n-3 Polyunsaturated Fatty Acids and Metabolic Syndrome Risk: A Meta-Analysis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Study Selection
2.2. Inclusive Criteria
2.3. Data Extraction
2.4. Statistical Analysis
3. Results
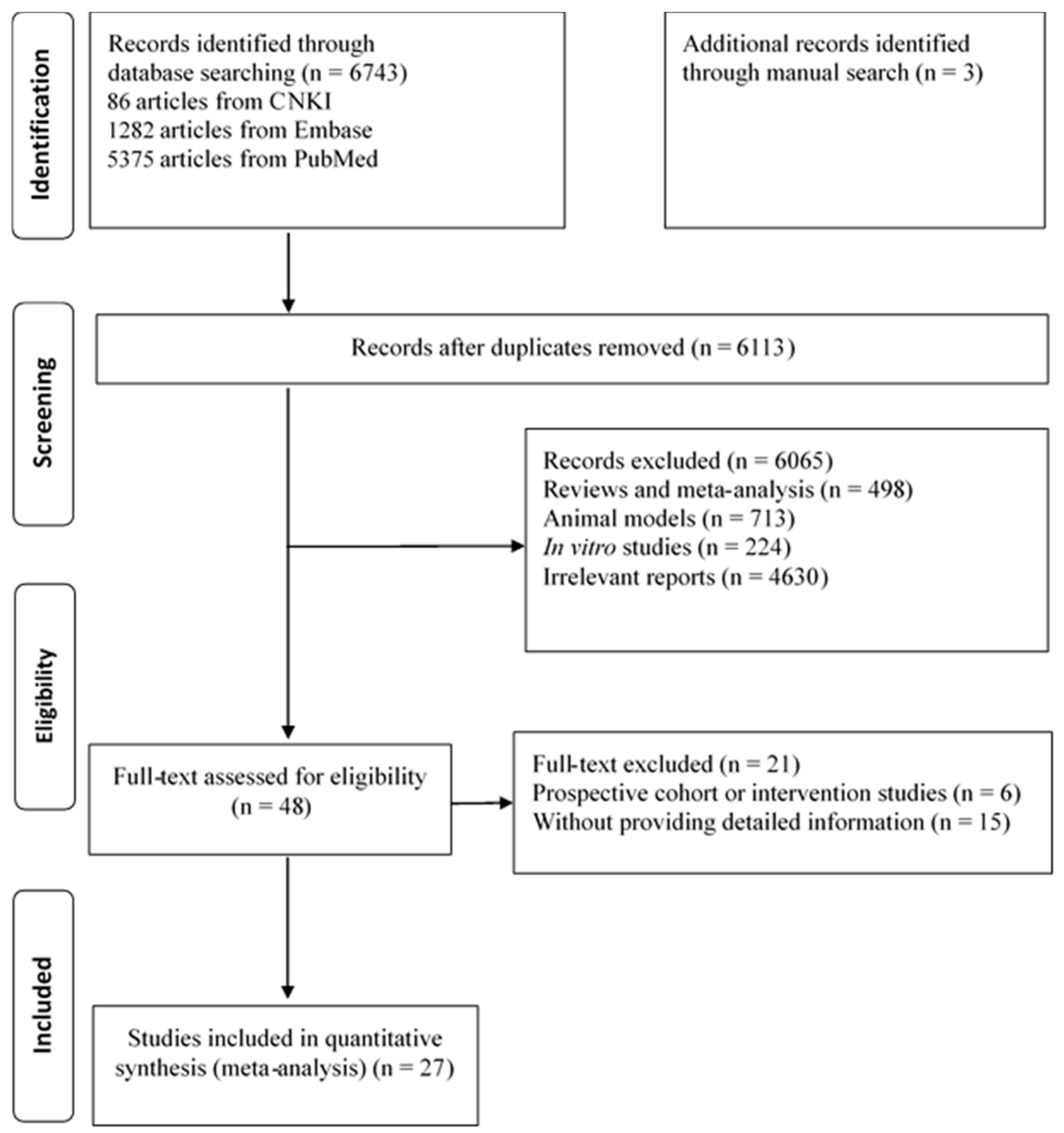
3.1. Literature Search
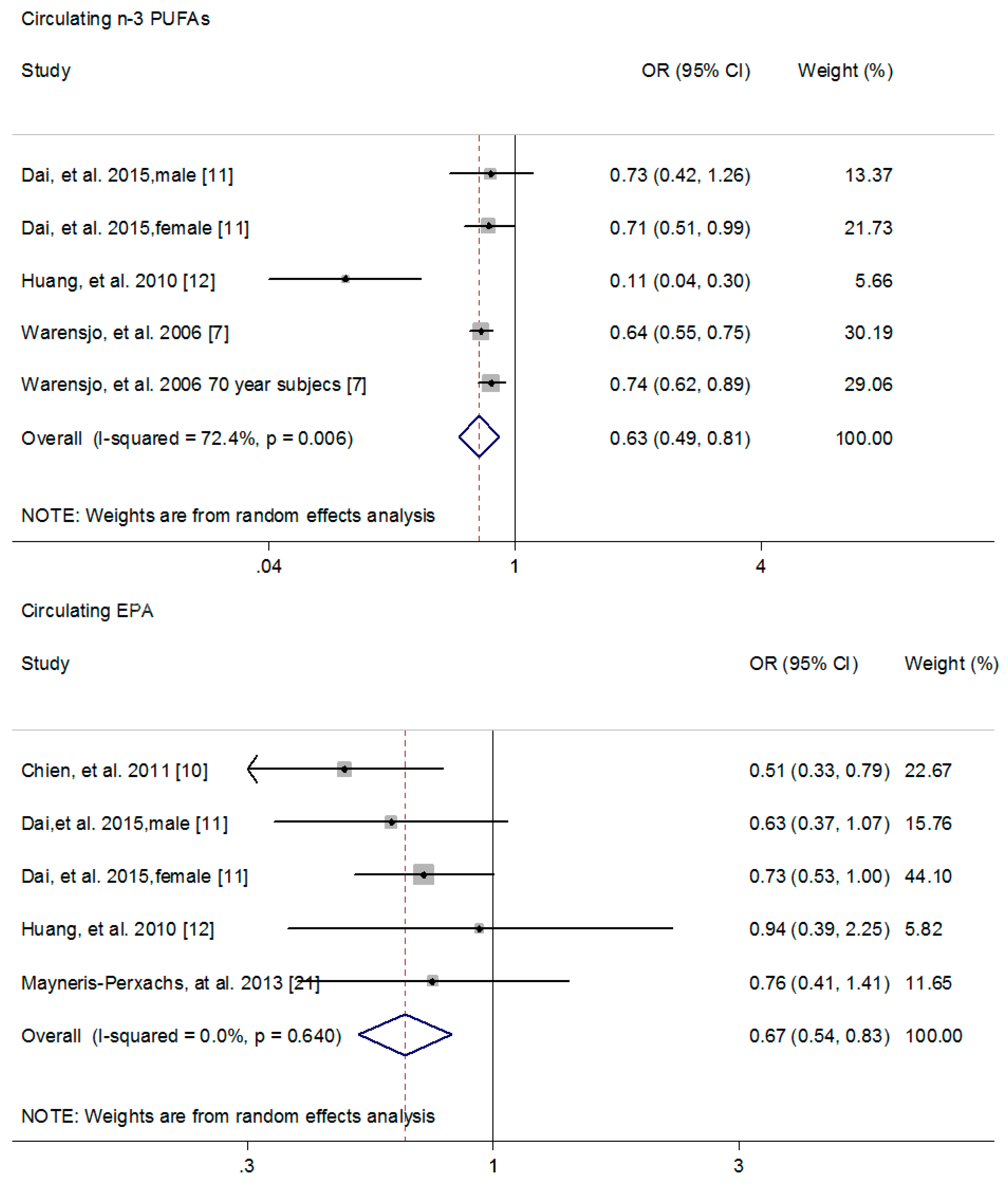
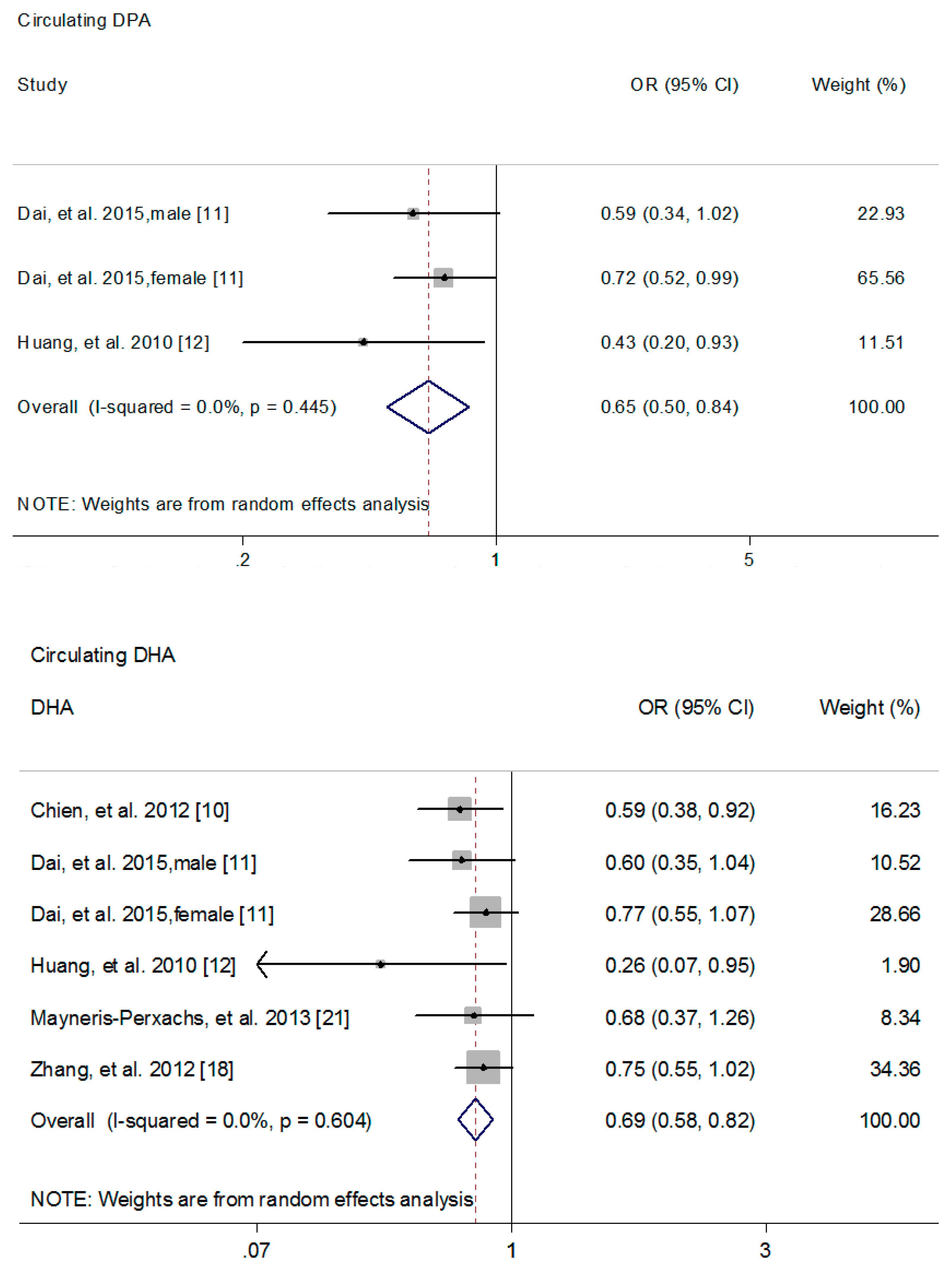
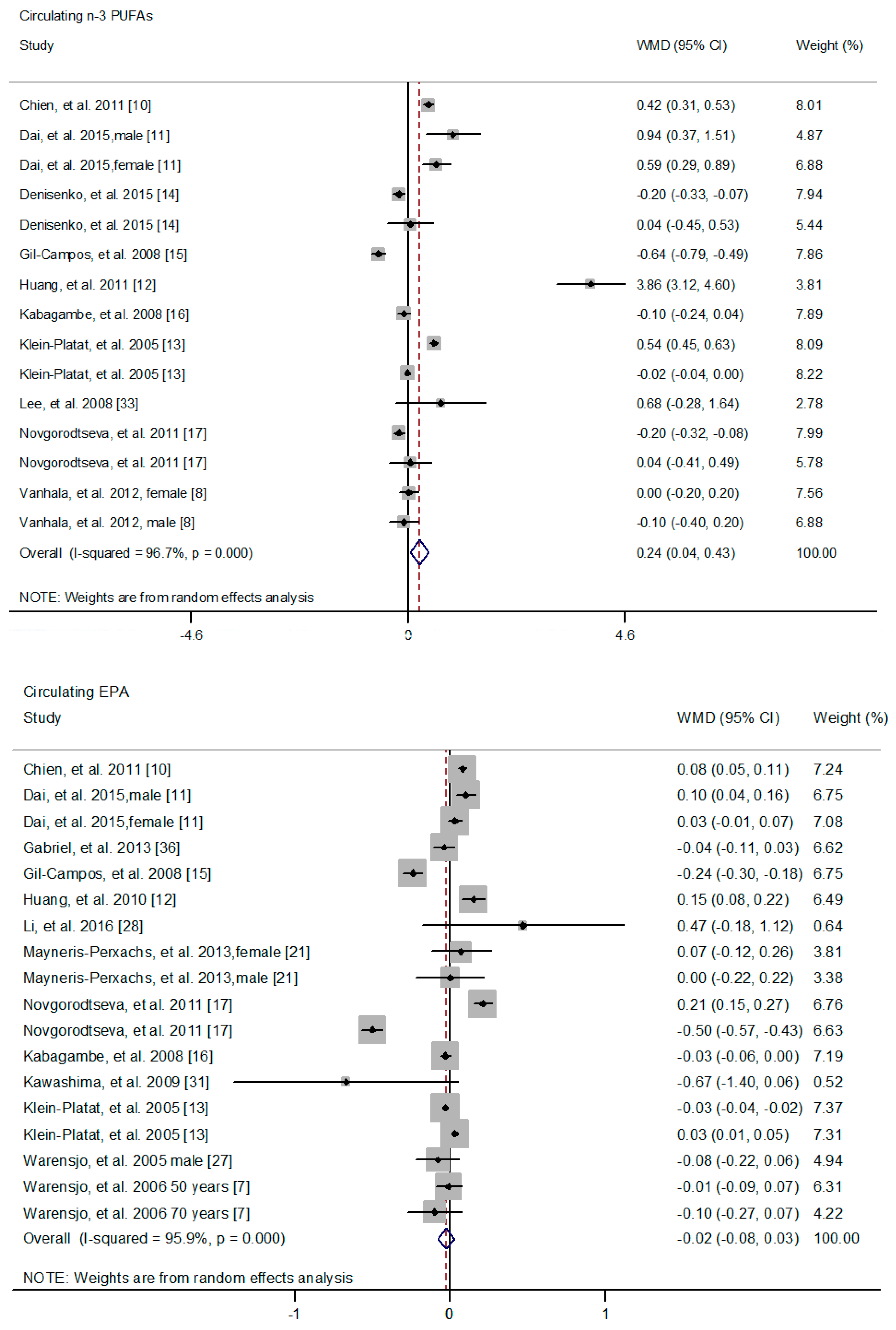
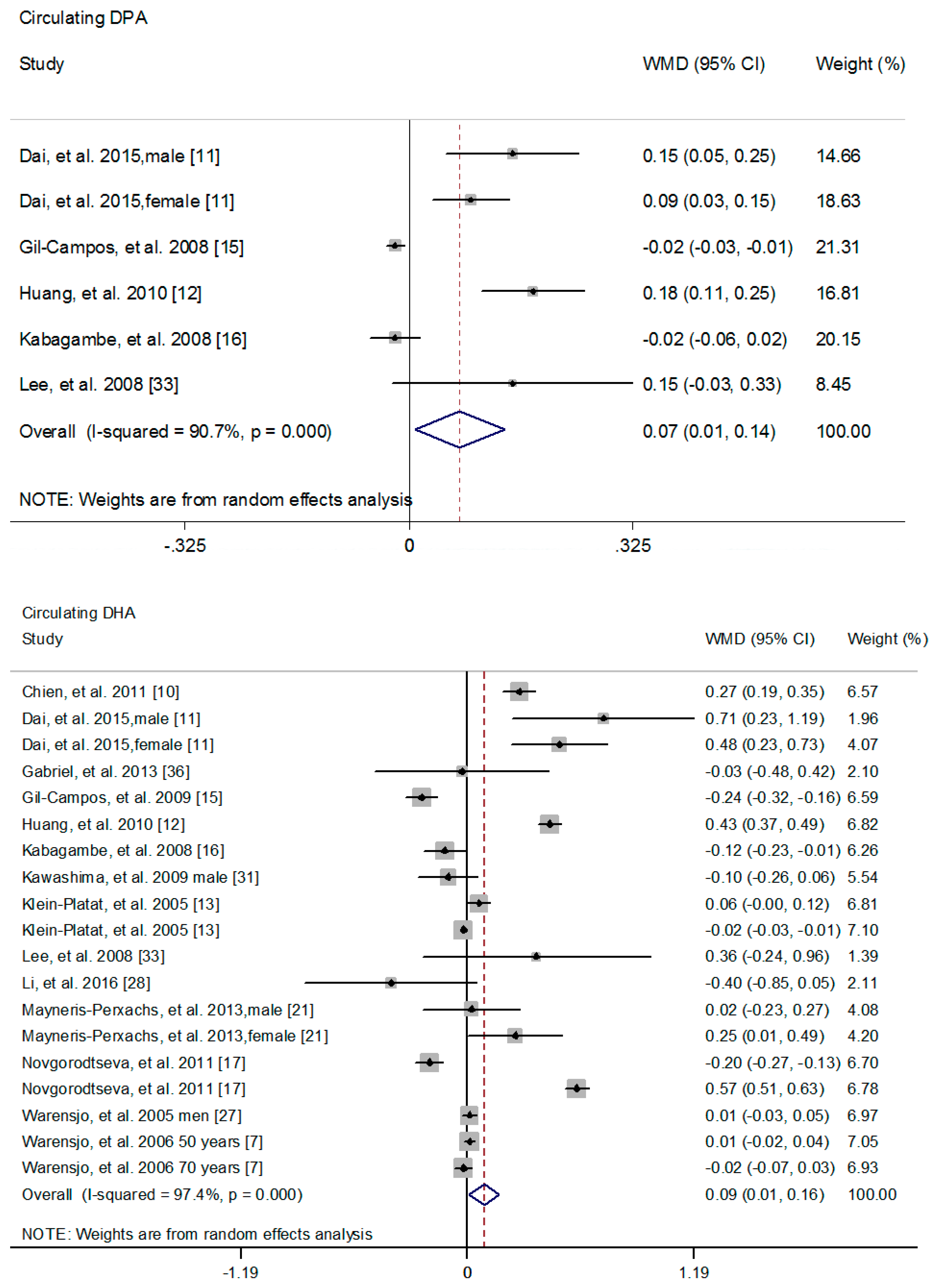
3.2. The Association of n-3 PUFAs with MetS Risk
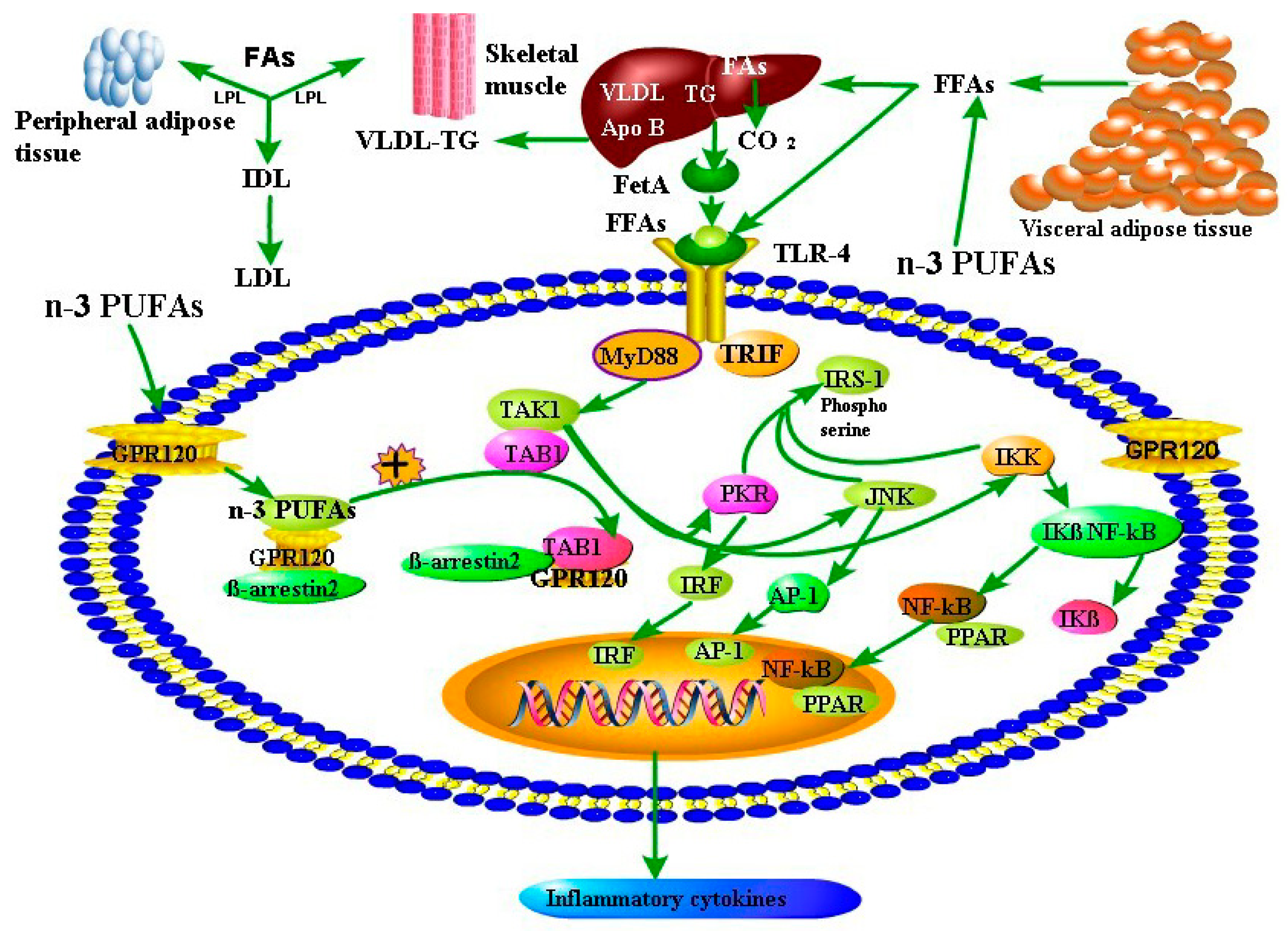
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Expert Panel on Detection, Evaluation, and Treatment of High Blood Cholesterol in Adults. Executive summary of the third report of the national cholesterol education program (NCEP) expert panel on detection, evaluation, and treatment of high blood cholesterol in adults (adult treatment panel III). JAMA 2001, 285, 2486–2497. [Google Scholar]
- Laaksonen, D.E.; Lakka, H.M.; Niskanen, L.K.; Kaplan, G.A.; Salonen, J.T.; Lakka, T.A. Metabolic syndrome and development of diabetes mellitus: Application and validation of recently suggested definitions of the metabolic syndrome in a prospective cohort study. Am. J. Epidemiol. 2002, 156, 1070–1077. [Google Scholar] [CrossRef] [PubMed]
- Lakka, H.M.; Laaksonen, D.E.; Lakka, T.A.; Niskanen, L.K.; Kumpusalo, E.; Tuomilehto, J.; Salonen, J.T. The metabolic syndrome and total and cardiovascular disease mortality in middle-aged men. JAMA 2002, 288, 2709–2716. [Google Scholar] [CrossRef] [PubMed]
- Carpentier, Y.A.; Portois, L.; Malaisse, W.J. n-3 fatty acids and the metabolic syndrome. Am. J. Clin. Nutr. 2006, 83, 481S–487S. [Google Scholar]
- Grundy, S.M. Diagnosis and management of the metabolic syndrome an American heart association/national heart, lung, and blood institute scientific statement. Curr. Opin. Cardiol. 2005, 21, 1–6. [Google Scholar] [CrossRef]
- Baik, I.; Abbott, R.D.; Curb, J.D.; Shin, C. Intake of fish and n-3 fatty acids and future risk of metabolic syndrome. J. Am. Diet. Assoc. 2010, 110, 1018–1026. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Xun, P.; Iribarren, C.; Horn, L.V.; Steffen, L.; Daviglus, M.L.; Siscovick, D.; Liu, K.; He, K. Intake of fish and long-chain omega-3 polyunsaturated fatty acids and incidence of metabolic syndrome among American young adults: A 25-year follow-up study. Eur. J. Nutr. 2016, 55, 1707–1716. [Google Scholar] [CrossRef] [PubMed]
- Vanhala, M.; Saltevo, J.; Soininen, P.; Kautiainen, H.; Kangas, A.J.; Alakorpela, M.; Mäntyselkä, P. Serum omega-6 polyunsaturated fatty acids and the metabolic syndrome: A longitudinal population-based cohort study. Am. J. Epidemiol. 2012, 176, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Warensjö, E.; Sundström, J.; Lind, L.; Vessby, B. Factor analysis of fatty acids in serum lipids as a measure of dietary fat quality in relation to the metabolic syndrome in men. Am. J. Clin. Nutr. 2006, 84, 442–448. [Google Scholar] [PubMed]
- Chien, K.L.; Chao, C.L.; Kuo, C.H.; Lin, H.J.; Liu, P.H.; Chen, P.R.; Hsu, H.C.; Lee, B.C.; Lee, Y.T.; Chen, M.F. Plasma fatty acids and the risk of metabolic syndrome in ethnic Chinese adults in Taiwan. Lipids Health Dis. 2011, 10, 33. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.W.; Chen, Y.M.; Zeng, F.F.; Sun, L.L.; Chen, C.G.; Su, Y.X. Association between n-3 polyunsaturated fatty acids in erythrocytes and metabolic syndrome in Chinese men and women. Eur. J. Nutr. 2015, 55, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Bhulaidok, S.; Cai, Z.; Xu, T.; Xu, F.; Wahlqvist, M.L.; Li, D. Plasma phospholipids n-3 polyunsaturated fatty acid is associated with metabolic syndrome. Mol. Nutr. Food Res. 2010, 54, 1628–1635. [Google Scholar] [CrossRef] [PubMed]
- Klein-Platat, C.; Drai, J.; Oujaa, M.; Schlienger, J.L.; Simon, C. Plasma fatty acid composition is associated with the metabolic syndrome and low-grade inflammation in overweight adolescents. Am. J. Clin. Nutr. 2005, 82, 1178–1184. [Google Scholar] [PubMed]
- Denisenko, Y.K.; Novgorodtseva, T.P.; Zhukova, N.V.; Gvozdenko, T.A.; Antonyuk, M.V.; Knyshova, V.V.; Nazarenko, A.V. Metabolic syndrome: Modification of the fatty acid composition and glucose-insulin homeostasis. Br. J. Med. Med. Res. 2015, 8, 975–987. [Google Scholar] [CrossRef]
- Gil-Campos, M.; Ramírez-Tortosa, M.D.C.; Larqué, E.; Linde, J.; Aguilera, C.M.; Cañete, R.; Gil, A. Metabolic syndrome affects fatty acid composition of plasma lipids in obese prepubertal children. Lipids 2008, 43, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Kabagambe, E.K.; Tsai, M.Y.; Hopkins, P.N.; Ordovas, J.M.; Peacock, J.M.; Borecki, I.B.; Arnett, D.K. Erythrocyte fatty acid composition and the metabolic syndrome: A national heart, lung, and blood institute goldn study. Clin. Chem. 2008, 54, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Novgorodtseva, T.P. Composition of fatty acids in plasma and erythrocytes and eicosanoids level in patients with metabolic syndrome. Lipids Health Dis. 2011, 10, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Sun, Q.; Hu, F.B.; Ye, X.; Yu, Z.; Zong, G.; Li, H.; Zhou, Y.; Lin, X. Erythrocyte n-3 fatty acids and metabolic syndrome in middle-aged and older Chinese. J. Clin. Endocrinol. Metab. 2012, 97, E973–E977. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Xun, P.; He, K. Fish consumption, long-chain omega-3 polyunsaturated fatty acid intake and risk of metabolic syndrome: A meta-analysis. Nutrients 2015, 7, 2085–2100. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.P.; Green, S. Cochrane Handbook for Systematic Reviews of Interventions: Cochrane Book Series; Wiley-Blackwell: Hoboken, NJ, USA, 2008; pp. 102–108. [Google Scholar]
- Mayneris-Perxachs, J.; Guerendiain, M.; Castellote, A.I.; Estruch, R.; Covas, M.I.; Fitó, M.; Salassalvadó, J.; Martínezgonzález, M.A.; Aros, F.; Lamuelaraventós, R.M. Plasma fatty acid composition, estimated desaturase activities, and their relation with the metabolic syndrome in a population at high risk of cardiovascular disease. Clin. Nutr. 2014, 33, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Mennen, L.I.; Lafay, L.; Feskens, E.J.M.; Novak, M.; Lépinay, P.; Balkau, B. Possible protective effect of bread and dairy products on the risk of the metabolic syndrome. Nutr. Res. 2000, 20, 335–347. [Google Scholar] [CrossRef]
- Kouki, R.; Schwab, U.; Hassinen, M.; Komulainen, P.; Heikkilä, H.; Lakka, T.A.; Rauramaa, R. Food consumption, nutrient intake and the risk of having metabolic syndrome: The DR’s Extra study. Eur. J. Clin. Nutr. 2011, 65, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Ruidavets, J.B.; Bongard, V.; Dallongeville, J.; Arveiler, D.; Ducimetière, P.; Perret, B.; Simon, C.; Amouyel, P.; Ferrières, J. High consumptions of grain, fish, dairy products and combinations of these are associated with a low prevalence of metabolic syndrome. J. Epidemiol. Commun. H 2007, 61, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Tørris, C.; Molin, M.; Cvancarova, M.S. Lean fish consumption is associated with lower risk of metabolic syndrome: A Norwegian cross sectional study. BMC Public Health 2016, 16, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tørris, C.; Molin, M.; Cvancarova, S.M. Associations between fish consumption and metabolic syndrome. A large cross-sectional study from the Norwegian Tromsø Study: Tromsø 4. Diabetol. Metab. Syndr. 2016, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Warensjö, E.; Risérus, U.; Vessby, B. Fatty acid composition of serum lipids predicts the development of the metabolic syndrome in men. Diabetologia 2005, 48, 1999–2005. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Dai, Y.; Liu, Y.; Zheng, J.; Liu, W. Comparison of serum adiponectin, free fatty acid composition, and other lipid metabolic parameters among patients with different component numbers of metabolic syndrome. Chin. J. Diabetes 2007, 15, 968. [Google Scholar]
- Mirmiran, P.; Hosseinpour-Niazi, S.; Naderi, Z.; Bahadoran, Z.; Sadeghi, M.; Azizi, F. Association between interaction and ratio of ω-3 and ω-6 polyunsaturated fatty acid and the metabolic syndrome in adults. Nutrition 2012, 28, 856–863. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Ahn, J.; Kim, N.S.; Lee, B.K. High carbohydrate diets are positively associated with the risk of metabolic syndrome irrespective to fatty acid composition in women: The KNHANES 2007–2014. Int. J. Food Sci. Nut. 2017, 68, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, A.; Sugawara, S.; Okita, M.; Akahane, T.; Fukui, K.; Hashiuchi, M.; Kataoka, C.; Tsukamoto, I. Plasma fatty acid composition, estimated desaturase activities, and intakes of energy and nutrient in Japanese men with abdominal obesity or metabolic syndrome. J. Nutr. Sci. Vitaminol. 2009, 55, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Zaribaf, F.; Falahi, E.; Barak, F.; Heidari, M.; Keshteli, A.H.; Yazdannik, A.; Esmaillzadeh, A. Fish consumption is inversely associated with the metabolic syndrome. Eur. J. Clin. Nutr. 2014, 68, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Park, S.L.Y. n-3 polyunsaturated fatty acids and trans fatty acids in patients with the metabolic syndrome: A case-control study in Korea. Br. J. Nutr. 2008, 100, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.L.; Petrone, A.B.; Pankow, J.S.; Arnett, D.K.; North, K.E.; Ellison, R.C.; Hunt, S.C.; Djoussé, L. Association of dietary omega-3 fatty acids with prevalence of metabolic syndrome: The national heart, lung, and blood institute family heart study. Clin. Nutr. 2013, 32, 966–969. [Google Scholar] [CrossRef] [PubMed]
- Noel, S.E.; Newby, P.K.; Ordovas, J.M.; Tucker, K.L. Adherence to an (n-3) fatty acid/fish intake pattern is inversely associated with metabolic syndrome among Puerto Rican adults in the greater Boston area. J. Nutr. 2010, 140, 1846–1854. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, M.; Azucena, M.; Alejandra, S.F.; Jorge, M.H. Association between metabolic syndrome and erythrocyte fatty acid profile in Mexican adolescents: A trans fatty acid approach. Food Nutr. Sci. 2013, 4, 51–58. [Google Scholar] [CrossRef]
- Hodson, L.; Skeaff, C.M.; Fielding, B.A. Fatty acid composition of adipose tissue and blood in humans and its use as a biomarker of dietary intake. Prog. Lipid Res. 2008, 47, 348–380. [Google Scholar] [CrossRef] [PubMed]
- Vessby, B.; Gustafsson, I.B.; Tengblad, S.; Boberg, M.; Andersson, A. Desaturation and elongation of fatty acids and insulin action. Ann. N. Y. Acad. Sci. 2002, 967, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Forouhi, N.G.; Fumiaki, I.; Sharp, S.J.; Albert, K.; Schulze, M.B.; Zheng, J.; Ye, Z.; Ivonne, S.; Marcela, G.; María, H.J. Association of plasma phospholipid n-3 and n-6 polyunsaturated fatty acids with type 2 diabetes: The epic-interact case-cohort study. PLoS Med. 2016, 13, e1002094. [Google Scholar] [CrossRef] [PubMed]
- Dembinska, A. Effects of dietary fat modification on insulin sensitivity and on other risk factors of the metabolic syndrome—LIPGENE: A European randomized dietary intervention. Int. J. Obes. 2011, 35, 800–809. [Google Scholar]
- Paniagua, J.A.; Pérez-Martinez, P.; Gjelstad, I.M.; Tierney, A.C.; Delgado-Lista, J.; Defoort, C.; Blaak, E.E.; Risérus, U.; Drevon, C.A.; Kiec-Wilk, B. A low-fat high-carbohydrate diet supplemented with long-chain n-3 PUFA reduces the risk of the metabolic syndrome. Atherosclerosis 2011, 218, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Simão, A.N.; Lozovoy, M.A.; Bahls, L.D.; Morimoto, H.K.; Simão, T.N.; Matsuo, T.; Dichi, I. Blood pressure decrease with ingestion of a soya product (kinako) or fish oil in women with the metabolic syndrome: Role of adiponectin and nitric oxide. Br. J. Nutr. 2012, 108, 1435–1442. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, C.; Botella-Carretero, J.I.; Corella, D.; Fiol, M.; Lage, M.; Lurbe, E.; Richart, C.; Fernández-Real, J.M.; Fuentes, F.; Ordóñez, A. White fish reduces cardiovascular risk factors in patients with metabolic syndrome: The wish-care study, a multicenter randomized clinical trial. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Venturini, D.; Simão, A.N.; Urbano, M.R.; Dichi, I. Effects of extra virgin olive oil and fish oil on lipid profile and oxidative stress in patients with metabolic syndrome. Nutrition 2015, 31, 834–840. [Google Scholar] [CrossRef] [PubMed]
- Musso, G.; Gambino, R.; Cassader, M. Recent insights into hepatic lipid metabolism in non-alcoholic fatty liver disease (NAFLD). Prog. Lipid Res. 2009, 48, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Scorletti, E.; Byrne, C.D. Omega-3 fatty acids, hepatic lipid metabolism, and nonalcoholic fatty liver disease. Annu. Rev. Nutr. 2013, 33, 231–248. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, Y.; Yahagi, N.; Izumida, Y.; Nishi, M.; Kubota, M.; Teraoka, Y.; Yamamoto, T.; Matsuzaka, T.; Nakagawa, Y.; Sekiya, M. Polyunsaturated fatty acids selectively suppress sterol regulatory element-binding protein-1 through proteolytic processing and autoloop regulatory circuit. J. Biol. Chem. 2010, 285, 11681–11691. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Dong, Q.; Bridges, D.; Raghow, R.; Park, E.A.; Elam, M.B. Docosahexaenoic acid inhibits proteolytic processing of sterol regulatory element-binding protein-1c (SREBP-1c) via activation of amp-activated kinase. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2015, 1851, 1521–1529. [Google Scholar] [CrossRef] [PubMed]
- Salem, N., Jr.; Litman, B.; Kim, H.Y.; Gawrisch, K. Mechanisms of action of docosahexaenoic acid. Lipids 2001, 36, 945–959. [Google Scholar]
- Bugianesi, E.; Mccullough, A.J.; Marchesini, G. Insulin resistance: A metabolic pathway to chronic liver disease. Hepatology 2005, 42, 987–1000. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Mechanisms of action of (n-3) fatty acids. J. Nutr. 2012, 142, 592S–599S. [Google Scholar] [CrossRef] [PubMed]
- Baker, R.G.; Hayden, M.S.; Ghosh, S. Nf-κb, inflammation and metabolic disease. Cell Metab. 2011, 13, 11–12. [Google Scholar] [CrossRef] [PubMed]
- Gregor, M.F.; Hotamisligil, G.S. Inflammatory mechanisms in obesity. Annu. Rev. Immunol. 2011, 29, 415–445. [Google Scholar] [CrossRef] [PubMed]
- De, C.R. n-3 fatty acids in cardiovascular disease. N. Engl. J. Med. 2011, 364, 2439–2450. [Google Scholar]
- Meydani, S.N.; Endres, S.; Woods, M.M.; Goldin, B.R.; Soo, C.; Morrilllabrode, A.; Dinarello, C.A.; Gorbach, S.L. Oral (n-3) fatty acid supplementation suppresses cytokine production and lymphocyte proliferation: Comparison between young and older women. J. Nutr. 1991, 121, 547–555. [Google Scholar] [PubMed]
- Endres, S.; Ghorbani, R.; Kelley, V.E.; Georgilis, K.; Lonnemann, G.; Jw, V.D.M.; Cannon, J.G.; Rogers, T.S.; Klempner, M.S.; Weber, P.C. The effect of dietary supplementation with n-3 polyunsaturated fatty acids on the synthesis of interleukin-1 and tumor necrosis factor by mononuclear cells. N. Engl. J. Med. 1989, 320, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Sohn, K.H.; Rhee, S.H.; Hwang, D. Saturated fatty acids, but not unsaturated fatty acids, induce the expression of cyclooxygenase-2 mediated through toll-like receptor 4. J. Biol. Chem. 2001, 276, 16683–16689. [Google Scholar] [CrossRef] [PubMed]
- Novak, T.E.; Babcock, T.A.; Jho, D.H.; Helton, W.S.; Espat, N.J. NF-kappa B inhibition by omega-3 fatty acids modulates LPS-stimulated macrophage TNF-alpha transcription. Am. J. Physiol. Lung Cell. Mol. Physiol. 2003, 284, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.Y. Gpr120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell 2010, 142, 687–698. [Google Scholar] [CrossRef] [PubMed]






© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, X.-f.; Li, X.; Shi, M.; Li, D. n-3 Polyunsaturated Fatty Acids and Metabolic Syndrome Risk: A Meta-Analysis. Nutrients 2017, 9, 703. https://doi.org/10.3390/nu9070703
Guo X-f, Li X, Shi M, Li D. n-3 Polyunsaturated Fatty Acids and Metabolic Syndrome Risk: A Meta-Analysis. Nutrients. 2017; 9(7):703. https://doi.org/10.3390/nu9070703
Chicago/Turabian StyleGuo, Xiao-fei, Xin Li, Meiqi Shi, and Duo Li. 2017. "n-3 Polyunsaturated Fatty Acids and Metabolic Syndrome Risk: A Meta-Analysis" Nutrients 9, no. 7: 703. https://doi.org/10.3390/nu9070703
APA StyleGuo, X. -f., Li, X., Shi, M., & Li, D. (2017). n-3 Polyunsaturated Fatty Acids and Metabolic Syndrome Risk: A Meta-Analysis. Nutrients, 9(7), 703. https://doi.org/10.3390/nu9070703