulation and new indication. Diabetes Obes. Metab. 2010, 12, 1048–1057. [Google Scholar] [CrossRef]
- Cincotta, A.H.; Meier, A.H. Bromocriptine (Ergoset) reduces body weight and improves glucose tolerance in obese subjects. Diabetes Care 1996, 19, 667–670. [Google Scholar] [CrossRef] [PubMed]
- Glass, M.J.; Billington, C.J.; Levine, A.S. Opioids and food intake: Distributed functional neural pathways? Neuropeptides 1999, 33, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, R.J. Endogenous opiates and behavior: 2020. Peptides 2022, 151, 170752. [Google Scholar] [CrossRef] [PubMed]
- Darcq, E.; Kieffer, B.L. Opioid receptors: Drivers to addiction? Nat. Rev. Neurosci. 2018, 19, 499–514. [Google Scholar] [CrossRef] [PubMed]
- Greenway, F.L.; Fujioka, K.; Plodkowski, R.A.; Mudaliar, S.; Guttadauria, M.; Erickson, J.; Kim, D.D.; Dunayevich, E.; Group, C.-I.S. Effect of naltrexone plus bupropion on weight loss in overweight and obese adults (COR-I): A multicentre, randomised, double-blind, placebo-controlled, phase 3 trial. Lancet 2010, 376, 595–605. [Google Scholar] [CrossRef] [PubMed]
- Sainsbury, A.; Lin, S.; McNamara, K.; Slack, K.; Enriquez, R.; Lee, N.J.; Boey, D.; Smythe, G.A.; Schwarzer, C.; Baldock, P.; et al. Dynorphin knockout reduces fat mass and increases weight loss during fasting in mice. Mol. Endocrinol. 2007, 21, 1722–1735. [Google Scholar] [CrossRef] [PubMed]
- Czyzyk, T.A.; Nogueiras, R.; Lockwood, J.F.; McKinzie, J.H.; Coskun, T.; Pintar, J.E.; Hammond, C.; Tschop, M.H.; Statnick, M.A. kappa-Opioid receptors control the metabolic response to a high-energy diet in mice. FASEB J. 2010, 24, 1151–1159. [Google Scholar] [CrossRef]
- Romero-Pico, A.; Vazquez, M.J.; Gonzalez-Touceda, D.; Folgueira, C.; Skibicka, K.P.; Alvarez-Crespo, M.; Van Gestel, M.A.; Velasquez, D.A.; Schwarzer, C.; Herzog, H.; et al. Hypothalamic kappa-opioid receptor modulates the orexigenic effect of ghrelin. Neuropsychopharmacology 2013, 38, 1296–1307. [Google Scholar] [CrossRef]
- Romero-Pico, A.; Sanchez-Rebordelo, E.; Imbernon, M.; Gonzalez-Touceda, D.; Folgueira, C.; Senra, A.; Ferno, J.; Blouet, C.; Cabrera, R.; van Gestel, M.; et al. Melanin-Concentrating Hormone acts through hypothalamic kappa opioid system and p70S6K to stimulate acute food intake. Neuropharmacology 2018, 130, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Seoane-Collazo, P.; Linares-Pose, L.; Rial-Pensado, E.; Romero-Pico, A.; Moreno-Navarrete, J.M.; Martinez-Sanchez, N.; Garrido-Gil, P.; Iglesias-Rey, R.; Morgan, D.A.; Tomasini, N.; et al. Central nicotine induces browning through hypothalamic kappa opioid receptor. Nat. Commun. 2019, 10, 4037. [Google Scholar] [CrossRef] [PubMed]
- Romero-Pico, A.; Novelle, M.G.; Al-Massadi, O.; Beiroa, D.; Tojo, M.; Heras, V.; Ruiz-Pino, F.; Senra, A.; Lopez, M.; Blouet, C.; et al. Kappa-Opioid Receptor Blockade Ameliorates Obesity Caused by Estrogen Withdrawal via Promotion of Energy Expenditure through mTOR Pathway. Int. J. Mol. Sci. 2022, 23, 3118. [Google Scholar] [CrossRef] [PubMed]
- Grimwood, S.; Lu, Y.; Schmidt, A.W.; Vanase-Frawley, M.A.; Sawant-Basak, A.; Miller, E.; McLean, S.; Freeman, J.; Wong, S.; McLaughlin, J.P.; et al. Pharmacological characterization of 2-methyl-N-((2′-(pyrrolidin-1-ylsulfonyl)biphenyl-4-yl)methyl)propan-1-amine (PF-04455242), a high-affinity antagonist selective for kappa-opioid receptors. J. Pharmacol. Exp. Ther. 2011, 339, 555–566. [Google Scholar] [CrossRef]
- Decara, J.; Rivera, P.; Arrabal, S.; Vargas, A.; Serrano, A.; Pavon, F.J.; Dieguez, C.; Nogueiras, R.; Rodriguez de Fonseca, F.; Suarez, J. Cooperative role of the glucagon-like peptide-1 receptor and beta3-adrenergic-mediated signalling on fat mass reduction through the downregulation of PKA/AKT/AMPK signalling in the adipose tissue and muscle of rats. Acta Physiol. 2018, 222, e13008. [Google Scholar] [CrossRef] [PubMed]
- Beiroa, D.; Imbernon, M.; Gallego, R.; Senra, A.; Herranz, D.; Villarroya, F.; Serrano, M.; Ferno, J.; Salvador, J.; Escalada, J.; et al. GLP-1 agonism stimulates brown adipose tissue thermogenesis and browning through hypothalamic AMPK. Diabetes 2014, 63, 3346–3358. [Google Scholar] [CrossRef]
- Folgueira, C.; Beiroa, D.; Callon, A.; Al-Massadi, O.; Barja-Fernandez, S.; Senra, A.; Ferno, J.; Lopez, M.; Dieguez, C.; Casanueva, F.F.; et al. Uroguanylin Action in the Brain Reduces Weight Gain in Obese Mice via Different Efferent Autonomic Pathways. Diabetes 2016, 65, 421–432. [Google Scholar] [CrossRef]
- Pena-Leon, V.; Folgueira, C.; Barja-Fernandez, S.; Perez-Lois, R.; Da Silva Lima, N.; Martin, M.; Heras, V.; Martinez-Martinez, S.; Valero, P.; Iglesias, C.; et al. Prolonged breastfeeding protects from obesity by hypothalamic action of hepatic FGF21. Nat. Metab. 2022, 4, 901–917. [Google Scholar] [CrossRef] [PubMed]
- Imbernon, M.; Sanchez-Rebordelo, E.; Romero-Pico, A.; Kallo, I.; Chee, M.J.; Porteiro, B.; Al-Massadi, O.; Contreras, C.; Ferno, J.; Senra, A.; et al. Hypothalamic kappa opioid receptor mediates both diet-induced and melanin concentrating hormone-induced liver damage through inflammation and endoplasmic reticulum stress. Hepatology 2016, 64, 1086–1104. [Google Scholar] [CrossRef]
- Stanley, B.G.; Lanthier, D.; Leibowitz, S.F. Multiple brain sites sensitive to feeding stimulation by opioid agonists: A cannula-mapping study. Pharmacol. Biochem. Behav. 1988, 31, 825–832. [Google Scholar] [CrossRef]
- DePaoli, A.M.; Bell, G.I.; Stoffel, M. G protein-activated inwardly rectifying potassium channel (GIRK1/KGA) mRNA in adult rat heart and brain by in situ hybridization histochemistry. Mol. Cell Neurosci. 1994, 5, 515–522. [Google Scholar] [CrossRef]
- Tabarin, A.; Diz-Chaves, Y.; Carmona Mdel, C.; Catargi, B.; Zorrilla, E.P.; Roberts, A.J.; Coscina, D.V.; Rousset, S.; Redonnet, A.; Parker, G.C.; et al. Resistance to diet-induced obesity in mu-opioid receptor-deficient mice: Evidence for a “thrifty gene”. Diabetes 2005, 54, 3510–3516. [Google Scholar] [CrossRef]
- Kotz, C.M.; Grace, M.K.; Billington, C.J.; Levine, A.S. The effect of norbinaltorphimine, beta-funaltrexamine and naltrindole on NPY-induced feeding. Brain Res. 1993, 631, 325–328. [Google Scholar] [CrossRef]
- Lambert, P.D.; Wilding, J.P.; al-Dokhayel, A.A.; Gilbey, S.G.; Bloom, S.R. The effect of central blockade of kappa-opioid receptors on neuropeptide Y-induced feeding in the rat. Brain Res. 1993, 629, 146–148. [Google Scholar] [CrossRef]
- Hagan, M.M.; Rushing, P.A.; Benoit, S.C.; Woods, S.C.; Seeley, R.J. Opioid receptor involvement in the effect of AgRP-(83-132) on food intake and food selection. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 280, R814–R821. [Google Scholar] [CrossRef]
- Abraham, A.D.; Fontaine, H.M.; Song, A.J.; Andrews, M.M.; Baird, M.A.; Kieffer, B.L.; Land, B.B.; Chavkin, C. kappa-Opioid Receptor Activation in Dopamine Neurons Disrupts Behavioral Inhibition. Neuropsychopharmacology 2018, 43, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Pirino, B.E.; Spodnick, M.B.; Gargiulo, A.T.; Curtis, G.R.; Barson, J.R.; Karkhanis, A.N. Kappa-opioid receptor-dependent changes in dopamine and anxiety-like or approach-avoidance behavior occur differentially across the nucleus accumbens shell rostro-caudal axis. Neuropharmacology 2020, 181, 108341. [Google Scholar] [CrossRef] [PubMed]
- Escobar, A.D.P.; Casanova, J.P.; Andres, M.E.; Fuentealba, J.A. Crosstalk Between Kappa Opioid and Dopamine Systems in Compulsive Behaviors. Front. Pharmacol. 2020, 11, 57. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, R. Bromocriptine mesylate: FDA-approved novel treatment for type-2 diabetes. Indian J. Pharmacol. 2009, 41, 197–198. [Google Scholar] [CrossRef] [PubMed]
- Helal, M.A.; Habib, E.S.; Chittiboyina, A.G. Selective kappa opioid antagonists for treatment of addiction, are we there yet? Eur. J. Med. Chem. 2017, 141, 632–647. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cicuéndez, B.; Pérez-García, J.; Folgueira, C. A Combination of a Dopamine Receptor 2 Agonist and a Kappa Opioid Receptor Antagonist Synergistically Reduces Weight in Diet-Induced Obese Rodents. Nutrients 2024, 16, 424. https://doi.org/10.3390/nu16030424
Cicuéndez B, Pérez-García J, Folgueira C. A Combination of a Dopamine Receptor 2 Agonist and a Kappa Opioid Receptor Antagonist Synergistically Reduces Weight in Diet-Induced Obese Rodents. Nutrients. 2024; 16(3):424. https://doi.org/10.3390/nu16030424
Chicago/Turabian StyleCicuéndez, Beatriz, Javier Pérez-García, and Cintia Folgueira. 2024. "A Combination of a Dopamine Receptor 2 Agonist and a Kappa Opioid Receptor Antagonist Synergistically Reduces Weight in Diet-Induced Obese Rodents" Nutrients 16, no. 3: 424. https://doi.org/10.3390/nu16030424
APA StyleCicuéndez, B., Pérez-García, J., & Folgueira, C. (2024). A Combination of a Dopamine Receptor 2 Agonist and a Kappa Opioid Receptor Antagonist Synergistically Reduces Weight in Diet-Induced Obese Rodents. Nutrients, 16(3), 424. https://doi.org/10.3390/nu16030424