Dose-Dependent Prebiotic Effect of Lactulose in a Computer-Controlled In Vitro Model of the Human Large Intestine

Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Product
2.2. Intestinal Conditions of the TIM-2 System
2.3. Addition of Test Product
2.4. Sampling from TIM-2
2.5. Sodium Hydroxide Usage (pH)
2.6. Short-Chain Fatty Acids and Branched-Chain Fatty Acids
2.7. Lactate and Ammonia
2.8. 16S rDNA Amplicon Sequencing
2.9. Statistical Analysis
3. Results
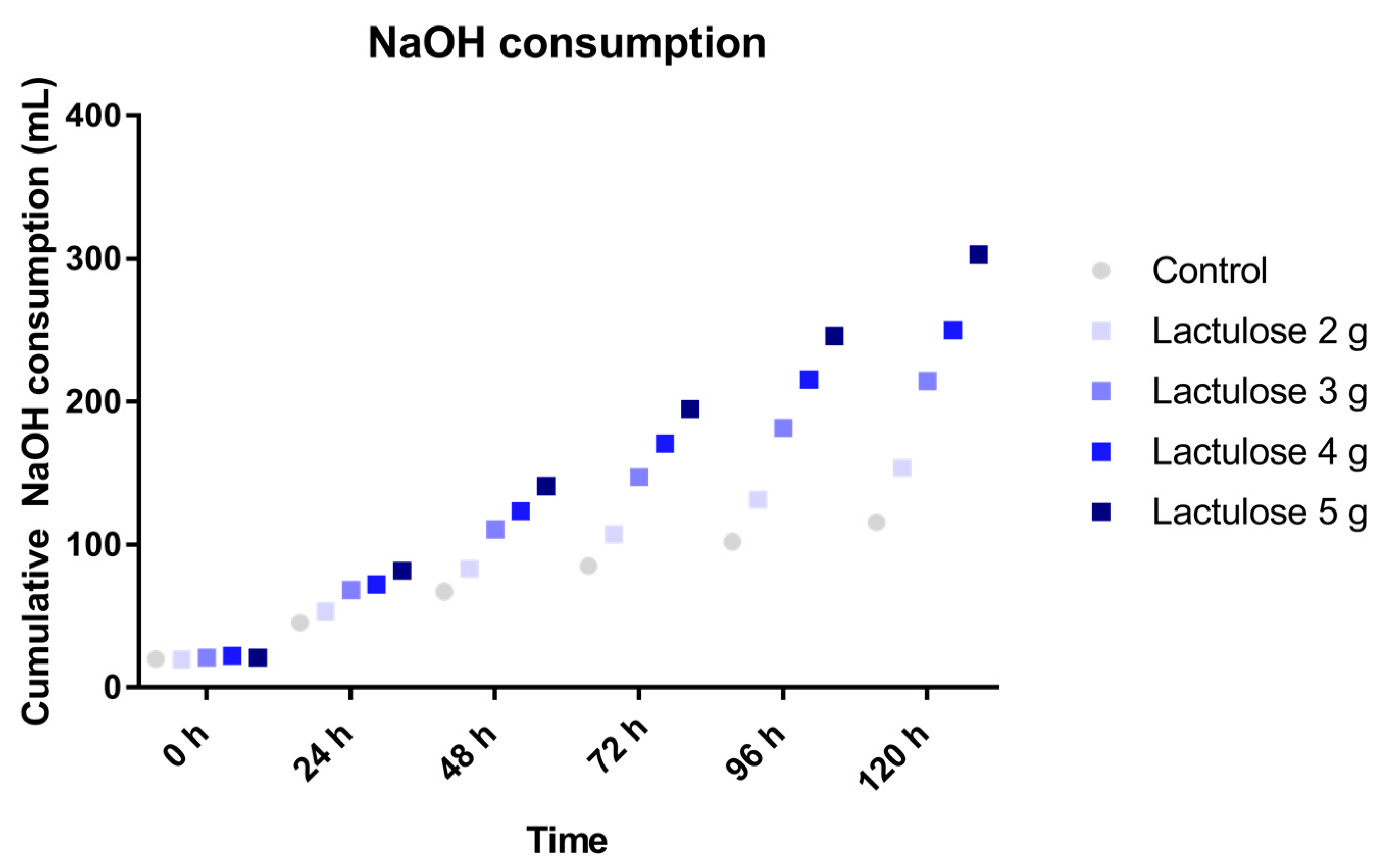
3.1. Sodium Hydroxide Usage
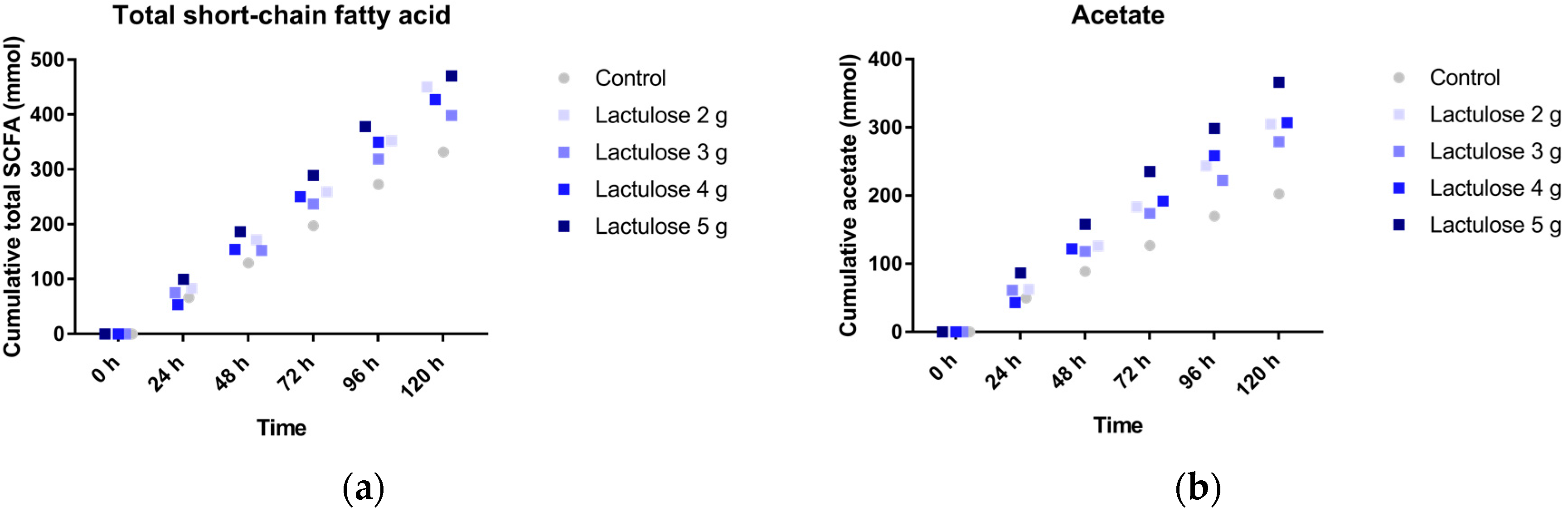
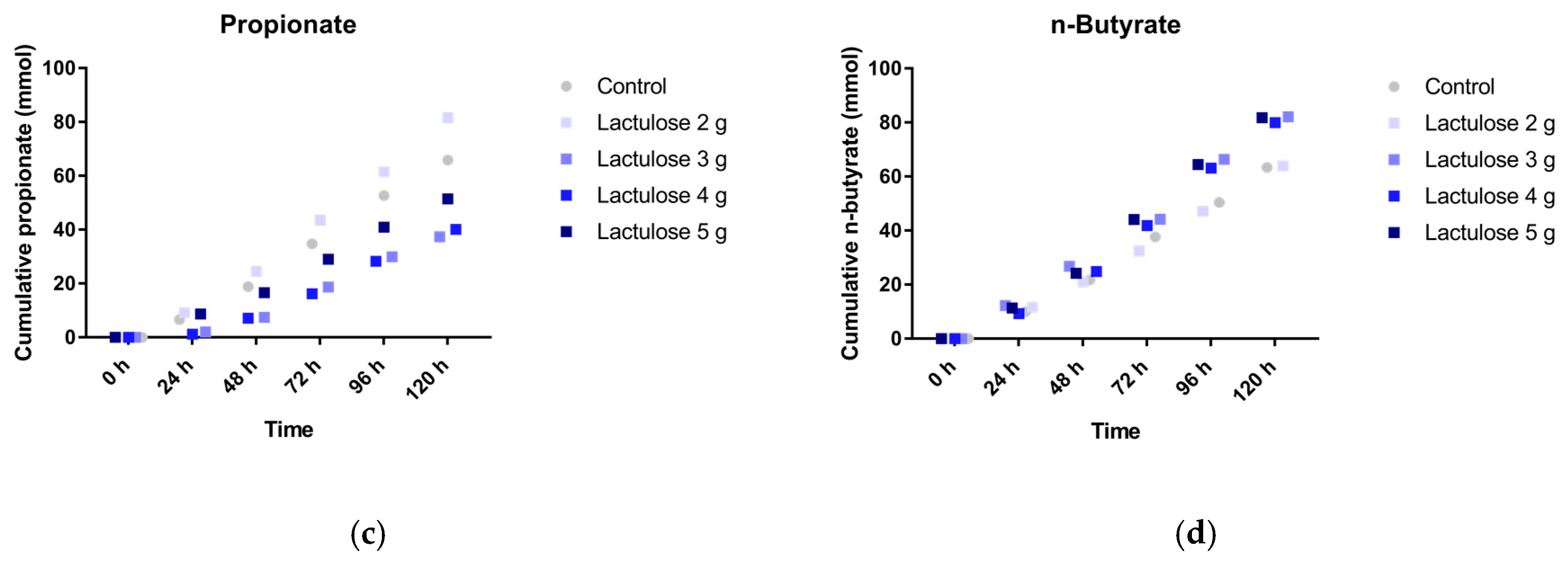
3.2. SCFA Production
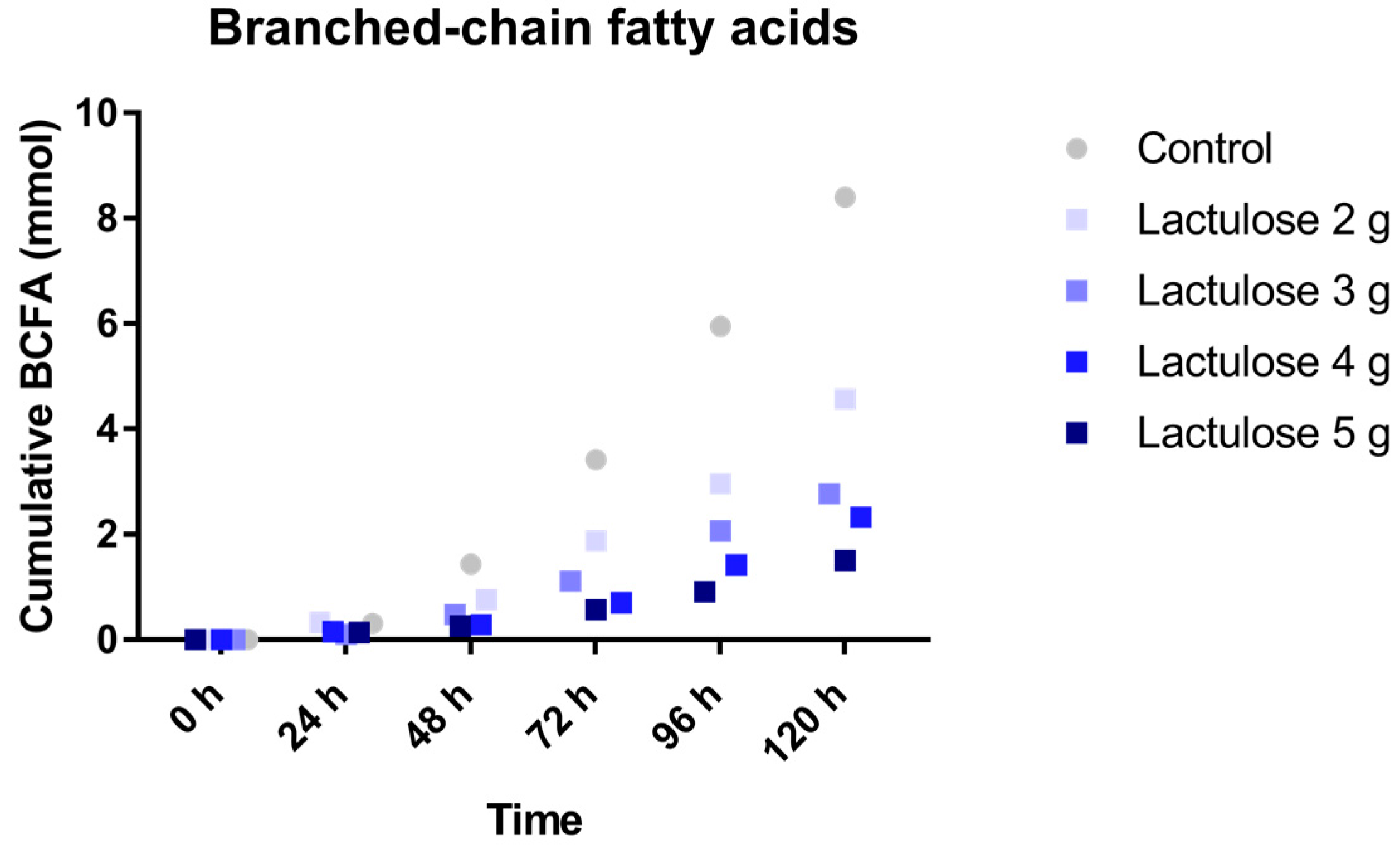
3.3. BCFA Production
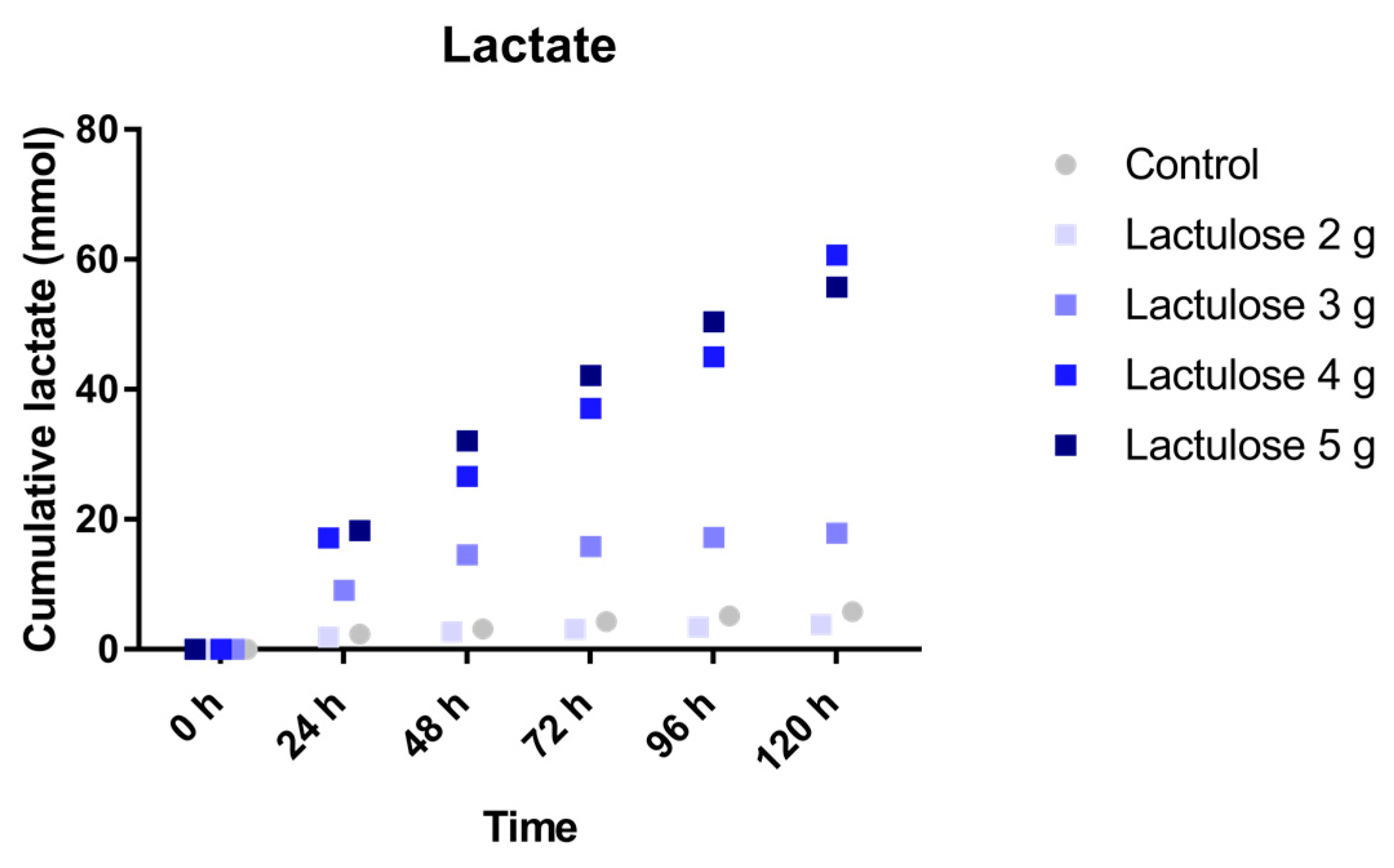
3.4. Lactate
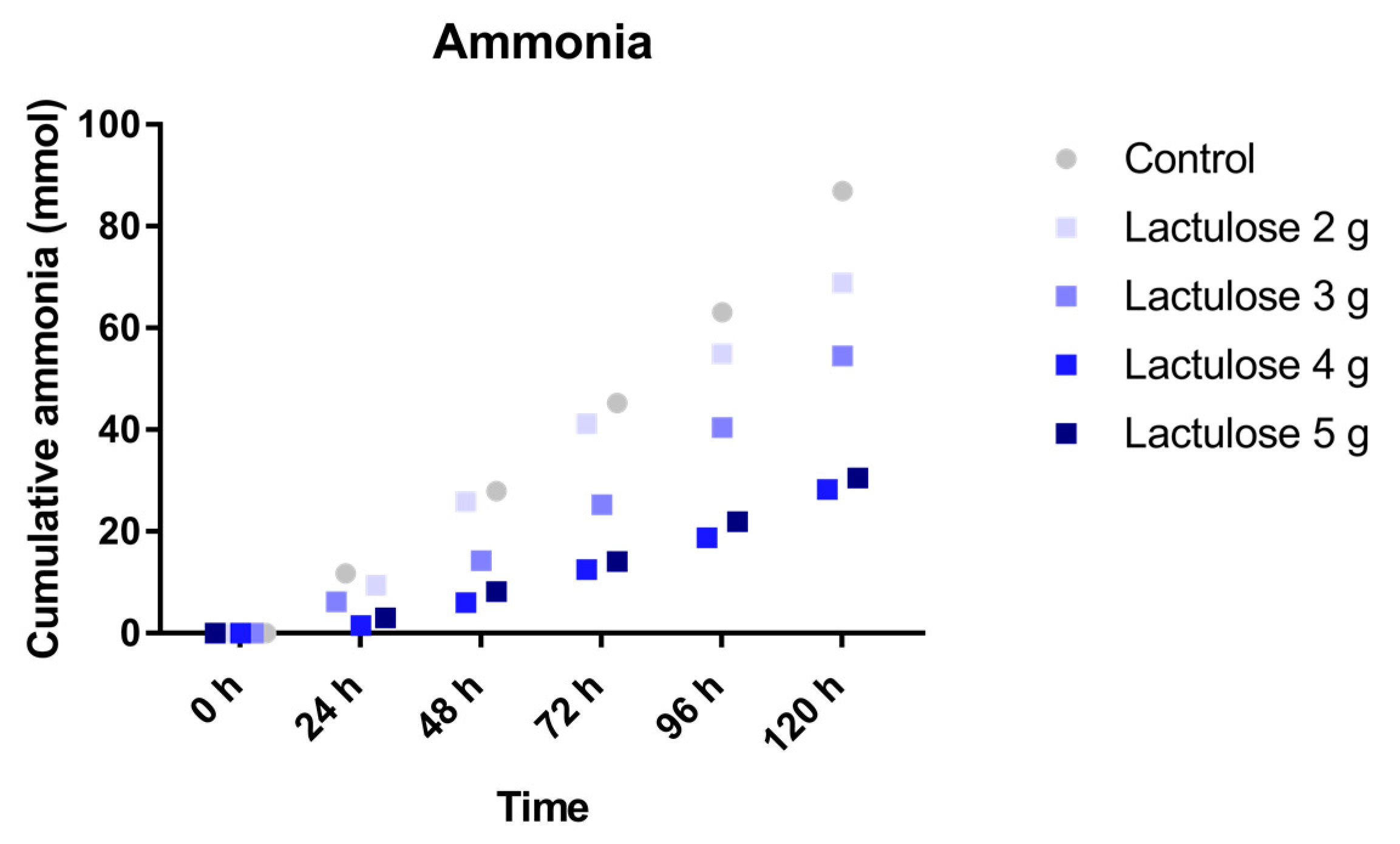
3.5. Ammonia
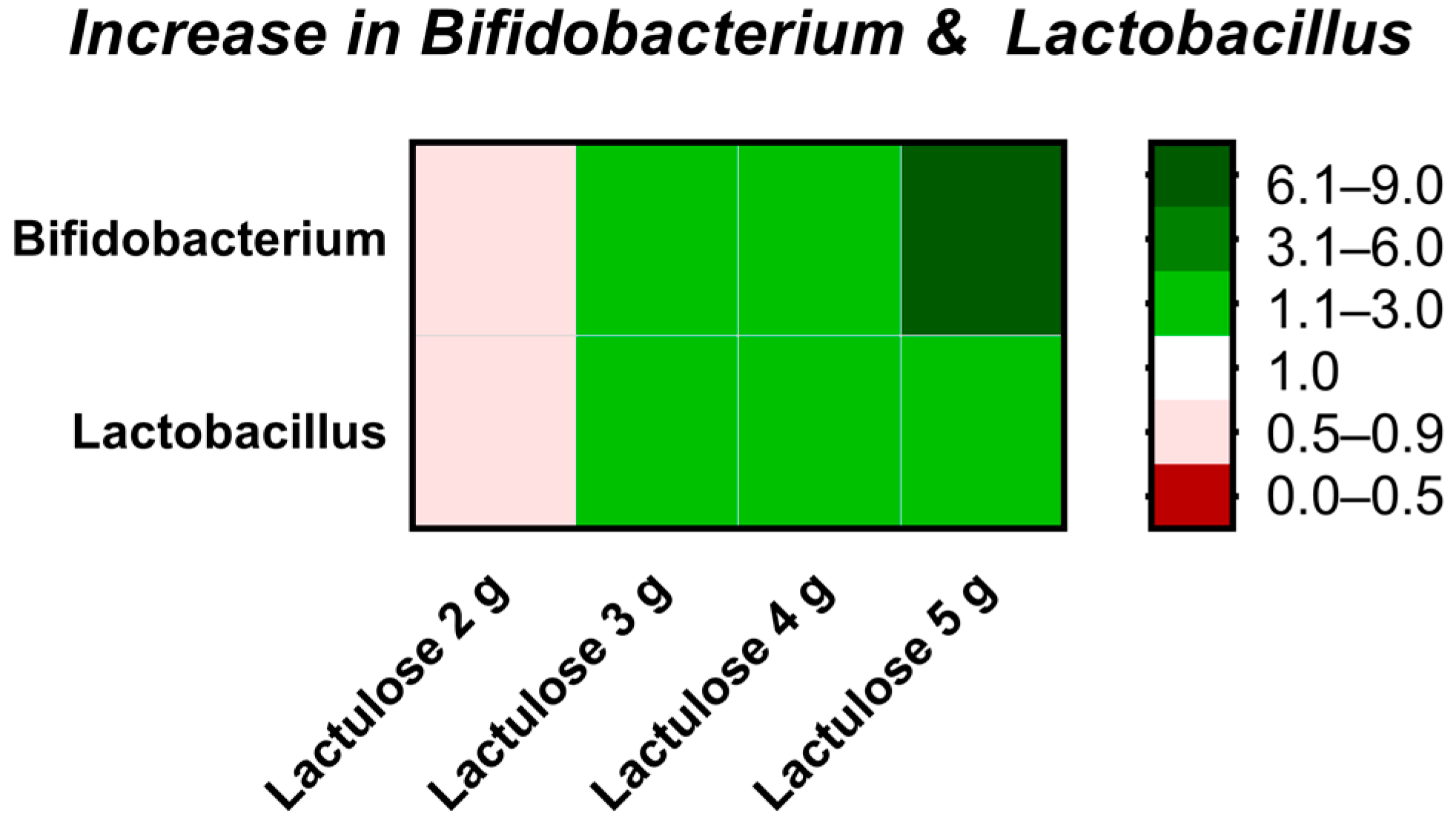
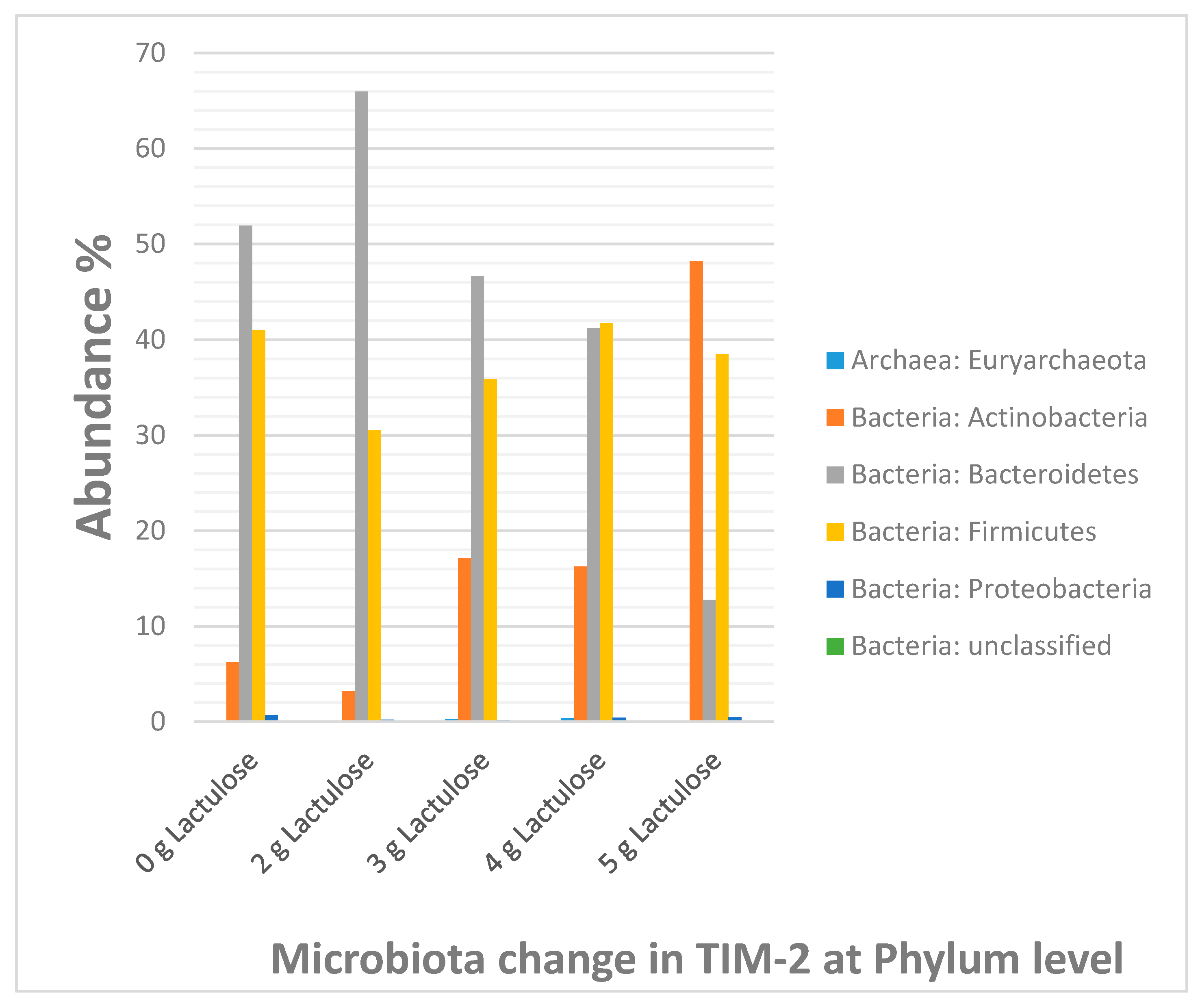
3.6. Microbiota Composition
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Aguirre, M.; Jonkers, D.M.; Troost, F.J.; Roeselers, G.; Venema, K. In vitro characterization of the impact of different substrates on metabolite production, energy extraction and composition of gut microbiota from lean and obese subjects. PLoS ONE 2014, 9, e113864. [Google Scholar] [CrossRef] [PubMed]
- Venema, K.; van Nuenen, M.; van den Heuvel, E.; Pool, W.; van der Vossen, J. The effect of lactulose on the composition of the intestinal microbiota and short-chain fatty acid production in human volunteers and a computer-controlled model of the proximal large intestine. Microb. Ecol. Health Dis. 2003, 15, 94–105. [Google Scholar] [CrossRef]
- Bouhnik, Y.; Attar, A.; Joly, F.A.; Riottot, M.; Dyard, F.; Flourie, B. Lactulose ingestion increases faecal bifidobacterial counts: A randomised double-blind study in healthy humans. Eur. J. Clin. Nutr. 2004, 58, 462–466. [Google Scholar] [CrossRef] [PubMed]
- Mizota, T.; Mori, T.; Yaeshima, T.; Yanagida, T.; Iwatsuki, K.; Ishibashi, N.; Tamura, Y.; Fukuwatari, Y. Effects of low dosages of lactulose on the intestinal function of healthy adults. Milchwissenschaft 2002, 57, 312–315. [Google Scholar]
- Terada, A.; Hara, H.; Kataoka, M.; Mitsuoka, T. Effects of lactulose on the composition and metabolic activity of human fecal microflora. Microb. Ecol. Health Dis. 1992, 5, 43–50. [Google Scholar] [CrossRef]
- Topping, D.L.; Clifton, P.M. Short-chain fatty acids and human colonic function: Roles of resistant starch and nonstarch polysaccharides. Physiol. Rev. 2001, 81, 1031–1064. [Google Scholar] [PubMed]
- Maathuis, A.; Hoffman, A.; Evans, A.; Sanders, L.; Venema, K. The effect of the undigested fraction of maize products on the activity and composition of the microbiota determined in a dynamic in vitro model of the human proximal large intestine. J. Am. Coll. Nutr. 2009, 28, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Minekus, M.; Smeets-Peeters, M.; Bernalier, A.; Marol-Bonnin, S.; Havenaar, R.; Marteau, P.; Alric, M.; Fonty, G.; Huis in’t Veld, J.H. A computer-controlled system to simulate conditions of the large intestine with peristaltic mixing, water absorption and absorption of fermentation products. Appl. Microbiol. Biotechnol. 1999, 53, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Maathuis, A.J.; van den Heuvel, E.G.; Schoterman, M.H.; Venema, K. Galacto-oligosaccharides have prebiotic activity in a dynamic in vitro colon model using a (13)c-labeling technique. J. Nutr. 2012, 142, 1205–1212. [Google Scholar] [CrossRef] [PubMed]
- Van Nuenen, M.H.; Meyer, P.; Venema, K. The effect of various inulins and clostridium difficile on the metabolic activity of the human colonic microbiota in vitro. Microb. Ecol. Health Dis. 2003, 15, 137–144. [Google Scholar] [CrossRef]
- Sims, G.; Ellsworth, T.; Mulvaney, R. Microscale determination of inorganic nitrogen in water and soil extracts. Commun. Soil Sci. Plant Anal. 1995, 26, 303–316. [Google Scholar] [CrossRef]
- Ladirat, S.E.; Schols, H.A.; Nauta, A.; Schoterman, M.H.; Keijser, B.J.; Montijn, R.C.; Gruppen, H.; Schuren, F.H. High-throughput analysis of the impact of antibiotics on the human intestinal microbiota composition. J. Microbiol. Methods 2013, 92, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the miseq illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16s rrna diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4516–4522. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Ballongue, J.; Schumann, C.; Quignon, P. Effects of lactulose and lactitol on colonic microflora and enzymatic activity. Scand. J. Gastroenterol. Suppl. 1997, 222, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Schwiertz, A.; Hold, G.L.; Duncan, S.H.; Gruhl, B.; Collins, M.D.; Lawson, P.A.; Flint, H.J.; Blaut, M. Anaerostipes caccae gen. Nov., sp. Nov., a new saccharolytic, acetate-utilising, butyrate-producing bacterium from human faeces. Syst. Appl. Microbiol. 2002, 25, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Bourriaud, C.; Robins, R.J.; Martin, L.; Kozlowski, F.; Tenailleau, E.; Cherbut, C.; Michel, C. Lactate is mainly fermented to butyrate by human intestinal microfloras but inter-individual variation is evident. J. Appl. Microbiol. 2005, 99, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Bouhnik, Y.; Neut, C.; Raskine, L.; Michel, C.; Riottot, M.; Andrieux, C.; Guillemot, F.; Dyard, F.; Flourie, B. Prospective, randomized, parallel-group trial to evaluate the effects of lactulose and polyethylene glycol-4000 on colonic flora in chronic idiopathic constipation. Aliment. Pharmacol. Ther. 2004, 19, 889–899. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Long, W.; Zhang, C.; Liu, S.; Zhao, L.; Hamaker, B.R. Fiber-utilizing capacity varies in prevotella- versus bacteroides-dominated gut microbiota. Sci. Rep. 2017, 7, 2594. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Checa, F.; Bejar, V.; Martinez-Canovas, M.J.; Llamas, I.; Quesada, E. Halomonas almeriensis sp. Nov., a moderately halophilic, exopolysaccharide-producing bacterium from cabo de gata, almeria, south-east Spain. Int. J. Syst. Evol. Microbiol. 2005, 55, 2007–2011. [Google Scholar] [CrossRef] [PubMed]
- Duncan, S.H.; Louis, P.; Flint, H.J. Lactate-utilizing bacteria, isolated from human feces, that produce butyrate as a major fermentation product. Appl. Environ. Microbiol. 2004, 70, 5810–5817. [Google Scholar] [CrossRef] [PubMed]
- Brutinel, E.D.; Gralnick, J.A. Preferential utilization of d-lactate by shewanella oneidensis. Appl. Environ. Microbiol. 2012, 78, 8474–8476. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.M.; Taylor, J.S.; Latour, D.J.; Iuchi, S.; Lin, E.C. Three overlapping lct genes involved in l-lactate utilization by escherichia coli. J. Bacteriol. 1993, 175, 6671–6678. [Google Scholar] [CrossRef] [PubMed]
- Reimer, R.A.; Maathuis, A.J.; Venema, K.; Lyon, M.R.; Gahler, R.J.; Wood, S. Effect of the novel polysaccharide polyglycoplex(r) on short-chain fatty acid production in a computer-controlled in vitro model of the human large intestine. Nutrients 2014, 6, 1115–1127. [Google Scholar] [CrossRef] [PubMed]
- Visek, W.J. Diet and cell growth modulation by ammonia. Am. J. Clin. Nutr. 1978, 31, S216–S220. [Google Scholar] [PubMed]
- Scheppach, W.; Luehrs, H.; Menzel, T. Beneficial health effects of low-digestible carbohydrate consumption. Br. J. Nutr. 2001, 85 (Suppl. 1), S23–S30. [Google Scholar] [CrossRef] [PubMed]
- De Preter, V.; Vanhoutte, T.; Huys, G.; Swings, J.; Rutgeerts, P.; Verbeke, K. Baseline microbiota activity and initial bifidobacteria counts influence responses to prebiotic dosing in healthy subjects. Aliment. Pharmacol. Ther. 2008, 27, 504–513. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Relative Abundance (%) | Lactulose 2 g | Lactulose 3 g | Lactulose 4 g | Lactulose 5 g |
|---|---|---|---|---|---|
| Growth enhancement | |||||
| Bifidobacterium | 19.8 | 0.5 | 2.62 | 2.37 | 7.96 |
| Lactobacillus | 6.7 | 0.86 | 1.86 | 2.84 | 1.98 |
| Blautia | 6.1 | 1.61 | 1.99 | 1.89 | 1.52 |
| unclassified_Ruminococcaceae | 1.6 | 0.54 | 1.88 | 1.23 | 1.11 |
| Collinsella | 0.8 | 0.71 | 5.26 | 7.41 | 3.11 |
| Allisonella | 0.3 | 0.66 | 1.38 | 1.46 | 1.06 |
| unclassified_Clostridiales | 0.3 | 0.74 | 1.01 | 1.27 | 1.14 |
| unclassified_Erysipelotrichaceae | 0.2 | 12 | 372 | 352 | 323 |
| Clostridium_XI | 0.1 | 0.8 | 1.2 | 1.6 | 2.4 |
| unclassified_Bacteria | 0.08 | 0 | 2 | 2 | 2 |
| Methanobrevibacter | 0.07 | 0.31 | 3.13 | 4.46 | 1.05 |
| Ruminococcus2 | 0.06 | 1.36 | 8.36 | 14.73 | 6.18 |
| Anaerostipes | 0.03 | >6 | >5.5 | >37 | >39.5 |
| Butyricococcus | 0.02 | 0 | 2.8 | 2.2 | 9.8 |
| Olsenella | 0.01 | 4.86 | 6 | 11.43 | 7.71 |
| unclassified_Coriobacteriaceae | 0.01 | 1 | 6 | 3 | 9 |
| Growth reduction | |||||
| Prevotella | 25.9 | 1.35 | 0.95 | 0.84 | 0.25 |
| Clostridium sensu stricto | 3.9 | 0.14 | 0.33 | 0.62 | 2.05 |
| Ruminococcus | 2.7 | 0.18 | 0.11 | 0.03 | 0.03 |
| Bacteroides | 1.6 | 0 | 0.01 | 0.01 | 0.03 |
| Weissella | 1.3 | 0.99 | 0.38 | 0.24 | 1.4 |
| Dialister | 1.2 | 0.76 | 0.35 | 0.23 | 0.25 |
| Acinetobacter | 0.6 | 0 | 0 | 0 | 0 |
| Escherichia/Shigella | 0.5 | 0 | 0 | 0.55 | 2.18 |
| Enterobacter | 0.3 | 0.32 | 0.31 | 0.78 | 0.45 |
| Peptoniphilus | 0.3 | 0.19 | 0.15 | 0.3 | 0.55 |
| Paraprevotella | 0.3 | 0 | 0.09 | 0.1 | 0.4 |
| unclassified_Enterobacteriaceae | 0.2 | 0.58 | 0.12 | 0.62 | 0.16 |
| Oscillobacter | 0.2 | 0.12 | 0.05 | 0.04 | 0.21 |
| Clostridium XIVa | 0.2 | 0.14 | 0.22 | 0.71 | 0.63 |
| Sutterella | 0.2 | 0.09 | 0.32 | 0.53 | 0.28 |
| Parabacteroides | 0.1 | 0 | 0 | 0 | 0 |
| unclassified_Prevotellaceae | 0.1 | 0.26 | 0.58 | 0.76 | 0.64 |
| Methanosphera | 0.08 | 0.49 | 0.23 | 0.3 | 0.19 |
| Succiniclasticum | 0.06 | 0.06 | 0.02 | 0.13 | 0.03 |
| Shewanella | 0.06 | 0.15 | 0.31 | 0.92 | 6.62 |
| unclassified_Clostridiales | 0.05 | 0.03 | 0.23 | 0.03 | 0.56 |
| unclassified_Firmicutes | 0.05 | 0 | 0 | 0 | 0 |
| Pseudomonas | 0.04 | 0 | 0 | 0 | 0 |
| Atopobium | 0.02 | 0.05 | 0.18 | 0.36 | 0.32 |
| Clostridium_IV | 0.01 | 0.33 | 0 | 0 | 0 |
| Alistipes | 0.01 | 0 | 0 | 0 | 0 |
| Finegoldia | 0.01 | 0 | 0 | 0.27 | 2.53 |
| No clear dose-dependent effect | |||||
| Enterococcus | 14.0 | 0.27 | 0.7 | 1.53 | 4.51 |
| Faecalibacterium | 5.9 | 1.15 | 0.91 | 1.26 | 0.86 |
| Dorea | 0.9 | 0.41 | 0.94 | 1.24 | 0.61 |
| unclassified_Lachnospiraceae | 0.5 | 0.45 | 0.39 | 1.79 | 0.51 |
| Staphylococcus | 0.4 | n/d | n/d | >0.5 | n/d |
| Streptococcus | 0.3 | n/d | n/d | >0.5 | >8 |
| Moraxella | 0.3 | n/d | n/d | n/d | n/d |
| Anaerococcus | 0.3 | 0.05 | 0.89 | 3.07 | 31.96 |
| Roseburia | 0.3 | 3.08 | 0.08 | 12.85 | 5.31 |
| Gemmiger | 0.3 | 0.13 | 0.39 | 2.08 | 0.18 |
| Coprococcus | 0.1 | 0.45 | 0.27 | 3.19 | 4.39 |
| Halomonas | 0.1 | 0.17 | 0.67 | 1.08 | 7 |
| Corynebacterium | 0.1 | n/d | n/d | >0.5 | n/d |
| Subdoligranulum | 0.06 | 0.26 | 1.13 | 0.65 | 0 |
| Lachnospira | 0.06 | 1.38 | 0.03 | 0.74 | 1.69 |
| Clostridium_XIVb | 0.03 | 2 | 0 | 0.1 | 1.14 |
| Catenibacterium | 0.02 | n/d | n/d | >2.5 | >84 |
| unclassified_Bacteroides | 0.02 | 2 | 0 | 10 | 24 |
| Sporobacter | 0.01 | n/d | n/d | n/d | n/d |
| Slackia | 0.01 | >0.5 | n/d | >3 | >2 |
| Akkermansia | 0.01 | n/d | n/d | n/d | n/d |
| Genus | Lactulose 0 g | Lactulose 2 g | Lactulose 3 g | Lactulose 4 g | Lactulose 5 g |
|---|---|---|---|---|---|
| Prevotella | 12,069 | 16,305 | 11,493 | 10,138 | 3072 |
| Bifidobacterium | 1468 | 733 | 3842 | 3474 | 11,689 |
| Enterococcus | 272 | 75 | 191 | 416 | 1225 |
| Lactobacillus | 135 | 116 | 251 | 384 | 267 |
| Blautia | 1934 | 3112 | 3849 | 3654 | 2940 |
| Faecalibacterium | 1941 | 2227 | 1766 | 2448 | 1660 |
| Clostridium_sensu_stricto | 50 | 7 | 17 | 31 | 103 |
| Ruminococcus | 2535 | 457 | 267 | 80 | 65 |
| Bacteroides | 660 | 3 | 7 | 7 | 17 |
| unclassified_Ruminococcaceae | 581 | 315 | 1094 | 715 | 647 |
| Weissella | 40 | 40 | 15 | 10 | 56 |
| Dialister | 740 | 566 | 256 | 167 | 184 |
| Dorea | 415 | 169 | 390 | 515 | 251 |
| Collinsella | 70 | 50 | 368 | 519 | 218 |
| Acinetobacter | 1 | 0 | 0 | 0 | 0 |
| unclassified_Lachnospiraceae | 298 | 134 | 117 | 534 | 151 |
| Escherichia/Shigella | 3 | 0 | 0 | 2 | 6 |
| Staphylococcus | 0 | 0 | 0 | 1 | 0 |
| Moraxella | 0 | 0 | 0 | 0 | 0 |
| Enterobacter | 43 | 14 | 13 | 33 | 19 |
| Anaerococcus | 29 | 2 | 26 | 88 | 911 |
| Peptoniphilus | 178 | 35 | 28 | 54 | 99 |
| Paraprevotella | 43 | 0 | 4 | 5 | 17 |
| Roseburia | 7 | 20 | 1 | 84 | 35 |
| Allisonella | 53 | 35 | 73 | 77 | 56 |
| Streptococcus | 0 | 0 | 0 | 1 | 8 |
| Gemmiger | 162 | 21 | 63 | 338 | 29 |
| unclassified_Clostridiales | 128 | 95 | 129 | 163 | 147 |
| unclassified_Enterobacteriaceae | 63 | 36 | 8 | 39 | 10 |
| Oscillibacter | 286 | 34 | 15 | 11 | 60 |
| Clostridium_XlVa | 153 | 22 | 34 | 109 | 96 |
| Sutterella | 47 | 4 | 15 | 25 | 13 |
| unclassified_Erysipelotrichaceae | 1 | 6 | 186 | 176 | 162 |
| Coprococcus | 30 | 14 | 8 | 95 | 131 |
| Parabacteroides | 4 | 0 | 0 | 0 | 0 |
| Halomonas | 6 | 1 | 4 | 7 | 42 |
| Corynebacterium | 0 | 0 | 0 | 1 | 0 |
| Clostridium_XI | 1 | 1 | 2 | 2 | 3 |
| unclassified_Prevotellaceae | 69 | 18 | 41 | 53 | 44 |
| Methanosphaera | 13 | 7 | 3 | 4 | 3 |
| unclassified_Bacteria | 0 | 0 | 1 | 1 | 1 |
| Methanobrevibacter | 20 | 6 | 61 | 87 | 21 |
| Ruminococcus2 | 6 | 8 | 46 | 81 | 34 |
| Subdoligranulum | 29 | 8 | 33 | 19 | 0 |
| Lachnospira | 20 | 27 | 1 | 15 | 33 |
| Succiniclasticum | 79 | 5 | 2 | 11 | 2 |
| Shewanella | 3 | 1 | 1 | 3 | 22 |
| unclassified_Clostridia | 15 | 1 | 4 | 1 | 9 |
| unclassified_Firmicutes | 3 | 0 | 0 | 0 | 0 |
| Pseudomonas | 0 | 0 | 0 | 0 | 0 |
| Anaerostipes | 0 | 6 | 6 | 37 | 40 |
| Clostridium_XlVb | 5 | 11 | 0 | 1 | 6 |
| Butyricicoccus | 3 | 0 | 7 | 6 | 25 |
| Catenibacterium | 0 | 0 | 0 | 3 | 84 |
| unclassified_Bacteroidetes | 0 | 1 | 0 | 3 | 6 |
| Atopobium | 11 | 1 | 2 | 4 | 4 |
| Sporobacter | 0 | 0 | 0 | 0 | 0 |
| Clostridium_IV | 2 | 1 | 0 | 0 | 0 |
| Olsenella | 2 | 9 | 11 | 20 | 14 |
| unclassified_Coriobacteriaceae | 1 | 1 | 3 | 2 | 5 |
| Alistipes | 0 | 0 | 0 | 0 | 0 |
| Finegoldia | 4 | 0 | 0 | 1 | 10 |
| Slackia | 0 | 1 | 0 | 3 | 2 |
| Akkermansia | 0 | 0 | 0 | 0 | 0 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bothe, M.K.; Maathuis, A.J.H.; Bellmann, S.; Van der Vossen, J.M.B.M.; Berressem, D.; Koehler, A.; Schwejda-Guettes, S.; Gaigg, B.; Kuchinka-Koch, A.; Stover, J.F. Dose-Dependent Prebiotic Effect of Lactulose in a Computer-Controlled In Vitro Model of the Human Large Intestine. Nutrients 2017, 9, 767. https://doi.org/10.3390/nu9070767
Bothe MK, Maathuis AJH, Bellmann S, Van der Vossen JMBM, Berressem D, Koehler A, Schwejda-Guettes S, Gaigg B, Kuchinka-Koch A, Stover JF. Dose-Dependent Prebiotic Effect of Lactulose in a Computer-Controlled In Vitro Model of the Human Large Intestine. Nutrients. 2017; 9(7):767. https://doi.org/10.3390/nu9070767
Chicago/Turabian StyleBothe, Melanie K., Annet J. H. Maathuis, Susann Bellmann, Jos M. B. M. Van der Vossen, Dirk Berressem, Annalena Koehler, Susann Schwejda-Guettes, Barbara Gaigg, Angelika Kuchinka-Koch, and John F. Stover. 2017. "Dose-Dependent Prebiotic Effect of Lactulose in a Computer-Controlled In Vitro Model of the Human Large Intestine" Nutrients 9, no. 7: 767. https://doi.org/10.3390/nu9070767
APA StyleBothe, M. K., Maathuis, A. J. H., Bellmann, S., Van der Vossen, J. M. B. M., Berressem, D., Koehler, A., Schwejda-Guettes, S., Gaigg, B., Kuchinka-Koch, A., & Stover, J. F. (2017). Dose-Dependent Prebiotic Effect of Lactulose in a Computer-Controlled In Vitro Model of the Human Large Intestine. Nutrients, 9(7), 767. https://doi.org/10.3390/nu9070767