Coagulotoxic Cobras: Clinical Implications of Strong Anticoagulant Actions of African Spitting Naja Venoms That Are Not Neutralised by Antivenom but Are by LY315920 (Varespladib)

 , , ,
, , , 
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Antivenom
4.2. Human Plasma
4.3. Human Fibrinogen
4.4. Coagulation Assays Using the Stago STA-R Max
4.5. Phylogenetic Comparative Analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chippaux, J. Snakebite envenomation turns again into a neglected tropical disease! J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 38. [Google Scholar] [CrossRef]
- Fry, B.G. Snakebite: When the human touch becomes a bad touch. Toxins 2018, 10, 170. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.M.; Theakston, R.D.; Warrell, D.A. Confronting the neglected problem of snake bite envenoming: The need for a global partnership. PLoS Med. 2006, 3, e150. [Google Scholar] [CrossRef]
- Kasturiratne, A.; Wickremasinghe, A.R.; de Silva, N.; Gunawardena, N.K.; Pathmeswaran, A.; Premaratna, R.; Savioli, L.; Lalloo, D.G.; de Silva, H.J. The global burden of snakebite: A literature analysis and modelling based on regional estimates of envenoming and deaths. PLoS Med. 2008, 5, e218. [Google Scholar] [CrossRef] [PubMed]
- Longbottom, J.; Shearer, F.M.; Devine, M.; Alcoba, G.; Chappuis, F.; Weiss, D.J.; Ray, S.E.; Ray, N.; Warrell, D.A.; Ruiz de Castaneda, R.; et al. Vulnerability to snakebite envenoming: A global mapping of hotspots. Lancet 2018, 392, 673–684. [Google Scholar] [CrossRef]
- Boyer, L.; Alagón, A.; Fry, B.G.; Jackson, T.N.W.; Sunagar, K.; Chippaux, J.P. Signs, Symptoms and Treatment of Envenomation. In Venomous Reptiles and Their Toxins: Evolution, Pathophysiology and Biodiscovery; Fry, B.G., Ed.; Oxford University Press: New York, NY, USA, 2015; pp. 32–60. [Google Scholar]
- Harrison, R.A.; Hargreaves, A.; Wagstaff, S.C.; Faragher, B.; Lalloo, D.G. Snake envenoming: A disease of poverty. PLoS Negl. Trop. Dis. 2009, 3, e569. [Google Scholar] [CrossRef] [PubMed]
- Bénard-Valle, M.; Neri-Castro, E.E.; Fry, B.G.; Boyer, L.; Cochran, C.; Alam, M.; Jackson, T.N.W.; Paniagua, D.; Olvera-Rodríguez, F.; Koludarov, I.; et al. Antivenom research and development. In Venomous Reptiles and Their Toxins: Evolution, Pathophysiology and Biodiscovery; Fry, B.G., Ed.; Oxford University Press: New York, NY, USA, 2015; pp. 61–72. [Google Scholar]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The toxicogenomic multiverse: Convergent recruitment of proteins into animal venoms. Ann. Rev. Genomics Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef]
- Osipov, A.V.; Filkin, S.Y.; Makarova, Y.V.; Tsetlin, V.I.; Utkin, Y.N. A new type of thrombin inhibitor, noncytotoxic phospholipase A2, from the Naja haje cobra venom. Toxicon 2010, 55, 186–194. [Google Scholar] [CrossRef]
- Mukherjee, A.K.; Kalita, B.; Thakur, R. Two acidic, anticoagulant PLA2 isoenzymes purified from the venom of monocled cobra Naja kaouthia exhibit different potency to inhibit thrombin and factor Xa via phospholipids independent, non-enzymatic mechanism. PloS ONE 2014, 9, e101334. [Google Scholar] [CrossRef]
- Kerns, R.T.; Kini, R.M.; Stefansson, S.; Evans, H.J. Targeting of venom phospholipases: The strongly anticoagulant phospholipase A(2) from Naja nigricollis venom binds to coagulation factor Xa to inhibit the prothrombinase complex. Arch. Biochem. Biophys. 1999, 369, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Kini, R.M. Structure-function relationships and mechanism of anticoagulant phospholipase A2 enzymes from snake venoms. Toxicon 2005, 45, 1147–1161. [Google Scholar] [CrossRef] [PubMed]
- Stefansson, S.; Kini, R.M.; Evans, H.J. The basic phospholipase A2 from Naja nigricollis venom inhibits the prothrombinase complex by a novel nonenzymatic mechanism. Biochemistry 1990, 29, 7742–7746. [Google Scholar] [CrossRef] [PubMed]
- Mladic, M.; Zietek, B.M.; Iyer, J.K.; Hermarij, P.; Niessen, W.M.; Somsen, G.W.; Kini, R.M.; Kool, J. At-line nanofractionation with parallel mass spectrometry and bioactivity assessment for the rapid screening of thrombin and factor Xa inhibitors in snake venoms. Toxicon 2016, 110, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Panagides, N.; Jackson, T.N.; Ikonomopoulou, M.P.; Arbuckle, K.; Pretzler, R.; Yang, D.C.; Ali, S.A.; Koludarov, I.; Dobson, J.; Sanker, B.; et al. How the cobra got its flesh-eating venom: Cytotoxicity as a defensive innovation and its co-evolution with hooding, aposematic marking, and spitting. Toxins 2017, 9, 103. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Sanders, K.L.; King, B.; Palci, A. Diversification rates and phenotypic evolution in venomous snakes (Elapidae). R. Soc. Open Sci. 2016, 3, 150277. [Google Scholar] [CrossRef] [Green Version]
- Wuster, W.; Crookes, S.; Ineich, I.; Mane, Y.; Pook, C.E.; Trape, J.F.; Broadley, D.G. The phylogeny of cobras inferred from mitochondrial DNA sequences: Evolution of venom spitting and the phylogeography of the African spitting cobras (Serpentes: Elapidae: Naja nigricollis complex). Mol. Phylogenet. Evol. 2007, 45, 437–453. [Google Scholar] [CrossRef] [PubMed]
- Warrell, D.A.; Greenwood, B.M.; Davidson, N.M.; Ormerod, L.D.; Prentice, C.R. Necrosis, haemorrhage and complement depletion following bites by the spitting cobra (Naja nigricollis). Q. J. Med. 1976, 45, 1–22. [Google Scholar] [PubMed]
- Lewin, M.; Samuel, S.; Merkel, J.; Bickler, P. Varespladib (LY315920) appears to be a potent, broad-spectrum, inhibitor of snake venom phospholipase A2 and a possible pre-referral treatment for envenomation. Toxins 2016, 8, 248. [Google Scholar] [CrossRef] [PubMed]
- Saaiman, E.; Buys, C.; Fry, B.G.; Windhoek, Namibia. Haemorrhage following Naja nigricincta envenomation. Personal communication, 2018. [Google Scholar]
- Kandiwa, E.; Mushonga, B.; Samkange, A.; Fabiano, E. Quantitative characterization of the hemorrhagic, necrotic, coagulation-altering properties and edema-forming effects of zebra snake (Naja nigricincta nigricincta) venom. J. Toxicol. 2018, 2018, 6940798. [Google Scholar] [CrossRef]
- Mukherjee, A.K.; Maity, C.R. Biochemical composition, lethality and pathophysiology of venom from two cobras—Naja naja and N. kaouthia. Comp. Biochem. Physiol. Part B Biochem. Molecular Biol. 2002, 131, 125–132. [Google Scholar] [CrossRef]
- Sunagar, K.; Jackson, T.N.W.; Reeks, T.; Fry, B.G. Group I Phospholipase A2 Enzymes. In Venomous Reptiles and Their Toxins: Evolution, Pathophysiology and Biodiscovery; Fry, B.G., Ed.; Oxford University Press: New York, NY, USA, 2015; pp. 324–327. [Google Scholar]
- Lewin, M.R.; Gutierrez, J.M.; Samuel, S.P.; Herrera, M.; Bryan-Quiros, W.; Lomonte, B.; Bickler, P.E.; Bulfone, T.C.; Williams, D.J. Delayed Oral LY333013 Rescues Mice from Highly Neurotoxic, Lethal Doses of Papuan Taipan (Oxyuranus scutellatus) Venom. Toxins 2018, 10, 380. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, J.; Zhang, D.; Xiao, H.; Xiong, S.; Huang, C. Exploration of the inhibitory potential of varespladib for snakebite envenomation. Molecules 2018, 23, 391. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Li, H.; Zhang, D.; Li, Y.; Sun, S.; Huang, C. Inactivation of Venom PLA(2) Alleviates myonecrosis and facilitates muscle regeneration in envenomed mice: A time course observation. Molecules 2018, 23, 1911. [Google Scholar] [CrossRef] [PubMed]
- Tilbury, C.R. Observations on the bite of the Mozambique spitting cobra (Naja mossambica mossambica). S. Afr. Med. J. 1982, 61, 308–313. [Google Scholar] [PubMed]
- Vermaak, S.S. A deadly bed partner: M’fess (Mozambique spitting cobra). SA Orthop. J. 2010, 58–62. [Google Scholar]
- Pattinson, J.P.; Kong, V.Y.; Bruce, J.L.; Oosthuizen, G.V.; Bekker, W.; Laing, G.L.; Wood, D.; Brysiewicz, P.; Clarke, D.L. Defining the need for surgical intervention following a snakebite still relies heavily on clinical assessment: The experience in Pietermaritzburg, South Africa. S. Afr. Med. J. 2017, 107, 1082–1085. [Google Scholar] [CrossRef]
- Müller, G.J. Snake bite in southern Africa: Diagnosis and Management. CME 2012, 30, 362–391. [Google Scholar]
- Rivel, M.; Solano, D.; Herrera, M.; Vargas, M.; Villalta, M.; Segura, A.; Arias, A.S.; Leon, G.; Gutierrez, J.M. Pathogenesis of dermonecrosis induced by venom of the spitting cobra, Naja nigricollis: An experimental study in mice. Toxicon 2016, 119, 171–179. [Google Scholar] [CrossRef]
- Mendez, I.; Gutierrez, J.M.; Angulo, Y.; Calvete, J.J.; Lomonte, B. Comparative study of the cytolytic activity of snake venoms from African spitting cobras (Naja spp., Elapidae) and its neutralization by a polyspecific antivenom. Toxicon 2011, 58, 558–564. [Google Scholar] [CrossRef]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef]
- Revell, L.J. phytools: An R package for phylogenetic comparative biology (and other things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Symonds, M.R.E.; Blomberg, S.P. A Primer on Phylogenetic Generalised Least Squares. In Modern Phylogenetic Comparative Methods and Their Application in Evolutionary Biology; Garamszegi, L.Z., Ed.; Springer: Berlin/Heidelberg, Germay, 2014; pp. 105–130. [Google Scholar]
- Orme, D.; Freckleton, R.; Thomas, G.; Petzoldt, T.; Fritz, S.; Isaac, N.; Pearse, W. Caper: Comparative Analyses of Phylogenetics and Evolution in R. R Package Version 0.5.2. Methods Ecol. Evol. 2013, 3, 145–151. [Google Scholar]
- Lister, C.; Arbuckle, K.; Jackson, T.N.W.; Debono, J.; Zdenek, C.N.; Dashevsky, D.; Dunstan, N.; Allen, L.; Hay, C.; Bush, B.; et al. Catch a tiger snake by its tail: Differential toxicity, co-factor dependence and antivenom efficacy in a procoagulant clade of Australian venomous snakes. Comp. Biochem. Physiol. Toxicol. Pharmacol. CBP 2017, 202, 39–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogalski, A.; Soerensen, C.; Op den Brouw, B.; Lister, C.; Dashevsky, D.; Arbuckle, K.; Gloria, A.; Zdenek, C.N.; Casewell, N.R.; Gutierrez, J.M.; et al. Differential procoagulant effects of saw-scaled viper (Serpentes: Viperidae: Echis) snake venoms on human plasma and the narrow taxonomic ranges of antivenom efficacies. Toxicol. Lett. 2017, 280, 159–170. [Google Scholar] [CrossRef] [PubMed]





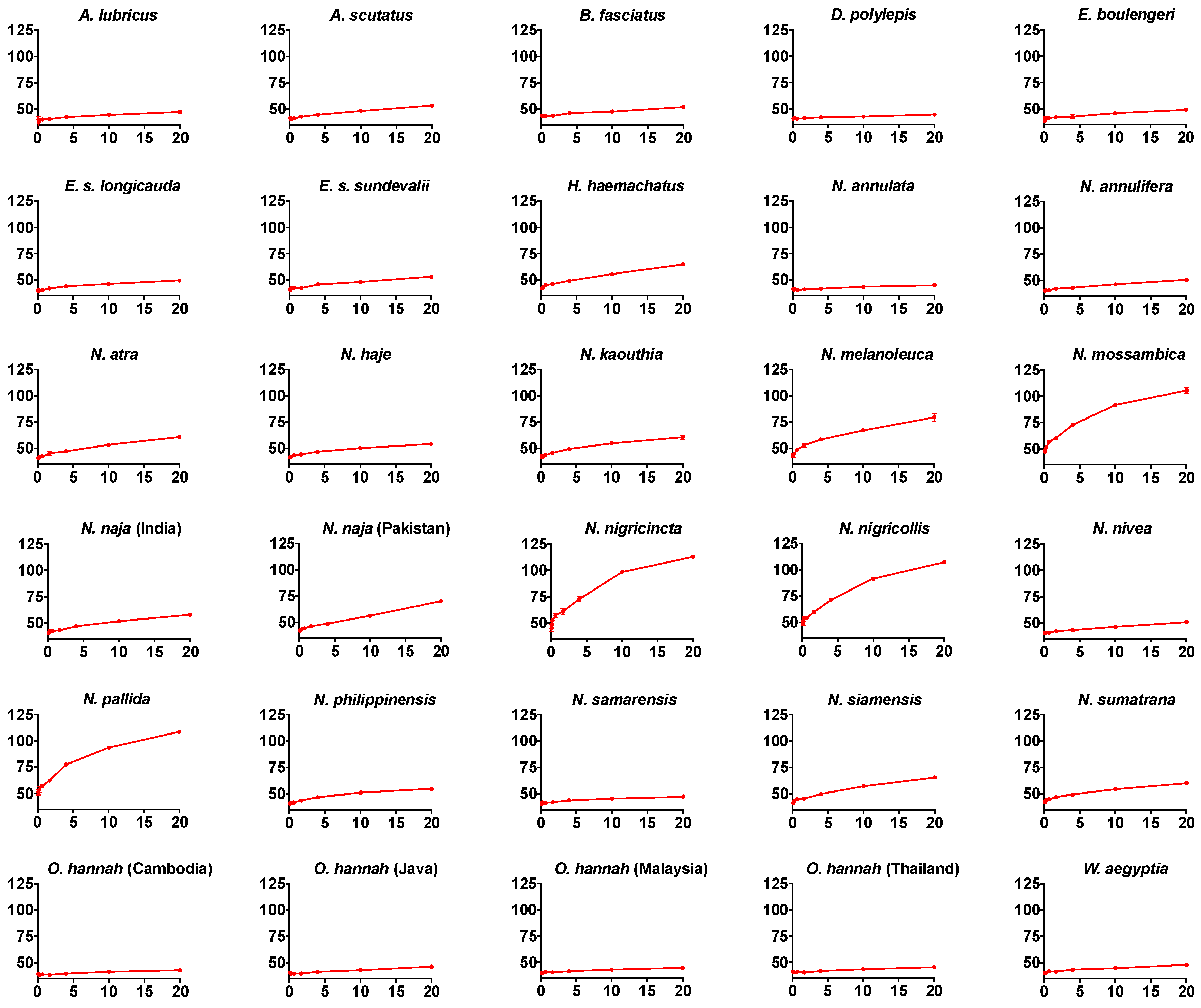{kind=link}
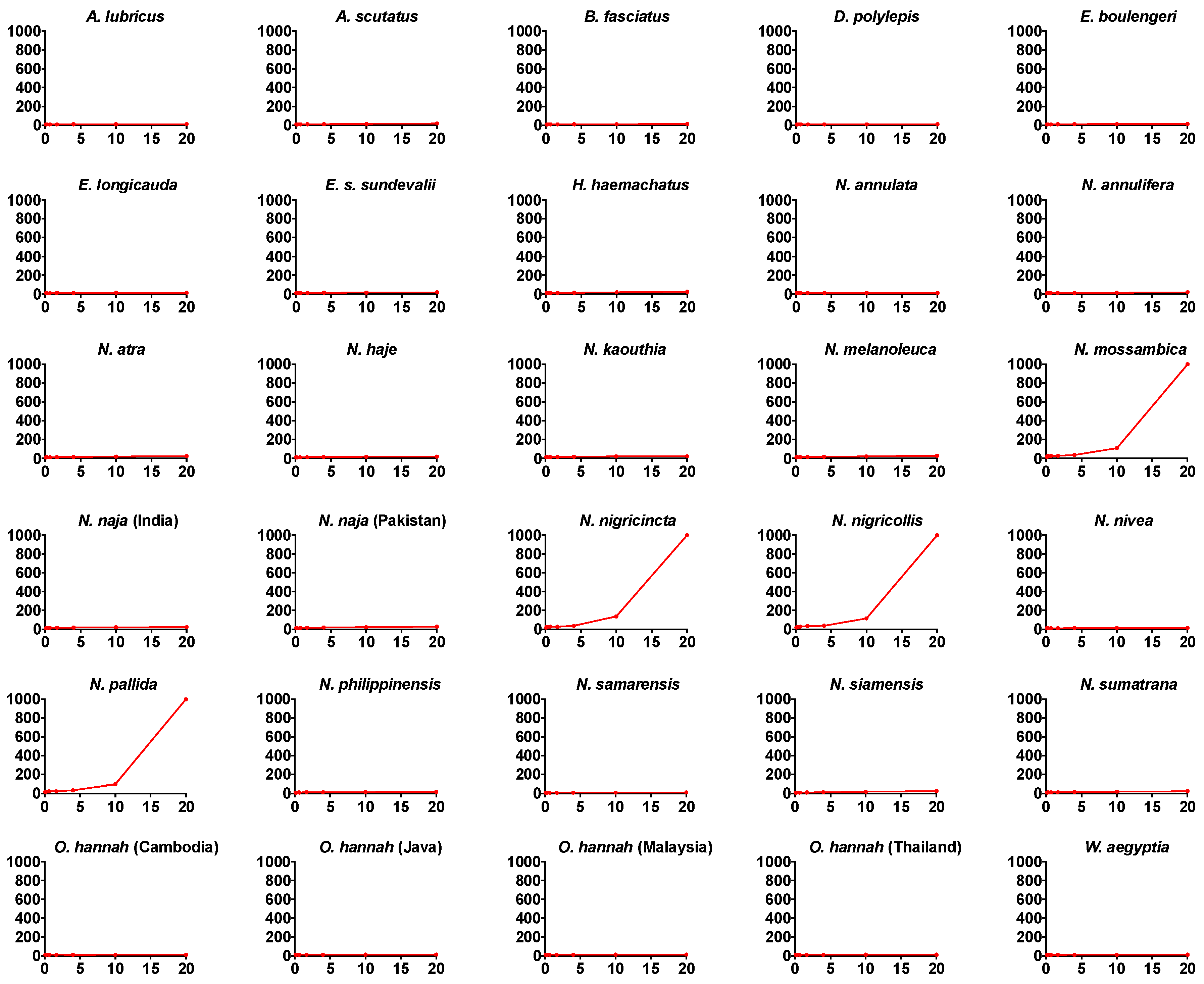{kind=link}
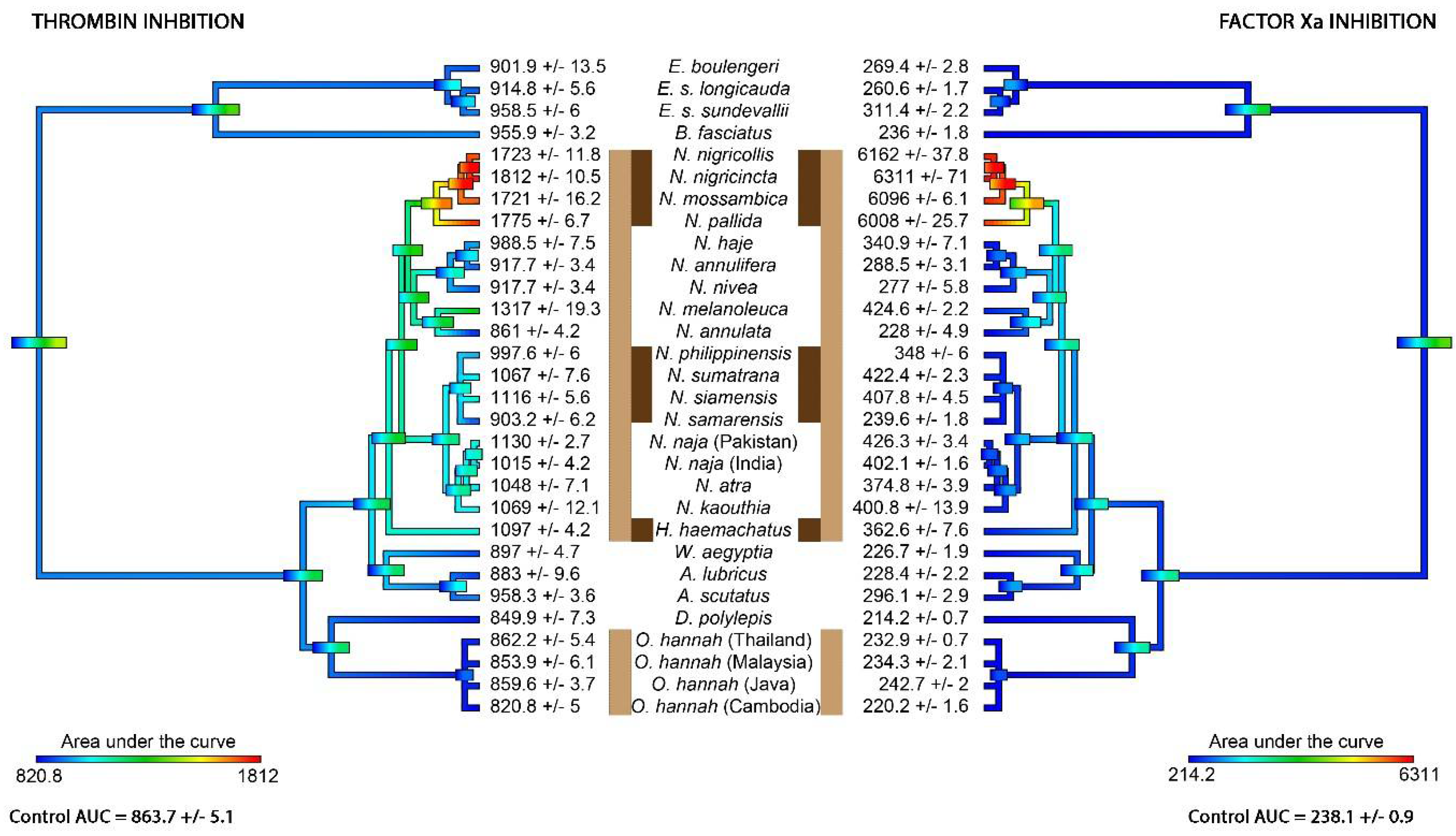{kind=link}
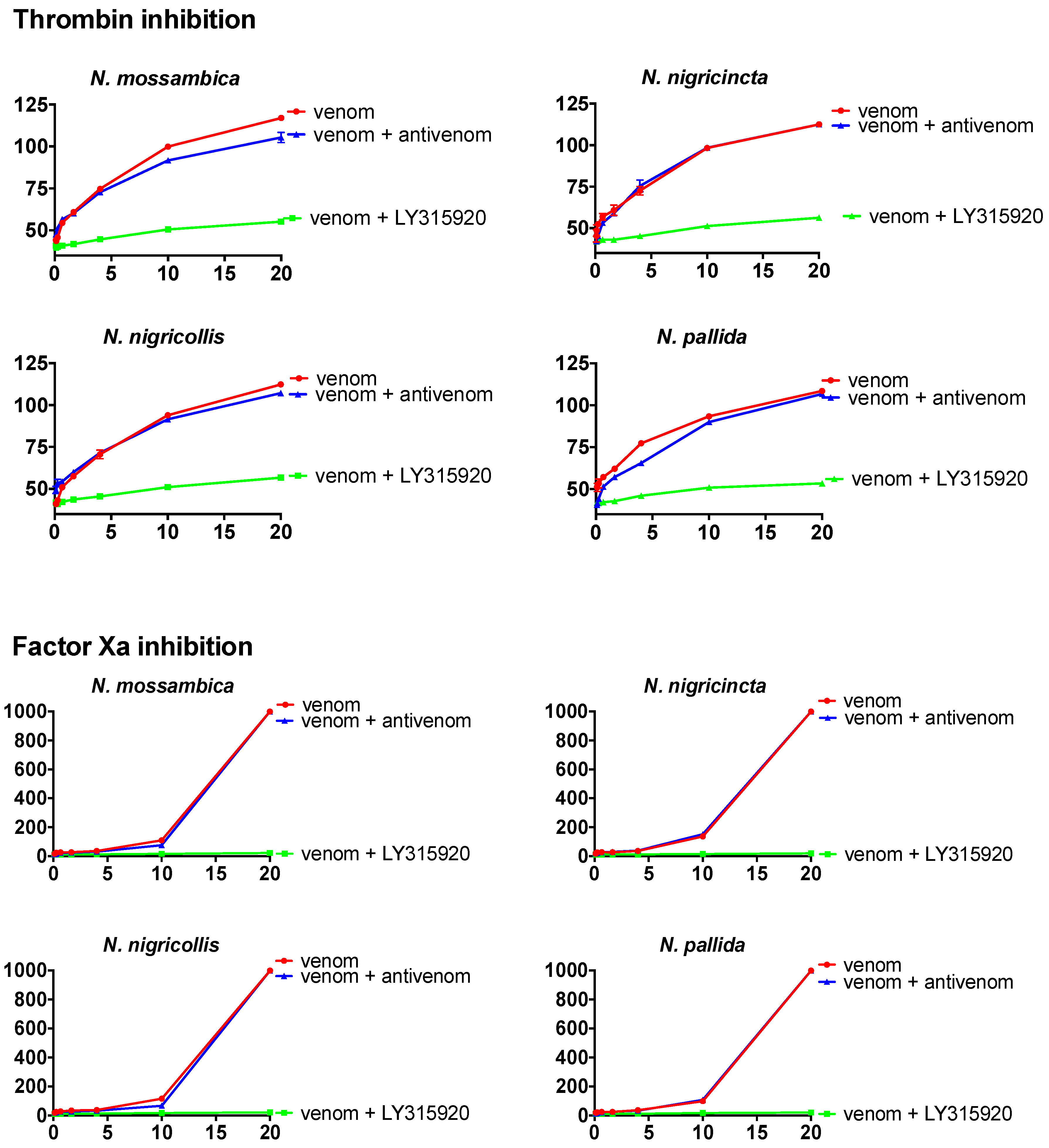{kind=link}
| Thrombin inhibition assay | Step 1. 25 µL of 0.2 mg/mL venom (1 mg/mL 50% glycerol stock diluted with Owren Koller (OK) Buffer (Stago Catalog # 00360) + 75 µL of [50 µL 0.025 M calcium (Stago catalog # 00367 + 25 µL of OK buffer] + 50 µL of phospholipid (Stago catalog #00597) + 25 µL of thrombin (Stago catalog # 00611). Step 2. 120 second incubation. Step 3. Addition of 75 µL of 4 mg/mL fibrinogen. |
| Thrombin inhibition assay (antivenom neutralisation tests) | Step 1. 25 µL of 0.2 mg/mL venom (1 mg/mL 50% glycerol stock diluted with OK Buffer + 75 µL of [50 µL 0.025 M calcium + 25 µL of 5% concentration antivenom] + 50 µL phospholipid + 25 µL of thrombin. Step 2. 120 s incubation. Step 3. Addition of 75 µL of 4 mg/mL fibrinogen. |
| Thrombin inhibition assay (LY315920 [varespladib] neutralisation tests) | Step 1. 25 µL of 0.2 mg/mL venom (1 mg/mL 50% glycerol stock diluted with OK Buffer + 75 µL of [50 µL 0.025 M calcium + 25 µL of 0.025 mg/mL LY315920] + 50 µL phospholipid + 25 µL of thrombin. Step 2. 120 s incubation. Step 3. Addition of 75 µL of 4 mg/mL fibrinogen. |
| FXa inhibition assay | Step 1. 25 µL 0.2 mg/mL venom (1 mg/mL 50% glycerol stock diluted with OK Buffer + 75 μl of [50 µL 0.025 M calcium +25 µL OK buffer] + 50 µL of phospholipid + 25 µL of FXa (Stago catalog # 00311). Step 2. 120 s incubation. Step 3. Addition of 75 µL plasma. |
| FXa inhibition assay (antivenom neutralisation tests) | Step 1. 25 µL 0.2 mg/mL of venom (1 mg/mL 50% glycerol stock diluted with OK Buffer + 75 µL of [50 µL 0.025 M calcium + 25 µL of 5% concentration antivenom] + 50 µL of phospholipid + 50 µL of FXa. Step 2. 120 s incubation. Step 3. Addition of 75 µL of plasma. |
| FXa inhibition assay (LY315920 neutralisation tests) | Step 1. 25 µL of 0.2 mg/mL venom (1 mg/mL 50% glycerol stock diluted with OK Buffer + 75 µL of [50 µL 0.025 M calcium + 25 µL of 0.025 mg/mL LY315920] + 50 µL of phospholipid + 25 µL of FXa. Step 2. 120 s incubation. Step 3. Addition of 75 µL of plasma. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bittenbinder, M.A.; Zdenek, C.N.; Op den Brouw, B.; Youngman, N.J.; Dobson, J.S.; Naude, A.; Vonk, F.J.; Fry, B.G. Coagulotoxic Cobras: Clinical Implications of Strong Anticoagulant Actions of African Spitting Naja Venoms That Are Not Neutralised by Antivenom but Are by LY315920 (Varespladib). Toxins 2018, 10, 516. https://doi.org/10.3390/toxins10120516
Bittenbinder MA, Zdenek CN, Op den Brouw B, Youngman NJ, Dobson JS, Naude A, Vonk FJ, Fry BG. Coagulotoxic Cobras: Clinical Implications of Strong Anticoagulant Actions of African Spitting Naja Venoms That Are Not Neutralised by Antivenom but Are by LY315920 (Varespladib). Toxins. 2018; 10(12):516. https://doi.org/10.3390/toxins10120516
Chicago/Turabian StyleBittenbinder, Mátyás A., Christina N. Zdenek, Bianca Op den Brouw, Nicholas J. Youngman, James S. Dobson, Arno Naude, Freek J. Vonk, and Bryan G. Fry. 2018. "Coagulotoxic Cobras: Clinical Implications of Strong Anticoagulant Actions of African Spitting Naja Venoms That Are Not Neutralised by Antivenom but Are by LY315920 (Varespladib)" Toxins 10, no. 12: 516. https://doi.org/10.3390/toxins10120516
APA StyleBittenbinder, M. A., Zdenek, C. N., Op den Brouw, B., Youngman, N. J., Dobson, J. S., Naude, A., Vonk, F. J., & Fry, B. G. (2018). Coagulotoxic Cobras: Clinical Implications of Strong Anticoagulant Actions of African Spitting Naja Venoms That Are Not Neutralised by Antivenom but Are by LY315920 (Varespladib). Toxins, 10(12), 516. https://doi.org/10.3390/toxins10120516