N-3 Polyunsaturated Fatty Acids Decrease the Protein Expression of Soluble Epoxide Hydrolase via Oxidative Stress-Induced P38 Kinase in Rat Endothelial Cells

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Animals
2.3. Endothelium-Dependent Vasorelaxation
2.4. Cell Culture
2.5. Fatty Acid Treatments
2.6. Western Blot Analysis
2.7. mRNA Extraction and Real-Time qPCR Analysis
2.8. Quantitative Analysis of 4-HHE in Biological Samples
2.9. Quantitative Analysis of the Epoxyeicosatrienoic Acid/Dihydroxyeicosatrienoic Acid Ratio in RAECs
2.10. Statistical Analysis
3. Results
3.1. Fish Oil Treatment Improves Acetylcholine-Induced Vasodilation in Rat Aorta
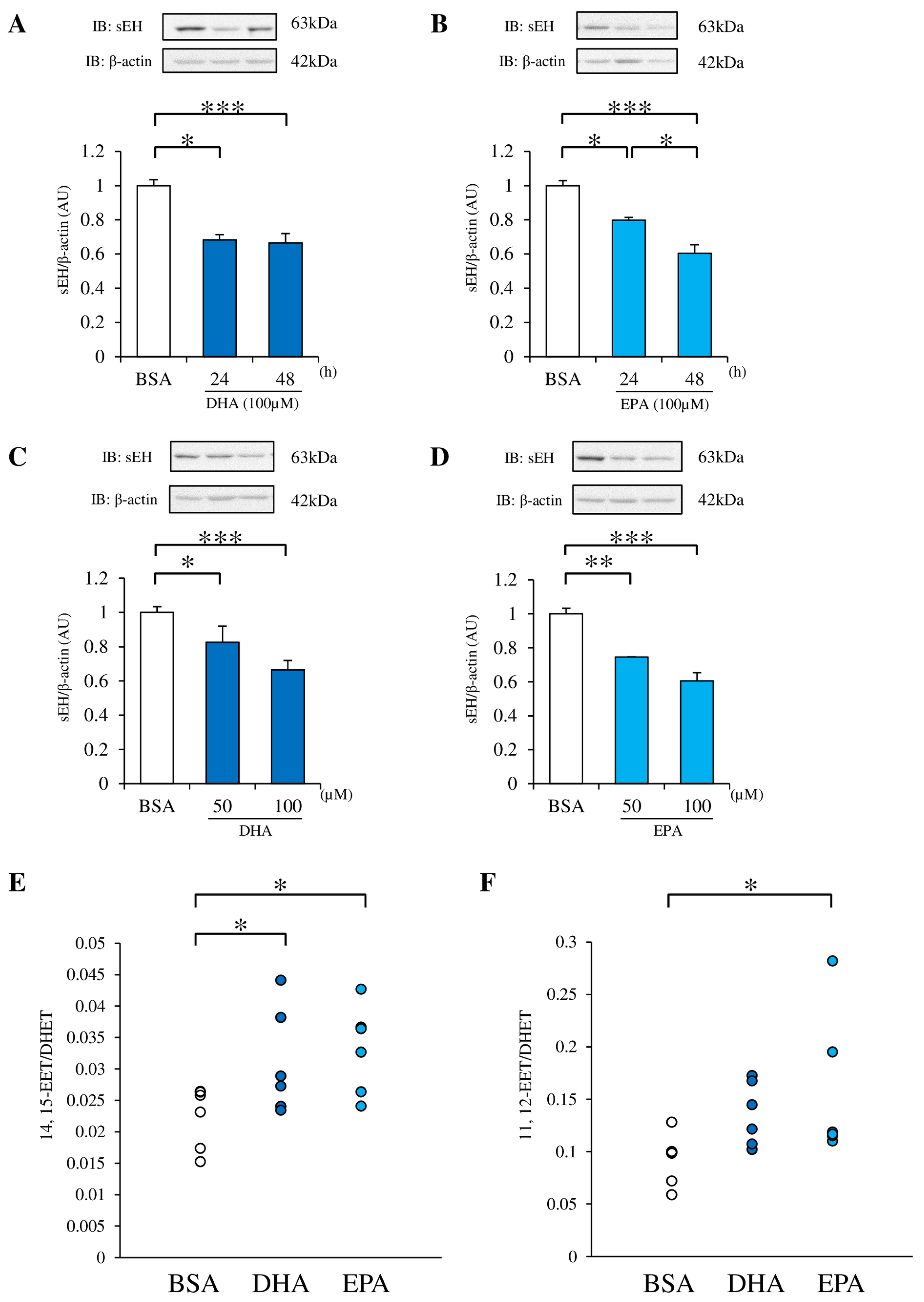
3.2. DHA and EPA Downregulate sEH Protein Expression in RAECs
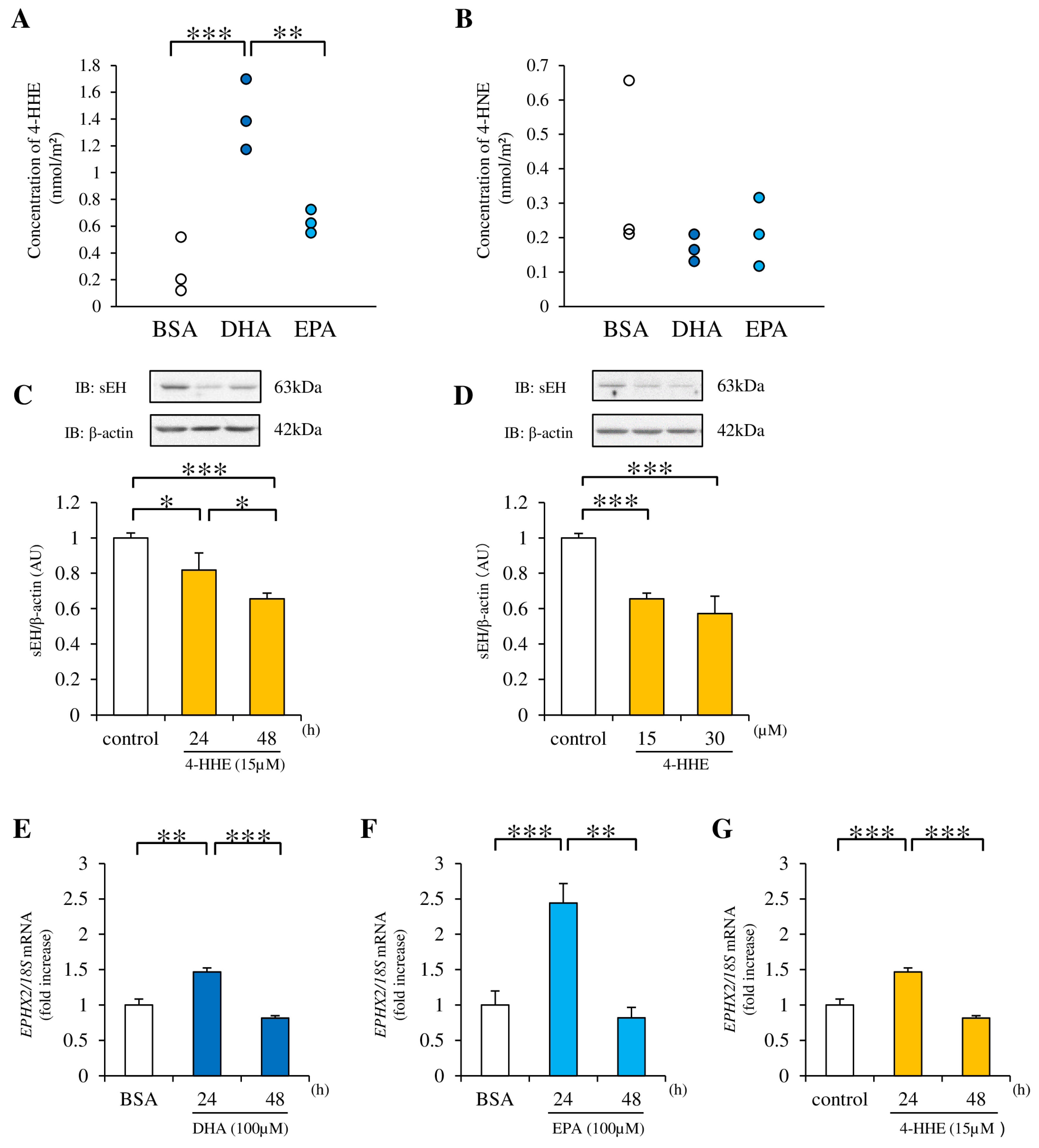
3.3. 4-HHE Downregulates sEH Protein Expression
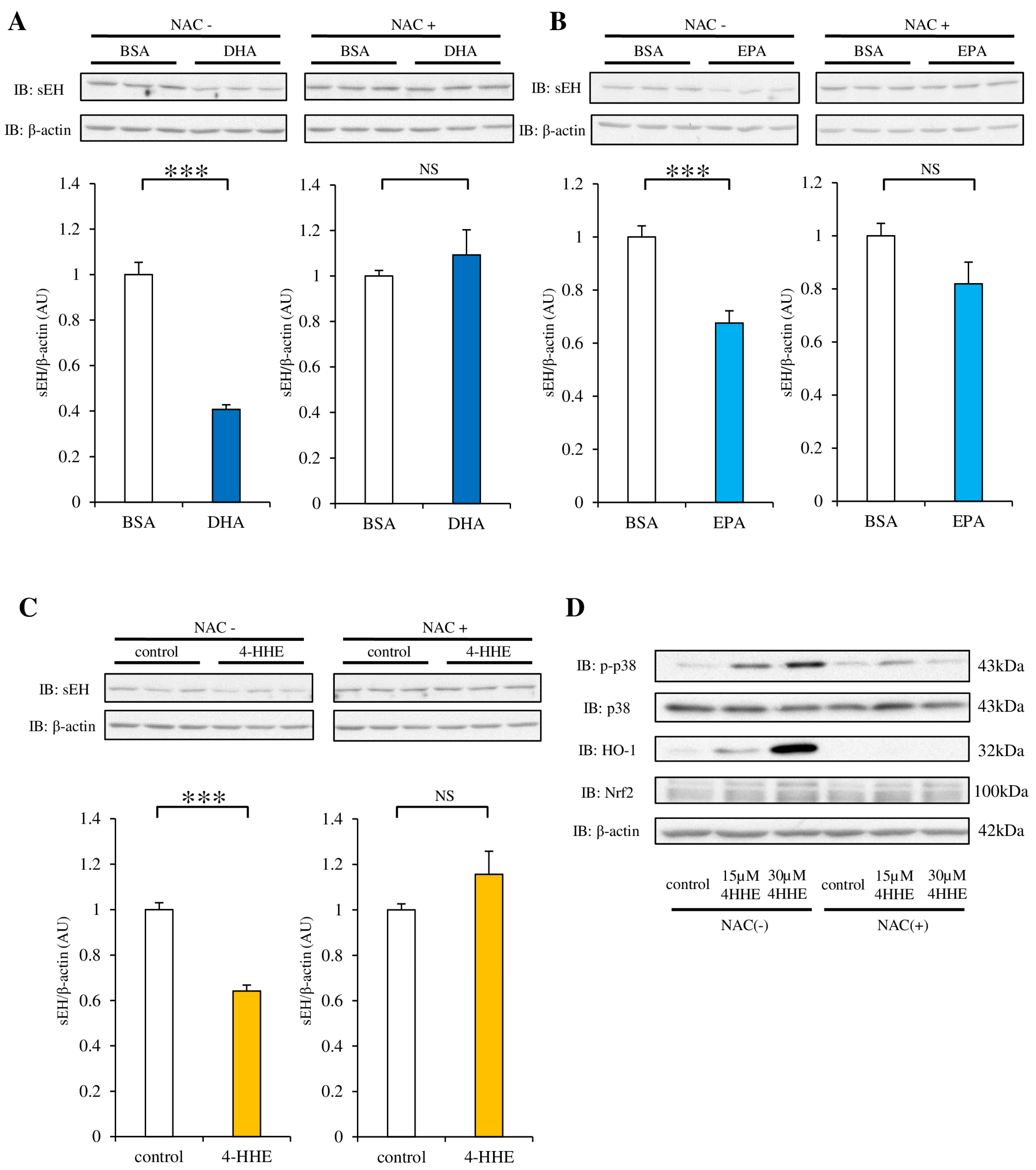
3.4. NAC Pretreatment Inhibits the Decrease in sEH Protein Induced by DHA, EPA or 4-HHE
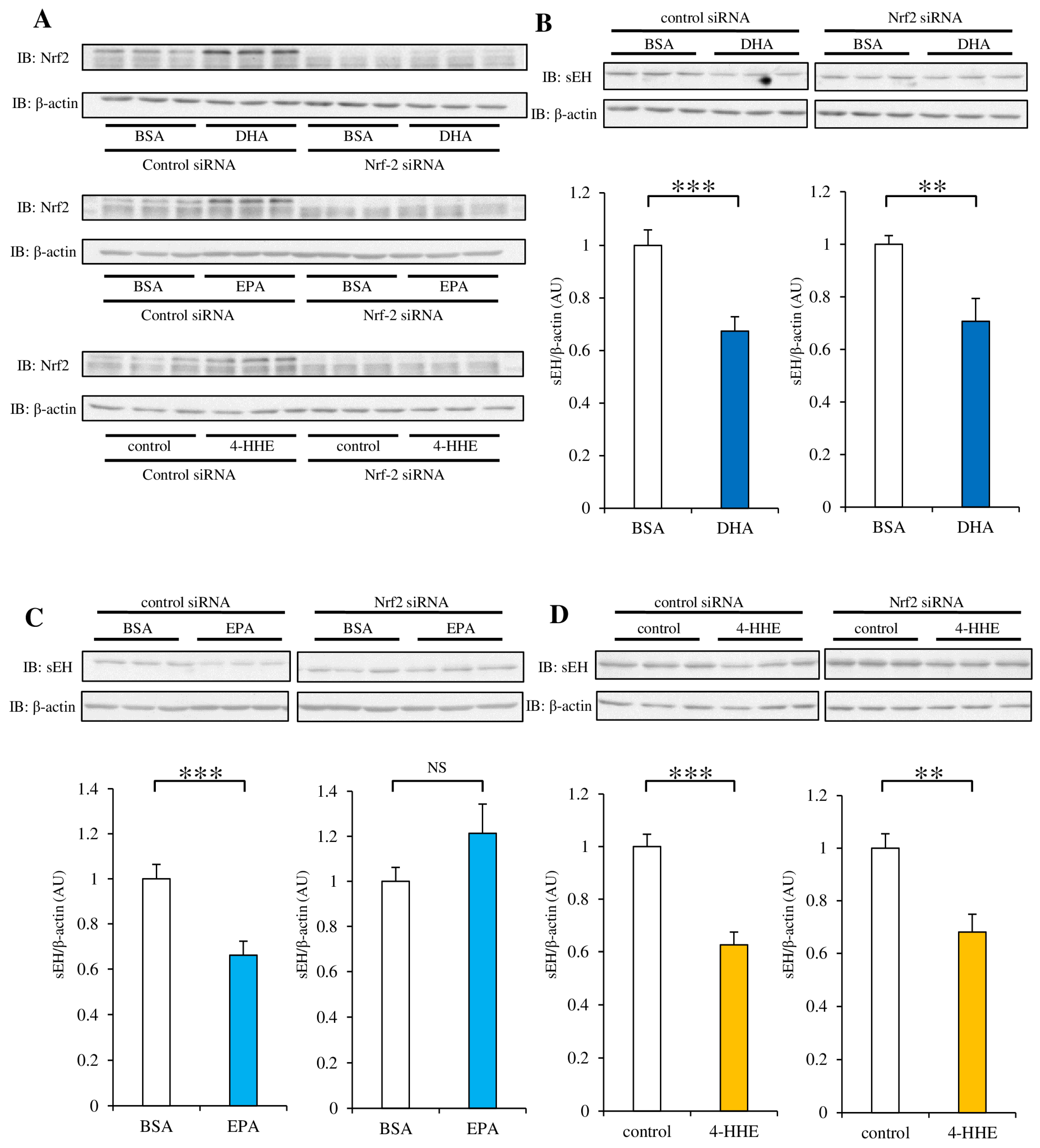
3.5. siRNA Against Nrf2 Inhibits the Decrease in sEH Protein Expression Induced by DHA, EPA, or 4-HHE
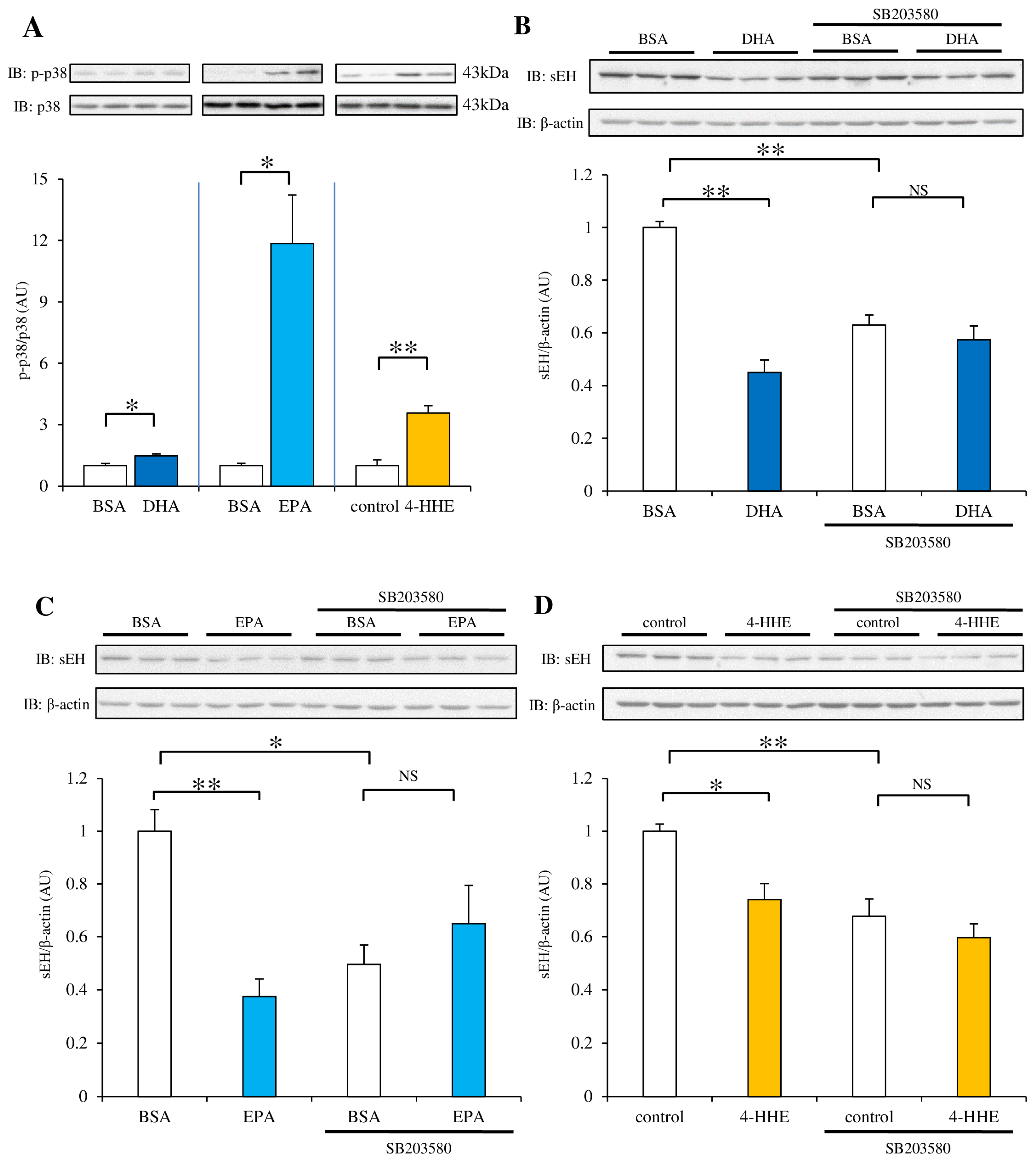
3.6. Pretreatment with the p38 Kinase Inhibitor SB203580 Rescues the Decrease in sEH Protein Expression Induced by DHA, EPA or 4-HHE
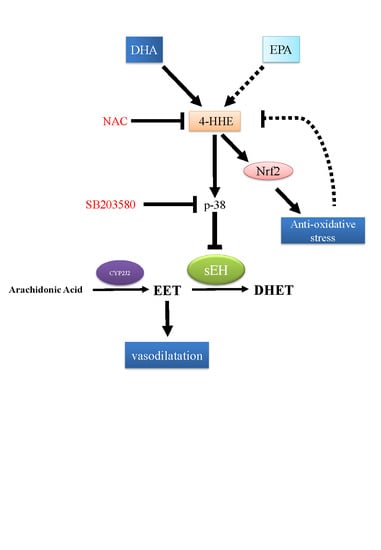
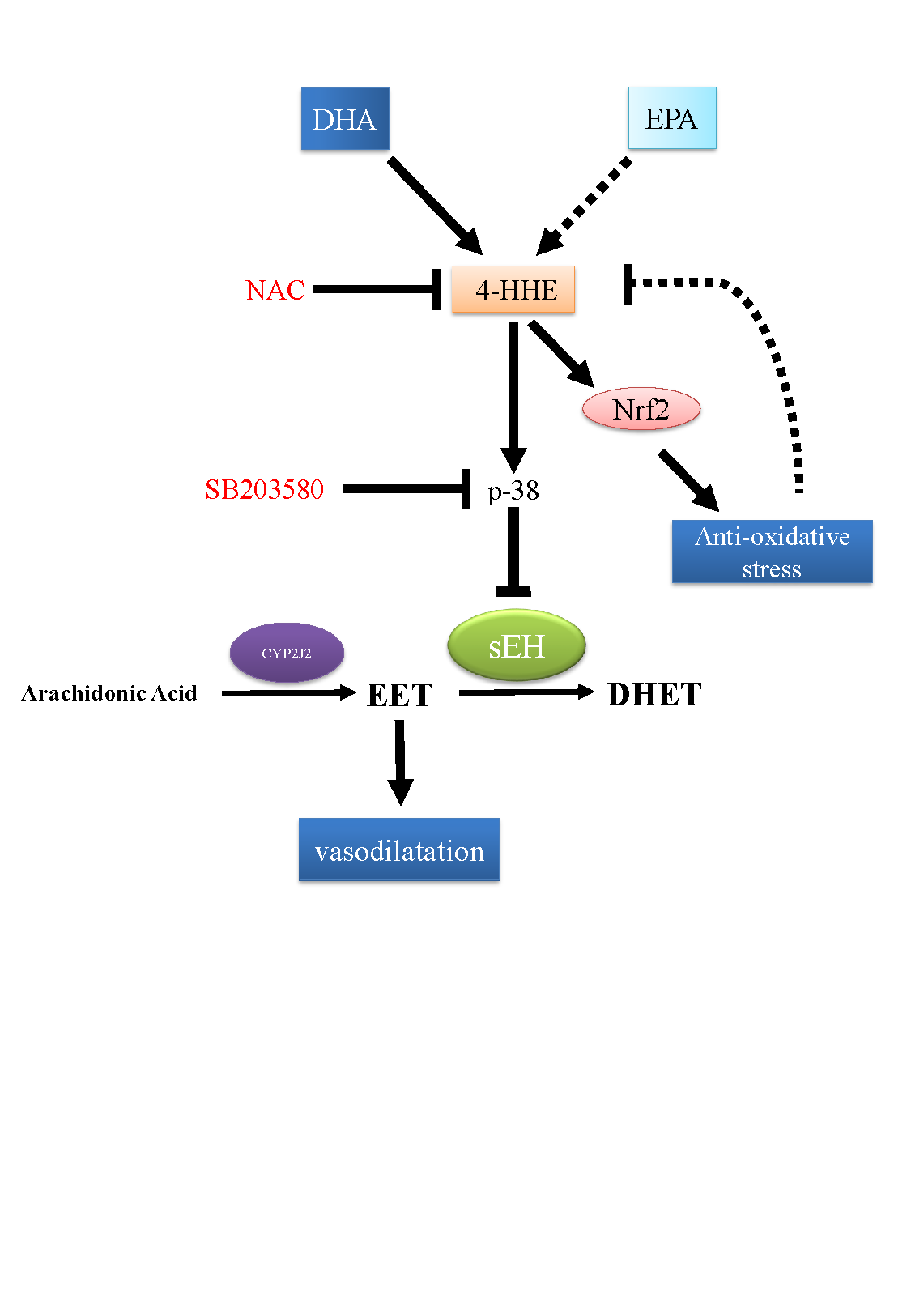
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| sEH | Soluble epoxide hydrolase |
| EET | epoxyeicosatrienoic acid |
| PECAM-1 | platelet endothelial cell adhesion molecule-1 |
| Nrf2 | nuclear factor erythroid 2-related factor 2 |
| 4-HHE | 4-hydroxy hexenal |
| PUFAs | polyunsaturated fatty acids |
| DHA | docosahexaenoic acid |
| EPA | eicosapentaenoic acid |
| HO-1 | heme oxygenase-1 |
| p38 | p38 kinase |
References
- Flammer, A.J.; Anderson, T.; Celermajer, D.S.; Creager, M.A.; Deanfield, J.; Ganz, P.; Hamburg, N.M.; Lüscher, T.F.; Shechter, M.; Taddei, S.; et al. The assessment of endothelial function: From research into clinical practice. Circulation 2012, 126, 753–767. [Google Scholar] [CrossRef] [PubMed]
- Félétou, M. The endothelium, part II: EDHF-mediated responses “The classical pathway”. In Colloquium Series on Integrated Systems Physiology: From Molecule to Function; Morgan & Claypool Publishers: Williston, FL, USA, 2011. [Google Scholar]
- Luksha, L.; Agewall, S.; Kublickiene, K. Endothelium-derived hyperpolarizing factor in vascular physiology and cardiovascular disease. Atherosclerosis 2009, 202, 330–344. [Google Scholar] [CrossRef] [PubMed]
- Campbell, W.B.; Fleming, I. Epoxyeicosatrienoic acids and endothelium-dependent responses. Pflugers. Arch. 2010, 459, 881–895. [Google Scholar] [CrossRef] [PubMed]
- Ayajiki, K.; Fujioka, H.; Toda, N.; Okada, S.; Minamiyama, Y.; Imaoka, S.; Funae, Y.; Watanabe, S.; Nakamura, A.; Okamura, T. Mediation of arachidonic acid metabolite(s) produced by endothelial cytochrome P-450 3A4 in monkey arterial relaxation. Hypertens. Res. 2003, 26, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Kaduce, T.L.; Weintraub, N.L.; Harmon, S.; Teesch, L.M.; Morisseau, C.; Thompson, D.A.; Hammock, B.D.; Spector, A.A. Pathways of epoxyeicosatrienoic acid metabolism in endothelial cells. Implications for the vascular effects of soluble epoxide hydrolase inhibition. J. Biol. Chem. 2001, 276, 14867–14874. [Google Scholar] [CrossRef] [PubMed]
- Lazaar, A.L.; Yang, L.; Boardley, R.L.; Goyal, N.S.; Robertson, J.; Baldwin, S.J.; Newby, D.E.; Wilkinson, I.B.; Tal-Singer, R.; Mayer, R.J.; et al. Pharmacokinetics, pharmacodynamics and adverse event profile of GSK2256294, a novel soluble epoxide hydrolase inhibitor. Br. J. Clin. Pharmacol. 2016, 81, 971–979. [Google Scholar] [CrossRef] [PubMed]
- Kromhout, D.; Bosschieter, E.B.; de Lezenne Coulander, C. The inverse relation between fish consumption and 20-year mortality from coronary heart disease. N. Engl. J. Med. 1985, 312, 1205–1209. [Google Scholar] [CrossRef] [PubMed]
- Iso, H.; Kobayashi, M.; Ishihara, J.; Sasaki, S.; Okada, K.; Kita, Y.; Kokubo, Y.; Tsugane, S.; Group, J.S. Intake of fish and n3 fatty acids and risk of coronary heart disease among Japanese: The Japan public health center-based (JPHC) study cohort I. Circulation 2006, 113, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Weitz, D.; Weintraub, H.; Fisher, E.; Schwartzbard, A.Z. Fish oil for the treatment of cardiovascular disease. Cardiol. Rev. 2010, 18, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Ellulu, M.S.; Khaza’ai, H.; Abed, Y.; Rahmat, A.; Ismail, P.; Ranneh, Y. Role of fish oil in human health and possible mechanism to reduce the inflammation. Inflammopharmacology 2015, 23, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Kondo, K.; Morino, K.; Nishio, Y.; Kondo, M.; Nakao, K.; Nakagawa, F.; Ishikado, A.; Sekine, O.; Yoshizaki, T.; Kashiwagi, A.; et al. A fish-based diet intervention improves endothelial function in postmenopausal women with type 2 diabetes mellitus: A randomized crossover trial. Metabolism 2014, 63, 930–940. [Google Scholar] [CrossRef] [PubMed]
- Sawada, T.; Tsubata, H.; Hashimoto, N.; Takabe, M.; Miyata, T.; Aoki, K.; Yamashita, S.; Oishi, S.; Osue, T.; Yokoi, K.; et al. Effects of 6-month eicosapentaenoic acid treatment on postprandial hyperglycemia, hyperlipidemia, insulin secretion ability, and concomitant endothelial dysfunction among newly-diagnosed impaired glucose metabolism patients with coronary artery disease. An open label, single blinded, prospective randomized controlled trial. Cardiovasc. Diabetol. 2016, 15, 121. [Google Scholar] [PubMed]
- Yagi, S.; Aihara, K.; Fukuda, D.; Takashima, A.; Hara, T.; Hotchi, J.; Ise, T.; Yamaguchi, K.; Tobiume, T.; Iwase, T.; et al. Effects of docosahexaenoic acid on the endothelial function in patients with coronary artery disease. J. Atheroscler. Thromb. 2015, 22, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.A.; Watts, G.F.; Burke, V.; Hilme, E.; Puddey, I.B.; Beilin, L.J. Differential effects of eicosapentaenoic acid and docosahexaenoic acid on vascular reactivity of the forearm microcirculation in hyperlipidemic, overweight men. Circulation 2000, 102, 1264–1269. [Google Scholar] [CrossRef] [PubMed]
- Ishikado, A.; Morino, K.; Nishio, Y.; Nakagawa, F.; Mukose, A.; Sono, Y.; Yoshioka, N.; Kondo, K.; Sekine, O.; Yoshizaki, T.; et al. 4-hydroxy hexenal derived from docosahexaenoic acid protects endothelial cells via Nrf2 activation. PLoS ONE 2013, 8, e69415. [Google Scholar] [CrossRef] [PubMed]
- López, D.; Orta, X.; Casós, K.; Sáiz, M.P.; Puig-Parellada, P.; Farriol, M.; Mitjavila, M.T. Upregulation of endothelial nitric oxide synthase in rat aorta after ingestion of fish oil-rich diet. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H567–572. [Google Scholar] [CrossRef] [PubMed]
- Okamura, T.; Tawa, M.; Geddawy, A.; Shimosato, T.; Iwasaki, H.; Shintaku, H.; Yoshida, Y.; Masada, M.; Shinozaki, K.; Imamura, T. Effects of atorvastatin, amlodipine, and their combination on vascular dysfunction in insulin-resistant rats. J. Pharmacol. Sci. 2014, 124, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Nagayama, K.; Morino, K.; Sekine, O.; Nakagawa, F.; Ishikado, A.; Iwasaki, H.; Okada, T.; Tawa, M.; Sato, D.; Imamura, T.; et al. Duality of n-3 polyunsaturated fatty acids on Mcp-1 expression in vascular smooth muscle: A potential role of 4-hydroxy hexenal. Nutrients 2015, 7, 8112–8126. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, F.; Morino, K.; Ugi, S.; Ishikado, A.; Kondo, K.; Sato, D.; Konno, S.; Nemoto, K.; Kusunoki, C.; Sekine, O.; et al. 4-hydroxy hexenal derived from dietary n-3 polyunsaturated fatty acids induces anti-oxidative enzyme heme oxygenase-1 in multiple organs. Biochem. Biophys. Res. Commun. 2014, 443, 991–996. [Google Scholar] [CrossRef] [PubMed]
- O’Brien-Coker, I.C.; Perkins, G.; Mallet, A.I. Aldehyde analysis by high performance liquid chromatography/tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2001, 15, 920–928. [Google Scholar] [CrossRef] [PubMed]
- Mavrommatis, Y.; Ross, K.; Rucklidge, G.; Reid, M.; Duncan, G.; Gordon, M.J.; Thies, F.; Sneddon, A.; de Roos, B. Intervention with fish oil, but not with docosahexaenoic acid, results in lower levels of hepatic soluble epoxide hydrolase with time in apoE knockout mice. Br. J. Nutr. 2010, 103, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Caligiuri, S.P.; Aukema, H.M.; Ravandi, A.; Guzman, R.; Dibrov, E.; Pierce, G.N. Flaxseed consumption reduces blood pressure in patients with hypertension by altering circulating oxylipins via an α-linolenic acid-induced inhibition of soluble epoxide hydrolase. Hypertension 2014, 64, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Dey, A.; Romanko, O.P.; Stepp, D.W.; Wang, M.H.; Zhou, Y.; Jin, L.; Pollock, J.S.; Webb, R.C.; Imig, J.D. Decreased epoxygenase and increased epoxide hydrolase expression in the mesenteric artery of obese zucker rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Förstermann, U.; Münzel, T. Endothelial nitric oxide synthase in vascular disease: From marvel to menace. Circulation 2006, 113, 1708–1714. [Google Scholar] [CrossRef] [PubMed]
- Grune, T.; Davies, K.J. The proteasomal system and HNE-modified proteins. Mol. Aspects Med. 2003, 24, 195–204. [Google Scholar] [CrossRef]
- Huot, J.; Houle, F.; Marceau, F.; Landry, J. Oxidative stress-induced actin reorganization mediated by the p38 mitogen-activated protein kinase/heat shock protein 27 pathway in vascular endothelial cells. Circ. Res. 1997, 80, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Oguro, A.; Oida, S.; Imaoka, S. Down-regulation of EPHX2 gene transcription by Sp1 under high-glucose conditions. Biochem. J. 2015, 470, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Dasari, A.; Bartholomew, J.N.; Volonte, D.; Galbiati, F. Oxidative stress induces premature senescence by stimulating caveolin-1 gene transcription through p38 mitogen-activated protein kinase/Sp1-mediated activation of two GC-rich promoter elements. Cancer Res. 2006, 66, 10805–10814. [Google Scholar] [CrossRef] [PubMed]
- Shechter, M.; Sharir, M.; Labrador, M.J.; Forrester, J.; Silver, B.; Bairey Merz, C.N. Oral magnesium therapy improves endothelial function in patients with coronary artery disease. Circulation 2000, 102, 2353–2358. [Google Scholar] [CrossRef] [PubMed]
- Daiber, A.; Steven, S.; Weber, A.; Shuvaev, V.V.; Muzykantov, V.R.; Laher, I.; Li, H.; Lamas, S.; Munzel, T. Targeting vascular (endothelial) dysfunction. Br. J. Pharmacol. 2017, 174, 1591–1619. [Google Scholar] [CrossRef] [PubMed]
- Marsh, S.A.; Coombes, J.S. Exercise and the endothelial cell. Int. J. Cardiol. 2005, 99, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Fleming, I. The pharmacology of the cytochrome P450 epoxygenase/soluble epoxide hydrolase axis in the vasculature and cardiovascular disease. Pharmacol. Rev. 2014, 66, 1106–1140. [Google Scholar] [CrossRef] [PubMed]






© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okada, T.; Morino, K.; Nakagawa, F.; Tawa, M.; Kondo, K.; Sekine, O.; Imamura, T.; Okamura, T.; Ugi, S.; Maegawa, H. N-3 Polyunsaturated Fatty Acids Decrease the Protein Expression of Soluble Epoxide Hydrolase via Oxidative Stress-Induced P38 Kinase in Rat Endothelial Cells. Nutrients 2017, 9, 654. https://doi.org/10.3390/nu9070654
Okada T, Morino K, Nakagawa F, Tawa M, Kondo K, Sekine O, Imamura T, Okamura T, Ugi S, Maegawa H. N-3 Polyunsaturated Fatty Acids Decrease the Protein Expression of Soluble Epoxide Hydrolase via Oxidative Stress-Induced P38 Kinase in Rat Endothelial Cells. Nutrients. 2017; 9(7):654. https://doi.org/10.3390/nu9070654
Chicago/Turabian StyleOkada, Takashi, Katsutaro Morino, Fumiyuki Nakagawa, Masashi Tawa, Keiko Kondo, Osamu Sekine, Takeshi Imamura, Tomio Okamura, Satoshi Ugi, and Hiroshi Maegawa. 2017. "N-3 Polyunsaturated Fatty Acids Decrease the Protein Expression of Soluble Epoxide Hydrolase via Oxidative Stress-Induced P38 Kinase in Rat Endothelial Cells" Nutrients 9, no. 7: 654. https://doi.org/10.3390/nu9070654
APA StyleOkada, T., Morino, K., Nakagawa, F., Tawa, M., Kondo, K., Sekine, O., Imamura, T., Okamura, T., Ugi, S., & Maegawa, H. (2017). N-3 Polyunsaturated Fatty Acids Decrease the Protein Expression of Soluble Epoxide Hydrolase via Oxidative Stress-Induced P38 Kinase in Rat Endothelial Cells. Nutrients, 9(7), 654. https://doi.org/10.3390/nu9070654