Soluble Fiber with High Water-Binding Capacity, Swelling Capacity, and Fermentability Reduces Food Intake by Promoting Satiety Rather Than Satiation in Rats

Abstract
:1. Introduction
2. Experimental Section
2.1. Animals and Diets
2.2. Behavioral Satiety Sequence
2.3. Meal Pattern Analysis
2.4. Samples Collection
2.5. Chemical Analysis
2.6. Quantitative PCR (QPCR)
2.7. Statistical Analysis
3. Results
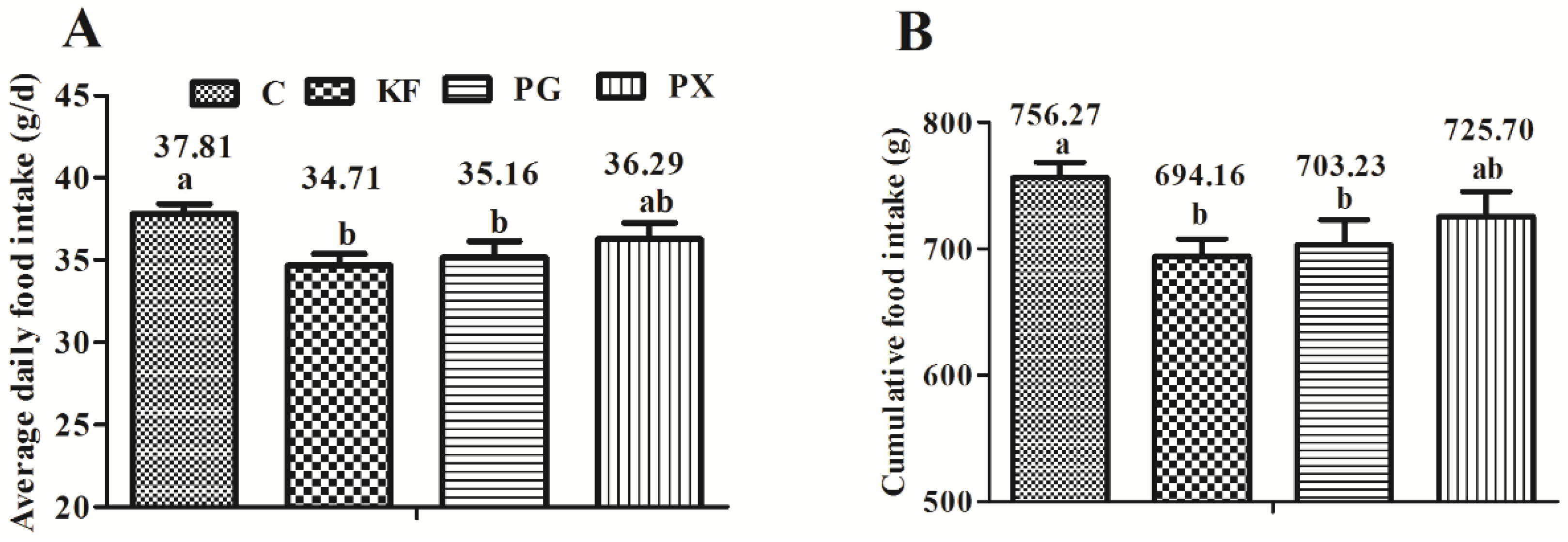
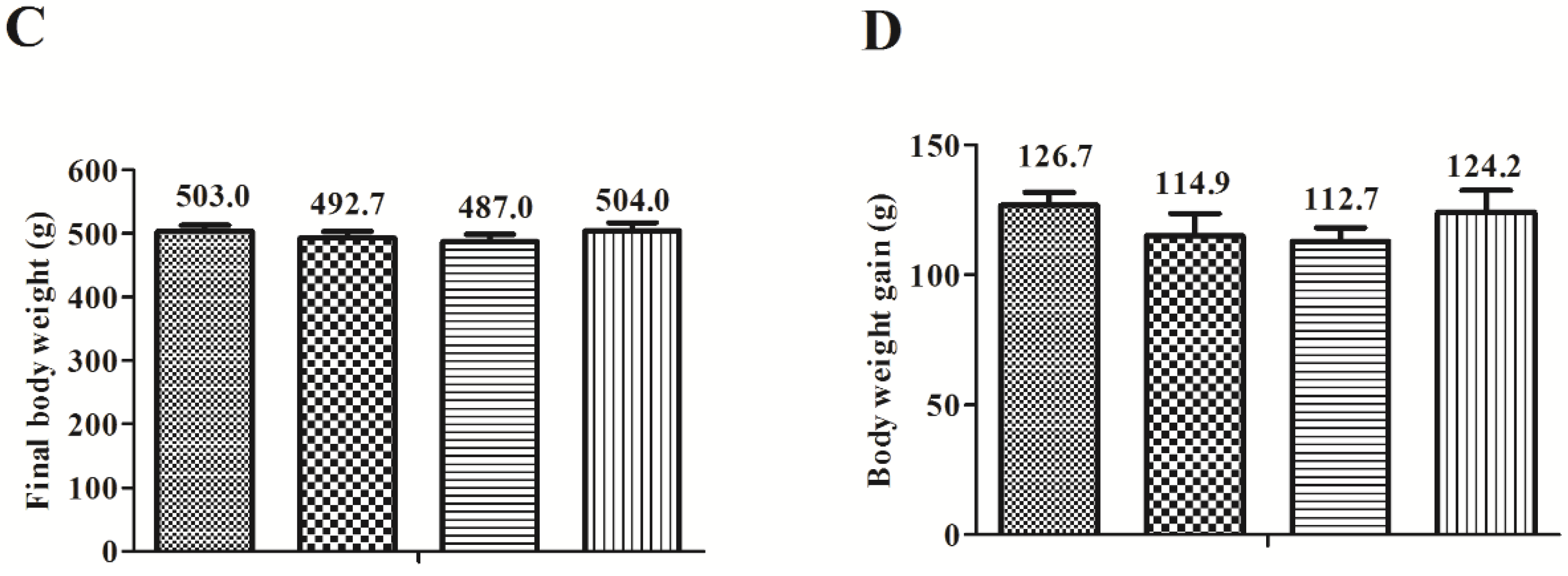
3.1. Food Intake and Body Weight
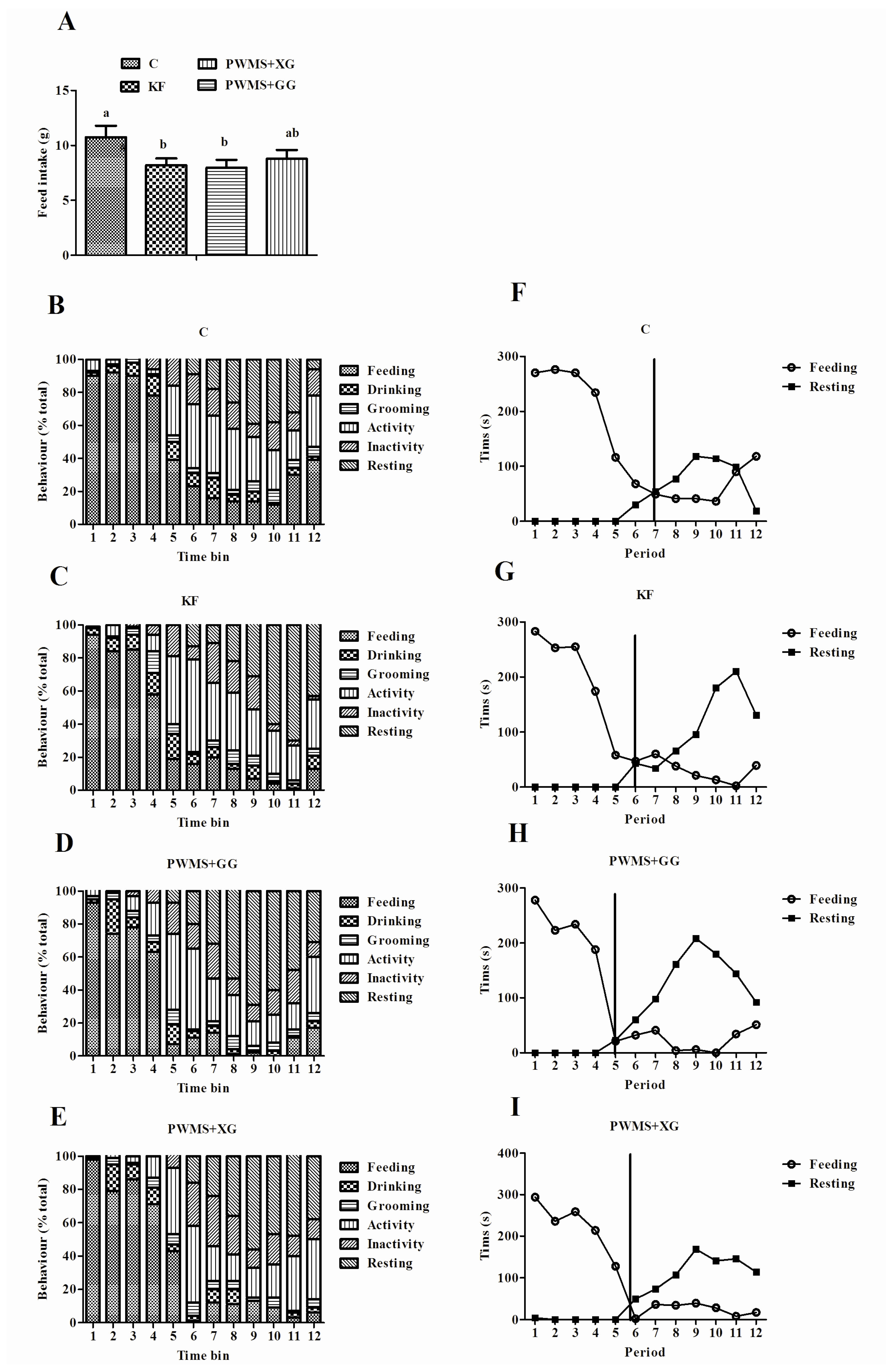
3.2. Behavioral Satiety Sequence
3.3. Meal Pattern Parameters
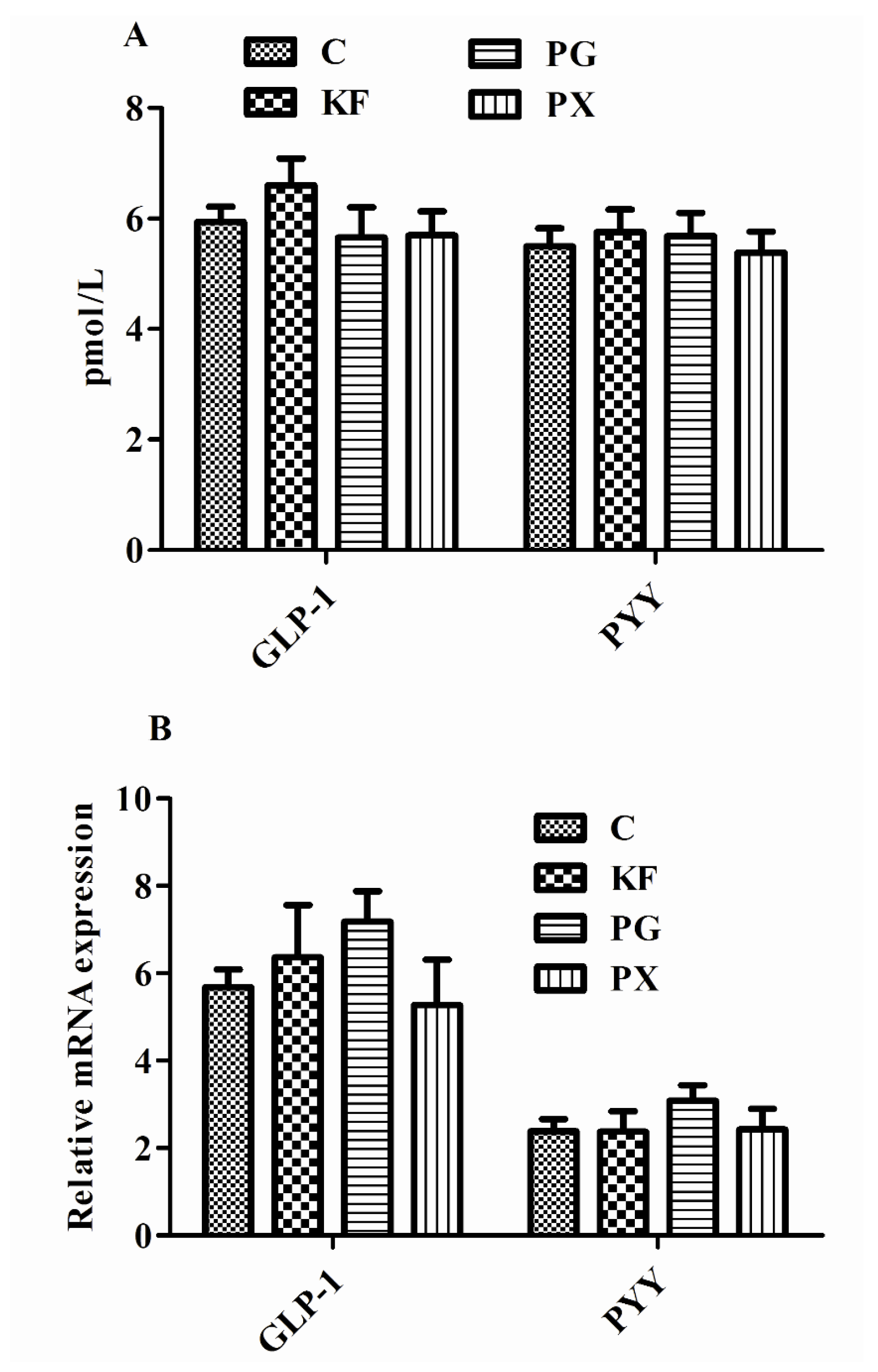
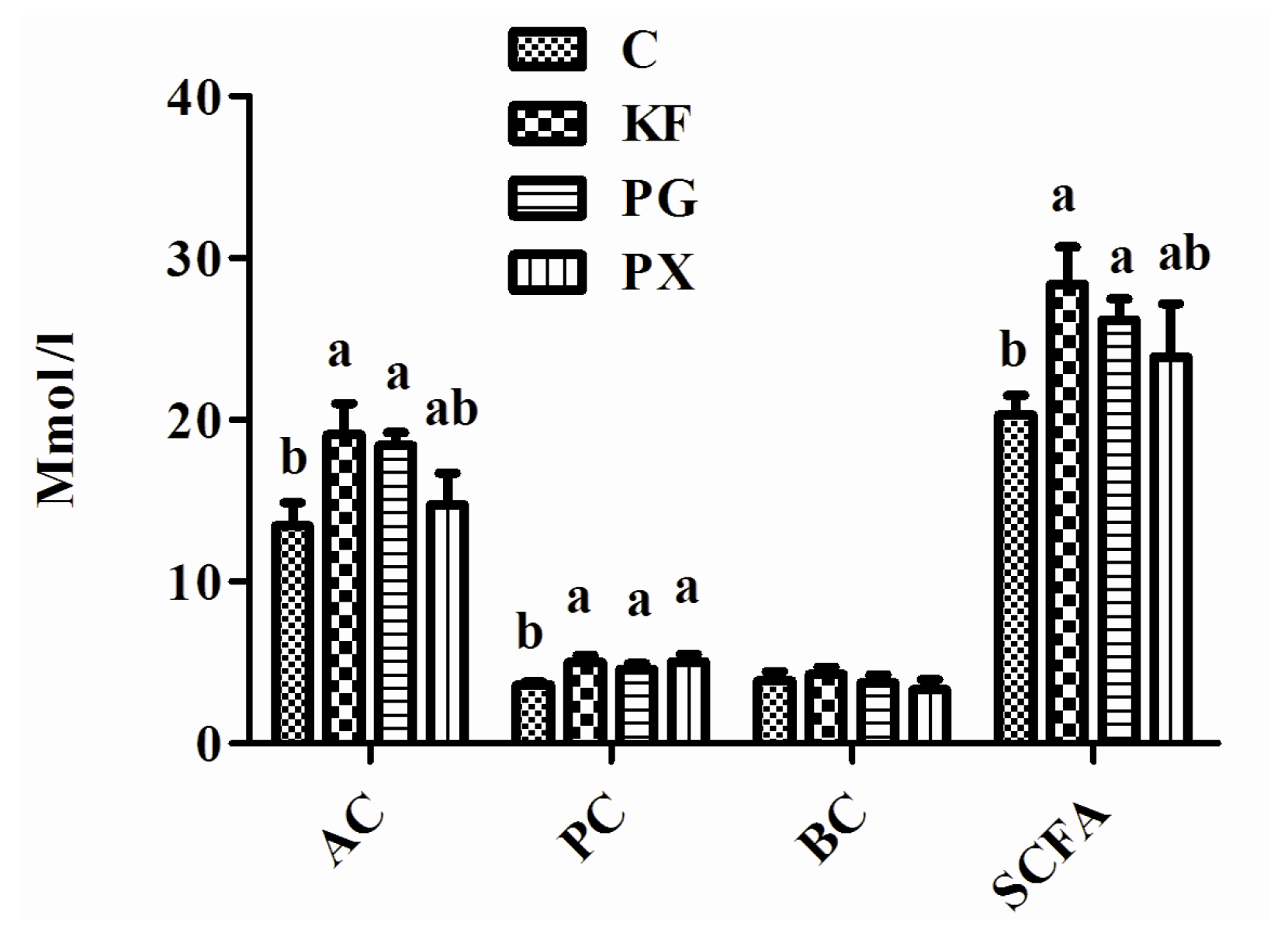
3.4. Plasma Concentrations of GLP-1/PYY, mRNA Abundance in Distal Ileum of GLP-1/PYY, and Concentrations of SCFAs in Cecal Contents
4. Discussion
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Raghad, A.; Hala, G.; Mohamad, A.; Omar, O. Meal pattern of male rats maintained on amino acid supplemented diets: The effect of tryptophan, lysine, arginine, proline and threonine. Nutrients 2014, 6, 2509–2522. [Google Scholar]
- Schroeder, N.; Marquart, L.F.; Gallaher, D.D. The role of viscosity and fermentability of dietary fibers on satiety- and adiposity-related hormones in rats. Nutrients 2013, 5, 2093–2113. [Google Scholar] [CrossRef] [PubMed]
- Wanders, A.J.; Jonathan, M.C.; van den Borne, J.J.; Mars, M.; Schols, H.A.; Feskens, E.J.; de Graaf, C. The effects of bulking, viscous and gel-forming dietary fibres on satiation. Br. J. Nutr. 2013, 109, 1330–1337. [Google Scholar] [CrossRef] [PubMed]
- Bortolotti, M.; Levorato, M.; Lugli, A.; Mazzero, G. Effect of a balanced mixture of dietary fibers on gastric emptying, intestinal transit and body weight. Ann. Nutr. Metab. 2008, 52, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Rendon-Huerta, J.A.; Juarez-Flores, B.; Pinos-Rodriguez, J.M.; Aguirre-Rivera, J.R.; Delgado-Portales, R.E. Effects of different sources of fructans on body weight, blood metabolites and fecal bacteria in normal and obese non-diabetic and diabetic rats. Plant Foods Hum. Nutr. 2012, 67, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Howarth, N.C.; Saltzman, E.; Roberts, S.B. Dietary fiber and weight regulation. Nutr. Rev. 2001, 59, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, N.; Hu, L.; Li, Z.; Li, R.; Li, C.; Wang, S. Improvement in chewing activity reduces energy intake in one meal and modulates plasma gut hormone concentrations in obese and lean young Chinese men. Am. J. Clin. Nutr. 2011, 94, 709–716. [Google Scholar] [CrossRef] [PubMed]
- Zijlstra, N.; Mars, M.; de Wijk, R.A.; Westerterp-Plantenga, M.S.; de Graaf, C. The effect of viscosity on ad libitum food intake. Int. J. Obes. 2008, 32, 676–683. [Google Scholar] [CrossRef] [PubMed]
- Dabin, B. Fibres for fertility: The inclusion of raw fibre in sow diets has many positive effects. In Feed Mix; Misset International: Doetinchem, The Netherlands, 2006; pp. 18–21. [Google Scholar]
- Bergmann, J.F.; Chassany, O.; Petit, A.; Triki, R.; Caulin, C.; Segrestaa, J.M. Correlation between echographic gastric emptying and appetite: Influence of psyllium. Gut 1992, 33, 1042–1043. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.J.; Powley, T.L. Tension and stretch receptors in gastrointestinal smooth muscle: Re-evaluating vagal mechanoreceptor electrophysiology. Brain Res. Brain Res. Rev. 2000, 34, 1–26. [Google Scholar] [CrossRef]
- Heijboer, A.C.; Pijl, H.; van den Hoek, A.M.; Havekes, L.M.; Romijn, J.A.; Corssmit, E.P.M. Gut-brain axis: Regulation of glucose metabolism. J. Neuroendocrinol. 2006, 18, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Silva, E.; Birkenhake, M.; Scholten, E.; Sagis, L.M.C.; Linden, E.V.D. Controlling rheology and structure of sweet potato starch noodles with high broccoli powder content by hydrocolloids. Food Hydrocoll. 2013, 30, 42–52. [Google Scholar] [CrossRef]
- Sun, H.Q.; Zhou, Y.F.; Tan, C.Q.; Zheng, L.F.; Peng, J.; Jiang, S.W. Effects of konjac flour inclusion in gestation diets on the nutrient digestibility, lactation feed intake and reproductive performance of sows. Animal 2014, 8, 1089–1094. [Google Scholar] [CrossRef] [PubMed]
- Katsuraya, K.; Okuyama, K.; Hatanaka, K.; Oshima, R.; Sato, T.; Matsuzaki, K. Constitution of konjac glucomannan: Chemical analysis and 13 C NMR spectroscopy. Carbohydr. Polym. 2003, 53, 183–189. [Google Scholar] [CrossRef]
- Chua, M.; Baldwin, T.C.; Hocking, T.J.; Chan, K. Traditional uses and potential health benefits of Amorphophallus konjac K. Koch ex NEBr. J. Ethnopharmacol. 2010, 128, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Shah, B.R.; Li, B.; Wang, L.; Liu, S.; Li, Y.; Wei, X.; Jin, W.; Li, Z. Health benefits of konjac glucomannan with special focus on diabetes. Bioact. Carbohydr. Diet. Fibre 2015, 5, 179–187. [Google Scholar] [CrossRef]
- Tan, C.Q.; Wei, H.K.; Sun, H.Q.; Long, G.; Ao, J.T.; Jiang, S.W.; Peng, J. Effects of supplementing sow diets during two gestations with konjac flour and Saccharomyces boulardii on constipation in peripartal period, lactation feed intake and piglet performance. Anim. Feed Sci. Technol. 2015, 210, 254–262. [Google Scholar] [CrossRef]
- Tan, C.Q.; Wei, H.K.; Ao, J.T.; Long, G.; Peng, J. Inclusion of konjac flour in the gestation diet changes the gut microbiota, alleviates oxidative stress, and improves insulin sensitivity in sows. Appl. Environ. Microbiol. 2016, 82, 5899–5909. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Tan, C.; Wei, H.; Zou, Y.; Long, G.; Ao, J.; Xue, H.; Jiang, S.; Peng, J. Effects of different amounts of konjac flour inclusion in gestation diets on physio-chemical properties of diets, postprandial satiety in pregnant sows, lactation feed intake of sows and piglet performance. Anim. Reprod. Sci. 2015, 152, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Copeland, L.; Blazek, J.; Salman, H.; Tang, M.C.M. Form and functionality of starch. Food Hydrocoll. 2009, 23, 1527–1534. [Google Scholar] [CrossRef]
- Shimotoyodome, A.; Suzuki, J.; Kameo, Y.; Hase, T. Dietary supplementation with hydroxypropyl-distarch phosphate from waxy maize starch increases resting energy expenditure by lowering the postprandial glucose-dependent insulinotropic polypeptide response in human subjects. Br. J. Nutr. 2011, 106, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Fuongfuchat, A.; Seetapan, N.; Makmoon, T.; Pongjaruwat, W.; Methacanon, P.; Gamonpilas, C. Linear and non-linear viscoelastic behaviors of crosslinked tapioca starch/polysaccharide systems. J. Food Eng. 2012, 109, 571–578. [Google Scholar] [CrossRef]
- Mandala, I.G.; Bayas, E. Xanthan effect on swelling, solubility and viscosity of wheat starch dispersions. Food Hydrocoll. 2004, 18, 191–201. [Google Scholar] [CrossRef]
- Paquet, É.; Bédard, A.; Lemieux, S.; Turgeon, S.L. Effects of apple juice-based beverages enriched with dietary fibres and xanthan gum on the glycemic response and appetite sensations in healthy men. Bioact. Carbohydr. Diet. Fibre 2014, 4, 39–47. [Google Scholar] [CrossRef]
- Tan, C.Q.; Wei, H.K.; Zhao, X.C.; Xu, C.H.; Peng, J. Effects of Water Binding Capacity and Swelling Capacity of Soluble Fibers on Physicochemical Properties of Gastrointestinal Digesta and Food Intake in Rats. Food Nutr. Res. 2015, in press. [Google Scholar]
- Glendinning, J.I.; Smith, J.C. Consistency of meal patterns in laboratory rats. Physiol. Behav. 1994, 56, 7–16. [Google Scholar] [CrossRef]
- Halford, J.C.; Wanninayake, S.C.; Blundell, J.E. Behavioral satiety sequence (BSS) for the diagnosis of drug action on food intake. Pharmacol. Biochem. Behav. 1998, 61, 159–168. [Google Scholar] [CrossRef]
- Lira, L.A.; Almeida, L.C.A.; Silva, A.A.M.D.; Cavalcante, T.C.F.; Melo, D.D.C.B.D.; Souza, J.A.D.; Campina, R.C.F.; Souza, S.L.D. Perinatal undernutrition increases meal size and neuronal activation of the nucleus of the solitary tract in response to feeding stimulation in adult rats. Int. J. Dev. Neurosci. 2014, 38, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Hadri, Z.; Chaumontet, C.; Fromentin, G.; Even, P.C.; Darcel, N.; Bouras, A.D.; Tome, D.; Rasoamanana, R. Long term ingestion of a preload containing fructo-oligosaccharide or guar gum decreases fat mass but not food intake in mice. Physiol. Behav. 2015, 147, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Association of Official Analytical Chemists. Official Methods of Analysis, 16th ed.; AOAC International: Arlington, VA, USA, 1999. [Google Scholar]
- Johansen, H.N.; Bach Knudsen, K.E.; Wood, P.J.; Fulcher, R.G. Physico-Chemical Properties and the Degradation of Oat Bran Polysaccharides in the Gut of Pigs. J. Sci. Food Agric. 1997, 73, 81–92. [Google Scholar] [CrossRef]
- Serena, A.; Jorgensen, H.; Knudsen, K.E.B. Digestion of carbohydrates and utilization of energy in sows fed diets with contrasting levels and physicochemical properties of dietary fiber. J. Anim. Sci. 2008, 86, 2208–2216. [Google Scholar] [CrossRef] [PubMed]
- Bosch, G.; Pellikaan, W.F.; Rutten, P.G.P.; van der Poel, A.F.B.; Verstegen, M.W.A.; Hendriks, W.H. Comparative in vitro fermentation activity in the canine distal gastrointestinal tract and fermentation kinetics of fiber sources. J. Anim. Sci. 2008, 86, 2979–2989. [Google Scholar] [CrossRef] [PubMed]
- Orozco-Solis, R.; Lopes-de-Souza, S.; Barbosa Matos, R.J.; Grit, I.; Le-Bloch, J.; Nguyen, P.; Manhaes-de-Castro, R.; Bolanos-Jimenez, F. Perinatal undernutrition-induced obesity is independent of the developmental programming of feeding. Physiol. Behav. 2009, 96, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Labouré, H.; Saux, S.; Nicolaidis, S. Effects of food texture change on metabolic parameters: Short- and long-term feeding patterns and body weight. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 280, R780–R789. [Google Scholar] [PubMed]
- Rasoamanana, R.; Even, P.C.; Darcel, N.; Tomé, D.; Fromentin, G. Dietary fibers reduce food intake by satiation without conditioned taste aversion in mice. Physiol. Behav. 2013, 17, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Alles, M.S.; de Roos, N.M.; Bakx, J.C.; van de Lisdonk, E.; Zock, P.L.; Hautvast, G.A. Consumption of fructooligosaccharides does not favorably affect blood glucose and serum lipid concentrations in patients with type 2 diabetes. Am. J. Clin. Nutr. 1999, 69, 64–69. [Google Scholar] [PubMed]
- Da Silva, C.S.; Bosch, G.; Bolhuis, J.E.; Stappers, L.J.N.; van Hees, H.M.J.; Gerrits, W.J.J.; Kemp, B. Effects of alginate and resistant starch on feeding patterns, behaviour and performance in ad libitum-fed growing pigs. Animal 2014, 8, 1917–1927. [Google Scholar] [CrossRef] [PubMed]
- Eidam, D.C.D.; Kuhn, K.; Stute, R. Formation of Maize Starch Gels Selectively Regulated by the Addition of Hydrocolloids. Starch Stärke 1995, 47, 378–384. [Google Scholar] [CrossRef]
- Sudhakar, V.; Singhal, R.S.; Kulkarni, P.R. Effect of salts on interactions of starch with guar gum. Food Hydrocoll. 1996, 10, 329–334. [Google Scholar] [CrossRef]
- Biliaderis, C.G.; Arvanitoyannis, I.; Izydorczyk, M.S.; Prokopowich, D.J. Effect of Hydrocolloids on Gelatinization and Structure Formation in Concentrated Waxy Maize and Wheat Starch Gels. Starch Stärke 1997, 49, 278–283. [Google Scholar] [CrossRef]
- Eerlingen, R.C.; Jacobs, H.; Delcour, J.A. Enzyme-resistant starch. V. Effect of retrogradation of waxy maize starch on enzyme susceptibility. Cereal Chem. 1994, 71, 351–355. [Google Scholar]
- Liljeberg, H.G.; Akerberg, A.K.; Björck, I.M. Effect of the glycemic index and content of indigestible carbohydrates of cereal-based breakfast meals on glucose tolerance at lunch in healthy subjects. Am. J. Clin. Nutr. 1999, 69, 647–655. [Google Scholar] [PubMed]
- Raben, A.; Tagliabue, A.; Christensen, N.J.; Madsen, J.; Holst, J.J.; Astrup, A. Resistant starch: The effect on postprandial glycemia, hormonal response, and satiety. Am. J. Clin. Nutr. 1994, 60, 544–551. [Google Scholar] [PubMed]
- Burton-Freeman, B. Dietary fiber and energy regulation. J. Nutr. 2000, 130, 272S–275S. [Google Scholar] [PubMed]
- Kristensen, M.; Jensen, M.G. Dietary fibres in the regulation of appetite and food intake. Importance of viscosity. Appetite 2011, 56, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Moran, T.H.; Ladenheim, E.E.; Schwartz, G.J. Within-meal gut feedback signaling. Int. J. Obes. Relat. Metab. Disord. 2001, 25, S39–S41. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, G. Integrative capacity of the caudal brainstem in the control of food intake. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1275–1280. [Google Scholar] [CrossRef] [PubMed]
- Blackwood, A.D.; Salter, J.; Dettmar, P.W.; Chaplin, M.F. Dietary fibre, physicochemical properties and their relationship to health. J. R. Soc. Promot. Health 2000, 120, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Slavin, J.; Savarino, V.; Paredes-Diaz, A.; Fotopoulos, G. A review of the role of soluble fiber in health with specific reference to wheat dextrin. J. Int. Med. Res. 2009, 37, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Fiszman, S.; Varela, P. The role of gums in satiety/satiation. A review. Food Hydrocoll. 2013, 32, 147–154. [Google Scholar] [CrossRef]
- Yang, H.-F.; Chen, H.-L. Utilization of Gum Dietary Fibers by the Human Fecal Inoculum in the Static Fermentation Culture System. Taiwan J. Agric. Chem. Food Sci. 2008, 46, 183–189. [Google Scholar]
- Tilg, H.; Moschen, A.R. Microbiota and diabetes: An evolving relationship. Gut 2014, 63, 1513–1521. [Google Scholar] [CrossRef] [PubMed]
- Del, P.A.; Iadevaia, M.; Loguercio, C. The role of gut hormones in controlling the food intake: What is their role in emerging diseases? Endocrinol. Nutr. 2012, 59, 197–206. [Google Scholar]
- Woodward, A.D.; Regmi, P.R.; Gänzle, M.G.; van Kempen, T.A.; Zijlstra, R.T. Slowly digestible starch influences mRNA abundance of glucose and short-chain fatty acid transporters in the porcine distal intestinal tract. J. Anim. Sci. 2012, 90, 80–82. [Google Scholar] [CrossRef] [PubMed]
- Frost, G.; Sleeth, M.L.; Sahuri-Arisoylu, M.; Lizarbe, B.; Cerdan, S.; Brody, L.; Anastasovska, J.; Ghourab, S.; Hankir, M.; Shuai, Z. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat. Commun. 2014, 5, 3611. [Google Scholar] [CrossRef] [PubMed]
- Byrne, C.S.; Chambers, E.S.; Alhabeeb, H.; Chhina, N.; Morrison, D.J.; Preston, T.; Tedford, C.; Fitzpatrick, J.; Irani, C.; Busza, A. Increased colonic propionate reduces anticipatory reward responses in the human striatum to high-energy foods123. Am. J. Clin. Nutr. 2016, 188, 724–732. [Google Scholar]
- Higgins, J.A. Resistant starch: Metabolic effects and potential health benefits. J. AOAC Int. 2004, 87, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Leeuw, J.A.D.; Jongbloed, A.W.; Spoolder, H.A.M.; Verstegen, M.W.A. Effects of hindgut fermentation of non-starch polysaccharides on the stability of blood glucose and insulin levels and physical activity in empty sows. Livest. Prod. Sci. 2005, 96, 165–174. [Google Scholar] [CrossRef]
- De Leeuw, J.A.; Jongbloed AWVerstegen, M.W. Dietary Fiber Stabilizes Blood Glucose and Insulin Levels and Reduces Physical Activity in Sows (Sus scrofa). J. Nutr. 2004, 134, 1481–1486. [Google Scholar] [PubMed]
- Sengupta, P. The Laboratory Rat: Relating Its Age With Human’s. Int. J. Prev. Med. 2013, 4, 624–630. [Google Scholar] [PubMed]
- Food and Drug Administration. Guidance for Industry: Estimating the Maximum Safe Starting Dose in Initial Clinical Trials for Therapeutics in Adult Healthy Volunteers; Food and Drug Administration: Washington, DC, USA, 2005.
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient (% w/w) | Control 1 | KF 1 | PG 1 | PX 1 |
|---|---|---|---|---|
| Corn | 52.2 | 52.2 | 52.2 | 52.2 |
| Soybean meal | 15.9 | 15.9 | 15.9 | 15.9 |
| Fish meal | 10.0 | 10.0 | 10.0 | 10.0 |
| Wheat bran | 12.0 | 10.0 | 10.0 | 10.0 |
| Sucrose | 5.0 | 5.0 | 5.0 | 5.0 |
| Fiber source | 2.0 | 2.0 | 2.0 | |
| AIN-93 Mineral mix 2 | 3.5 | 3.5 | 3.5 | 3.5 |
| AIN-93 Vitamin mix 3 | 1.0 | 1.0 | 1.0 | 1.0 |
| Composition | ||||
| Crude protein (%) | 20.04 | 19.94 | 19.78 | 19.90 |
| Energy (kcal/g) | 3.81 | 3.83 | 3.76 | 3.74 |
| Soluble fiber (%) | 1.91 | 2.89 | 3.09 | 2.57 |
| Insoluble fiber (%) | 10.46 | 10.58 | 10.61 | 11.10 |
| Viscosity (mPa/s) | 1.53 | 1.66 | 1.60 | 1.57 |
| Swelling (mL/g) | 1.81 | 2.63 | 3.03 | 2.18 |
| Water-binding capacity (g/g) | 2.05 | 2.58 | 2.87 | 2.26 |
| Item | Control 2 | KF 2 | PG 2 | PX 2 | p-Value |
|---|---|---|---|---|---|
| Behavior (% time) | |||||
| Feeding | 44.69 ± 3.45 a | 34.53 ± 3.06 b | 30.83 ± 3.19 b | 35.94 ± 2.94 a,b | 0.03 |
| Drinking | 6.04 ± 0.69 | 6.88 ± 0.70 | 5.47 ± 0.48 | 5.36 ± 0.80 | 0.38 |
| Activity | 21.15 ± 2.75 | 24.11 ± 3.54 | 21.82 ± 3.99 | 20.78 ± 2.12 | 0.88 |
| Inactivity | 10.42 ± 0.87 | 8.85 ± 1.52 | 10.73 ± 1.30 | 11.51 ± 2.01 | 0.64 |
| Grooming | 3.49 ± 0.47 | 4.53 ± 0.91 | 4.32 ± 1.09 | 4.11 ± 0.31 | 0.79 |
| Resting | 14.22 ± 4.08 | 21.09 ± 4.06 | 26.82 ± 4.28 | 22.29 ± 5.28 | 0.21 |
| Item | Control 2 | KF 2 | PG 2 | PX 2 | p-Value |
|---|---|---|---|---|---|
| Food intake (g/day) | |||||
| Total | 51.68 ± 1.50 a | 46.58 ± 0.86 b | 46.01 ± 1.07 b | 48.47 ± 0.79 b | <0.01 |
| Nocturnal | 23.40 ± 1.20 | 20.55 ± 1.04 | 20.79 ± 0.96 | 20.18 ± 1.53 | 0.24 |
| Diurnal | 28.29 ± 0.67 | 26.03 ± 1.33 | 25.22 ± 0.64 | 28.29 ± 1.62 | 0.16 |
| Feeding rate (mg/s) | |||||
| Total | 6.59 ± 0.93 | 6.54 ± 0.68 | 8.41 ± 1.29 | 5.89 ± 0.57 | 0.26 |
| Nocturnal | 5.67 ± 0.88 | 6.04 ± 0.44 | 7.09 ± 0.52 | 5.37 ± 0.62 | 0.26 |
| Diurnal | 6.59 ± 1.08 | 7.38 ± 1.25 | 8.47 ± 0.85 | 6.32 ± 0.60 | 0.45 |
| Meal size (g/day) | |||||
| Total | 3.12 ± 0.20 | 3.21 ± 0.13 | 3.39 ± 0.19 | 2.92 ± 0.19 | 0.33 |
| Nocturnal | 2.36 ± 0.14 | 2.53 ± 0.16 | 2.52 ± 0.15 | 2.20 ± 0.10 | 0.30 |
| Diurnal | 4.36 ± 0.41 | 4.22 ± 0.33 | 4.89 ± 0.47 | 3.73 ± 0.30 | 0.22 |
| Meal duration (s) | |||||
| Total | 507.45 ± 39.86 | 517.79 ± 41.60 | 437.35 ± 35.27 | 517.51 ± 41.47 | 0.43 |
| Nocturnal | 449.70 ± 38.00 | 440.70 ± 49.95 | 365.73 ± 28.00 | 437.96 ± 37.82 | 0.35 |
| Diurnal | 584.90 ± 49.69 | 620.19 ± 51.21 | 591.17 ± 40.64 | 612.64 ± 49.50 | 0.94 |
| Meal number (meals/day) | |||||
| Total | 16.88 ± 0.74 a | 14.63 ± 0.50 b | 13.88 ± 0.83 b | 17.00 ± 0.89 a | 0.01 |
| Nocturnal | 10.00 ± 0.38 a | 8.25 ± 0.41 b | 8.38 ± 0.50 b | 9.25 ± 0.70 a,b | 0.07 |
| Diurnal | 6.88 ± 0.61a b | 6.38 ± 0.50 a,b | 5.50 ± 0.53 b | 7.75 ± 0.41 a | 0.03 |
| Inter-meal interval (min) | |||||
| Total | 73.49 ± 4.57 b | 87.72 ± 4.16 a,b | 98.23 ± 9.48 a | 76.53 ± 4.03 b | 0.03 |
| Nocturnal | 58.34 ± 4.70 | 73.24 ± 6.40 | 70.90 ± 6.68 | 65.10 ± 4.74 | 0.27 |
| Diurnal | 106.65 ± 13.23 | 113.15 ± 9.48 | 133.19 ± 13.33 | 93.65 ± 5.61 | 0.09 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, C.; Wei, H.; Zhao, X.; Xu, C.; Zhou, Y.; Peng, J. Soluble Fiber with High Water-Binding Capacity, Swelling Capacity, and Fermentability Reduces Food Intake by Promoting Satiety Rather Than Satiation in Rats. Nutrients 2016, 8, 615. https://doi.org/10.3390/nu8100615
Tan C, Wei H, Zhao X, Xu C, Zhou Y, Peng J. Soluble Fiber with High Water-Binding Capacity, Swelling Capacity, and Fermentability Reduces Food Intake by Promoting Satiety Rather Than Satiation in Rats. Nutrients. 2016; 8(10):615. https://doi.org/10.3390/nu8100615
Chicago/Turabian StyleTan, Chengquan, Hongkui Wei, Xichen Zhao, Chuanhui Xu, Yuanfei Zhou, and Jian Peng. 2016. "Soluble Fiber with High Water-Binding Capacity, Swelling Capacity, and Fermentability Reduces Food Intake by Promoting Satiety Rather Than Satiation in Rats" Nutrients 8, no. 10: 615. https://doi.org/10.3390/nu8100615
APA StyleTan, C., Wei, H., Zhao, X., Xu, C., Zhou, Y., & Peng, J. (2016). Soluble Fiber with High Water-Binding Capacity, Swelling Capacity, and Fermentability Reduces Food Intake by Promoting Satiety Rather Than Satiation in Rats. Nutrients, 8(10), 615. https://doi.org/10.3390/nu8100615