Administration of Inulin-Supplemented Gluten-Free Diet Modified Calcium Absorption and Caecal Microbiota in Rats in a Calcium-Dependent Manner



Abstract
:1. Introduction
2. Materials and Methods
2.1. Composition of Experimental Gluten-Free Diets
2.2. Animals
2.3. Experimental Design
2.4. Analysis of Food Intake, Body Weight Gain and Minerals Bioavailability
2.5. Sampling Procedures
2.6. Analytical Procedures
2.6.1. Ammonia and Dry Matter (DM) Content
2.6.2. Short Chain Fatty Acids
2.6.3. Activity of Bacterial Enzymes
2.6.4. Microbiota Characteristics with PCR-DGGE
2.6.5. Quantification of Caecal Microbiota by Real-Time PCR
2.6.6. Mineral Concentration
2.7. Statistic Analysis
3. Results
3.1. Effect of a Gluten-Free Diet Enriched with Inulin on a Daily Food Intake, Body Weight Gain and Gastrointestinal Tract Parameters
3.2. Effect of a Gluten-Free Diet Enriched with Inulin on the Concentration of Short Chain Fatty Acids (SCFAs), Putrefactive SCFAs (PSCFAs) and Their Profiles in the Caecal Digesta
3.3. Effect of a Gluten-Free Diet Enriched with Inulin on the Activity of Bacterial Enzymes in the Caecal Digesta
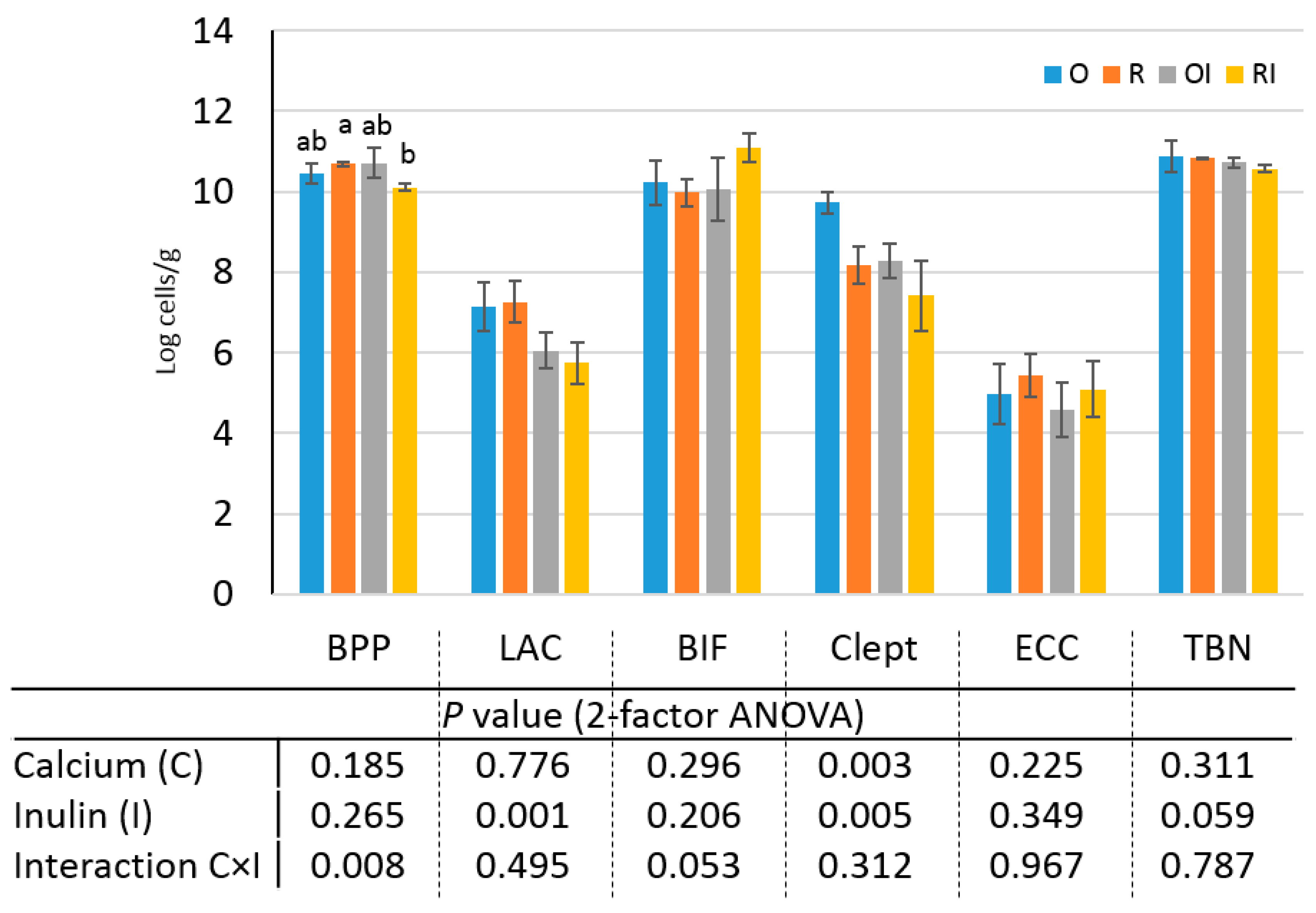
3.4. Effect of a Gluten-Free Diet Enriched with Inulin on the Quantitative Profile of Caecal Microbiota
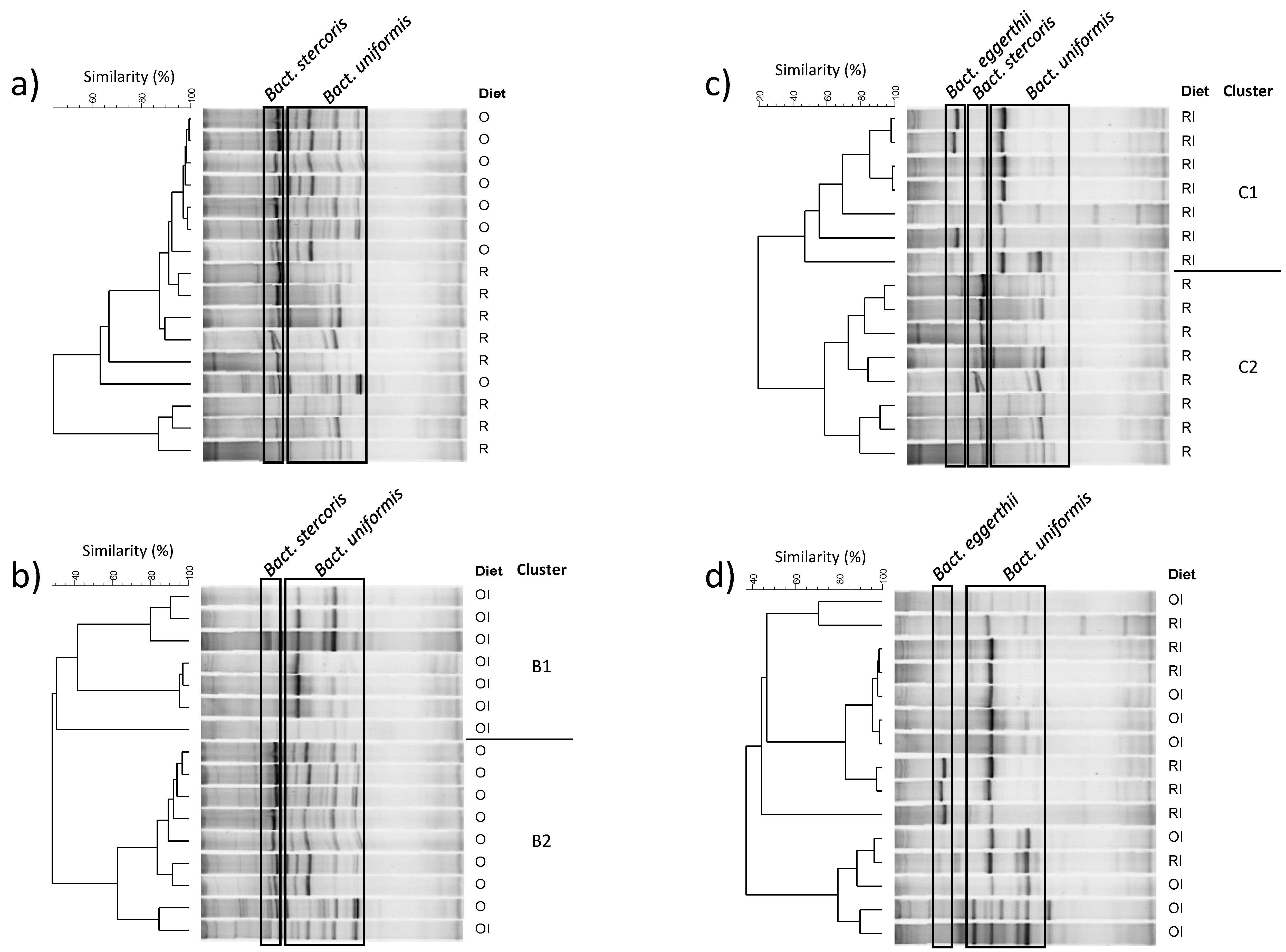
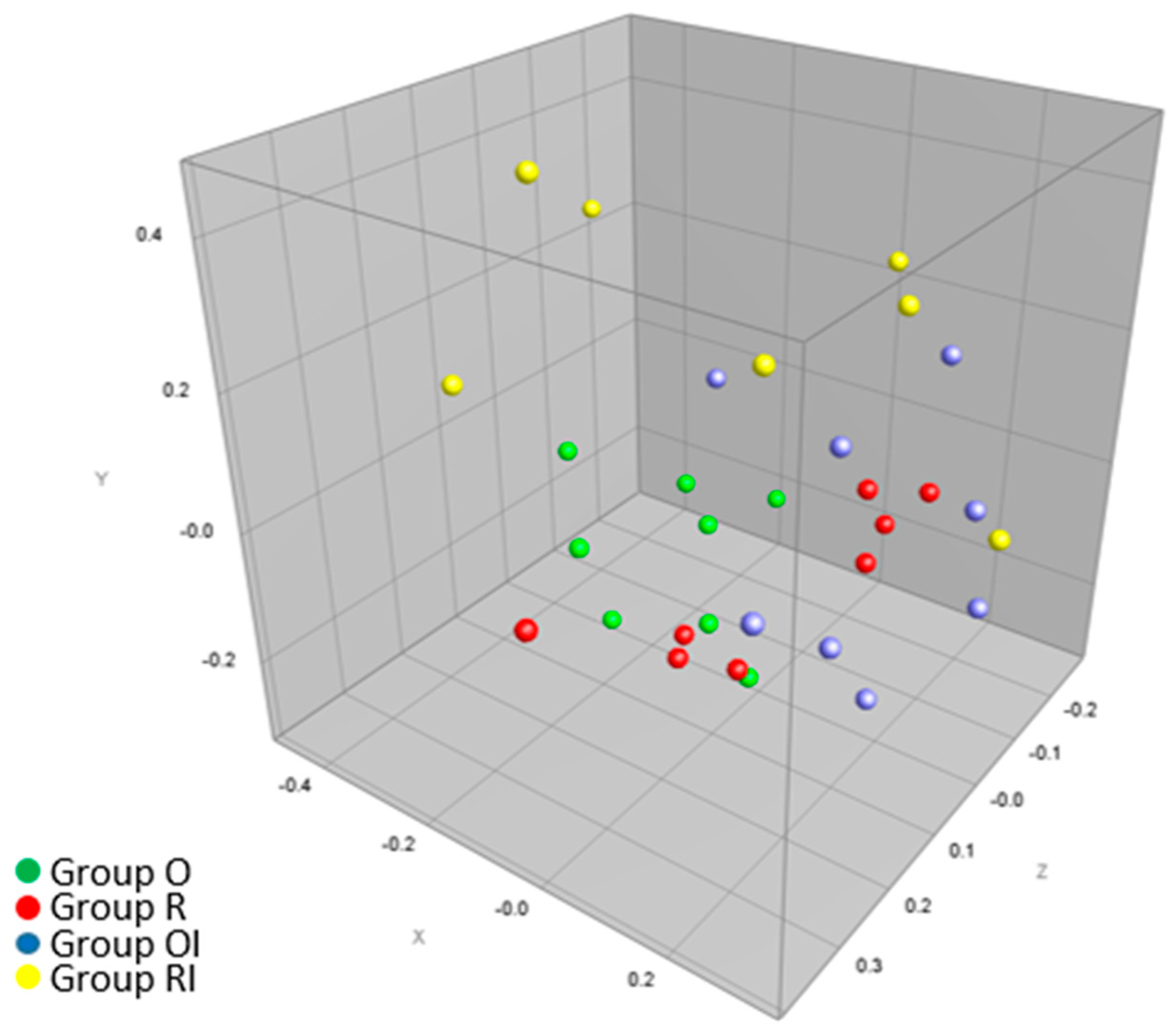
3.5. Effect of a Gluten-Free Diet Enriched with Inulin on the Qualitative Profile of Caecal Microbiota
3.6. Effect of a Gluten-Free Diet Enriched with Inulin on Intestinal Absorption and Retention of Calcium, Magnesium and Phosphorus
3.7. Effect of a Gluten-Free Diet Enriched with Inulin on Intestinal Absorption and Retention of Calcium, Magnesium and Phosphorus
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Khanal, R.C.; Nemere, I. Regulation of intestinal calcium transport. Ann. Rev. Nutr. 2008, 28, 179–196. [Google Scholar] [CrossRef] [PubMed]
- Krupa-Kozak, U.; Drabińska, N. Calcium in Gluten-Free Life: Health-Related and Nutritional Implications. Foods 2016, 5, E51. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.M.; Bachrach, L.K.; Carpenter, T.O.; Karsenty, G.; Rauch, F. Bone health in children and adolescents: A symposium at the annual meeting of the Pediatric Academic Societies/Lawson Wilkins Pediatric Endocrine Society, May 2003. Curr. Probl. Pediatr. Adolesc. Health Care 2004, 34, 226–242. [Google Scholar] [CrossRef] [PubMed]
- Green, P.H.; Rostami, K.; Marsh, M.N. Diagnosis of coeliac disease. Best Pract. Res. Clin. Gastroenterol. 2005, 19, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Krupa-Kozak, U. Pathologic bone alterations in celiac disease: Etiology, epidemiology, and treatment. Nutrition 2014, 30, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Lebwohl, B.; Michaëlsson, K.; Green, P.H.; Ludvigsson, J.F. Persistent mucosal damage and risk of fracture in celiac disease. J. Clin. Endocrinol. Metab. 2014, 99, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Vilppula, A.; Kaukinen, K.; Luostarinen, L.; Krekelä, I.; Patrikainen, H.; Valve, R.; Luostarinen, M.; Laurila, K.; Mäki, M.; Collin, P. Clinical benefit of gluten-free diet in screen-detected older celiac disease patients. BMC Gastroenterol. 2011, 11, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Margoni, D.; Chouliaras, G.; Duscas, G.; Voskaki, I.; Voutsas, N.; Papadopoulou, A.; Panayiotou, J.; Roma, E. Bone health in children with celiac disease assessed by dual x-ray absorptiometry: Effect of gluten-free diet and predictive value of serum biochemical indices. J. Pediatr. Gastroenterol. Nutr. 2012, 54, 680–684. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, S.J.; Gibson, P.R. Nutritional inadequacies of the gluten-free diet in both recently-diagnosed and long-term patients with coeliac disease. J. Hum. Nutr. Diet. 2013, 26, 349–358. [Google Scholar] [CrossRef] [PubMed]
- De Palma, G.; Nadal, I.; Collado, M.C.; Sanz, Y. Effects of a gluten-free diet on gut microbiota and immune function in healthy adult human subjects. Br. J. Nutr. 2009, 102, 1154–1160. [Google Scholar] [CrossRef] [PubMed]
- Mahadev, S.; Murray, J.A.; Wu, T.T.; Chandan, V.S.; Torbenson, M.S.; Kelly, C.P.; Maki, M.; Green, P.H.R.; Adelman, D.C.; Lebwohl, B. Factors associated with villus atrophy in symptomatic coeliac disease patients on a gluten-free diet. Aliment. Pharmacol. Ther. 2017, 45, 1084–1093. [Google Scholar] [CrossRef] [PubMed]
- Sdepanian, V.L.; de Miranda Carvalho, C.N.; de Morais, M.B.; Colugnati, F.A.B.; Fagundes-Neto, U. Bone mineral density of the lumbar spine in children and adolescent with celiac disease on a gluten free diet in Sao Paulo, Brazil. J. Pediatr. Gastroenterol. Nutr. 2003, 37, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Blazina, S.; Bratanic, N.; Campa, A.S.; Blagus, R.; Orel, R. Bone mineral density and importance of strict gluten-free diet in children and adolescents with celiac disease. Bone 2010, 47, 598–603. [Google Scholar] [CrossRef] [PubMed]
- Saturni, L.; Ferretti, G.; Bacchetti, T. The Gluten-Free Diet: Safety and Nutritional Quality. Nutrients 2010, 2, 16–34. [Google Scholar] [CrossRef] [PubMed]
- Theethira, T.G.; Dennis, M.; Leffler, D.A. Nutritional consequences of celiac disease and the gluten-free diet. Expert Rev. Gastroenterol. Hepatol. 2014, 8, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Cashman, K.D. Diet, nutrition and bone health. J. Nutr. 2007, 137, 2507S–2512S. [Google Scholar] [PubMed]
- Weaver, C.M.; Martin, B.R.; Nakatsu, C.H.; Armstrong, A.P.; Clavijo, A.; McCabe, L.D.; McCabe, G.P.; Duignan, S.; Schoterman, M.H.; van den Heuvel, E.G. Galactooligosaccharides improve mineral absorption and bone properties in growing rats through gut fermentation. J. Agric. Food Chem. 2011, 22, 6501–6510. [Google Scholar] [CrossRef] [PubMed]
- Bryk, G.; Coronel, M.Z.; Pellegrini, G.; Mandalunis, P.; Rio, M.E.; de Portela, M.L.; Zeni, S.N. Effect of a combination GOS/FOS® prebiotic mixture and interaction with calcium intake on mineral absorption and bone parameters in growing rats. Eur. J. Nutr. 2015, 54, 913–923. [Google Scholar] [CrossRef] [PubMed]
- Drabińska, N.; Krupa-Kozak, U.; Zieliński, H. Technological benefits of inulin-type fructans application in gluten-free products—A review. Trends Food Sci. Technol. 2016, 56, 149–157. [Google Scholar] [CrossRef]
- Rossi, M.; Corradini, C.; Amaretti, A.; Nicolini, M.; Pompei, A.; Zanoni, S.; Matteuzzi, D. Fermentation of Fructooligosaccharides and Inulin by Bifidobacteria: A Comparative Study of Pure and Fecal Cultures. Appl. Environ. Microbiol. 2005, 71, 6150–6158. [Google Scholar] [CrossRef] [PubMed]
- Ohta, A.; Ohtsuki, M.; Baba, S.; Adachi, T.; Sakata, T.; Sakaguchi, E. Calcium and magnesium absorption from the colon and rectum are increased in rats fed fructooligosaccharides. J. Nutr. 1995, 125, 2417–2424. [Google Scholar] [PubMed]
- Younes, H.; Coudray, C.; Bellanger, J.; Demigne, C.; Rayssiguier, Y.; Remesy, C. Effects of two fermentable carbohydrates (inulin and resistant starch) and their combination on calcium and magnesium balance in rats. Br. J. Nutr. 2001, 86, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Coudray, C.; Feillet-Coudray, C.; Tressol, J.C.; Gueux, E.; Thien, S.; Jaffrelo, L.; Mazur, A.; Rayssiguier, Y. Stimulatory effect of inulin on intestinal absorption of calcium and magnesium in rats is modulated by dietary calcium intakes—Short- and long-term balance studies. Eur. J. Nutr. 2005, 44, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Abrams, S.A.; Griffin, I.J.; Hawthorne, K.M.; Liang, L.; Gunn, S.K.; Darlington, G.; Ellis, K.J. A combination of prebiotic short- and long-chain inulin-type fructans enhances calcium absorption and bone mineralization in young adolescents. Am. J. Clin. Nutr. 2005, 82, 471–476. [Google Scholar] [PubMed]
- Tahiri, M.; Tressol, J.C.; Arnaud, Y.; Bornet, F.R.; Bouteloup-Demange, C.; Feillet-Coudray, C.; Brandolini, M.; Ducros, V.; Pépin, D.; Brouns, F.; et al. Effect of short chain fructooligosaccharides on intestinal Ca absorption and Ca status in postmenopausal women: A stable isotope study. Am. J. Clin. Nutr. 2003, 77, 449–457. [Google Scholar] [PubMed]
- Slevin, M.M.; Allsopp, P.J.; Magee, P.J.; Bonham, M.P.; Naughton, V.R.; Strain, J.J.; Duffy, M.E.; Wallace, J.M.; Mc Sorley, E.M. Supplementation with calcium and short-chain fructo-oligosaccharides affects markers of bone turnover but not bone mineral density in postmenopausal women. J. Nutr. 2014, 144, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Krupa-Kozak, U.; Troszyńska, A.; Bączek, N.; Soral-Śmietana, M. Effect of organic calcium supplements on the technological characteristic and sensory properties of gluten-free bread. Eur. Food Res. Technol. 2011, 232, 497–508. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO) Recommendations. Food Energy—Methods of Analysis and Conversion Factors; Food and Agriculture Organization of the United Nations (FAO) Recommendations: Rome, Italy, 2003; pp. 57–60. Available online: http://www.fao.org/uploads/media/FAO_2003_Food_Energy_02.pdf (accessed on 4 July 2017).
- Reeves, P.G. Components of the AIN-93 diets as improvements in the AIN-76A diet. J. Nutr. 1997, 127, 838–841. [Google Scholar]
- Close, B.; Banister, K.; Baumans, V.; Bernoth, E.M.; Bromage, N.; Bunyan, J.; Erhardt, W.; Flecknell, P.; Gregory, N.; Hackbarth, H.; et al. Recommendation for euthanasia of experimental animals: Part 2. Lab. Anim. 1997, 31, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Hofirek, B.; Haas, D. Comparative studies of ruminal fluid collected by oral tube or by puncture of the caudorental ruminal sac. Acta Vet. Brno 2001, 70, 27–33. [Google Scholar] [CrossRef]
- Krupa-Kozak, U.; Juśkiewicz, J.; Wronkowska, M.; Soral-Śmietana, M.; Zduńczyk, Z. Native and microwaved bean and pea starch preparations: Physiological effects on the intestinal ecosystem, caecal tissue and serum lipids in rats. Br. J. Nutr. 2010, 103, 1118–1126. [Google Scholar] [CrossRef] [PubMed]
- Markiewicz, L.H.; Honke, J.; Haros, M.; Świątecka, D.; Wróblewska, B. Diet shapes the ability of human intestinal microbiota to degrade phytate-in vitro studies. J. Appl. Microbiol. 2013, 115, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Fotschki, J.; Szyc, A.M.; Laparra, J.M.; Markiewicz, L.H.; Wróblewska, B. Immune-modulating properties of horse milk administered to mice sensitized to cow milk. J. Dairy Sci. 2016, 99, 9395–9404. [Google Scholar] [CrossRef] [PubMed]
- Hollants, J.; Decleyre, H.; Leliaert, F.; De Clerck, O.; Willems, A. Life without a cell membrane: Challenging the specificity of bacterial endophytes within Bryopsis (Bryopsidales, Chlorophyta). BMC Microbiol. 2011, 11, 255. [Google Scholar] [CrossRef] [PubMed]
- Van den Bogert, B.; Erkus, O.; Boekhorst, J.; de Goffau, M.; Smid, E.J.; Zoetendal, E.G.; Kleerebezem, M.F. Diversity of human small intestinal Streptococcus and Veillonella populations. EMS Microbiol. Ecol. 2013, 85, 376–388. [Google Scholar] [CrossRef] [PubMed]
- Van den Bogert, B.; Boekhorst, J.; Smid, E.J.; Zoetendal, E.G.; Kleerebezem, M. Draft Genome Sequence of Enterococcus sp. Strain HSIEG1, Isolated from the Human Small Intestine. Genome Announc. 2013, 12, e01013. [Google Scholar] [CrossRef] [PubMed]
- Roberfroid, M.B. Inulin-type fructans: Functional food ingredients. J. Nutr. 2007, 137, 2493–2502. [Google Scholar]
- Cummings, J.H.; Macfarlane, G.T. The control and consequences of bacterial fermentation in the human colon. J. Appl. Bacteriol. 1991, 70, 443–459. [Google Scholar] [CrossRef] [PubMed]
- Scholz-Ahrens, K.E.; Schrezenmeir, J. Inulin, oligofructose and mineral metabolism: Experimental data and mechanism. Br. J. Nutr. 2002, 87, 179–186. [Google Scholar] [CrossRef]
- Nyman, M. Fermentation and bulking capacity of indigestible carbohydrates: The case of inulin and oligofructose. Br. J. Nutr. 2002, 87, S163–S168. [Google Scholar] [CrossRef] [PubMed]
- Jurgoński, A.; Juśkiewicz, J.; Zduńczyk, Z. Comparative effects of different dietary levels of cellulose and fructooligosaccharides on fermentative processes in the caecum of rats. J. Anim. Feed Sci. 2008, 17, 88–99. [Google Scholar] [CrossRef]
- Shemarova, I.V.; Nesterov, V.P. Ca2+ signaling in prokaryotes. Microbiology 2014, 83, 431–437. [Google Scholar] [CrossRef]
- Onoda, T.; Enokizono, J.; Kaya, H.; Oshima, A.; Freestone, P.; Norris, V. Effects of calcium and calcium chelators on growth and morphology of Escherichia coli L-form Nc-7. J. Bacteriol. 2000, 182, 1419–1422. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.A.; Macfarlane, G.T. Enumeration of amino acid fermenting bacteria in the human large intestine: Effects of pH and starch on peptide metabolism and dissimilation of amino acids. FEMS Microbiol. Ecol. 1998, 25, 355–368. [Google Scholar] [CrossRef]
- Lin, H.C.; Visek, W.J. Colon mucosal cell damage by ammonia in rats. J. Nutr. 1991, 121, 887–893. [Google Scholar] [PubMed]
- Nilsson, U.; Nyman, M. Short-chain fatty acid formation in the hindgut of rats fed oligosaccharides varying in monomeric composition, degree of polymerisation and solubility. Br. J. Nutr. 2005, 94, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Salonen, A.; Lahti, L.; Salojarvi, J.; Holtrop, G.; Korpela, K.; Duncan, S.H.; Date, P.; Farquharson, F.; Johnstone, A.M.; Lobley, G.E.; et al. Impact of diet and individual variation on intestinal microbiota composition and fermentation products in obese men. ISME J. 2014, 8, 2218–2230. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Duncan, S.H.; Mccrae, S.I.; Millar, J.; Jackson, M.S.; Flint, H.J. Restricted distribution of the butyrate kinase pathway among butyrate-producing bacteria from the human colon. J. Bacteriol. 2004, 186, 2099–2106. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, D.C.; Guragain, M.; Patrauchan, M. Calcium binding proteins and calcium signaling in prokaryotes. Cell Calcium 2015, 57, 151–165. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Duncan, S.H.; Scott, K.P.; Louis, P. Links between diet, gut microbiota composition and gut metabolism. Proc. Nutr. Soc. 2015, 74, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.; Reusch, R.N. Poly-3-hydroxybutyrate synthase from the periplasm of Escherichia coli. Biochem. Biophys. Res. Commun. 2008, 374, 485–489. [Google Scholar] [CrossRef] [PubMed]
- Reusch, R.N. Transmembrane ion transport by polyphosphate/poly-(R)-3-hydroxybutyrate complexes. Biochemistry (Moscow) 2000, 65, 280–295. [Google Scholar]
- Topping, D.L.; Clifton, P.M. Short-chain fatty acids and human colonic function: Roles of resistant starch and nonstarch polysaccharides. Physiol. Rev. 2001, 81, 1031–1064. [Google Scholar] [PubMed]
- Machiels, K.; Joossens, M.; Sabino, J.; DePreter, V.; Arijs, I.; Eeckhaut, V.; Ballet, V.; Claes, K.; Van Immerseel, F.; Verbeke, K.; et al. A decrease of the butyrate-producing species Roseburia hominis and Faecali bacterium prausnitzii defines dysbiosis in patients with ulcerative colitis. Gut 2014, 63, 1275–1283. [Google Scholar] [CrossRef] [PubMed]
- Montesi, A.; García-Albiach, R.; Pozuelo, M.J.; Pintado, C.; Goñi, I.; Rotger, R. Molecular and microbiological analysis of caecal microbiota in rats fed with diets supplemented either with prebiotics or probiotics. Int. J. Food Microbiol. 2005, 98, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Dziarski, R.; Park, S.Y.; Kashyap, D.R.; Dowd, S.E.; Gupta, D. Pglyrp-Regulated Gut Microflora Prevotella falsenii, Parabacteroides distasonis and Bacteroides eggerthii Enhance and Alistipes finegoldii Attenuates Colitis in Mice. PLoS ONE 2016, 11, e0146162. [Google Scholar] [CrossRef] [PubMed]
- Juśkiewicz, J.; Wróblewska, M.; Jarosławska, J.; Zduńczyk, Z. Effects of inulin supplemented to cellulose-free or cellulose-rich diets on caecal environment and biochemical blood parameters in rats. J. Anim. Feed Sci. 2009, 18, 709–722. [Google Scholar] [CrossRef]
- Roberton, A.M.; Lee, S.P.; Lindop, R.; Stanley, R.A.; Thomsen, L.; Tasman-Jones, C. Biliary control of beta-glucuronidase activity in the luminal contents of the rat ileum, cecum, and rectum. Cancer Res. 1982, 42, 5165–5166. [Google Scholar] [PubMed]
- Gråsten, S.M.; Pajarib, A.M.; Liukkonenc, K.H.; Karppinenc, S.; Mykkänena, H.M. Fibers with different solubility characteristics alter similarly the metabolic activity of intestinal microbiota in rats fed cereal brans and inulin. Nutr. Res. 2002, 12, 1435–1444. [Google Scholar] [CrossRef]
- Scholz-Ahrens, K.E.; Adolphi, B.; Rochat, F.; Barclay, D.V.; de Vrese, M.; Açil, Y.; Schrezenmeir, J. Effects of probiotics, prebiotics, and synbiotics on mineral metabolism in ovariectomized rats—Impact of bacterial mass, intestinal absorptive area and reduction of bone turn-over. NFS J. 2016, 3, 41–50. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| O | R | OI | RI | |
|---|---|---|---|---|
| Bread ingredients as diet compounds (%) | ||||
| Corn starch | 48.4 | 49.8 | 40.4 | 41.8 |
| Potato starch | 11.7 | 11.7 | 11.7 | 11.7 |
| Pectin | 3.0 | 3.0 | 3.0 | 3.0 |
| Sugar | 3.7 | 3.7 | 3.7 | 3.7 |
| Salt | 1.0 | 1.0 | 1.0 | 1.0 |
| Sunflower oil | 1.7 | 1.7 | 1.7 | 1.7 |
| Yeast | 3.7 | 3.7 | 3.7 | 3.7 |
| CAS * | 12.0 | 12.0 | 12.0 | 12.0 |
| CIT & | 2.0 | 0.6 | 2.0 | 0.6 |
| Inulin # | 0.0 | 0.0 | 8.0 | 8.0 |
| Other diet compounds (%) | ||||
| dl-Methionine | 0.3 | 0.3 | 0.3 | 0.3 |
| Soya oil | 8.0 | 8.0 | 8.0 | 8.0 |
| Ca-free mineral mix † | 3.5 | 3.5 | 3.5 | 3.5 |
| Vitamin mix § | 1.0 | 1.0 | 1.0 | 1.0 |
| Energy density (kcal/g) | 4.01 | 4.00 | 3.82 | 3.80 |
| Diets | Ca Effect | Inulin Effect | Ca × I | ||||
|---|---|---|---|---|---|---|---|
| O | R | OI | RI | ||||
| Daily food intake (g/animal) | 13.90 ± 1.22 | 13.15 ± 1.56 | 13.83 ± 0.54 | 13.26 ± 0.95 | 0.0688 | 0.7049 | 0.9054 |
| Daily BW gain (g/animal) | 4.55 ± 0.45 | 4.61 ± 0.53 | 4.42 ± 0.30 | 4.38 ± 0.34 | 0.9477 | 0.2312 | 0.7594 |
| Small intestinal parameters | |||||||
| Weight (g/100 g BW) | 3.10 ± 0.13 | 2.86 ± 0.19 | 3.26 ± 0.32 | 3.19 ± 0.34 | 0.0983 | 0.0139 | 0.3633 |
| pH of small intestinal digesta | 7.03 ± 0.22 | 7.02 ± 0.18 | 6.86 ± 0.20 | 6.67 ± 0.24 | 0.1991 | 0.0015 | 0.2282 |
| Caecum parameters | |||||||
| Weight of tissue (g/100 g BW) | 0.320 ± 0.03 c | 0.332 ± 0.03 c | 0.436 ± 0.04 b | 0.587 ± 0.08 a | <0.001 | <0.001 | <0.001 |
| Weight of digesta (g/100 g BW) | 1.124 ± 0.198 b | 1.112 ± 0.202 b | 1.441 ± 0.240 b | 3.110 ± 0.694 a | <0.001 | <0.001 | <0.001 |
| DM content of digesta (%) | 16.79 ± 1.34 | 15.65 ± 1.42 | 15.50 ± 1.33 | 14.79 ± 1.40 | 0.0671 | 0.0346 | 0.6537 |
| Ammonia (mg/g digesta) | 0.191 ± 0.026 | 0.183 ± 0.018 | 0.141 ± 0.016 | 0.154 ± 0.022 | 0.7069 | <0.001 | 0.1704 |
| pH of digesta | 6.79 ± 0.13 | 6.53 ± 0.18 | 6.18 ± 0.24 | 5.83 ± 0.39 | 0.0019 | <0.001 | 0.5949 |
| Colonic parameters | |||||||
| Weight of tissue (g/100 g BW) | 0.531 ± 0.072 a,b | 0.467 ± 0.042 b | 0.535 ± 0.055 a,b | 0.621 ± 0.094 a | 0.6517 | 0.0028 | 0.0044 |
| Weight of digesta (g/100 g BW) | 0.519 ± 0.108 b | 0.389 ± 0.153 c | 0.570 ± 0.095 a,b | 0.658 ± 0.078 a | 0.5933 | <0.001 | 0.0104 |
| pH of digesta | 6.71 ± 0.186 | 6.11 ± 0.178 | 6.11 ± 0.168 | 5.66 ± 0.132 | <0.001 | <0.001 | 0.2046 |
| Diets | Ca Effect | Inulin Effect | Ca × I | ||||
|---|---|---|---|---|---|---|---|
| O | R | OI | RI | ||||
| SCFAs (μM/g digesta) | |||||||
| Acetic | 55.17 ± 7.19 b | 53.22 ± 9.46 b | 58.49 ± 8.56 a | 35.14 ± 8.94 c | <0.001 | 0.0216 | 0.0015 |
| Propionic | 16.30 ± 1.39 c | 28.51 ± 6.15 b | 39.91 ± 7.65 a | 27.57 ± 7.48 b | 0.9768 | <0.001 | <0.001 |
| Iso-butyric | 0.41 ± 0.22 | 0.17 ± 0.07 | 0.29 ± 0.13 | 0.05 ± 0.03 | <0.001 | 0.0187 | 0.9336 |
| Butyric | 6.09 ± 2.02 b | 5.68 ± 0.72 b | 8.92 ± 2.79 a | 2.31 ± 1.64 c | <0.001 | 0.6953 | <0.001 |
| Iso-valeric | 0.70 ± 0.21 | 0.44 ± 0.17 | 0.25 ± 0.13 | 0.14 ± 0.06 | 0.0016 | <0.001 | 0.1738 |
| Valeric | 1.28 ± 0.24 | 0.67 ± 0.14 | 0.64 ± 0.18 | 0.05 ± 0.02 | <0.001 | <0.001 | 0.5737 |
| PSCFAs | 2.39 ± 0.50 | 1.28 ± 0.21 | 1.19 ± 0.29 | 0.23 ± 0.08 | <0.001 | <0.001 | 0.4801 |
| Total SCFAs | 79.95 ± 9.43 b | 88.69 ± 13.98 b | 108.50 ± 13.95 a | 65.31 ± 17.43 c | 0.0017 | 0.6097 | <0.001 |
| C2:C3:C4 profile (%) | |||||||
| C2 | 69 a | 60 b | 54 c | 54 c | <0.001 | <0.001 | <0.001 |
| C3 | 21 c | 32 b | 37 b | 42 a | <0.001 | <0.001 | 0.0070 |
| C4 | 8 a | 6 b | 8 a | 4 c | <0.001 | 0.1097 | 0.0086 |
| Diets | Ca Effect | Inulin Effect | Ca × I | ||||
|---|---|---|---|---|---|---|---|
| O | R | OI | RI | ||||
| α-Glucosidase (μmol/h/g) | 13.86 ± 7.28 | 31.80 ± 13.76 | 29.74 ± 10.55 | 40.72 ± 10.99 | <0.001 | 0.0032 | 0.3737 |
| β-Glucosidase (μmol/h/g) | 4.94 ± 1.49 b | 7.53 ± 1.30 a | 6.90 ± 1.74 a | 3.82 ± 0.71 b | 0.6205 | 0.0803 | <0.001 |
| α-Galactosidase (μmol/h/g) | 7.86 ± 1.71 | 4.48 ± 0.92 | 8.22 ± 3.58 | 7.84 ± 4.92 | 0.1066 | 0.1096 | 0.1939 |
| β-Galactosidase (μmol/h/g) | 27.37 ± 8.85 | 44.19 ± 10.08 | 48.99 ± 19.22 | 56.56 ± 19.59 | 0.0319 | 0.0039 | 0.3995 |
| β-Glucuronidase (μmol/h/g) | 18.82 ± 6.14 a | 4.77 ± 1.98 b | 5.67 ± 2.71 b | 1.85 ± 0.98 c | <0.001 | <0.001 | <0.001 |
| Diets | Ca Effect | Inulin Effect | Ca × I | ||||
|---|---|---|---|---|---|---|---|
| O | R | OI | RI | ||||
| Calcium | |||||||
| Intake (mg/day) | 86.21 ± 7.87 a | 35.64 ± 4.24 b | 80.46 ± 5.37 a | 33.95 ± 2.44 b | <0.001 | 0.0024 | 0.0220 |
| Faecal excretion (mg/day) | 22.68 ± 4.08 | 2.01 ± 0.43 | 20.05 ± 4.43 | 0.84 ± 0.32 | <0.001 | 0.0232 | 0.2026 |
| Absorption (%) † | 73.28 ± 3.13 | 94.36 ± 0.80 | 74.60 ± 4.86 | 97.51 ± 0.92 | <0.001 | 0.0414 | 0.3879 |
| Urinary excretion (mg/day) | 6.92 ± 1.98 | 0.54 ± 0.15 | 9.44 ± 2.98 | 0.52 ± 0.18 | <0.001 | 0.0179 | 0.0673 |
| Retention (%) ‡ | 65.63 ± 2.23 | 93.05 ± 1.84 | 62.62 ± 6.18 | 94.18 ± 3.80 | <0.001 | 0.5916 | 0.0753 |
| Magnesium | |||||||
| Intake (mg/day) | 6.35 ± 0.56 | 6.30 ± 0.75 | 6.46 ± 0.25 | 6.21 ± 0.43 | 0.1695 | 0.9289 | 0.2754 |
| Faecal excretion (mg/day) | 1.72 ± 0.37 | 0.42 ± 0.07 | 1.48 ± 0.19 | 0.32 ± 0.06 | <0.001 | 0.0173 | 0.3777 |
| Absorption (%) † | 72.94 ± 5.06 | 93.28 ± 0.99 | 76.54 ± 1.50 | 95.21 ± 1.26 | <0.001 | 0.0078 | 0.3920 |
| Urinary excretion (mg/day) | 3.03 ± 0.24 | 3.11 ± 0.41 | 3.25 ± 0.28 | 3.27 ± 0.47 | 0.4112 | 0.0321 | 0.2432 |
| Retention (%) ‡ | 28.54 ± 8.57 | 43.84 ± 3.30 | 28.51 ± 4.04 | 43.68 ± 5.07 | <0.001 | 0.4611 | 0.7504 |
| Phosphorus | |||||||
| Intake (mg/day) | 35.26 ± 3.10 | 34.74 ± 4.13 | 35.44 ± 2.42 | 32.79 ± 2.35 | 0.1515 | 0.4407 | 0.3269 |
| Faecal excretion (mg/day) | 13.19 ± 1.54 | 4.68 ± 0.73 | 12.62 ± 1.83 | 4.57 ± 0.24 | <0.001 | 0.0517 | 0.9248 |
| Absorption (%) † | 62.66 ± 2.79 | 86.42 ± 2.25 | 63.23 ± 1.05 | 88.90 ± 3.48 | <0.001 | 0.0994 | 0.2963 |
| Urinary excretion (mg/day) | 0.37 ± 0.07 | 9.37 ± 1.39 | 0.37 ± 0.15 | 8.91 ± 0.74 | <0.001 | 0.2644 | 0.2606 |
| Retention (%) ‡ | 61.60 ± 2.79 | 60.21 ± 3.01 | 63.62 ± 4.13 | 62.30 ± 8.00 | 0.0927 | 0.0314 | 0.7850 |
| Diets | Ca Effect | Inulin Effect | Ca × I | ||||
|---|---|---|---|---|---|---|---|
| O | R | OI | RI | ||||
| Calcium (mM/L) | 2.522 ± 0.084 | 2.337 ± 0.059 | 2.545 ± 0.204 | 2.537 ± 0.238 | 0.1083 | 0.0662 | 0.1393 |
| Phosphorus (mM/L) | 1.861 ± 0.247 b | 2.202 ± 0.166 a | 1.835 ± 0.211 b | 2.670 ± 0.215 a | <0.001 | 0.0063 | 0.0026 |
| Ca:P | 1.4 | 1.1 | 1.4 | 1.0 | <0.001 | 0.3116 | 0.1177 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krupa-Kozak, U.; Markiewicz, L.H.; Lamparski, G.; Juśkiewicz, J. Administration of Inulin-Supplemented Gluten-Free Diet Modified Calcium Absorption and Caecal Microbiota in Rats in a Calcium-Dependent Manner. Nutrients 2017, 9, 702. https://doi.org/10.3390/nu9070702
Krupa-Kozak U, Markiewicz LH, Lamparski G, Juśkiewicz J. Administration of Inulin-Supplemented Gluten-Free Diet Modified Calcium Absorption and Caecal Microbiota in Rats in a Calcium-Dependent Manner. Nutrients. 2017; 9(7):702. https://doi.org/10.3390/nu9070702
Chicago/Turabian StyleKrupa-Kozak, Urszula, Lidia H. Markiewicz, Grzegorz Lamparski, and Jerzy Juśkiewicz. 2017. "Administration of Inulin-Supplemented Gluten-Free Diet Modified Calcium Absorption and Caecal Microbiota in Rats in a Calcium-Dependent Manner" Nutrients 9, no. 7: 702. https://doi.org/10.3390/nu9070702
APA StyleKrupa-Kozak, U., Markiewicz, L. H., Lamparski, G., & Juśkiewicz, J. (2017). Administration of Inulin-Supplemented Gluten-Free Diet Modified Calcium Absorption and Caecal Microbiota in Rats in a Calcium-Dependent Manner. Nutrients, 9(7), 702. https://doi.org/10.3390/nu9070702