
B Cells Dynamic in Aging and the Implications of Nutritional Regulation
 ,
, 
Abstract
:1. Introduction
2. A Brief Overview of the Key Types of B Cells Involved in the Aging Process
3. Changes in the Number and Function of B Cells during Aging
3.1. Aging and B-1 Cells
3.2. Aging and B-2 Cells
3.3. Aging and Regulatory B Cells
3.4. Aging and Age-Associated B Cells
4. The Role of B Cells in Autoimmune Diseases
4.1. Multiple Functions of B Cells in the Pathogenesis of Systemic Lupus Erythematosus
4.1.1. ABCs in SLE
4.1.2. Innate-like B Cells in SLE
4.1.3. Bregs in SLE
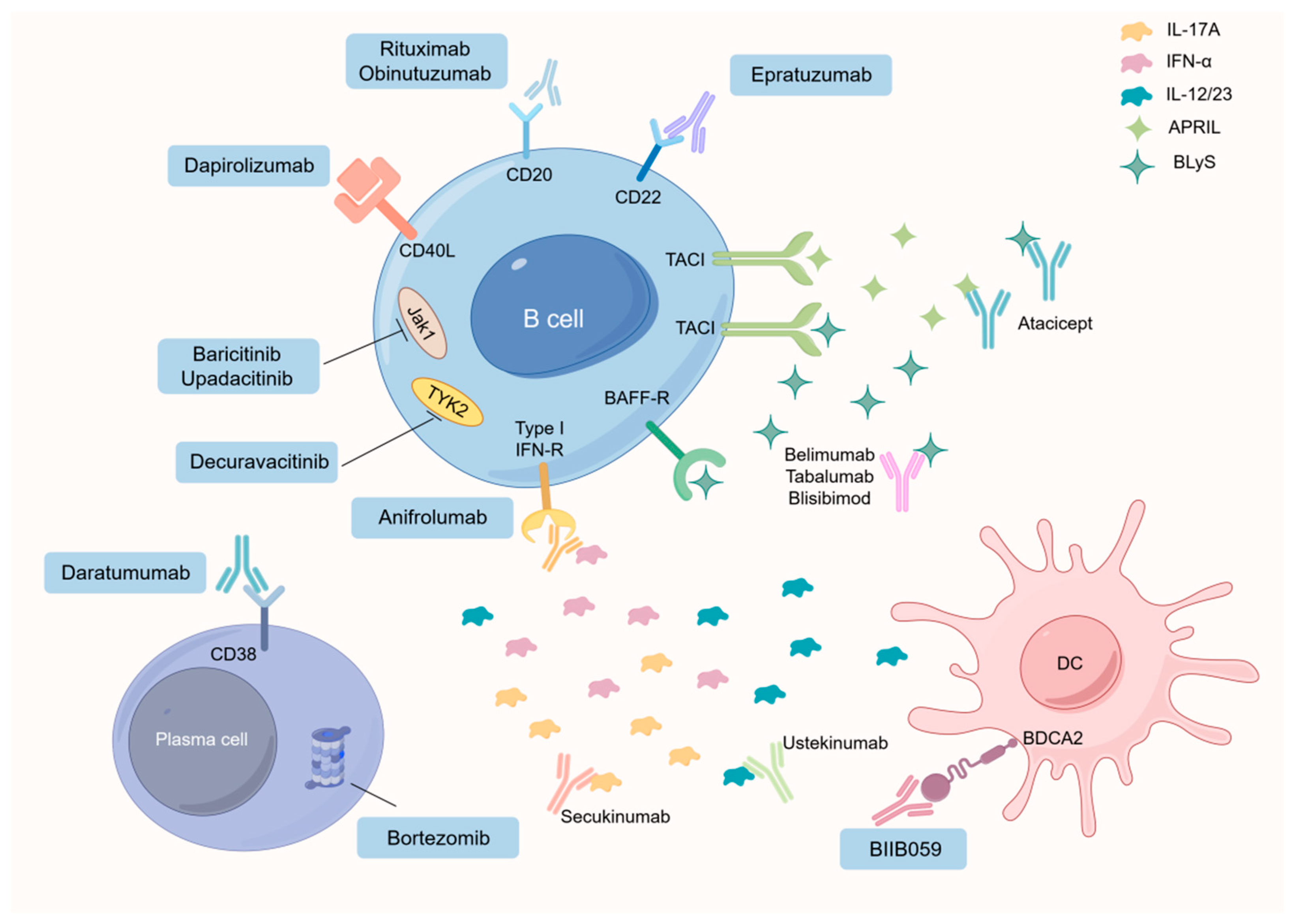
4.1.4. Advancements in SLE Treatment of B Cells
4.2. Multiple Functions of B Cells in the Pathogenesis of Primary Sjögren’s Syndrome
4.2.1. Bregs in pSS
4.2.2. CD11c+ ABCs in pSS
4.2.3. Marginal Zone B Cells in pSS
4.2.4. Memory B Cells in pSS
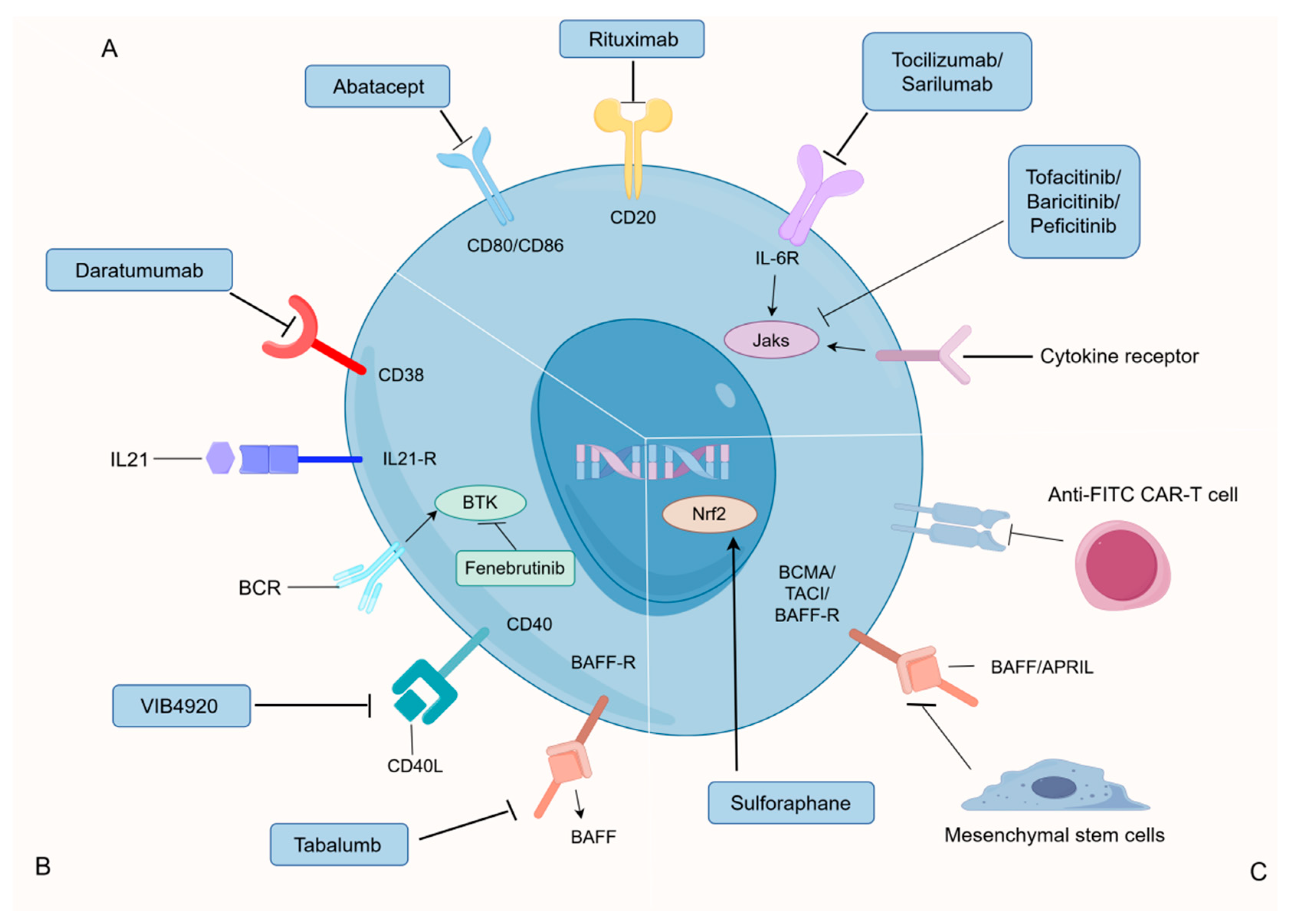
4.2.5. Advancements in pSS Treatment of B Cells
4.3. Multiple Functions of B Cells in the Pathogenesis of Rheumatoid Arthritis
4.3.1. Bregs in RA
4.3.2. Age-Associated B Cells in RA
4.3.3. Plasma Cell in RA
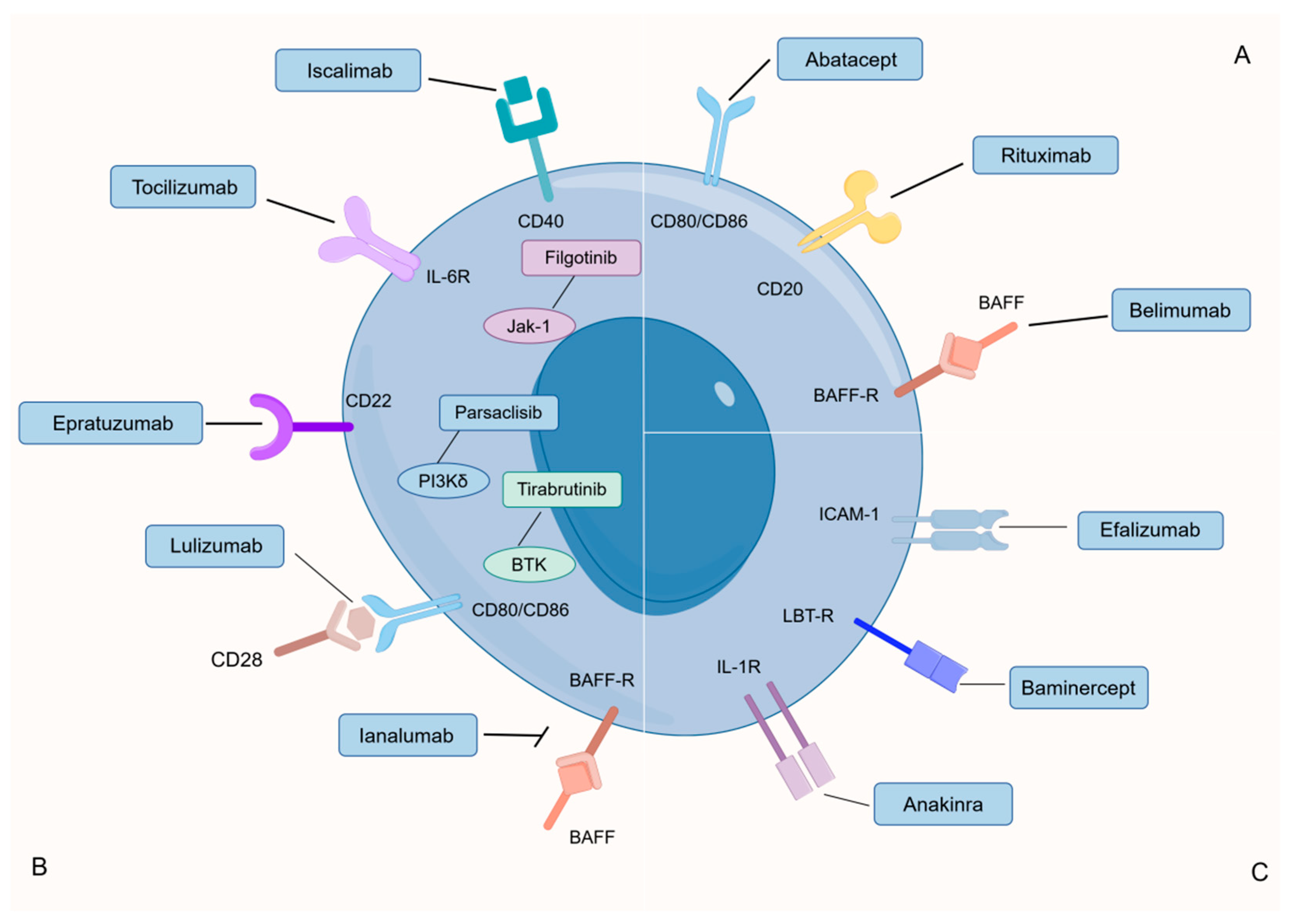
4.3.4. Advancements in RA Treatment of B Cells
5. Nutrients Regulate B Cells and Their Implications for Autoimmune Diseases
5.1. Effects of Energy on B Cells Dynamic Changes
5.2. Effects of Carbohydrates on B Cells Dynamic Changes
5.3. Effects of Amino Acids on B Cells Dynamic Changes
5.4. Effects of Vitamins on B Cells Dynamic Changes
5.5. Effects of Lipids on B Cells Dynamic Changes
5.5.1. Effects of Lipid Synthesis on B Cells Function
5.5.2. Effect of Lipid Uptake and Catabolism on B Cells Function
5.5.3. Potential Therapeutic Strategies for Lipids in Autoimmune Diseases
6. Conclusions and Future Perspectives
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yousefzadeh, M.J.; Flores, R.R.; Zhu, Y.; Schmiechen, Z.C.; Brooks, R.W.; Trussoni, C.E.; Cui, Y.X.; Angelini, L.; Lee, K.A.; McGowan, S.J.; et al. An aged immune system drives senescence and ageing of solid organs. Nature 2021, 594, 100. [Google Scholar] [CrossRef]
- Rodriguez-Zhurbenko, N.; Quach, T.D.; Hopkins, T.J.; Rothstein, T.L.; Hemandez, A.M. Human B-1 Cells and B-1 Cell Antibodies Change with Advancing Age. Front. Immunol. 2019, 10, 483. [Google Scholar] [CrossRef]
- Ma, S.; Wang, C.; Mao, X.; Hao, Y. B Cell Dysfunction Associated with Aging and Autoimmune Diseases. Front. Immunol. 2019, 10, 318. [Google Scholar] [CrossRef]
- Baumgarth, N. B-1 Cell Heterogeneity and the Regulation of Natural and Antigen-induced IgM Production. Front. Immunol. 2016, 7, 324. [Google Scholar] [CrossRef] [PubMed]
- Tsiantoulas, D.; Diehl, C.J.; Witztum, J.L.; Binder, C.J. B Cells and Humoral Immunity in Atherosclerosis. Circ. Res. 2014, 114, 1743–1756. [Google Scholar] [CrossRef] [PubMed]
- Halperin, S.T.; T Hart, B.A.; Luchicchi, A.; Schenk, G.J. The Forgotten Brother: The Innate-like B1 Cell in Multiple Sclerosis. Biomedicines 2022, 10, 606. [Google Scholar] [CrossRef]
- Ma, K.Y.; Du, W.H.; Wang, S.Y.; Xiao, F.; Li, J.Y.; Tian, J.; Xing, Y.D.; Kong, X.D.; Rui, K.; Qin, R.C.; et al. B1-cell-produced anti-phosphatidylserine antibodies contribute to lupus nephritis development via TLR-mediated Syk activation. Cell. Mol. Immunol. 2023, 20, 881–894. [Google Scholar] [CrossRef]
- Torring, C.; Petersen, C.C.; Bjerg, L.; Kofod-Olsen, E.; Petersen, T.; Hollsberg, P. The B1-cell subpopulation is diminished in patients with relapsing-remitting multiple sclerosis. J. Neuroimmunol. 2013, 262, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Rauch, M.; Tussiwand, R.; Bosco, N.; Rolink, A.G. Crucial Role for BAFF-BAFF-R Signaling in the Survival and Maintenance of Mature B Cells. PLoS ONE 2009, 4, e5456. [Google Scholar] [CrossRef]
- Zouali, M.; Richard, Y. Marginal zone B-cells, a gatekeeper of innate immunity. Front. Immunol. 2011, 2, 63. [Google Scholar] [CrossRef]
- Tanigakik, K.; Han, H.; Yamamoto, N.; Tashiro, K.; Ikegawa, M.; Kuroda, K.; Suzuki, A.; Nakano, T.; Honjo, T. Notch-RBP-J signaling is involved in cell fate determination of marginal zone B cells. Nat. Immunol. 2002, 3, 443–450. [Google Scholar] [CrossRef]
- Weisel, F.J.; Zuccarino-Catania, G.V.; Chikina, M.; Shlomchik, M.J. A Temporal Switch in the Germinal Center Determines Differential Output of Memory B and Plasma Cells. Immunity 2016, 44, 116–130. [Google Scholar] [CrossRef] [PubMed]
- Rosser, E.C.; Mauri, C. Regulatory B Cells: Origin, Phenotype, and Function. Immunity 2015, 42, 607–612. [Google Scholar] [CrossRef] [PubMed]
- Yanaba, K.; Bouaziz, J.D.; Haas, K.M.; Poe, J.C.; Fujimoto, M.; Tedder, T.F. A regulatory B cell subset with a unique CD1d(hi)CD5(+) phenotype controls T cell-dependent inflammatory responses. Immunity 2008, 28, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Sachinidis, A.; Xanthopoulos, K.; Garyfallos, A. Age-Associated B Cells (ABCs) in the Prognosis, Diagnosis and Therapy of Systemic Lupus Erythematosus (SLE). Mediterr. J. Rheumatol. 2020, 31, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Deng, J.C.; Goldstein, D.R. How aging impacts vaccine efficacy: Known molecular and cellular mechanisms and future directions. Trends Mol. Med. 2022, 28, 1100–1111. [Google Scholar] [CrossRef] [PubMed]
- Barber, C.L.; Montecino-Rodriguez, E.; Dorshkind, K. Reduced production of B-1-specified common lymphoid progenitors results in diminished potential of adult marrow to generate B-1 cells. Proc. Natl. Acad. Sci. USA 2011, 108, 13700–13704. [Google Scholar] [CrossRef] [PubMed]
- Alter-Wolf, S.; Blomberg, B.B.; Riley, R.L. Old mice retain bone marrow B1 progenitors, but lose B2 precursors, and exhibit altered immature B cell phenotype and light chain usage. Mech. Ageing Dev. 2009, 130, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Savage, H.P.; Yenson, V.M.; Sawhney, S.S.; Mousseau, B.J.; Lund, F.E.; Baumgarth, N. Blimp-1-dependent and -independent natural antibody production by B-1 and B-1-derived plasma cells. J. Exp. Med. 2017, 214, 2777–2794. [Google Scholar] [CrossRef]
- Inui, M.; Hirota, S.; Hirano, K.; Fujii, H.; Sugahara-Tobinai, A.; Ishii, T.; Harigae, H.; Takai, T. Human CD43+ B cells are closely related not only to memory B cells phenotypically but also to plasmablasts developmentally in healthy individuals. Int. Immunol. 2015, 27, 345–355. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, C.L.; Yang, Q.Y.; Kantor, A.B.; Chu, H.T.; Ghosn, E.; Qin, G.; Mazmanian, S.K.; Han, J.; Herzenberg, L.A. Distinct mechanisms define murine B cell lineage immunoglobulin heavy chain (IgH) repertoires. elife 2015, 4, e09083. [Google Scholar] [CrossRef]
- Wong, J.B.; Hewitt, S.L.; Heltemes-Harris, L.M.; Mandal, M.; Johnson, K.; Rajewsky, K.; Koralov, S.B.; Clark, M.R.; Farrar, M.A.; Skok, J.A. B-1a cells acquire their unique characteristics by bypassing the pre-BCR selection stage. Nat. Commun. 2019, 10, 4768. [Google Scholar] [CrossRef]
- Wang, H.; Shin, D.; Abbasi, S.; Jain, S.; Kovalchuk, A.L.; Beaty, N.; Chen, S.; Gonzalez-Garcia, I.; Morse, H.C. Expression of plasma cell alloantigen 1 defines layered development of B-1a B-cell subsets with distinct innate-like functions. Proc. Natl. Acad. Sci. USA 2012, 109, 20077–20082. [Google Scholar] [CrossRef]
- Adler, H.; Ferreira, D.M.; Gordon, S.B.; Rylance, J. Pneumococcal Capsular Polysaccharide Immunity in the Elderly. Clin. Vaccine Immunol. 2017, 24, e00004-17. [Google Scholar] [CrossRef]
- Haas, K.M.; Poe, J.C.; Steeber, D.A.; Tedder, T.F. B-1a and b-1b cells exhibit distinct developmental requirements and have unique functional roles in innate and adaptive immunity to S-pneumoniae. Immunity 2005, 23, 7–18. [Google Scholar] [CrossRef]
- Holodick, N.E.; Vizconde, T.; Hopkins, T.J.; Rothstein, T.L. Age-Related Decline in Natural IgM Function: Diversification and Selection of the B-1a Cell Pool with Age. J. Immunol. 2016, 196, 4348–4357. [Google Scholar] [CrossRef] [PubMed]
- Holodick, N.E.; Repetny, K.; Zhong, X.; Rothstein, T.L. Adult BM generates CD5(+) B1 cells containing abundant N-region additions. Eur. J. Immunol. 2009, 39, 2383–2394. [Google Scholar] [CrossRef] [PubMed]
- Hu, A.; Ehleiter, D.; Benyehuda, A.; Schwab, R.; Russo, C.; Szabo, P.; Weksler, M.E. Effect of Age on The Expressed B-Cell Repertoire-Role of B-Cell Subsets. Int. Immunol. 1993, 5, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Frasca, D.; Van Der Put, E.; Riley, R.L.; Blomberg, B.B. Reduced Ig class switch in aged mice correlates with decreased E47 and activation-induced cytidine deaminase. J. Immunol. 2004, 172, 2155–2162. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.P.; Cancro, M.P. B cells and aging: Balancing the homeostatic equation. Exp. Gerontol. 2007, 42, 396–399. [Google Scholar] [CrossRef] [PubMed]
- Bulati, M.; Buffa, S.; Candore, G.; Caruso, C.; Dunn-Walters, D.K.; Pellicano, M.; Wu, Y.C.; Romano, G.C. B cells and immunosenescence: A focus on IgG(+)IgD(−)CD27(−) (DN) B cells in aged humans. Ageing Res. Rev. 2011, 10, 274–284. [Google Scholar] [CrossRef]
- Pillai, S.; Cariappa, A.; Moran, S.T. Marginal zone B cells. Annu. Rev. Immunol. 2005, 23, 161–196. [Google Scholar] [CrossRef] [PubMed]
- Mebius, R.E.; Nolte, M.A.; Kraal, G. Development and function of the splenic marginal zone. Crit. Rev. Immunol. 2004, 24, 449–464. [Google Scholar] [CrossRef] [PubMed]
- Kraal, G.; Mebius, R. New insights into the cell biology of the marginal zone of the spleen. Int. Rev. Cytol. 2006, 205, 175–215. [Google Scholar]
- Birjandi, S.Z.; Ippolito, J.A.; Ramadorai, A.K.; Witte, P.L. Alterations in Marginal Zone Macrophages and Marginal Zone B Cells in Old Mice. J. Immunol. 2011, 186, 3441–3451. [Google Scholar] [CrossRef] [PubMed]
- Frasca, D.; Landin, M.; Lechner, S.C.; Ryan, J.G.; Schwartz, R.; Riley, R.L.; Blomberg, B.B. Aging down-regulates the transcription factor E2A, activation-induced cytidine deaminase, and Ig class switch in human B cells. J. Immunol. 2008, 180, 5283–5290. [Google Scholar] [CrossRef]
- Cortegano, I.; Rodriguez, M.; Martin, I.; Carmen Prado, M.; Ruiz, C.; Hortiguela, R.; Alia, M.; Vilar, M.; Mira, H.; Cano, E.; et al. Altered marginal zone and innate-like B cells in aged senescence-accelerated SAMP8 mice with defective IgG1 responses. Cell Death Dis. 2017, 8, e3000. [Google Scholar] [CrossRef] [PubMed]
- Allman, D.; Miller, J.P. B cell development and receptor diversity during aging. Curr. Opin. Immunol. 2005, 17, 463–467. [Google Scholar] [CrossRef]
- Frasca, D.; Nguyen, D.; Van Der Put, E.; Riley, R.L.; Blomberg, B.B. The age-related decrease in E47 DNA-binding does not depend on increased Id inhibitory proteins in bone marrow-derived B cell precursors. Front. Biosci. 2003, 8, A110–A116. [Google Scholar] [CrossRef]
- Frasca, D.; Nguyen, D.; Riley, R.L.; Blomberg, B.B. Effects of aging on DNA-binding activity of the E47 transcription factor in splenic B cells. Mech. Ageing Dev. 2004, 125, 111–112. [Google Scholar] [CrossRef]
- Jin, R.; Kaneko, H.; Suzuki, H.; Arai, T.; Teramoto, T.; Fukao, T.; Kondo, N. Age-related changes in BAFF and APRIL profiles and upregulation of BAFF and APRIL expression in patients with primary antibody deficiency. Int. J. Mol. Med. 2008, 21, 233–238. [Google Scholar] [CrossRef]
- Gray, M.; Miles, K.; Salter, D.; Gray, D.; Savill, J. Apoptotic cells protect mice from autoimmune inflammation by the induction of regulatory B cells. Proc. Natl. Acad. Sci. USA 2007, 104, 14080–14085. [Google Scholar] [CrossRef]
- Matsumoto, M.; Baba, A.; Yokota, T.; Nishikawa, H.; Ohkawa, Y.; Kayama, H.; Kallies, A.; Nutt, S.L.; Sakaguchi, S.; Takeda, K.; et al. Interleukin-10-Producing Plasmablasts Exert Regulatory Function in Autoimmune Inflammation. Immunity 2014, 41, 1040–1051. [Google Scholar] [CrossRef]
- Liu, X.M.; Jiang, X.C.; Liu, R.H.; Wang, L.M.; Qian, T.T.; Zheng, Y.J.; Deng, Y.T.; Huang, E.Y.; Xu, F.K.; Wang, J.Y.; et al. B cells expressing CD11b effectively inhibit CD4+T-cell responses and ameliorate experimental autoimmune hepatitis in mice. Hepatology 2015, 62, 1563–1575. [Google Scholar] [CrossRef]
- Johnson, B.A.; Kahler, D.J.; Baban, B.; Chandler, P.R.; Kang, B.L.; Shimoda, M.; Koni, P.A.; Pihkala, J.; Vilagos, B.; Busslinger, M.; et al. B-lymphoid cells with attributes of dendritic cells regulate T cells via indoleamine 2,3-dioxygenase. Proc. Natl. Acad. Sci. USA 2010, 107, 10644–10648. [Google Scholar] [CrossRef]
- Hao, Y.; O’neill, P.; Naradikian, M.S.; Scholz, J.L.; Cancro, M.P. A B-cell subset uniquely responsive to innate stimuli accumulates in aged mice. Blood 2011, 118, 1294–1304. [Google Scholar] [CrossRef] [PubMed]
- Rubtsov, A.V.; Rubtsova, K.; Fischer, A.; Meehan, R.T.; Gillis, J.Z.; Kappler, J.W.; Marrack, P. Toll-like receptor 7 (TLR7)-driven accumulation of a novel CD11c(+) B-cell population is important for the development of autoimmunity. Blood 2011, 118, 1305–1315. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Z.; Li, Z.G.; Hu, F.L. Double-negative (DN) B cells: An under-recognized effector memory B cell subset in autoimmunity. Clin. Exp. Immunol. 2021, 205, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Sanz, I.; Wei, C.; Jenks, S.A.; Cashman, K.S.; Tipton, C.; Woodruff, M.C.; Hom, J.; Lee, F.E. Challenges and Opportunities for Consistent Classification of Human B Cell and Plasma Cell Populations. Front. Immunol. 2019, 10, 2458. [Google Scholar] [CrossRef] [PubMed]
- Claes, N.; Fraussen, J.; Vanheusden, M.; Hellings, N.; Stinissen, P.; Van Wijmeersch, B.; Hupperts, R.; Somers, V. Age-Associated B Cells with Proinflammatory Characteristics Are Expanded in a Proportion of Multiple Sclerosis Patients. J. Immunol. 2016, 197, 4576–4583. [Google Scholar] [CrossRef] [PubMed]
- Adlowitz, D.G.; Barnard, J.; Biear, J.N.; Cistrone, C.; Owen, T.; Wang, W.S.; Palanichamy, A.; Ezealah, E.; Campbell, D.; Wei, C.W.; et al. Expansion of Activated Peripheral Blood Memory B Cells in Rheumatoid Arthritis, Impact of B Cell Depletion Therapy, and Biomarkers of Response. PLoS ONE 2015, 10, e0128269. [Google Scholar] [CrossRef]
- Frasca, D.; Blomberg, B.B. Aging induces B cell defects and decreased antibody responses to influenza infection and vaccination. Immun. Ageing 2020, 17, 1–10. [Google Scholar] [CrossRef]
- Naradikian, M.S.; Hao, Y.; Cancro, M.P. Age-associated B cells: Key mediators of both protective and autoreactive humoral responses. Immunol. Rev. 2016, 269, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Rubtsova, K.; Rubtsov, A.V.; Cancro, M.P.; Marrack, P. Age-Associated B Cells: A T-bet Dependent Effector with Roles in Protective and Pathogenic Immunity. J. Immunol. 2015, 195, 1933–1937. [Google Scholar] [CrossRef]
- Riley, R.L.; Khomtchouk, K.; Blomberg, B.B. Age-associated B cells (ABC) inhibit B lymphopoiesis and alter antibody repertoires in old age. Cell. Immunol. 2017, 321, 61–67. [Google Scholar] [CrossRef]
- Naradikian, M.S.; Myles, A.; Beiting, D.P.; Roberts, K.J.; Dawson, L.; Herati, R.S.; Bengsch, B.; Linderman, S.L.; Stelekati, E.; Spolski, R.; et al. Cutting Edge: IL-4, IL-21, and IFN-gamma Interact to Govern T-bet and CD11c Expression in TLR-Activated B Cells. J. Immunol. 2016, 197, 1023–1028. [Google Scholar] [CrossRef]
- Karnell, J.L.; Kumar, V.; Wang, J.Y.; Wang, S.; Voynova, E.; Ettinger, R. Role of CD11c(+) T-bet(+) B cells in human health and disease. Cell. Immunol. 2017, 321, 40–45. [Google Scholar] [CrossRef]
- Cancro, M.P. Age-Associated B Cells. Annu. Rev. Immunol. 2020, 38, 315–340. [Google Scholar] [CrossRef]
- Ratliff, M.; Alter, S.; Frasca, D.; Blomberg, B.B.; Riley, R.L. In senescence, age-associated B cells secrete TNF and inhibit survival of B-cell precursors. Aging Cell 2013, 12, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Phalke, S.; Marrack, P. Age (autoimmunity) associated B cells (ABCs) and their relatives. Curr. Opin. Immunol. 2018, 55, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Winer, D.A.; Winer, S.; Shen, L.; Wadia, P.P.; Yantha, J.; Paltser, G.; Tsui, H.; Wu, P.; Davidson, M.G.; Alonso, M.N.; et al. B cells promote insulin resistance through modulation of T cells and production of pathogenic IgG antibodies. Nat. Med. 2011, 17, 134–610. [Google Scholar] [CrossRef]
- Zeng, Q.H.; Wei, Y.; Lao, X.M.; Chen, D.P.; Huang, C.X.; Lin, Q.Y.; He, M.; Liao, Y.; Zheng, L.M.; Li, B.; et al. B cells polarize pathogenic inflammatory T helper subsets through ICOSL-dependent glycolysis. Sci. Adv. 2020, 6, eabb6296. [Google Scholar] [CrossRef] [PubMed]
- Frasca, D.; Romero, M.; Garcia, D.; Diaz, A.; Blomberg, B.B. Hyper-metabolic B cells in the spleens of old mice make antibodies with autoimmune specificities. Immun. Ageing 2021, 18, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lescale, C.; Dias, S.; Maes, J.; Cumano, A.; Szabo, P.; Charron, D.; Weksler, M.E.; Dosquet, C.; Vieira, P.; Goodhardt, M. Reduced EBF expression underlies loss of B-cell potential of hematopoietic progenitors with age. Aging Cell 2010, 9, 410–419. [Google Scholar] [CrossRef] [PubMed]
- King, A.M.; Keating, P.; Prabhu, A.; Blomberg, B.B.; Riley, R.L. NK cells in the CD19(−) B220(+) bone marrow fraction are increased in senescence and reduce E2A and surrogate light chain proteins in B cell precursors. Mech. Ageing Dev. 2009, 130, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Nemazee, D. Mechanisms of central tolerance for B cells. Nat. Rev. Immunol. 2017, 17, 281–294. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Han, M.; Rui, K.; Ai, X.Y.; Tian, J.; Zhang, W.; Zhao, F.T.; Zhao, Y.; Jiang, Q.; Lu, L.W. New insights into follicular helper T cell response and regulation in autoimmune pathogenesis. Cell. Mol. Immunol. 2021, 18, 1610–1612. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.Y.; Du, W.H.; Wang, X.H.; Yuan, S.W.; Cai, X.Y.; Liu, D.Z.; Li, J.Y.; Lu, L.W. Multiple Functions of B Cells in the Pathogenesis of Systemic Lupus Erythematosus. Int. J. Mol. Sci. 2019, 20, 6021. [Google Scholar] [CrossRef] [PubMed]
- Du, W.H.; Han, M.; Zhu, X.X.; Xiao, F.; Huang, E.Y.; Che, N.; Tang, X.P.; Zou, H.J.; Jiang, Q.; Lu, L.W. The Multiple Roles of B Cells in the Pathogenesis of Sjogren’s Syndrome. Front. Immunol. 2021, 12, 684999. [Google Scholar] [CrossRef]
- Medzhitov, R.; Janeway, C.A. Decoding the patterns of self and nonself by the innate immune system. Science 2002, 296, 298–300. [Google Scholar] [CrossRef]
- Tipton, C.M.; Fucile, C.F.; Darce, J.; Chida, A.; Ichikawa, T.; Gregoretti, I.; Schieferl, S.; Hom, J.; Jenks, S.; Feldman, R.J.; et al. Diversity, cellular origin and autoreactivity of antibody-secreting cell population expansions in acute systemic lupus erythematosus. Nat. Immunol. 2015, 16, 755. [Google Scholar] [CrossRef]
- Cancro, M.P. Expanding roles for the Tbet plus B cell subset in health and disease. Cell. Immunol. 2017, 321, 1–2. [Google Scholar] [CrossRef]
- Ma, K.Y.; Li, J.Y.; Fang, Y.F.; Lu, L.W. Roles of B Cell-Intrinsic TLR Signals in Systemic Lupus Erythematosus. Int. J. Mol. Sci. 2015, 16, 13084–13105. [Google Scholar] [CrossRef]
- Martin, F.; Oliver, A.M.; Kearney, J.F. Marginal zone and B1B cells unite in the early response against T-independent blood-borne particulate antigens. Immunity 2001, 14, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Lenert, P.; Brummel, R.; Field, E.H.; Ashman, R.F. TLR-9 activation of marginal zone B cells in lupus mice regulates immunity through increased IL-10 production. J. Clin. Immunol. 2005, 25, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Barr, T.A.; Shen, P.; Brown, S.; Lampropoulou, V.; Roch, T.; Lawrie, S.; Fan, B.; O’Connor, R.A.; Anderton, S.M.; Bar-Or, A.; et al. B cell depletion therapy ameliorates autoimmune disease through ablation of IL-6-producing B cells. J. Exp. Med. 2012, 209, 1001–1010. [Google Scholar] [CrossRef]
- Duddy, M.E.; Alter, A.; Bar-Or, A. Distinct profiles of human B cell effector cytokines: A role in immune regulation? J. Immunol. 2004, 172, 3422–3427. [Google Scholar] [CrossRef] [PubMed]
- Miyagaki, T. Regulatory B cells in human autoimmune diseases. Nihon Rinsho Men’eki Gakkai Kaishi Jpn. J. Clin. Immunol. 2015, 38, 390–397. [Google Scholar] [CrossRef]
- Green, N.M.; Marshak-Rothstein, A. Toll-like receptor driven B cell activation in the induction of systemic autoimmunity. Semin. Immunol. 2011, 23, 106–112. [Google Scholar] [CrossRef]
- Miura, M.; Hasegawa, N.; Noguchi, M.; Sugimoto, K.; Touma, M. The atypical I kappa B protein I kappa B-NS is important for Toll-like receptor-induced interleukin-10 production in B cells. Immunology 2016, 147, 453–463. [Google Scholar] [CrossRef]
- Tian, J.; Zekzer, D.; Hanssen, L.; Lu, Y.X.; Olcott, A.; Kaufman, D.L. Lipopolysaccharide-activated B cells down-regulate Th1 immunity and prevent autoimmune diabetes in nonobese diabetic mice. J. Immunol. 2001, 167, 1081–1089. [Google Scholar] [CrossRef]
- Miyazaki, D.; Kuo, C.H.; Tominaga, T.; Inoue, Y.; Ono, S.J. Regulatory Function of CpG-Activated B Cells in Late-Phase Experimental Allergic Conjunctivitis. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1626–1635. [Google Scholar] [CrossRef]
- Toubi, E.; Vadasz, Z. The Expansion of Cd25(High)Il-10(High)Foxp3(High) B Regulatory Cells Is in Association with Sle Disease Activity. Ann. Rheum. Dis. 2016, 75, A15. [Google Scholar] [CrossRef]
- Dambuza, I.M.; He, C.; Choi, J.K.; Yu, C.R.; Wang, R.X.; Mattapallil, M.J.; Wingfield, P.T.; Caspi, R.R.; Egwuagu, C.E. IL-12p35 induces expansion of IL-10 and IL-35-expressing regulatory B cells and ameliorates autoimmune disease. Nat. Commun. 2017, 8, 719. [Google Scholar] [CrossRef] [PubMed]
- Harvey, P.R.; Gordon, C. B-Cell Targeted Therapies in Systemic Lupus Erythematosus Successes and Challenges. Biodrugs 2013, 27, 85–95. [Google Scholar] [CrossRef]
- Tokunaga, M.; Saito, K.; Kawabata, D.; Imura, Y.; Fujii, T.; Nakayamada, S.; Tsujimura, S.; Nawata, M.; Iwata, S.; Azuma, T.; et al. Efficacy of rituximab (anti-CD20) for refractory systemic lupus erythematosus involving the central nervous system. Ann. Rheum. Dis. 2007, 66, 470–475. [Google Scholar]
- Almaani, S.; Rovin, B.H. B-cell therapy in lupus nephritis: An overview. Nephrol. Dial. Transplant. 2019, 34, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.Q.; Moisini, I.; Bethunaickan, R.; Sahu, R.; Akerman, M.; Eilat, D.; Lesser, M.; Davidson, A. BAFF/APRIL Inhibition Decreases Selection of Naive but Not Antigen-Induced Autoreactive B Cells in Murine Systemic Lupus Erythematosus. J. Immunol. 2011, 187, 6571–6580. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.J.; Deng, Y.X.; Feng, Y.; Long, D.; Ma, K.Y.; Wang, X.H.; Zhao, M.; Lu, L.W.; Lu, Q.J. Epigenetic regulation in B-cell maturation and its dysregulation in autoimmunity. Cell. Mol. Immunol. 2018, 15, 676–684. [Google Scholar] [CrossRef] [PubMed]
- Ramanujam, M.; Wang, X.B.; Huang, W.Q.; Schiffer, L.; Grimaldi, C.; Akkerman, A.; Diamond, B.; Madaio, M.P.; Davidson, A. Mechanism of action of transmembrane activator and calcium modulator ligand interactor-Ig in murine systemic lupus erythematosus. J. Immunol. 2004, 173, 3524–3534. [Google Scholar] [CrossRef] [PubMed]
- Dörner, T.; Kaufmann, J.; Wegener, W.A.; Teoh, N.; Goldenberg, D.M.; Burmester, G.R. Initial clinical trial of epratuzumab (humanized anti-CD22 antibody) for immunotherapy of systemic lupus erythematosus. Arthritis Res. Ther. 2008, 10, 406. [Google Scholar] [CrossRef]
- Alexander, T.; Sarfert, R.; Klotsche, J.; Kühl, A.A.; Rubbert-Roth, A.; Lorenz, H.M.; Rech, J.; Hoyer, B.F.; Cheng, Q.Y.; Waka, A.; et al. The proteasome inhibitior bortezomib depletes plasma cells and ameliorates clinical manifestations of refractory systemic lupus erythematosus. Ann. Rheum. Dis. 2015, 74, 1474–1478. [Google Scholar] [CrossRef]
- Wang, T.; Mei, Y.J.; Li, Z.J. Research Progress on Regulatory B Cells in Systemic Lupus Erythematosus. Biom. Res. Int. 2019, 2019, 7948687. [Google Scholar] [CrossRef]
- Lal, S. Primary Sjogren’s Syndrome. N. Engl. J. Med. 2018, 379, 96. [Google Scholar]
- Voulgarelis, M.; Tzioufas, A.G. Pathogenetic mechanisms in the initiation and perpetuation of Sjogren’s syndrome. Nat. Rev. Rheumatol. 2010, 6, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Nocturne, G.; Mariette, X. B cells in the pathogenesis of primary Sjogren syndrome. Nat. Rev. Rheumatol. 2018, 14, 133–145. [Google Scholar] [CrossRef]
- Miceli-Richard, C.; Wang-Renault, S.; Boudaoud, S.; Busato, F.; Lallemand, C.; Bethune, K.; Belkhir, R.; Nocturne, G.; Mariette, X.; Tost, J. Overlap between differentially methylated DNA regions in blood B lymphocytes and genetic at-risk loci in primary Sjogren’s syndrome. Ann. Rheum. Dis. 2016, 75, 933–940. [Google Scholar] [CrossRef]
- Imgenberg-Kreuz, J.; Sandling, J.K.; Almlof, J.C.; Nordlund, J.; Signer, L.; Norheim, K.B.; Omdal, R.; Ronnblom, L.; Eloranta, M.L.; Syvanen, A.C.; et al. Genome-wide DNA methylation analysis in multiple tissues in primary Sjogren’s syndrome reveals regulatory effects at interferon-induced genes. Ann. Rheum. Dis. 2016, 75, 2029–2036. [Google Scholar] [CrossRef]
- Blair, P.A.; Norena, L.Y.; Flores-Borja, F.; Rawlings, D.J.; Isenberg, D.A.; Ehrenstein, M.R.; Mauri, C. CD19(+)CD24(hi)CD38(hi) B Cells Exhibit Regulatory Capacity in Healthy Individuals but Are Functionally Impaired in Systemic Lupus Erythematosus Patients. Immunity 2010, 32, 129–140. [Google Scholar] [CrossRef]
- Lin, X.; Wang, X.; Xiao, F.; Ma, K.; Liu, L.; Wang, X.; Xu, D.; Wang, F.; Shi, X.; Liu, D.; et al. IL-10-producing regulatory B cells restrain the T follicular helper cell response in primary Sjogren’s syndrome. Cell. Mol. Immunol. 2019, 16, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Shen, P.; Roch, T.; Lampropoulou, V.; O’Connor, R.A.; Stervbo, U.; Hilgenberg, E.; Ries, S.; Dang, V.D.; Jaimes, Y.; Daridon, C.; et al. IL-35-producing B cells are critical regulators of immunity during autoimmune and infectious diseases. Nature 2014, 507, 366. [Google Scholar] [CrossRef]
- Wang, R.X.; Yu, C.R.; Dambuza, I.M.; Mahdi, R.M.; Dolinska, M.B.; Sergeev, Y.V.; Wingfield, P.T.; Kim, S.H.; Egwuagu, C.E. Interleukin-35 induces regulatory B cells that suppress autoimmune disease. Nat. Med. 2014, 20, 633–641. [Google Scholar] [CrossRef]
- Papp, G.; Gyimesi, E.; Szabo, K.; Zoeld, E.; Zeher, M. Increased IL-21 Expression Induces Granzyme B in Peripheral CD5(+) B Cells as a Potential Counter-Regulatory Effect in Primary Sjogren’s Syndrome. Mediat. Inflamm. 2016, 2016, 4328372. [Google Scholar] [CrossRef] [PubMed]
- Rubtsova, K.; Rubtsov, A.V.; Van Dyk, L.F.; Kappler, J.W.; Marrack, P. T-box transcription factor T-bet, a key player in a unique type of B-cell activation essential for effective viral clearance. Proc. Natl. Acad. Sci. USA 2013, 110, E3216–E3224. [Google Scholar] [CrossRef] [PubMed]
- Ramskold, D.; Parodis, I.; Lakshmikanth, T.; Sippl, N.; Khademi, M.; Chen, Y.; Zickert, A.; Mikes, J.; Achour, A.; Amara, K.; et al. B cell alterations during BAFF inhibition with belimumab in SLE. Ebiomedicine 2019, 40, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Knode, L.; Naradikian, M.S.; Myles, A.; Scholz, J.L.; Hao, Y.; Liu, D.Y.; Ford, M.L.; Tobias, J.W.; Cancro, M.P.; Gearhart, P.J. Age-Associated B Cells Express a Diverse Repertoire of V-H and V kappa Genes with Somatic Hypermutation. J. Immunol. 2017, 198, 1921–1927. [Google Scholar] [CrossRef] [PubMed]
- Lorenzetti, R.; Janowska, I.; Smulski, C.R.; Frede, N.; Henneberger, N.; Walter, L.; Schleyer, M.T.; Hüppe, J.M.; Staniek, J.; Salzer, U.; et al. Abatacept modulates CD80 and CD86 expression and memory formation in human B-cells. J. Autoimmun. 2019, 101, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Kanayama, N.; Cascalho, M.; Ohmori, H. Analysis of marginal zone B cell development in the mouse with limited B cell diversity: Role of the antigen receptor signals in the recruitment of B cells to the marginal zone. J. Immunol. 2005, 174, 1438–1445. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Haji, Y.; Hozumi, M.; Amari, Y.; Mizuno, Y.; Ito, T.; Kato, M.; Okada, M. Combined B-cell immunomodulation with rituximab and belimumab in severe, refractory TAFRO syndrome associated with Sjogren’s syndrome: A case report. Mod. Rheumatol. Case Rep. 2023, 7, 475–479. [Google Scholar] [CrossRef]
- Hansen, A.; Reiter, K.; Ziprian, T.; Jacobi, A.; Hoffmann, A.; Gosemann, M.; Scholze, J.; Lipsky, P.E.; Dorner, T. Dysregulation of chemokine receptor expression and function by B cells of patients with primary Sjogren’s syndrome. Arthritis Rheum. 2005, 52, 2109–2119. [Google Scholar] [CrossRef]
- Theander, E.; Jonsson, R.; Sjostrom, B.; Brokstad, K.; Olsson, P.; Henriksson, G. Prediction of Sjogren’s Syndrome Years Before Diagnosis and Identification of Patients with Early Onset and Severe Disease Course by Autoantibody Profiling. Arthritis Rheumatol. 2015, 67, 2427–2436. [Google Scholar] [CrossRef]
- Shen, L.; Gao, C.; Suresh, L.; Xian, Z.; Song, N.; Chaves, L.D.; Yu, M.; Ambrus, J.L., Jr. Central role for marginal zone B cells in an animal model of Sjogren’s syndrome. Clin. Immunol. 2016, 168, 30–36. [Google Scholar] [CrossRef]
- Youinou, P.; Pers, J. Disturbance of cytokine networks in Sjogren’s syndrome. Arthritis Res. Ther. 2011, 13, 227. [Google Scholar] [CrossRef]
- Adler, L.N.; Jiang, W.; Bhamidipati, K.; Millican, M.; Macaubas, C.; Hung, S.C.; Mellins, E.D. The Other Function: Class II-Restricted Antigen Presentation by B Cells. Front. Immunol. 2017, 8, 319. [Google Scholar] [CrossRef]
- Ma, K.; Wang, X.; Shi, X.; Lin, X.; Xiao, F.; Ma, X.; Liu, D.; Lu, L. The expanding functional diversity of plasma cells in immunity and inflammation. Cell. Mol. Immunol. 2020, 17, 421–422. [Google Scholar] [CrossRef]
- Guo, J.; Gu, M.; Zhang, W.; Liu, Y.; Qian, C.; Deng, A. Aberrant IL-35 levels in patients with primary Sjogren’s syndrome. Scand. J. Immunol. 2018, 88, e12718. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Jin, L.X.; Chen, H.; Wu, Q.J.; Fei, Y.Y.; Zheng, W.J.; Wang, Q.; Li, P.; Li, Y.Z.; Zhang, W.; et al. B cell subsets and dysfunction of regulatory B cells in IgG4-related diseases and primary Sjogren’s syndrome: The similarities and differences. Arthritis Res. Ther. 2014, 16, R118. [Google Scholar] [CrossRef] [PubMed]
- Szabo, K.; Papp, G.; Szanto, A.; Tarr, T.; Zeher, M. A comprehensive investigation on the distribution of circulating follicular T helper cells and B cell subsets in primary Sjogren’s syndrome and systemic lupus erythematosus. Clin. Exp. Immunol. 2016, 183, 76–89. [Google Scholar] [CrossRef]
- Julie, M.; Anaiz, N.; Philippe, G.; Rachel, A.; Cecile, G.V.; Bernard, C.; Jacques, M.; Claire, D. IL-10-producing regulatory B cells are present and functional in primary Sjogren patients. Immunol. Res. 2021, 69, 107–113. [Google Scholar]
- Chen, X.; Jiang, S.; Zhou, Z.; Xu, X.; Ying, S.; Du, L.; Qiu, K.; Xu, Y.; Wu, J.; Wang, X. Increased expression of interleukin-21-inducible genes in minor salivary glands are associated with primary Sjogren’s syndrome disease characteristics. Rheumatology 2021, 60, 2979–2989. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.Q.; Kim, H.; Cornelius, J.G.; Peck, A.B. Development of Sjogren’s syndrome in Nonobese diabetic-derived autoimmune-prone C57BL/6.NOD-Aec1Aec2 mice is dependent on complement component-3. J. Immunol. 2007, 179, 2318–2329. [Google Scholar] [CrossRef]
- Daridon, C.; Pers, J.O.; Devauchelle, V.; Martins-Carvalho, C.; Hutin, P.; Pennec, Y.L.; Saraux, A.; Youinou, P. Identification of transitional type IIB cells in the salivary glands of patients with Sjogren’s syndrome. Arthritis Rheum. 2006, 54, 2280–2288. [Google Scholar] [CrossRef] [PubMed]
- Burton, A.R.; Guillaume, S.M.; Foster, W.S.; Wheatley, A.K.; Hill, D.L.; Carr, E.J.; Linterman, M.A. The memory B cell response to influenza vaccination is impaired in older persons. Cell Rep. 2022, 41, 111613. [Google Scholar] [CrossRef] [PubMed]
- Pierpont, T.M.; Limper, C.B.; Richards, K.L. Past, Present, and Future of Rituximab-The world’s First Oncology Monoclonal Antibody Therapy. Front. Oncol. 2018, 8, 163. [Google Scholar] [CrossRef] [PubMed]
- Sacco, K.A.; Abraham, R.S. Consequences of B-cell-depleting therapy: Hypogammaglobulinemia and impaired B-cell reconstitution. Immunotherapy 2018, 10, 713–728. [Google Scholar] [CrossRef] [PubMed]
- Dörner, T.; Posch, M.G.; Li, Y.; Petricoul, O.; Cabanski, M.; Milojevic, J.M.; Kamphausen, E.; Valentin, M.A.; Simonett, C.; Mooney, L.; et al. Treatment of primary Sjogren’s syndrome with ianalumab (VAY736) targeting B cells by BAFF receptor blockade coupled with enhanced, antibody-dependent cellular cytotoxicity. Ann. Rheum. Dis. 2019, 78, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Fasano, S.; Isenberg, D.A. Present and novel biologic drugs in primary Sjogren’s syndrome. Clin. Exp. Rheumatol. 2019, 37, S167–S174. [Google Scholar]
- Fisher, B.A.; Szanto, A.; Ng, W.F.; Bombardieri, M.; Posch, M.G.; Papas, A.S.; Farag, A.M.; Daikeler, T.; Bannert, B.; Kyburz, D.; et al. Assessment of the anti-CD40 antibody iscalimab in patients with primary Sjogren’s syndrome: A multicentre, randomised, double-blind, placebo-controlled, proof-of-concept study. Lancet Rheumatol. 2020, 2, E142–E152. [Google Scholar] [CrossRef] [PubMed]
- Mavragani, C.P.; Moutsopoulos, H.M. Sjogren’s syndrome: Old and new therapeutic targets. J. Autoimmun. 2020, 110, 102364. [Google Scholar] [CrossRef]
- Wang, S.P.; Iwata, S.; Nakayamada, S.; Niiro, H.; Jabbarzadeh-Tabrizi, S.; Kondo, M.; Kubo, S.; Yoshikawa, M.; Tanaka, Y. Amplification of IL-21 signalling pathway through Bruton’s tyrosine kinase in human B cell activation. Rheumatology 2015, 54, 1488–1497. [Google Scholar] [CrossRef]
- Corneth, O.; Verstappen, G.; Paulissen, S.; de Bruijn, M.; Rip, J.; Lukkes, M.; van Hamburg, J.P.; Lubberts, E.; Bootsma, H.; Kroese, F.; et al. Enhanced Bruton’s Tyrosine Kinase Activity in Peripheral Blood B Lymphocytes from Patients with Autoimmune Disease. Arthritis Rheumatol. 2017, 69, 1313–1324. [Google Scholar]
- Scott, D.L.; Wolfe, F.; Huizinga, T.W.J. Rheumatoid arthritis. Lancet 2010, 376, 1094–1108. [Google Scholar] [CrossRef]
- Smolen, J.S.; Aletaha, D.; Barton, A.; Burmester, G.R.; Emery, P.; Firestein, G.S.; Kavanaugh, A.; McInnes, I.B.; Solomon, D.H.; Strand, V.; et al. Rheumatoid arthritis. Nat. Rev. Dis. Primers 2018, 4, 18001. [Google Scholar] [CrossRef]
- Ummarino, D. Defective IL-10-producing Breg cells. Nat. Rev. Rheumatol. 2017, 13, 132. [Google Scholar] [CrossRef]
- Flores-Borja, F.; Bosma, A.; Ng, D.; Reddy, V.; Ehrenstein, M.R.; Isenberg, D.A.; Mauri, C. CD19(+)CD24(hi)CD38(hi) B Cells Maintain Regulatory T Cells While Limiting T(H)1 and T(H)17 Differentiation. Sci. Transl. Med. 2013, 5, 173ra23. [Google Scholar] [CrossRef]
- Daien, C.I.; Gailhac, S.; Mura, T.; Combe, B.; Hahne, M.; Morel, J. Regulatory B10 CELLS ARE Decreased in Patients With Rheumatoid Arthritis and Inversely Correlated With Disease Activity. Arthritis Rheum. 2013, 65, S592. [Google Scholar] [CrossRef]
- Cui, D.H.; Zhang, L.L.; Chen, J.X.; Zhu, M.; Hou, L.; Chen, B.G.; Shen, B. Changes in regulatory B cells and their relationship with rheumatoid arthritis disease activity. Clin. Exp. Med. 2015, 15, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.Y.; Zhang, X.M.; Qin, M.T.; Wang, X.F. Changes in peripheral CD19(+)Foxp3(+) and CD19(+)TGF beta(+) regulatory B cell populations in rheumatoid arthritis patients with interstitial lung disease. J. Thorac. Dis. 2015, 7, 471–477. [Google Scholar] [PubMed]
- Salomon, S.; Guignant, C.; Morel, P.; Flahaut, G.; Brault, C.; Gourguechon, C.; Fardellone, P.; Marolleau, J.P.; Gubler, B.; Goeb, V. Th17 and CD24(hi)CD27(+) regulatory B lymphocytes are biomarkers of response to biologics in rheumatoid arthritis. Arthritis Res. Ther. 2017, 19, 33. [Google Scholar] [CrossRef] [PubMed]
- Manni, M.; Gupta, S.; Ricker, E.; Chinenov, Y.; Park, S.H.; Shi, M.; Pannellini, T.; Jessberger, R.; Ivashkiv, L.B.; Pernis, A.B. Regulation of age-associated B cells by IRF5 in systemic autoimmunity. Nat. Immunol. 2018, 19, 407. [Google Scholar] [CrossRef] [PubMed]
- Bao, W.; Xie, M.; Ye, Y. Age-associated B cells indicate disease activity in rheumatoid arthritis. Cell. Immunol. 2022, 377, 104533. [Google Scholar] [CrossRef]
- Nakayamada, S.; Kubo, S.; Yoshikawa, M.; Miyazaki, Y.; Yunoue, N.; Iwata, S.; Miyagawa, I.; Hirata, S.; Nakano, K.; Saito, K.; et al. Differential effects of biological DMARDs on peripheral immune cell phenotypes in patients with rheumatoid arthritis. Rheumatology 2018, 57, 164–174. [Google Scholar] [CrossRef]
- Ajeganova, S.; De Faire, U.; Jogestrand, T.; Frostegard, J.; Hafstrom, I. Carotid Atherosclerosis, Disease Measures, Oxidized Low-density Lipoproteins, and Atheroprotective Natural Antibodies for Cardiovascular Disease in Early Rheumatoid Arthritis—An Inception Cohort Study. J. Rheumatol. 2012, 39, 1146–1154. [Google Scholar] [CrossRef]
- Chirivi, R.G.S. Anti-Citrullinated Protein Antibodies as Novel Therapeutic Drugs in Rheumatoid Arthritis. J. Clin. Cell. Immunol. 2013, 4, 2. [Google Scholar]
- De Faire, U.; Su, J.; Hua, X.; Frostegard, A.; Halldin, M.; Hellenius, M.L.; Wikstrom, M.; Dahlbom, I.; Gronlund, H.; Frostegard, J. Low levels of IgM antibodies to phosphorylcholine predict cardiovascular disease in 60-year old men: Effects on uptake of oxidized LDL in macrophages as a potential mechanism. J. Autoimmun. 2010, 34, 73–79. [Google Scholar] [CrossRef]
- Chirivi, R.G.S.; Van Rosmalen, J.W.G.; Van Der Linden, M.; Euler, M.; Schmets, G.; Bogatkevich, G.; Kambas, K.; Hahn, J.; Braster, Q.; Soehnlein, O.; et al. Therapeutic ACPA inhibits NET formation: A potential therapy for neutrophil-mediated inflammatory diseases. Cell. Mol. Immunol. 2021, 18, 1528–1544. [Google Scholar] [CrossRef]
- Teng, Y.K.O.; Levarht, E.W.N.; Toes, R.E.M.; Huizinga, T.W.J.; Van Laar, J.M. Residual inflammation after rituximab treatment is associated with sustained synovial plasma cell infiltration and enhanced B cell repopulation. Ann. Rheum. Dis. 2009, 68, 1011–1016. [Google Scholar] [CrossRef]
- Cole, S.; Walsh, A.; Yin, X.; Wechalekar, M.D.; Smith, M.D.; Proudman, S.M.; Veale, D.J.; Fearon, U.; Pitzalis, C.; Humby, F.; et al. Integrative analysis reveals CD38 as a therapeutic target for plasma cell-rich pre-disease and established rheumatoid arthritis and systemic lupus erythematosus. Arthritis Res. Ther. 2018, 20, 85. [Google Scholar] [CrossRef]
- Bag-Ozbek, A.; Hui-Yuen, J.S. Emerging B-Cell Therapies in Systemic Lupus Erythematosus. Ther. Clin. Risk Manag. 2021, 17, 39–54. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, Y.; Yuan, Y.; Sun, J.; Liu, L.; Huang, D.; Hu, J.; Wang, M.; Li, S.; Song, W.; et al. In vitro elimination of autoreactive B cells from rheumatoid arthritis patients by universal chimeric antigen receptor T cells. Ann. Rheum. Dis. 2021, 80, 176–184. [Google Scholar] [CrossRef]
- Lopez-Santalla, M.; Fernandez-Perez, R.; Garin, M.I. Mesenchymal Stem/Stromal Cells for Rheumatoid Arthritis Treatment: An Update on Clinical Applications. Cells 2020, 9, 1852. [Google Scholar] [CrossRef]
- Cohen, S.; Tuckwell, K.; Katsumoto, T.R.; Zhao, R.; Galanter, J.; Lee, C.; Rae, J.; Toth, B.; Ramamoorthi, N.; Hackney, J.A.; et al. Fenebrutinib Versus Placebo or Adalimumab in Rheumatoid Arthritis: A Randomized, Double-Blind, PhaseIITrial. Arthritis Rheumatol. 2020, 72, 1435–1446. [Google Scholar] [CrossRef]
- Banko, Z.; Pozsgay, J.; Gati, T.; Rojkovich, B.; Ujfalussy, I.; Sarmay, G. Regulatory B cells in rheumatoid arthritis: Alterations in patients receiving anti-TNF therapy. Clin. Immunol. 2017, 184, 63–69. [Google Scholar] [CrossRef]
- Kubo, S.; Nakayamada, S.; Sakata, K.; Kitanaga, Y.; Ma, X.; Lee, S.; Ishii, A.; Yamagata, K.; Nakano, K.; Tanaka, Y. Janus Kinase inhibitor Baricitinib Modulates human innate and adaptive immune system. Front. Immunol. 2018, 9, 1852. [Google Scholar] [CrossRef]
- Noguchi, A.; Yasuda, S.; Hisada, R.; Kato, M.; Oku, K.; Bohgaki, T.; Suzuki, M.; Matsumoto, Y.; Atsumi, T. Anti-cyclic citrullinated peptide antibody titers decrease in rheumatoid arthritis patients treated with tocilizumab: A pilot study. Mod. Rheumatol. 2020, 30, 276–281. [Google Scholar] [CrossRef]
- Sadras, T.; Chan, L.N.; Xiao, G.; Müschen, M. Metabolic Gatekeepers of Pathological B Cell Activation. Annu. Rev. Pathol. 2021, 16, 323–349. [Google Scholar] [CrossRef]
- Müschen, M. Metabolic gatekeepers to safeguard against autoimmunity and oncogenic B cell transformation. Nat. Rev. Immunol. 2019, 19, 337–348. [Google Scholar] [CrossRef]
- Steri, M.; Orrù, V.; Idda, M.L.; Pitzalis, M.; Pala, M.; Zara, I.; Sidore, C.; Faà, V.; Floris, M.; Deiana, M.; et al. Overexpression of the Cytokine BAFF and Autoimmunity Risk. N. Engl. J. Med. 2017, 376, 1615–1626. [Google Scholar] [CrossRef]
- Gelelete, C.B.; Pereira, S.H.; Azevedo, A.; Thiago, L.S.; Mundim, M.; Land, M.G.P.; Costa, E.S. Overweight as a Prognostic Factor in Children with Acute Lymphoblastic Leukemia. Obesity 2011, 19, 1908–1911. [Google Scholar] [CrossRef]
- Crowson, C.S.; Matteson, E.L.; Davis, J.M.; Gabriel, S.E. Contribution of obesity to the rise in incidence of rheumatoid arthritis. Arthritis Care Res. 2013, 65, 71–77. [Google Scholar] [CrossRef]
- Kazazian, N.H.; Wang, Y.W.; Roussel-Queval, A.; Marcadet, L.; Chasson, L.; Laprie, C.; Desnues, B.; Charaix, J.; Irla, M.; Alexopoulou, L. Lupus Autoimmunity and Metabolic Parameters Are Exacerbated Upon High Fat Diet-Induced Obesity Due to TLR7 Signaling. Front. Immunol. 2019, 10, 2015. [Google Scholar] [CrossRef]
- Myers, M.G.; Leibel, R.L.; Seeley, R.J.; Schwartz, M.W. Obesity and leptin resistance: Distinguishing cause from effect. Trends Endocrinol. Metab. 2010, 21, 643–651. [Google Scholar] [CrossRef]
- Amarilyo, G.; Iikuni, N.; Liu, A.J.; Matarese, G.; La Cava, A. Leptin Enhances Availability of Apoptotic Cell-Derived Self-Antigen in Systemic Lupus Erythematosus. PLoS ONE 2014, 9, e112826. [Google Scholar] [CrossRef]
- Lam, Q.; Wang, S.J.; Ko, O.; Kincade, P.W.; Lu, L.W. Leptin signaling maintains B-cell homeostasis via induction of Bcl-2 and Cyclin D1. Proc. Natl. Acad. Sci. USA 2010, 107, 13812–13817. [Google Scholar] [CrossRef]
- Lourenço, E.V.; Liu, A.J.; Matarese, G.; La Cava, A. Leptin promotes systemic lupus erythematosus by increasing autoantibody production and inhibiting immune regulation. Proc. Natl. Acad. Sci. USA 2016, 113, 10637–10642. [Google Scholar] [CrossRef]
- Monzo, C.; Gkioni, L.; Beyer, A.; Valenzano, D.R.; Gronke, S.; Partridge, L. Dietary restriction mitigates the age-associated decline in mouse B cell receptor repertoire diversity. Cell Rep. 2023, 42, 112722. [Google Scholar] [CrossRef]
- Tan, J.; Ni, D.; Wali, J.A.; Cox, D.A.; Pinget, G.V.; Taitz, J.; Daïen, C.I.; Senior, A.; Read, M.N.; Simpson, S.J.; et al. Dietary carbohydrate, particularly glucose, drives B cell lymphopoiesis and function. iScience 2021, 24, 102835. [Google Scholar] [CrossRef]
- Nguyen, V.; Mendelsohn, A.; Larrick, J.W. Interleukin-7 and Immunosenescence. J. Immunol. Res. 2017, 2017, 4807853. [Google Scholar] [CrossRef]
- Lam, W.Y.; Becker, A.M.; Kennerly, K.M.; Wong, R.; Curtis, J.D.; Llufrio, E.M.; McCommis, K.S.; Fahrmann, J.; Pizzato, H.A.; Nunley, R.M.; et al. Mitochondrial Pyruvate Import Promotes Long-Term Survival of Antibody-Secreting Plasma Cells. Immunity 2016, 45, 60–73. [Google Scholar] [CrossRef] [PubMed]
- Solon-Biet, S.M.; Mitchell, S.J.; Coogan, S.; Cogger, V.C.; Gokarn, R.; McMahon, A.C.; Raubenheimer, D.; De Cabo, R.; Simpson, S.J.; Le Couteur, D.G. Dietary Protein to Carbohydrate Ratio and Caloric Restriction: Comparing Metabolic Outcomes in Mice. Cell Rep. 2015, 11, 1529–1534. [Google Scholar] [CrossRef]
- Ang, Q.Y.; Alexander, M.; Newman, J.C.; Tian, Y.; Cai, J.W.; Upadhyay, V.; Turnbaugh, J.A.; Verdin, E.; Hall, K.D.; Leibel, R.L.; et al. Ketogenic Diets Alter the Gut Microbiome Resulting in Decreased Intestinal Th17 Cells. Cell 2020, 181, 1263. [Google Scholar] [CrossRef]
- Bock, M.; Karber, M.; Kuhn, H. Ketogenic diets attenuate cyclooxygenase and lipoxygenase gene expression in multiple sclerosis. Ebiomedicine 2018, 36, 293–303. [Google Scholar] [CrossRef]
- Boothby, M.R.; Brookens, S.K.; Raybuck, A.L.; Cho, S.H. Supplying the trip to antibody production-nutrients, signaling, and the programming of cellular metabolism in the mature B lineage. Cell. Mol. Immunol. 2022, 19, 352–369. [Google Scholar] [CrossRef]
- Sancak, Y.; Bar-Peled, L.; Zoncu, R.; Markhard, A.L.; Nada, S.; Sabatini, D.M. Ragulator-Rag Complex Targets mTORC1 to the Lysosomal Surface and Is Necessary for Its Activation by Amino Acids. Cell 2010, 141, 290–303. [Google Scholar] [CrossRef]
- Jewell, J.L.; Russell, R.C.; Guan, K.L. Amino acid signalling upstream of mTOR. Nat. Rev. Mol. Cell Biol. 2013, 14, 133–139. [Google Scholar] [CrossRef]
- Takahara, T.; Amemiya, Y.; Sugiyama, R.; Maki, M.; Shibata, H. Amino acid-dependent control of mTORC1 signaling: A variety of regulatory modes. J. Biomed. Sci. 2020, 27, 87. [Google Scholar] [CrossRef]
- Yoo, H.C.; Yu, Y.C.; Sung, Y.; Han, J.M. Glutamine reliance in cell metabolism. Exp. Mol. Med. 2020, 52, 1496–1516. [Google Scholar] [CrossRef] [PubMed]
- Indrevaer, R.L.; Moskaug, J.O.; Paur, I.; Bohn, S.K.; Jorgensen, S.F.; Blomhoff, R.; Aukrust, P.; Fevang, B.; Blomhoff, H.K. IRF4 Is a Critical Gene in Retinoic Acid-Mediated Plasma Cell Formation and Is Deregulated in Common Variable Immunodeficiency-Derived B Cells. J. Immunol. 2015, 195, 2601–2611. [Google Scholar] [CrossRef] [PubMed]
- Qi, T.; Sun, M.M.; Zhang, C.; Chen, P.D.; Xiao, C.C.; Chang, X. Ascorbic Acid Promotes Plasma Cell Differentiation through Enhancing TET2/3-Mediated DNA Demethylation. Cell Rep. 2020, 33, 108452. [Google Scholar] [CrossRef] [PubMed]
- Good-Jacobson, K.L. Strength in diversity: Phenotypic, functional, and molecular heterogeneity within the memory B cell repertoire. Immunol. Rev. 2018, 284, 67–78. [Google Scholar] [CrossRef]
- Charoenngam, N.; Holick, M.F. Immunologic Effects of Vitamin D on Human Health and Disease. Nutrients 2020, 12, 2097. [Google Scholar] [CrossRef]
- Ebers, G.C.; Sadovnick, A.D.; Veith, R. Vitamin D intake and incidence of multiple sclerosis. Neurology 2004, 63, 939. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhu, X.X.; Li, C.F.; Li, Y.F.; Ye, Z.Q.; Shapiro, V.S.; Copland, J.A.; Hitosugi, T.; Bernlohr, D.A.; Sun, J.; et al. Stearoyl-CoA Desaturase-Mediated Monounsaturated Fatty Acid Availability Supports Humoral Immunity. Cell Rep. 2021, 34, 108601. [Google Scholar] [CrossRef]
- Clarke, A.J.; Riffelmacher, T.; Braas, D.; Cornall, R.J.; Simon, A.K. B1a B cells require autophagy for metabolic homeostasis and self-renewal. J. Exp. Med. 2018, 215, 399–413. [Google Scholar] [CrossRef]
- Won, W.J.; Bachmann, M.F.; Kearney, J.F. CD36 is differentially expressed on B cell subsets during development and in responses to antigen. J. Immunol. 2008, 180, 230–237. [Google Scholar] [CrossRef] [PubMed]
- He, C.F.; Wang, S.; Zhou, C.K.; He, M.H.; Wang, J.; Ladds, M.; Lianoudaki, D.; Sedimbi, S.K.; Lane, D.P.; Westerberg, L.S.; et al. CD36 and LC3B initiated autophagy in B cells regulates the humoral immune response. Autophagy 2021, 17, 3577–3591. [Google Scholar] [CrossRef]
- Weisel, F.J.; Mullett, S.J.; Elsner, R.A.; Menk, A.V.; Trivedi, N.; Luo, W.; Wikenheiser, D.; Hawse, W.F.; Chikina, M.; Smita, S.; et al. Germinal center B cells selectively oxidize fatty acids for energy while conducting minimal glycolysis. Nat. Immunol. 2020, 21, 331. [Google Scholar] [CrossRef] [PubMed]
- Vásquez, A.; Baena, A.; González, L.A.; Restrepo, M.; Muñoz, C.H.; Vanegas-García, A.; Ortiz-Reyes, B.; Abdoel, N.; Rojas, M.; García, L.F.; et al. Altered recruitment of Lyn, Syk and ZAP-70 into lipid rafts of activated B cells in Systemic Lupus Erythematosus. Cell. Signal. 2019, 58, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.N.; Gui, M.; Wang, Y.J.; Mani, N.; Chaudhuri, S.; Gao, B.X.; Li, H.B.; Kanwar, Y.S.; Lewis, S.A.; Dumas, S.N.; et al. Inositol-Requiring Enzyme 1α-Mediated Synthesis of Monounsaturated Fatty Acids as a Driver of B Cell Differentiation and Lupus-like Autoimmune Disease. Arthritis Rheumatol. 2021, 73, 2314–2326. [Google Scholar] [CrossRef]
- Sanchez, H.N.; Moroney, J.B.; Gan, H.Q.; Shen, T.; Im, J.L.; Li, T.B.; Taylor, J.R.; Zan, H.; Casali, P. B cell-intrinsic epigenetic modulation of antibody responses by dietary fiber-derived short-chain fatty acids. Nat. Commun. 2020, 11, 60. [Google Scholar] [CrossRef]
- Schäfer, A.L.; Eichhorst, A.; Hentze, C.; Kraemer, A.N.; Amend, A.; Sprenger, D.T.L.; Fluhr, C.; Finzel, S.; Daniel, C.; Salzer, U.; et al. Low dietary fiber intake links development of obesity and lupus pathogenesis. Eur. J. Immunol. 2022, 52, 95. [Google Scholar] [CrossRef]
- Navarini, L.; Afeltra, A.; Afflitto, G.G.; Margiotta, D.P.E. Polyunsaturated fatty acids: Any role in rheumatoid arthritis? Lipids Health Dis. 2017, 16, 197. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Bae, S.C.; Song, G.G. Omega-3 Polyunsaturated Fatty Acids and the Treatment of Rheumatoid Arthritis: A Meta-analysis. Arch. Med. Res. 2012, 43, 356–362. [Google Scholar] [CrossRef]
- Tanaka, Y.; Kondo, K.; Ichibori, A.; Yanai, Y.; Susuta, Y.; Inoue, S.; Takeuchi, T. Amiselimod, a sphingosine 1-phosphate receptor-1 modulator, for systemic lupus erythematosus: A multicenter, open-label exploratory study. Lupus 2020, 29, 1902–1913. [Google Scholar] [CrossRef] [PubMed]
- Hermann, V.; Batalov, A.; Smakotina, S.; Juif, P.E.; Cornelisse, P. First use of cenerimod, a selective S1P1 receptor modulator, for the treatment of SLE: A double-blind, randomised, placebo-controlled, proof-of-concept study. Lupus Sci. Med. 2019, 6, e000354. [Google Scholar] [CrossRef]
- Nakamura, M.; Matsuoka, T.; Chihara, N.; Miyake, S.; Sato, W.; Araki, M.; Okamoto, T.; Lin, Y.W.; Ogawa, M.; Murata, M.; et al. Differential effects of fingolimod on B-cell populations in multiple sclerosis. Mult. Scler. J. 2014, 20, 1371–1380. [Google Scholar] [CrossRef] [PubMed]
- Van Leuven, S.I.; Mendez-Fernandez, Y.V.; Stroes, E.S.; Tak, P.P.; Major, A.S. Statin therapy in lupus-mediated atherogenesis: Two birds with one stone? Ann. Rheum. Dis. 2011, 70, 245–248. [Google Scholar] [CrossRef]
- Yu, H.H.; Chen, P.C.; Yang, Y.H.; Wang, L.C.; Lee, J.H.; Lin, Y.T.; Chiang, B.L. Statin reduces mortality and morbidity in systemic lupus erythematosus patients with hyperlipidemia: A nationwide population-based cohort study. Atherosclerosis 2015, 243, 11–18. [Google Scholar] [CrossRef]
- Brookens, S.K.; Boothby, M.R. AMPK Metabolism in the B Lineage Modulates Humoral Responses. Immunometabolism 2021, 3, e210011. [Google Scholar] [CrossRef]
- Jones, D.D.; Gaudette, B.T.; Wilmore, J.R.; Chernova, I.; Bortnick, A.; Weiss, B.M.; Allman, D. mTOR has distinct functions in generating versus sustaining humoral immunity. J. Clin. Investig. 2016, 126, 4250–4261. [Google Scholar] [CrossRef]
- Brookens, S.K.; Cho, S.H.; Basso, P.J.; Boothby, M.R. AMPKα1 in B Cells Dampens Primary Antibody Responses yet Promotes Mitochondrial Homeostasis and Persistence of B Cell Memory. J. Immunol. 2020, 205, 3011. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Park, H.; Lim, J.; Jin, H.S.; Park, Y.; Jung, Y.J.; Ko, H.J.; Yoon, S.I.; Lee, G.S.; Kim, P.H.; et al. GSK3 Restrains Germinal Center B Cells to Form Plasma Cells. J. Immunol. 2021, 206, 481. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| B Cell Subset | Markers | Function | Potential Therapy |
|---|---|---|---|
| IL-10+ Breg | CD24, CD38, CD1d, CD5, IL-10 [99,100] | Protective | - |
| IL-35+ Breg | CD138, TACI, CXCR4, IL-35 [101,102] | Protective | - |
| GrB+ Breg | CD5, GrB [103] | Protective | - |
| Age-Associated B cell | CD11c, T-bet, CXCR5, CD21, CD23 [104] | Pathogenic | Belimumab [105], Iscalimab [106], Abatacept [107] |
| Marginal Zone B cell | CD21, CD23, IgD [108] | Pathogenic | Rituximab [109] |
| Memory B cell | CD27, CXCR4, CXCR5 [110] | Pathogenic | - |
| Types of Nutrients | Effects of Nutrients on the Number and Function of B Cells |
|---|---|
| Carbohydrate |
|
| Amino acids |
|
| Vitamin |
|
| Lipid |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Y.; Lu, C.; Yu, W.; Lei, Y.; Sun, S.; Liu, P.; Bai, F.; Chen, Y.; Chen, J. B Cells Dynamic in Aging and the Implications of Nutritional Regulation. Nutrients 2024, 16, 487. https://doi.org/10.3390/nu16040487
Yu Y, Lu C, Yu W, Lei Y, Sun S, Liu P, Bai F, Chen Y, Chen J. B Cells Dynamic in Aging and the Implications of Nutritional Regulation. Nutrients. 2024; 16(4):487. https://doi.org/10.3390/nu16040487
Chicago/Turabian StyleYu, Yifei, Chenxu Lu, Weiru Yu, Yumei Lei, Siyuan Sun, Ping Liu, Feirong Bai, Yu Chen, and Juan Chen. 2024. "B Cells Dynamic in Aging and the Implications of Nutritional Regulation" Nutrients 16, no. 4: 487. https://doi.org/10.3390/nu16040487
APA StyleYu, Y., Lu, C., Yu, W., Lei, Y., Sun, S., Liu, P., Bai, F., Chen, Y., & Chen, J. (2024). B Cells Dynamic in Aging and the Implications of Nutritional Regulation. Nutrients, 16(4), 487. https://doi.org/10.3390/nu16040487