
Efficacy of Laurus nobilis L. for Tight Junction Protein Imbalance in Leaky Gut Syndrome


Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Extracts from Laurus nobilis L. (LNL)
2.2. In Vitro Experiments (Caco-2)
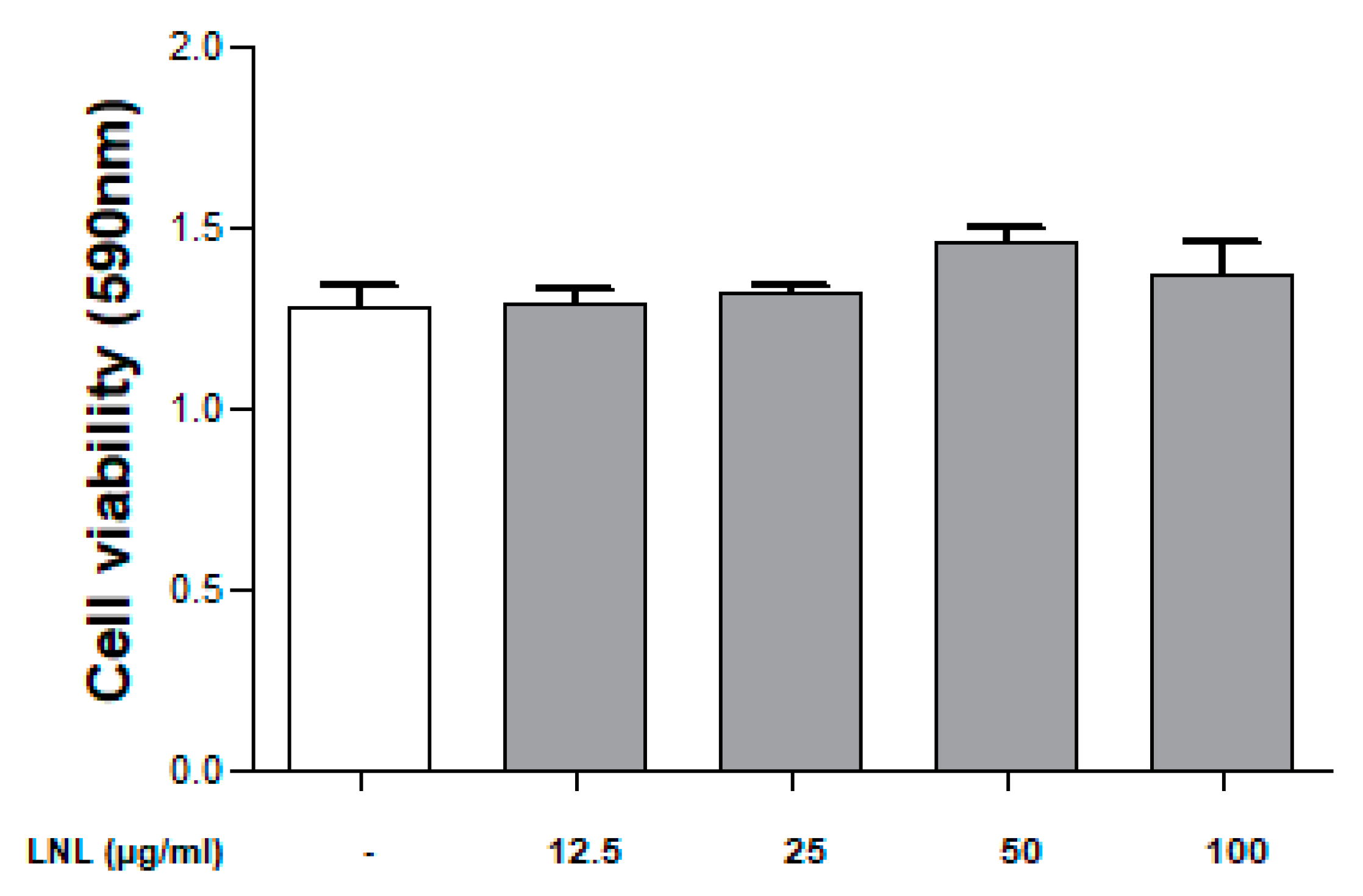
2.3. Cell Viability (MTT Assay)
2.4. Trans Epithelial Electrical Resistance (TEER) Measurements
2.5. Real-Time PCR (qPCR)
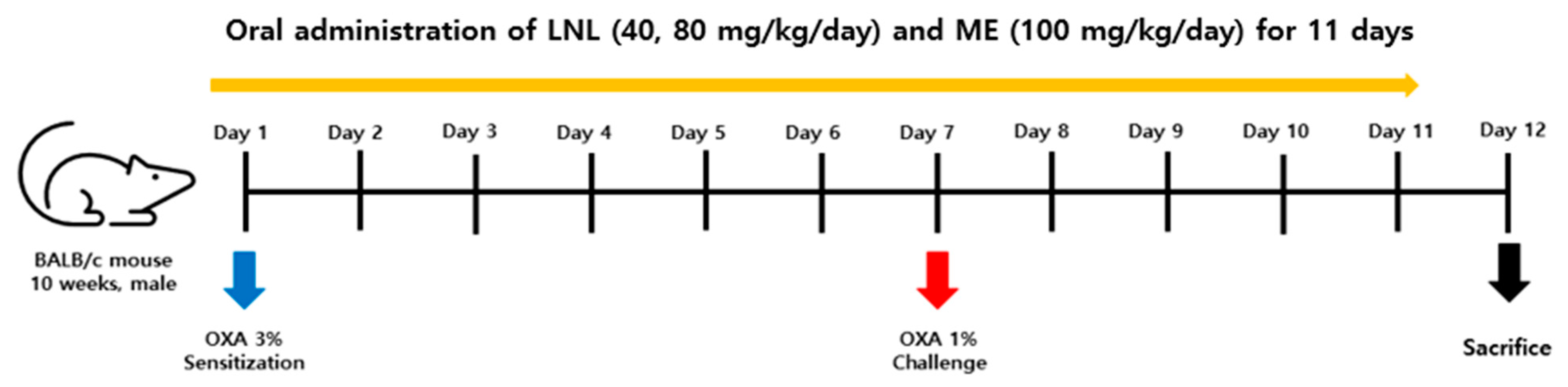
2.6. In Vivo Experiments
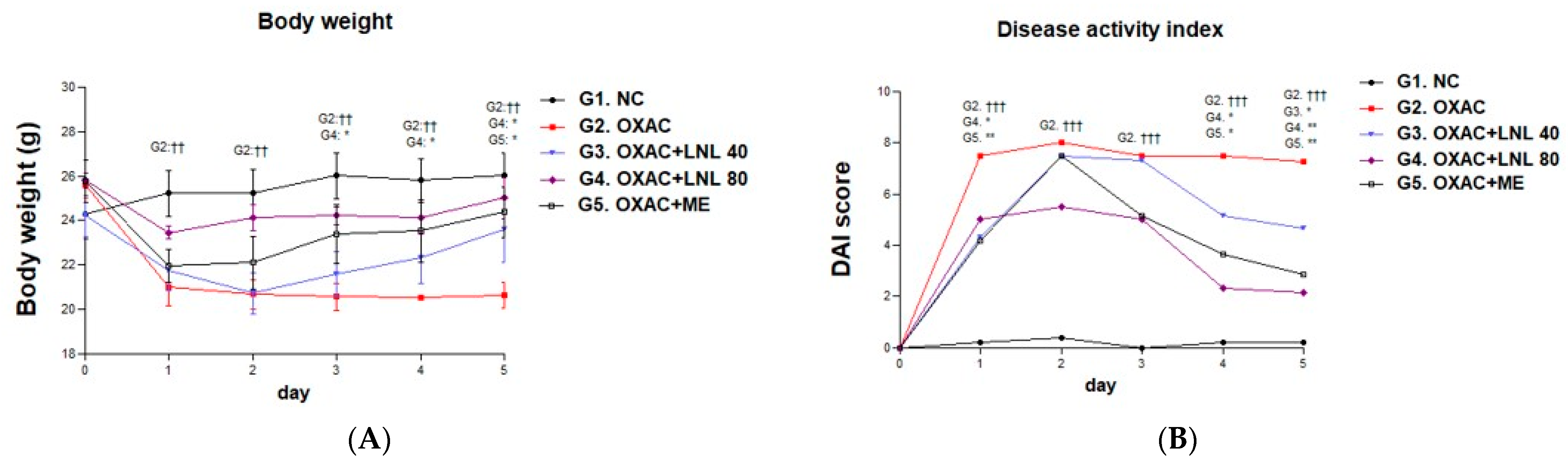
2.7. Disease Activity Index (DAI)
2.8. IL-5, IL-13, and MPO Enzyme-Linked Immunosorbent Assay (ELISA)
2.9. Western Blotting
2.10. Immunohistochemistry (IHC) Staining
2.11. Statistical Analysis
3. Results
3.1. In Vitro Experiments
3.1.1. Cell Viability
3.1.2. Efficacy of LNL on TEER in Caco-2 Cells Treated with IL-13
3.1.3. Effect of LNL on TJs Expression Mechanism in Caco-2 Cells
3.2. In Vivo Experiments
3.2.1. Effects of LNL on Body Weight Change and DAI in the OXAC Model
3.2.2. Effects of LNL on IL-5, IL-13, and MPO Secreted from the Large Intestine of OXAC Model
3.2.3. Effect of LNL on STAT6 Activation in Mouse Colon Tissue
3.2.4. Effect of LNL on Claudin-2 Expression in Mouse Colon Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Santhiravel, S.; Bekhit, A.E.A.; Mendis, E.; Jacobs, J.L.; Dunshea, F.R.; Rajapakse, N.; Ponnampalam, E.N. The impact of plant phytochemicals on the gut microbiota of humans for a balanced life. Int. J. Mol. Sci. 2022, 23, 8124. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Qiu, Y.; Yang, H. Intestinal intraepithelial lymphocytes: Maintainers of intestinal immune tolerance and regulators of intestinal immunity. J. Leukoc. Biol. 2021, 109, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.H.; Hasnain, S.Z.; Florin, T.H.; McGuckin, M.A. Mucins in inflammatory bowel diseases and colorectal cancer. J. Gastrol. Hepatol. 2012, 27, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Wlodarska, M.; Finlay, B.B. Host immune response to antibiotic perturbation of the microbiota. Mucosal Immunol. 2010, 3, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Otani, T.; Furuse, M. Tight junction structure and function revisited. Trends Cell Biol. 2020, 30, 805–817. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.-M.; Li, X.-M.; Qin, B.-Z.; Liu, B. Effect of tight junction protein of intestinal epithelium and permeability of colonic mucosa in pathogenesis of injured colonic barrier during chronic recovery stage of rats with inflammatory bowel disease. Asian Pac. J. Trop. Med. 2016, 9, 148–152. [Google Scholar] [CrossRef]
- Liang, G.H.; Weber, C.R. Molecular aspects of tight junction barrier function. Curr. Opin. Pharmacol. 2014, 19, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Soderholm, A.T.; Pedicord, V.A. Intestinal epithelial cells: At the interface of the microbiota and mucosal immunity. Immunology 2019, 158, 267–280. [Google Scholar] [CrossRef] [PubMed]
- Hoshiko, H.; Feskens, E.J.M.; Oosterink, E.; Ariens, R.M.C.; Mes, J.J.; de Wit, N.J.W. Identification of leaky gut-related markers as indicators of metabolic health in Dutch adults: The Nutrition Questionnaires plus (NQplus) study. PLoS ONE 2021, 16, e0252936. [Google Scholar] [CrossRef]
- Sheth, P.; Delos Santos, N.; Seth, A.; LaRusso, N.F.; Rao, R.K. Lipopolysaccharide disrupts tight junctions in cholangiocyte monolayers by a c-Src-, TLR4−, and LBP-dependent mechanism. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 293, G308–G318. [Google Scholar] [CrossRef]
- Capaldo, C.T.; Nusrat, A. Cytokine regulation of tight junctions. Biochim. Biophys. Acta 2009, 1788, 864–871. [Google Scholar] [CrossRef] [PubMed]
- de Kort, S.; Keszthelyi, D.; Masclee, A.A. Leaky gut and diabetes mellitus: What is the link? Obes. Rev. 2011, 12, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A. Leaky gut and autoimmune diseases. Clin. Rev. Allergy Immunol. 2021, 42, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Paray, B.A.; Albeshr, M.F.; Jan, A.T.; Rather, I.A. Leaky gut and autoimmunity: An intricate balance in individuals health and the diseased state. Int. J. Mol. Sci. 2020, 21, 9770. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Nagpal, R.; Wang, B. A human-origin probiotic cocktail ameliorates aging-related leaky gut and inflammation via modulating the microbiota/taurine/tight junction axis. JCI Insight 2020, 5, e132055. [Google Scholar] [CrossRef]
- Doron, S.; Snydman, D.R. Risk and safety of probiotics. Clin. Infect. Dis. 2015, 60, S129–S134. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Kuo, W.; Chang, K.; Yu, J.; Tsai, R. Effect of probiotics therapy on intestinal leakage: A meta-analysis of randomized controlled trials. Tungs Med. J. 2023, 17, 23–29. [Google Scholar] [CrossRef]
- He, L.; Wen, S.; Zhong, Z.; Wenig, S.; Jiang, Q.; Mi, H.; Liu, F. The synergistic effects of 5-aminosalicyclic acid and vorinostat in the treatment of ulcerative colitis. Front. Pharmacol. 2021, 12, 625543. [Google Scholar] [CrossRef] [PubMed]
- Adamowicz, M.; Milkiewicz, P.; Kempinska-Podhorodecka, A. 5-Aminosalicylic Acid Inhibits the Expression of OncomiRs and Pro-inflammatory microRNAs: An In Vitro Study. J. Physiol. Pharmacol. 2021, 72, 529–535. [Google Scholar] [CrossRef]
- Lichtenstein, G.R.; Rutgeerts, P. Importance of mucosal healing in ulcerative colitis. Inflam. Bowel Dis. 2010, 16, 338–346. [Google Scholar] [CrossRef]
- Brom, B. Integrative medicine and leaky gut syndrome. S. Afr. Fam. Pract. 2010, 52, 314–316. [Google Scholar] [CrossRef]
- Moss, A.C.; Peppercorn, M.A. The risks and the benefits of mesalazine as a treatment for ulcerative colitis. Expert. Opin. Drug Saf. 2007, 6, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, R.; Nikfar, S.; Rezaie, A.; Abdollahi, M. Comparison of Mesalazine and Balsalazide in Induction and Maintenance of Remission in Patients with Ulcerative Colitis: A Meta-Analysis. Dig. Dis. Sci. 2009, 54, 712–721. [Google Scholar] [CrossRef]
- Pica, R.; Cassieri, C.; Cocco, A.; Zippi, M.; Marcheggiano, A.; De Nitto, D.; Avallone, E.V.; Crispino, P.; Occhigrossi, G.; Paoluzi, P. A randomized trial comparing 4.8 vs. 2.4 g/day of oral mesalazine for maintenance of remission in ulcerative colitis. Dig. Liver Dis. 2015, 47, 933–937. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, A.N.; Muhammad, A.; Saliha, K.; Unal, K. Laurel (Laurus nobilis L.): A less-known medicinal plant to the world with diffusion, genomics, phenomics, and metabolomics for genetic improvement. Biotechnol. Approaches Med. Aromat. 2018, 8, 631–653. [Google Scholar] [CrossRef]
- Lee, E.H.; Shin, J.H.; Kim, S.S.; Joo, J.H.; Choi, E.; Seo, S.R. Suppression of Propionibacterium acnes-induced skin inflammation by Laurus nobilis extract and its major constituent eucalyptol. Int. J. Mol. Sci. 2019, 20, 3510. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.S.; Chi, L.W.; Jang, B.K.; Jeong, H.S.; Lee, C.H. Anti-microbial Activities of Ten Lauraceae Species against Propionibacterium acnes. Korean J. Plant Resour. 2018, 31, 423–432. [Google Scholar] [CrossRef]
- Paparella, A.; Nawade, B.; Shaltiel-Harpaz, L.; Ibdah, M. A Review of the Botany, Volatile Composition, Biochemical and Molecular Aspects, and Traditional Uses of Laurus nobilis. Plants 2022, 11, 1209. [Google Scholar] [CrossRef]
- Barla, A.; Topcu, G.; Oksuz, S.; Tumen, G. Identification of cytotoxic sesquiterpenes from Laurus nobilis L. Food Chem. 2007, 104, 1478–1484. [Google Scholar] [CrossRef]
- Guedouari, R.; Nabiev, M. Anti-inflammatory activity of different extracts from Laurus nobilis L. growing in Algeria. Alger. J. Environ. Sci. Technol. 2021, 7, 2115–2120. [Google Scholar]
- Ramos, C.; Teixeira, B.; Batista, I.; Matos, O.; Serrano, C.; Neng, N.R.; Nogueira, J.M.; Nunes, M.L.; Marques, A. Antioxidant and antibacterial activity of essential oil and extracts of bay laurel Laurus nobilis Linnaeus (Lauraceae) from Portugal. Nat. Prod. Res. 2021, 26, 518–529. [Google Scholar] [CrossRef]
- Mohammed, R.R.; Omer, A.K.; Yener, Z.; Uyar, A.; Ahmed, A.K. Biomedical effects of Laurus nobilis L. leaf extract on vital organs in streptozotocin-induced diabetic rats: Experimental research. Ann. Med. Surg. 2021, 61, 188–197. [Google Scholar] [CrossRef]
- Nam, Y.J.; Lee, D.H.; Lee, M.S.; Lee, C.S. Sesquiterpene lactone parthenolide attenuates production of inflammatory mediators by suppressing the Toll-like receptor-4-mediated activation of the Akt, mTOR, and NF-κB pathways. Naunyn-Schmiedebergs Arch. Pharmacol. 2015, 388, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo-Cantabrana, C.; Algieri, F.; Rodriguez-Nogales, A.; Vezza, T.; Martínez-Camblor, P.; Margolles, A.; Ruas-Madiedo, P.; Gálvez, J. Effect of a Ropy Exopolysaccharide-Producing Bifidobacterium animalis subsp. lactis Strain Orally Administered on DSS-Induced Colitis Mice Model. Front. Microbiol. 2016, 7, 868. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Nalle, S.C.; Shen, L.; Turner, E.S.; Singh, G.; Breskin, L.A.; Khramtsova, E.A.; Khramtsova, G.; Tsai, P.Y.; Fu, Y.X.; et al. TNFR2 activates MLCK-dependent tight junction dysregulation to cause apoptosis-mediated barrier loss and experimental colitis. Gastroenterology 2013, 145, 407–415. [Google Scholar] [CrossRef]
- Suzuki, H.; Tani, K.; Tamura, A.; Tsukita, S.; Fujiyoshi, Y. Model for the architecture of claudin-based paracellular ion channels through tight junctions. J. Mol. Biol. 2015, 427, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Kuo, W.T.; Odenwald, M.A.; Turner, J.R.; Zuo, L. Tight junction proteins occludin and ZO-1 as regulators of epithelial proliferation and survival. Ann. N. Y Acad. Sci. 2022, 1514, 21–33. [Google Scholar] [CrossRef]
- Schulzke, J.D.; Ploeger, S.; Amasheh, M.; Fromm, A.; Zeissig, S.; Troeger, H.; Richter, J.; Bojarski, C.; Schumann, M.; Fromm, M. Epithelial tight junctions in intestinal inflammation. Ann. N. Y Acad. Sci. 2009, 1165, 294–300. [Google Scholar] [CrossRef]
- Magro, F.; Cordeiro, G.; Dias, A.M.; Estevinho, M.M. Inflammatory bowel disease-Non-biological treatment. Pharmacol. Res. 2020, 160, 105075. [Google Scholar] [CrossRef]
- Cai, Z.; Wang, S.; Li, J. Treatment of Inflammatory Bowel Disease: A Comprehensive Review. Front. Med. 2021, 8, 765474. [Google Scholar] [CrossRef]
- Fuss, I.J.; Heller, F.; Boirivant, M.; Leon, F.; Yoshida, M.; Fichtner-Feigl, S.; Yang, Z.; Exley, M.; Kitani, A.; Blumberg, R.S.; et al. Nonclassical CD1d-restricted NK T cells that produce IL-13 characterize an atypical Th2 response in ulcerative colitis. J. Clin. Investig. 2004, 113, 1490–1497. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Liu, H.-Y.; Jaeson, M.I.; Deimel, L.P.; Ranasinghe, C. Unique IL-13Rα2/STAT3 mediated IL-13 regulation detected in lung conventional dendritic cells, 24 h post viral vector vaccination. Sci. Rep. 2020, 10, 1017. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Li, J.; Fu, M.; Zhao, X.; Wang, W. The JAK/STAT signaling pathway: From bench to clinic. Signal Transduct. Target. Ther. 2021, 6, 402. [Google Scholar] [CrossRef] [PubMed]
- Pickert, G.; Neufert, C.; Leppkes, M.; Zheng, Y.; Wittkopf, N.; Warntjen, M.; Lehr, H.-A.; Hirth, S.; Weigmann, B.; Wirtz, S.; et al. STAT3 links IL-22 signaling in intestinal epithelial cells to mucosal wound healing. J. Exp. Med. 2009, 206, 1465–1472. [Google Scholar] [CrossRef]
- Heller, F.; Florian, P.; Bojarski, C.; Richter, J.; Christ, M.; Hillenbrand, B.; Mankertz, J.; Gitter, A.H.; Bürgel, N.; Fromm, M.; et al. Interleukin-13 is the key effector Th2 cytokine in ulcerative colitis that affects epithelial tight junctions, apoptosis, and cell restitution. Gastroenterology 2005, 129, 550–564. [Google Scholar] [CrossRef] [PubMed]
- Domazetovic, V.; Iantomasi, T.; Bonanomi, A.G.; Stio, M. Vitamin D regulates claudin-2 and claudin-4 expressi on in active ulcerative colitis by p-Stat-6 and Smad-7 signaling. Int. J. Color. Dis. 2020, 35, 1231–1242. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Oshima, T.; Tomita, T.; Fukui, H.; Miwa, H. Butyrate Alleviates Cytokine-Induced Barrier Dysfunction by Modifying Claudin-2 Levels. Biology 2021, 10, 205. [Google Scholar] [CrossRef] [PubMed]
- Weber, C.R.; Raleigh, D.R.; Su, L.; Shen, L.; Sullivan, E.A.; Wang, Y.; Turner, J.R. Epithelial Myosin Light Chain Kinase Activation Induces Mucosal Interleukin-13 Expression to Alter Tight Junction Ion Selectivity. J. Biol. Chem. 2010, 285, 12037–12046. [Google Scholar] [CrossRef] [PubMed]
- Raju, P.; Shashikanth, N.; Tsai, P.Y.; Pongkorpsakol, P.; Chanez-Paredes, S.; Steinhagen, P.R.; Kuo, W.T.; Singh, G.; Tsukita, S.; Turner, J.R. Inactivation of paracellular cation-selective claudin-2 channels attenuates immune-mediated experimental colitis in mice. J. Clin. Investig. 2020, 130, 5197–5208. [Google Scholar] [CrossRef]
- Kim, B.E.; Leung, D.Y.; Boguniewicz, M.; Howell, M.D. Loricrin and involucrin expression is down-regulated by Th2 cytokines through STAT-6. Clin. Immunol. 2008, 126, 332–337. [Google Scholar] [CrossRef]
- Oueslati, M.; Bettaieb, I.; Ben Younes, R.; Gamoudi, A.; Rahal, K.; Oueslati, R. STAT-5 and STAT-6 in Breast Cancer: Potential Crosstalk with Estrogen and Progesterone Receptors Can Affect Cell Proliferation and Metastasis. J. Clin. Med. Res. 2022, 14, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Shi, Y.; Du, J.; Ge, X.; Teng, X.; Liu, L.; Wang, E.; Zhao, Q. Vitamin D treatment attenuates 2,4,6-trinitrobenzene sulphonic acid (TNBS)-induced colitis but not oxazolone-induced colitis. Sci. Rep. 2016, 6, 32889. [Google Scholar] [CrossRef] [PubMed]
- Rosen, M.J.; Frey, M.R.; Washington, M.K.; Chaturvedi, R.; Kuhnhein, L.A.; Matta, P.; Revetta, F.L.; Wilson, K.T.; Polk, D.B. STAT6 activation in ulcerative colitis: A new target for prevention of IL-13-induced colon epithelial cell dysfunction. Inflamm. Bowel Dis. 2011, 17, 2224–2234. [Google Scholar] [CrossRef]
- Gwiggner, M.; Martinez-Nunez, R.T.; Whiteoak, S.R.; Bondanese, V.P.; Claridge, A.; Collins, J.E.; Cummings, J.R.F.; Sanchez-Elsner, T. MicroRNA-31 and MicroRNA-155 Are Overexpressed in Ulcerative Colitis and Regulate IL-13 Signaling by Targeting Interleukin 13 Receptor α-1. Genes 2018, 9, 85. [Google Scholar] [CrossRef]
- Piliponsky, A.M.; Gleich, G.J.; Bar, I.; Levi-Schaffer, F. Effects of eosinophils on mast cells: A new pathway for the perpetuation of allergic inflammation. Mol. Immunol. 2002, 38, 1369. [Google Scholar] [CrossRef]
- Kawashima, R.; Kawamura, Y.I.; Oshio, T.; Son, A.; Yamazaki, M.; Hagiwara, T.; Okada, T.; Inagaki-Ohara, K.; Wu, P.; Szak, S.; et al. Interleukin-13 damages intestinal mucosa via TWEAK and Fn14 in mice-a pathway associated with ulcerative colitis. Gastroenterology 2011, 141, 2119–2129.e8. [Google Scholar] [CrossRef] [PubMed]
- Chami, B.; Martin, N.J.J.; Dennis, J.M.; Witting, P.K. Myeloperoxidase in the inflamed colon: A novel target for treating inflammatory bowel disease. Arch. Biochem. Biophys. 2018, 645, 61–71. [Google Scholar] [CrossRef]
- Li, M.; Ge, Q.; Du, H.; Jiang, P.; Bao, Z.; Chen, D.; Lin, S. Potential Mechanisms Mediating the Protective Effects of Tricholoma matsutake-Derived Peptides in Mitigating DSS-Induced Colitis. J. Agric. Food Chem. 2021, 69, 5536–5546. [Google Scholar] [CrossRef]
- Hohmann, M.S.N.; Longhi-Balbinot, D.T.; Guazelli, C.F.S.; Navarro, S.A.; Zarpelon, A.C.; Casagrande, R.; Arakawa, N.S.; Verri, W.A., Jr. Sesquiterpene Lactones: Structural Diversity and Perspectives as Anti-Inflammatory Molecules. Stud. Nat. Prod. Chem. 2016, 49, 243–264. [Google Scholar] [CrossRef]
- Park, E.; Song, J.H.; Kim, M.S.; Park, S.H.; Kim, T.S. Costunolide, a sesquiterpene lactone, inhibits the differentiation of pro-inflammatory CD4+ T cells through the modulation of mitogen-activated protein kinases. Int. Immunopharmacol. 2016, 40, 508–516. [Google Scholar] [CrossRef]
- Lee, B.K.; Park, S.J.; Nam, S.Y.; Kang, S.; Hwang, J.; Lee, S.J.; Im, D.S. Anti-allergic effects of sesquiterpene lactones from Saussurea costus (Falc.) Lipsch. determined using in vivo and in vitro experiments. J. Ethnopharmacol. 2018, 213, 256–261. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Sequence 5′ > 3′ | |
|---|---|---|
| Forward | Reverse | |
| claudin-1 | CCTCCTGGGAGTGATAGCAAT | GGCAACTAAAATAGCCAGACCT |
| claudin-2 | GCCTCTGGATGGAATGTGCC | GCTCCGCCACTCTGTCTTTG |
| occludin | GACTTCAGGCAGCCTCGTTAC | GCCAGTTGTGTAGTCGTGCTCA |
| ZO-1 | ACCAGTAAGTCGTCCTGATCC | TCGGCCAAATCTTCTCACTCC |
| GAPDH | CAACTTTGGCATTGTGGAAG | GGAGGAAGAGGATGCGGCAGT |
| NC | OXAC | LNL 40 | LNL 80 | ME | |
|---|---|---|---|---|---|
| Totalarea (μm2) | 1,433,933.28 | 3,065,461.29 | 2,546,376.58 | 2,375,414.46 | 2,208,251.85 |
| claudin-2area (μm2) | 24,089.25 | 95,752.78 | 66,821.68 | 35,863.96 | 33,907.18 |
| claudin-2area (%) | 1.68 | 3.12 | 2.62 | 1.51 | 1.54 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, Y.; Kim, J.; Song, Y.; Kim, S.; Kong, H. Efficacy of Laurus nobilis L. for Tight Junction Protein Imbalance in Leaky Gut Syndrome. Nutrients 2024, 16, 1250. https://doi.org/10.3390/nu16091250
Shin Y, Kim J, Song Y, Kim S, Kong H. Efficacy of Laurus nobilis L. for Tight Junction Protein Imbalance in Leaky Gut Syndrome. Nutrients. 2024; 16(9):1250. https://doi.org/10.3390/nu16091250
Chicago/Turabian StyleShin, Yelim, Jiyeon Kim, Youngcheon Song, Sangbum Kim, and Hyunseok Kong. 2024. "Efficacy of Laurus nobilis L. for Tight Junction Protein Imbalance in Leaky Gut Syndrome" Nutrients 16, no. 9: 1250. https://doi.org/10.3390/nu16091250
APA StyleShin, Y., Kim, J., Song, Y., Kim, S., & Kong, H. (2024). Efficacy of Laurus nobilis L. for Tight Junction Protein Imbalance in Leaky Gut Syndrome. Nutrients, 16(9), 1250. https://doi.org/10.3390/nu16091250