Camelid Single-Domain Antibodies (VHHs) against Crotoxin: A Basis for Developing Modular Building Blocks for the Enhancement of Treatment or Diagnosis of Crotalic Envenoming

 , , ,
, , ,  ,
, 
 and
and
Abstract
:1. Introduction
2. Results
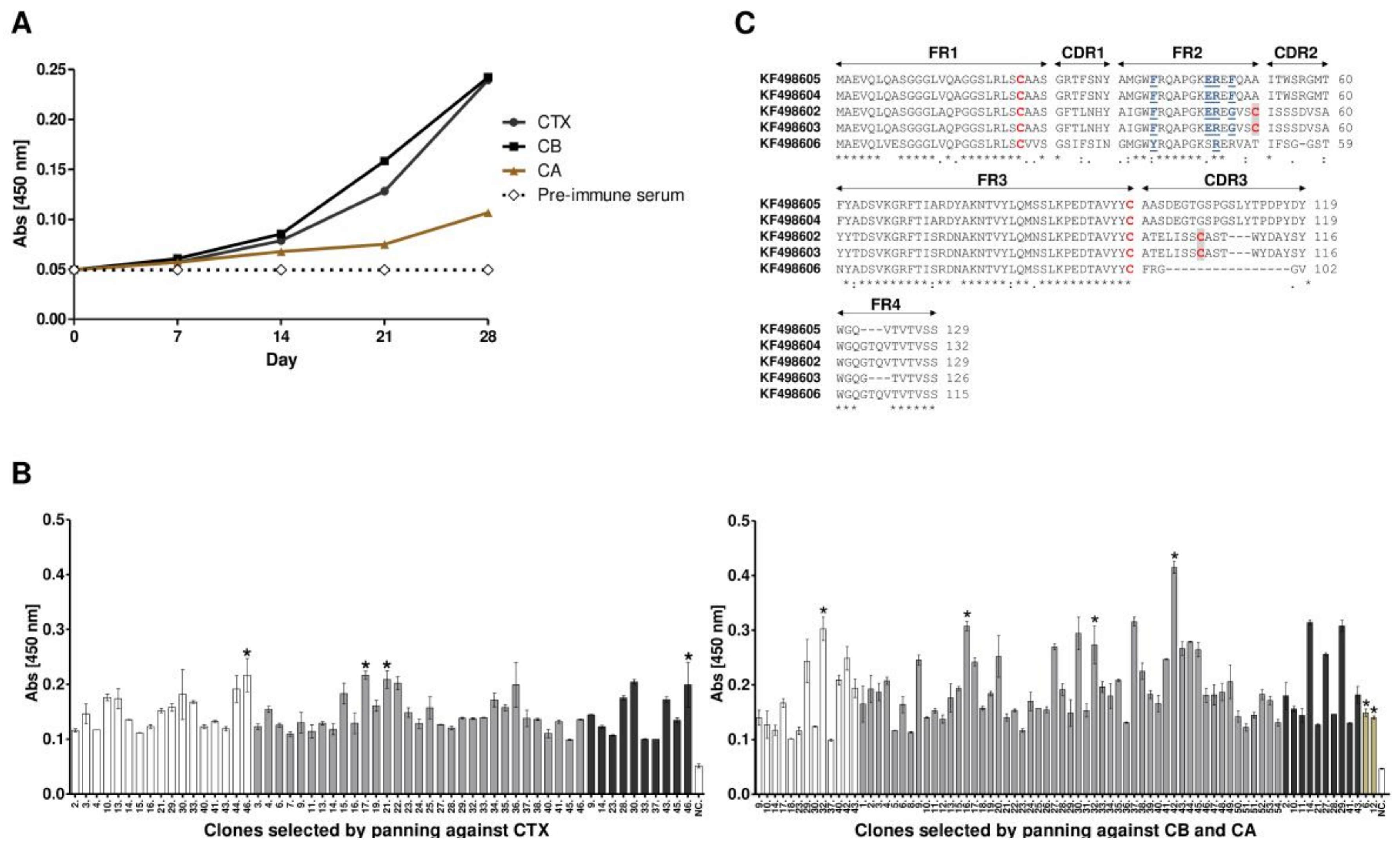
2.1. L. glama Immunization, Library Construction and Selection of Anti-Crotoxin VHHs
2.2. Sequence Analysis of Anti-Crotoxin VHHs
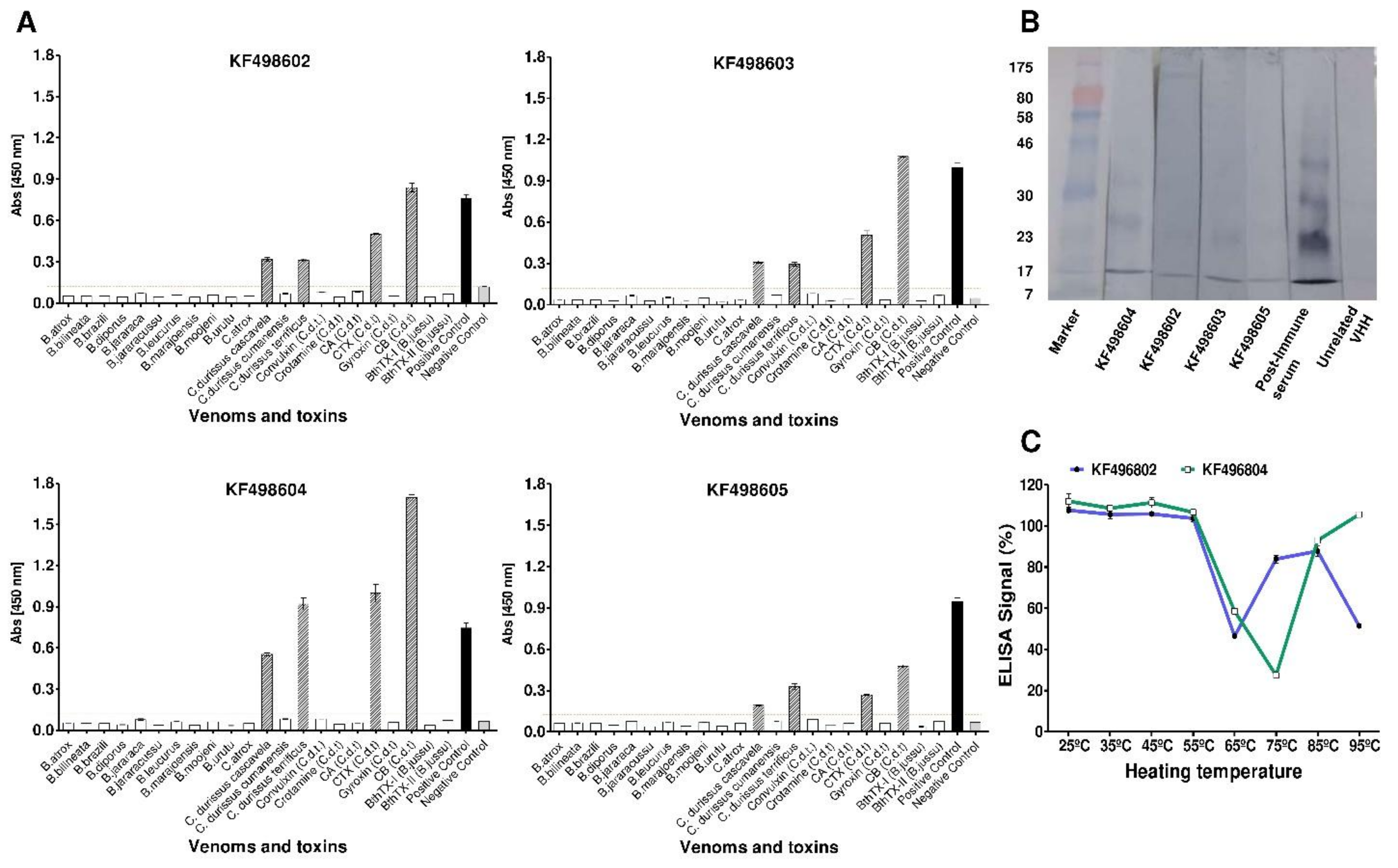
2.3. Cross-Immunoreactivity and Western Blot Analysis
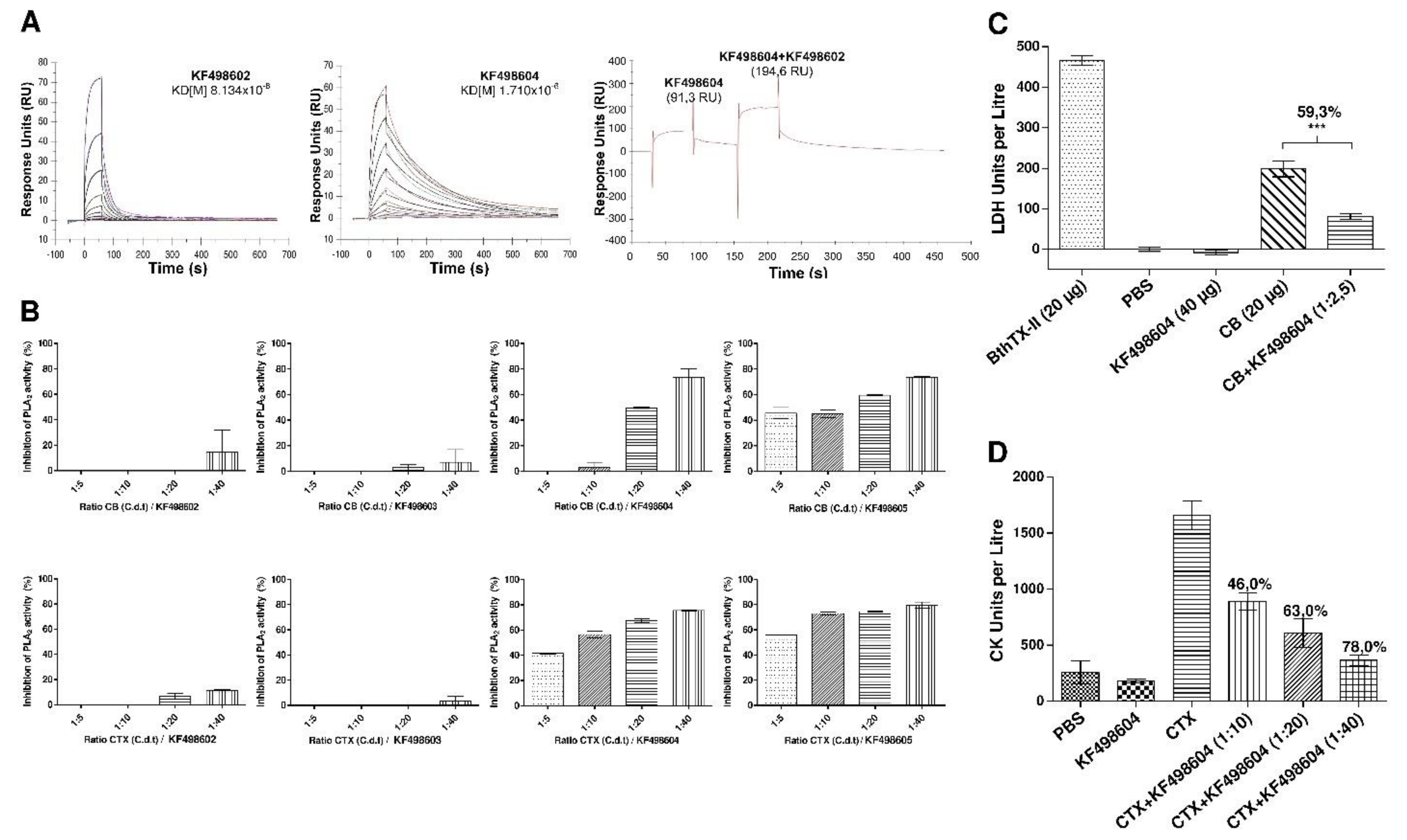
2.4. Thermal Stability Evaluation of Selected VHHs and Interaction Analysis by SPR
2.5. In Vitro and In Vivo Inhibition of CTX Activity by VHHs
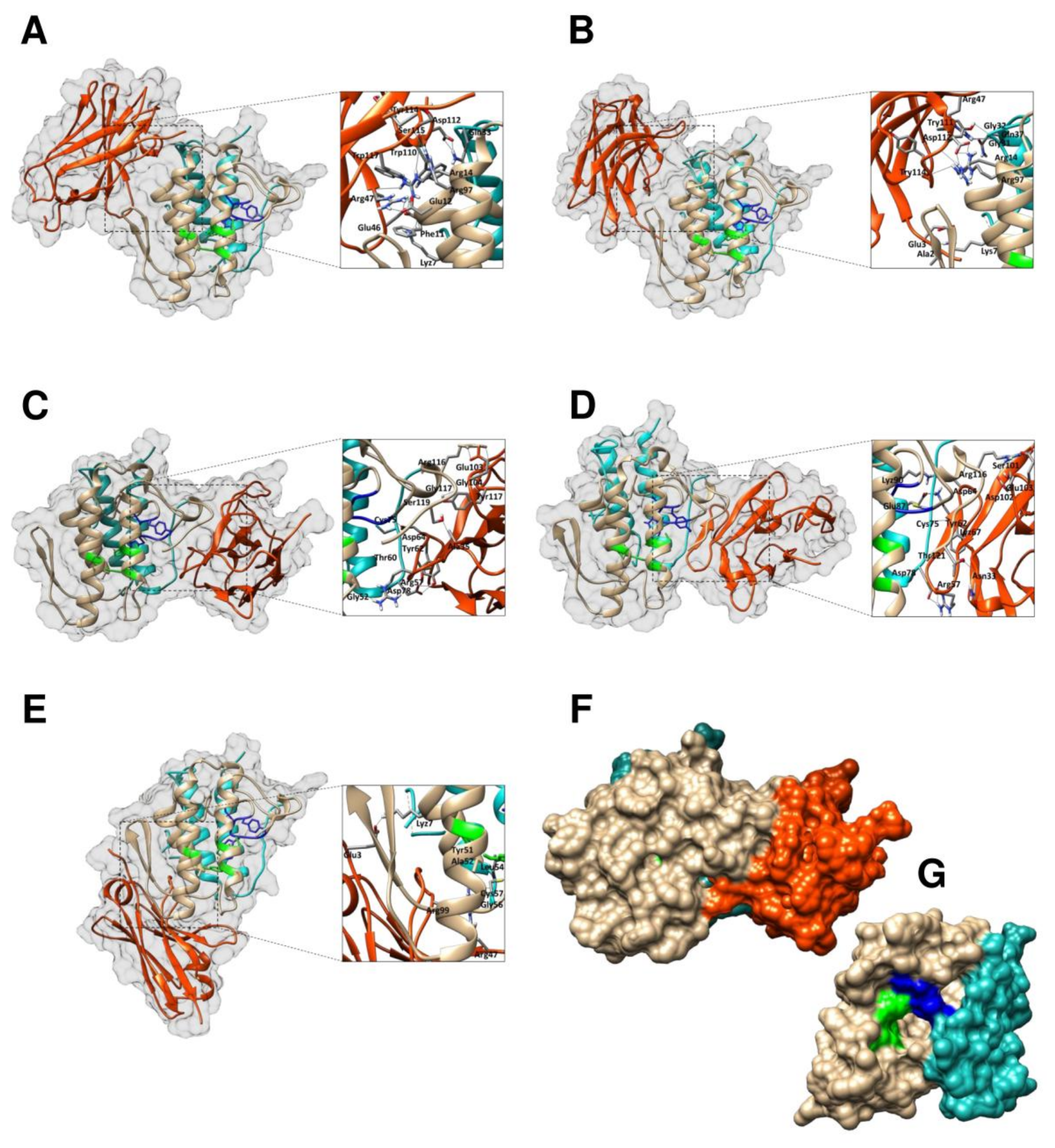
2.6. Modelling and Interface Binding of the VHHs and CTX
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Camelid Immunization and Humoral Immune Response Monitoring
4.3. Selection of Anti-Crotoxin VHHs
4.4. Expression and Purification of Anti-Crotoxin VHHs
4.5. Cross-Immunoreactivity Analysis of Anti-Crotoxin VHHs
4.6. Western Blot Analysis of Anti-Crotoxin VHHs
4.7. Thermal Stability Analysis of Anti-Crotoxin VHHs
4.8. Interaction Analyses by Surface Plasmon Resonance (SPR)
4.9. In Vitro Inhibition of CTX and CB Phospholipase Activities
4.10. In Vitro Inhibition of CB Cytotoxicity by Selected VHHs
4.11. In Vivo Neutralization of CTX Myotoxicity by VHHs
4.12. Modeling and Molecular Docking of Anti-Crotoxin VHHs
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- WHO. A Systematic Technically Driven Process for the Adoption of Additional Diseases as NTDs. Available online: https://www.who.int/neglected_diseases/diseases/systematic_technically_driven_process/en/ (accessed on 15 December 2017).
- Kasturiratne, A.; Wickremasinghe, A.R.; de Silva, N.; Gunawardena, N.K.; Pathmeswaran, A.; Premaratna, R.; Savioli, L.; Lalloo, D.G.; de Silva, H.J. The global burden of snakebite: A literature analysis and modelling based on regional estimates of envenoming and deaths. PLoS Med. 2008, 5, e218. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.P. Epidemiology of envenomations by terrestrial venomous animals in Brazil based on case reporting: From obvious facts to contingencies. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Bochner, R.; Struchiner, C.J. Snake bite epidemiology in the last 100 years in Brazil: A review. Cad. Saude Publica 2003, 19, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Brasil. Ministério da Saúde. Secretaria de Vigilância em Saúde. Guia de Vigilância Epidemiológica. Caderno 14. Acidentes por Animais Peçonhentos. Available online: https://portal.saude.gov.br/portal/arquivos/pdf/ gve_7ed_web_atual. pdf (accessed on 15 December 2017).
- Azevedo-marques, M.M.; Hering, S.E.; Cupo, P. Acidente Crotálico. In Animais Peçonhentos no Brasil: Biologia, Clínica e Terapêutica dos Acidentes, 2nd ed.; Cardoso, J.L.C., Ed.; Sarvier Editora, FAPESP: São Paulo, Brasil, 2009; pp. 91–97. ISBN 85-7378-133-5. [Google Scholar]
- Bucaretchi, F.; Herrera, S.R.F.; Hyslop, S.; Baracat, E.C.E.; Vieira, R.J. Snakebites by Crotalus durissus ssp. in children in Campinas, São Paulo, Brazil. Rev. Inst. Med. Trop. São Paulo 2002, 44, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Pinho, F.M.; Zanetta, D.M.; Burdmann, E.A. Acute renal failure after Crotalus durissus snakebite: A prospective survey on 100 patients. Kidney Int. 2005, 67, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, D.; Ohler, M.; Seifert, J.; von Bergen, M.; Arni, R.K.; Genov, N.; Betzel, C. Snake Venomic of Crotalus durissus terrificus correlation with pharmacological activities. J. Proteome Res. 2010, 9, 2302–2316. [Google Scholar] [CrossRef] [PubMed]
- Faure, G.; Xu, H.; Saul, F.A. Crystal structure of crotoxin reveals key residues involved in the stability and toxicity of this potent heterodimeric β-neurotoxin. J. Mol. Biol. 2011, 412, 176–191. [Google Scholar] [CrossRef] [PubMed]
- Theakston, R.D.; Warrell, D.A.; Griffiths, E. Report of a WHO workshop on the standardization and control of antivenoms. Toxicon 2003, 41, 541–557. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; León, G.; Lomonte, B. Pharmacokinetic-pharmacodynamic relationships of immunoglobulin therapy for envenomation. Clin. Pharmacokinet. 2003, 42, 721–741. [Google Scholar] [CrossRef] [PubMed]
- De Silva, H.A.; Ryan, N.M.; de Silva, H.J. Adverse reactions to snake antivenom, and their prevention and treatment. Br. J. Clin. Pharmacol. 2016, 81, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.I. Consequences of neglect: Analysis of the sub-saharan african snake antivenom market and the global context. PLoS Negl. Trop. Dis. 2012, 6, e1670. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M. Improving antivenom availability and accessibility: Science, technology, and beyond. Toxicon 2012, 60, 676–687. [Google Scholar] [CrossRef] [PubMed]
- Morais, V.; Massaldi, H. Economic evaluation of snake antivenom production in the public system. J. Venom. Anim. Toxins Incl. Trop. Dis. 2006, 12, 497–511. [Google Scholar] [CrossRef]
- Alvarenga, L.M.; Zahid, M.; di Tommaso, A.; Juste, M.O.; Aubrey, N.; Billiald, P.; Muzard, J. Engineering venom’s toxin-neutralizing antibody fragments and its therapeutic potential. Toxins 2014, 6, 2541–2567. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, A.H.; Solà, M.; Jappe, E.C.; Oscoz, S.; Lauridsen, L.P.; Engmark, M. Biotechnological trends in spider and scorpion antivenom development. Toxins 2016, 8, 226. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Phoolcharoen, W.; Lai, H.; Piensook, K.; Cardineau, G.; Zeitlin, L.; Whaley, K.J.; Arntzen, C.J.; Mason, H.S.; Chen, Q. High-level rapid production of full-size monoclonal antibodies in plants by a single-vector DNA replicon system. Biotechnol. Bioeng. 2010, 106, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.Z. Monoclonal Antibodies as Diagnostics; an Appraisal. Indian J. Pharm. Sci. 2010, 72, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.C.; Tuszynski, G.P.; Blackman, M.R.; Sharma, M. Long-term efficacy and downstream mechanism of anti-annexinA2 monoclonal antibody (anti-ANX A2 mAb) in a pre-clinical model of aggressive human breast cancer. Cancer Lett. 2016, 373, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.P.; Goyffon, M. Venoms, antivenoms and immunotherapy. Toxicon 1998, 36, 823–846. [Google Scholar] [CrossRef]
- Tjandra, J.J.; Ramadi, L.; McKenzie, I.F. Development of human anti-murine antibody (HAMA) response in patients. Immunol. Cell Biol. 1990, 68, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Nelson, A.L. Antibody fragments: Hope and hype. MAbs 2010, 2, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Roskos, L.K.; Davis, C.G.; Schwab, G.M. The clinical pharmacology of therapeutic monoclonal antibodies. Drug Dev. Res. 2004, 61, 108–120. [Google Scholar] [CrossRef]
- Batra, S.K.; Jain, M.; Wittel, U.A.; Chauhan, S.C.; Colcher, D. Pharmacokinetics and biodistribution of genetically engineered antibodies. Curr. Opin. Biotechnol. 2002, 13, 603–608. [Google Scholar] [CrossRef]
- Colcher, D.; Bird, R.; Roselli, M.; Hardman, K.D.; Johnson, S.; Pope, S.; Dodd, S.W.; Pantoliano, M.W.; Milenic, D.E.; Schlom, J. In vivo tumor targeting of a recombinant single-chain antigen-binding protein. J. Natl. Cancer Inst. 1990, 82, 1191–1197. [Google Scholar] [CrossRef] [PubMed]
- Nelson, A.L.; Reichert, J.M. Development trends for therapeutic antibody fragments. Nat. Biotechnol. 2009, 27, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Pavlinkova, G.; Beresford, G.W.; Booth, B.J.; Batra, S.K.; Colcher, D. Pharmacokinetics and biodistribution of engineered single-chain antibody constructs of MAb CC49 in colon carcinoma xenografts. J. Nucl. Med. 1999, 40, 1536–1546. [Google Scholar] [PubMed]
- Goel, A.; Colcher, D.; Baranowska-Kortylewicz, J.; Augustine, S.; Booth, B.J.; Pavlinkova, G.; Batra, S.K. Genetically engineered tetravalent single-chain Fv of the pancarcinoma monoclonal antibody CC49: Improved biodistribution and potential for therapeutic application. Cancer Res. 2000, 60, 6964–6971. [Google Scholar] [PubMed]
- Miller, B.R.; Demarest, S.J.; Lugovskoy, A.; Huang, F.; Wu, X.; Snyder, W.B.; Croner, L.J.; Wang, N.; Amatucci, A.; Michaelson, J.S.; et al. Stability engineering of scFvs for the development of bispecific and multivalent antibodies. Protein Eng. Des. Sel. 2010, 23, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Xiang, S.; Feng, Y.; Srinivas, S.; Zhang, Y.; Lin, M.; Wang, S. Engineering production of functional scFv antibody in E. coli by co-expressing the molecule chaperone Skp. Front. Cell. Infect. Microbiol. 2013, 3, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Hamers-Casterman, C.; Atarhouch, T.; Muyldermans, S.; Robinson, G.; Hamers, C.; Songa, E.B.; Bendahman, N.; Hamers, R. Naturally occurring antibodies devoid of light chains. Lett. Nat. 1993, 363, 446–448. [Google Scholar] [CrossRef] [PubMed]
- Muyldermans, S. Nanobodies: Natural single-domain antibodies. Annu. Rev. Biochem. 2013, 82, 775–797. [Google Scholar] [CrossRef] [PubMed]
- Kolkman, J.A.; Law, D.A. Nanobodies—From llamas to therapeutic proteins. Drug Discov. Today Technol. 2010, 7, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Cortez-Retamozo, V.; Lauwereys, M.; Hassanzadeh, G.G.; Gobert, M.; Conrath, K.; Muyldermans, S.; De Baetselier, P.; Revets, H. Efficient tumor targeting by single-domain antibody fragments of camels. Int. J. Cancer 2002, 98, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Harmsen, M.M.; De Haard, H.J. Properties, production, and applications of camelid single-domain antibody fragments. Appl. Microbiol. Biotechnol. 2007, 77, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Arbabi-Ghahroudi, M.; Desmyter, A.; Wyns, L.; Hamers, R.; Muyldermans, S. Selection and identification of single domain antibody fragments from camel heavy-chain antibodies. FEBS Lett. 1997, 414, 521–526. [Google Scholar] [CrossRef]
- Warrell, D.A. Venomous Bites, Stings, and Poisoning. Infect. Dis. Clin. N. Am. 2012, 26, 207–223. [Google Scholar] [CrossRef] [PubMed]
- Thanongsaksrikul, J.; Srimanote, P.; Maneewatch, S.; Choowongkomon, K.; Tapchaisri, P.; Makino, S.; Kurazono, H.; Chaicumpa, W. A VHH that neutralizes the zinc metalloproteinase activity of botulinum neurotoxin type A. J. Biol. Chem. 2010, 285, 9657–9666. [Google Scholar] [CrossRef] [PubMed]
- Richard, G.; Meyers, A.J.; McLean, M.D.; Arbabi-Ghahroudi, M.; MacKenzie, R.; Hall, J.C. In vivo neutralization of a-cobratoxin with high-affinity Llama single domain antibodies (VHHs) and a VHH-Fc antibody. PLoS ONE 2013, 8, e69495. [Google Scholar] [CrossRef] [PubMed]
- Prado, N.D.; Pereira, S.S.; da Silva, M.P.; Morais, M.S.; Kayano, A.M.; Moreira-Dill, L.S.; Luiz, M.B.; Zanchi, F.B.; Fuly, A.L.; Huacca, M.E.; et al. Inhibition of the myotoxicity induced by Bothrops jararacussu venom and isolated phospholipases A2 by Specific Camelid SingleDomain Antibody Fragments. PLoS ONE 2016, 11, e0151363. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, A.R.; Marks, J.D. Antibodies from phage antibody libraries. J. Immunol. Methods 2004, 290, 29–49. [Google Scholar] [CrossRef] [PubMed]
- Dumoulin, M.; Conrath, K.; Van Meirhaeghe, A.; Meersman, F.; Heremans, K.; Frenken, L.G.; Muyldermans, S.; Wyns, L.; Matagne, A. Single-domain antibody fragments with high conformational stability. Protein Sci. 2002, 11, 500–515. [Google Scholar] [CrossRef] [PubMed]
- Govaert, J.; Pellis, M.; Deschacht, N.; Vincke, C.; Conrath, K.; Muyldermans, S.; Saerens, D. Dual beneficial effect of interloop disulfide bond for single domain antibody fragments. J. Biol. Chem. 2012, 287, 1970–1979. [Google Scholar] [CrossRef] [PubMed]
- Vu, K.B.; Arbabi-Ghahroudi, M.; Wyns, L.; Muyldermans, S. Comparison of llama VH sequences from conventional and heavy chain antibodies. Mol. Immunol. 1997, 34, 1121–1131. [Google Scholar] [CrossRef]
- Sampaio, S.C.; Hyslop, S.; Fontes, M.R.; Prado-Franceschi, J.; Zambelli, V.O.; Magro, A.J.; Brigatte, P.; Gutierrez, V.P.; Cury, Y. Crotoxin: Novel activities for a classic beta-neurotoxin. Toxicon 2010, 55, 1045–1060. [Google Scholar] [CrossRef] [PubMed]
- Landucci, E.C.; Antunes, E.; Donato, J.L.; Faro, R.; Hyslop, S.; Marangoni, S.; Oliveira, B.; Cirino, G.; de Nucci, G. Inhibition of carrageenin-induced rat paw oedema by crotapotin, a polypeptide complexed with phospholipase A2. Br. J. Pharmacol. 1995, 114, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Garcia, F.; Toyama, M.H.; Castro, F.R.; Proença, P.L.; Marangoni, S.; Santos, L.M. Crotapotin induced modification of T lymphocyte proliferative response through interference with PGE2 synthesis. Toxicon 2003, 42, 433–437. [Google Scholar] [CrossRef]
- Janssen, M.; Freyvogel, T.A.; Meier, J. Antigenic relationship between the venom of the nigth adder Causus maculatus and venoms of other viperids. Toxicon 1990, 28, 975–983. [Google Scholar] [CrossRef]
- Tan, C.H.; Tan, N.H.; Tan, K.Y.; Kwong, K.O. Antivenom cross-neutralization of the venoms of Hydrophis schistosus and Hydrophis curtus, two common sea snakes in Malaysian waters. Toxins 2015, 7, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Boldrini-França, J.; Corrêa-Netto, C.; Silva, M.M.; Rodrigues, R.S.; De La Torre, P.; Pérez, A.; Soares, A.M.; Zingali, R.B.; Nogueira, R.A.; Rodrigues, V.M.; et al. Snake venomics and antivenomics of Crotalus durissus subspecies from Brazil: Assessment of geographic variation and its implication on snakebite management. J. Proteom. 2010, 73, 1758–1776. [Google Scholar] [CrossRef] [PubMed]
- Marchi-Salvador, D.P.; Corrêa, L.C.; Magro, A.J.; Oliveira, C.Z.; Soares, A.M.; Fontes, M.R. Insights into the role of oligomeric state on the biological activities of crotoxin: Crystal structure of a tetrameric phospholipase A2 formed by two isoforms of crotoxin B from Crotalus durissus terrificus venom. Proteins 2008, 72, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Baral, T.N.; Murad, Y.; Nguyen, T.D.; Iqbal, U.; Zhang, J. Isolation of functional single domain antibody by whole cell immunization: Implications for cancer treatment. J. Immunol. Methods 2011, 371, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Olichon, A.; Schweizer, D.; Muyldermans, S.; de Marco, A. Heating as a rapid purification method for recovering correctly-folded thermotolerant VH and VHH domains. BMC Biotechnol. 2007, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tabares-da Rosa, S.; Rossotti, M.; Carleiza, C.; Carrión, F.; Pritsch, O.; Ahn, K.C.; Last, J.A.; Hammock, B.D.; González-Sapienza, G. Competitive selection from single domain antibody libraries allows isolation of high-affinity antihapten antibodies that are not favored in the llama immune response. Anal. Chem. 2011, 11, 7213–7220. [Google Scholar] [CrossRef] [PubMed]
- Omidfar, K.; Rasaee, M.J.; Kashanian, S.; Paknejad, M.; Bathaie, Z. Studies of thermostability in Camelus bactrianus (Bactrian camel) single-domain antibody specific for the mutant epidermal-growth-factor receptor expressed by Pichia. Biotechnol. Appl. Biochem. 2007, 46, 41–49. [Google Scholar] [PubMed]
- Van der Linden, R.H.; Frenken, L.G.; de Geus, B.; Harmsen, M.M.; Ruuls, R.C.; Stok, W.; de Ron, L.; Wilson, S.; Davis, P.; Verrips, C.T. Comparison of physical chemical properties of llama VHH antibody fragments and mouse monoclonal antibodies. Biochim. Biophys. Acta 1999, 1431, 37–46. [Google Scholar] [CrossRef]
- Hussack, G.; Arbabi-Ghahroudi, M.; van Faassen, H.; Songer, J.G.; Ng, K.K.; MacKenzie, R.; Tanha, J. Neutralization of Clostridium difficile toxin A with single-domain antibodies targeting the cell receptor binding domain. J. Biol. Chem. 2011, 286, 8961–8976. [Google Scholar] [CrossRef] [PubMed]
- Hufton, S.E.; Risley, P.; Ball, C.R.; Major, D.; Engelhardt, O.G.; Poole, S. The breadth of cross sub-type neutralisation activity of a single domain antibody to influenza hemagglutinin can be increased by antibody valency. PLoS ONE 2014, 9, e103294. [Google Scholar] [CrossRef] [PubMed]
- Choumet, V.; Jiang, M.S.; Radvanyi, F.; Ownby, C.; Bon, C. Neutralization of lethal potency and inhibition of enzymatic activity of a phospholipase A2 neurotoxin, crotoxin, by non-precipitating antibodies (Fab). FEBS Lett. 1989, 244, 167–173. [Google Scholar] [CrossRef]
- Oliveira, J.G.; Soares, S.G.; Soares, A.M.; Giglio, J.R.; Teixeira, J.E.; Barbosa, J.E. Expression of human recombinant antibody fragments capable of partially inhibiting the phospholypase activity of Crotalus durissus terrificus venom. Basic Clin. Pharmacol. Toxicol. 2009, 105, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Ponce-Soto, L.A.; Lomonte, B.; Rodrigues-Simioni, L.; Novello, J.C.; Marangoni, S. Biological and structural characterization of crotoxin and new isoform of crotoxin B PLA2 (F6a) from Crotalus durissus collilineatus snake venom. Protein J. 2007, 26, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Lomonte, B.; Angulo, Y.; Rufini, S.; Cho, W.; Giglio, J.R.; Ohno, M.; Daniele, J.J.; Geoghegan, P.; Gutiérrez, J.M. Comparative study of the cytolytic activity of myotoxic phospholipases A2 on mouse endothelial (tEnd) and skeletal muscle (C2C12) cells in vitro. Toxicon 1999, 37, 145–158. [Google Scholar] [CrossRef]
- Soares, A.M.; Mancin, A.C.; Cecchini, A.L.; Arantes, E.C.; França, S.C.; Gutiérrez, J.M.; Giglio, J.R. Effects of chemical modifications of crotoxin B, the phospholipase A2 subunit of crotoxin from Crotalus durissus terrificus snake venom, on its enzymatic and pharmacological activities. Int. J. Biochem. Cell Biol. 2001, 33, 877–888. [Google Scholar] [CrossRef]
- Fernandes, C.A.; Pazin, W.M.; Dreyer, T.R.; Bicev, R.N.; Cavalcante, W.L.; Fortes-Dias, C.L.; Ito, A.S.; Oliveira, C.L.; Fernandez, R.M.; Fontes, M.R. Biophysical studies suggest a new structural arrangement of crotoxin and provide insights into its toxic mechanism. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Curin-Serbec, V.; Délot, E.; Faure, G.; Saliou, B.; Gubensek, F.; Bon, C.; Choumet, V. Antipeptide antibodies directed to the c-terminal part of ammodytoxin a react with the PLA2 subunit of crotoxin and neutralize its pharmacological activity. Toxicon 1994, 32, 1337–1348. [Google Scholar] [CrossRef]
- Azevedo-Marques, M.M.; Cupo, P.; Coimbra, T.M.; Hering, S.E.; Rossi, M.A.; Laure, C.J. Myonecrosis, myoglobinuria and acute renal failure induced by south american rattlesnake (Crotalus durissus terrificus) envenomation in Brazil. Toxicon 1985, 23, 631–636. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Ponce-Soto, L.A.; Marangoni, S.; Lomonte, B. Systemic and local myotoxicity induced by snake venom group II phospholipases A2: Comparison between crotoxin, crotoxin B and a Lys49 PLA2 homologue. Toxicon 2008, 51, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Amaral, C.F.; de Rezende, N.A.; da Silva, O.A.; Ribeiro, M.M.; Magalhães, R.A.; dos Reis, R.J.; Carneiro, J.G.; Castro, J.R. Insuficiência renal aguda secundária a acidentes ofídicos botrópico e crotálico. Análise de 63 casos. Rev. Inst. Med. Trop. Sao Paulo 1986, 28, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Jorge, M.T.; Ribeiro, L.A. The epidemiology and clinical picture of an accidental bite by the South American rattlesnake (Crotalus durissus). Rev. Inst. Med. Trop. Sao Paulo 1992, 34, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.S.; Moreira-Dill, L.S.; Morais, M.S.; Prado, N.D.; Barros, M.L.; Koishi, A.C.; Mazarrotto, G.A.; Gonçalves, G.M.; Zuliani, J.P.; Calderon, L.A.; et al. Novel camelid antibody fragments targeting recombinant nucleoprotein of Araucaria hantavirus: A prototype for an early diagnosis of hantavirus pulmonary syndrome. PLoS ONE 2014, 9, e108067. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Andersson, K.; Hämäläinen, M.; Malmqvist, M. Identification and optimization of regeneration conditions for affinity- based biosensor assays. A multivariate cocktail approach. Anal. Chem. 1999, 71, 2475–2481. [Google Scholar] [CrossRef] [PubMed]
- Lomonte, B.; Tarkowski, A.; Bagge, U.; Hanson, L.A. Neutralization of the cytolytic and myotoxic activities of phospholipases A2 from Bothrops asper snake venom by glycosaminoglycans of the heparina/heparan sulfate family. Biochem. Pharmacol. 1994, 47, 1509–1518. [Google Scholar] [CrossRef]
- Ebisui, C.; Tsujinaka, T.; Morimoto, T.; Kan, K.; Iijima, S.; Yano, M.; Kominami, E.; Tanaka, K.; Monden, M. Interleukin-6 induces proteolysis by activating intracellular proteases (cathepsins B and L, proteasome) in C2C12 myotubes. Clin. Sci. (Lond.) 1995, 89, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Li, L.; Weng, Z. Zdock: An initial-stage protein docking algorithm. Proteins 2003, 1, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Brenke, R.; Hall, D.R.; Chuang, G.Y.; Comeau, S.R.; Bohnuud, T.; Beglov, D.; Schueler-Furman, O.; Vajda, S.; Kozakov, D. Application of asymmetric statistical potentials to antibody-protein docking. Bioinformatics 2003, 28, 2608–2614. [Google Scholar] [CrossRef] [PubMed]
- Kozakov, D.; Brenke, R.; Comeau, S.R.; Vajda, S. PIPER: An FFT-based protein pocking program with pairwise potentials. Proteins 2006, 65, 392–406. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clone | VHH Domain | Amino Acids | CTX | Distances | |
|---|---|---|---|---|---|
| Antibody | Antigen | Domain | (Å) | ||
| KF498602 | FR2 | Arg-47 | Glu-12 | N-terminal α-helix A (CB) | 1.8/1.9/1.8 |
| CDR3 | Trp-117 | Glu-12 | N-terminal α-helix A (CB) | 2 | |
| FR2 | Arg-47 | Arg-14 | N-terminal α-helix A (CB) | 1.9 | |
| FR2 | Arg-47 | Phe-11 | N-terminal α-helix A (CB) | 1.7 | |
| CDR3 | Ser-115 | Arg-97 | α-helix D (CB) | 2.1 | |
| CDR3 | Tyr-114 | Arg-97 | α-helix D (CB) | 1.8 | |
| CDR3 | Trp-110 | Arg-14 | N-terminal α-helix A (CB) | 1.8 | |
| FR2 | Glu-46 | Lys-7 | N-terminal α-helix A (CB) | 1.7 | |
| CDR3 | Asp-112 | Gln-33 | α-Chain (CA) | 2.0/2.0 | |
| FR2 | Glu-46 | Arg-14 | N-terminal α-helix A (CB) | 1.8 | |
| KF498603 | CDR3 | Asp-112 | Arg-97 | α-helix D (CB) | 1.8/1.8/1.9 |
| CDR3 | Asp-112 | Arg-14 | N-terminal α-helix A (CB) | 1.9 | |
| CDR3 | Tyr-114 | Arg-14 | N-terminal α-helix A (CB) | 1.7 | |
| CDR3 | Tyr-111 | Gly-32 | Ca2+ binding loop (CB) | 1.8 | |
| FR2 | Arg-47 | Gly-31 | Ca2+ binding loop (CB) | 1.7/2.2 | |
| FR2 | Arg-47 | Gln-37 | Loop (CB) | 1.9 | |
| FR1 | Ala-2 | Lys-z-7 | N-terminal α-helix A (CB) | 1.7 | |
| FR1 | Glu-3 | Lys-7 | N-terminal α-helix A (CB) | 1.7 | |
| KF498604 | FR3 | Asp-64 | Cys-75 | β-wing (CB) | 2.1 |
| CDR2 | Arg57 | Gly-52 | α-helix C (CB) | 1.7 | |
| CDR3 | Tyr-117 | Gly117 | C-terminal extension (CB) | 1.8 | |
| FR3 | Try-62 | Asp78 | β-wing (CB) | 1.9/2.5 | |
| CDR2 | Thr-60 | Asp78 | β-wing (CB) | 2 | |
| CDR3 | Glu-103 | Arg-116 | C-terminal extension (CB) | 2.0/1.9 | |
| CDR3 | Gly-104 | Arg-116 | C-terminal extension (CB) | 1.7 | |
| CDR1 | Ala-35 | Ser-119 | C-terminal extension (CB) | 2.2 | |
| KF498605 | CDR2 | Arg-57 | Asp-78 | β-Chain (CA) | 1.8/1.8/2.0 |
| CDR1 | Asn-33 | Thr-121 | C-terminal extension (CB) | 1.8 | |
| CDR3 | Asp-102 | Arg-116 | C-terminal extension (CB) | 1.9 | |
| CDR3 | Ser-101 | Arg-116 | C-terminal extension (CB) | 2 | |
| CDR3 | Glu-103 | Arg-116 | C-terminal extension (CB) | 1.2/1.8/1.8 | |
| FR3 | Asp-64 | Lys-90 | β-Chain (CA) | 1.8 | |
| FR3 | Lyz67 | Glu-87 | β-Chain (CA) | 1.7/1.8 | |
| FR3 | Tyr-62 | Cys-75 | β-Chain (CA) | 2 | |
| KF498606 | FR1 | Glu-3 | Lys-7 | N-terminal α-helix A (CB) | 1.7 |
| CDR3 | Arg-99 | Ala-52 | α-Chain (CA) | 1.9 | |
| CDR3 | Arg-99 | Tyr-51 | α-Chain (CA) | 1.7/2.0 | |
| CDR3 | Arg-99 | Cys-57 | α-Chain (CA) | 1.9/2.4 | |
| CDR3 | Arg-99 | Leu-54 | α-Chain (CA) | 1.8 | |
| FR2 | Arg-47 | Gly-56 | α-Chain (CA) | 1.9 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luiz, M.B.; Pereira, S.S.; Prado, N.D.R.; Gonçalves, N.R.; Kayano, A.M.; Moreira-Dill, L.S.; Sobrinho, J.C.; Zanchi, F.B.; Fuly, A.L.; Fernandes, C.F.; et al. Camelid Single-Domain Antibodies (VHHs) against Crotoxin: A Basis for Developing Modular Building Blocks for the Enhancement of Treatment or Diagnosis of Crotalic Envenoming. Toxins 2018, 10, 142. https://doi.org/10.3390/toxins10040142
Luiz MB, Pereira SS, Prado NDR, Gonçalves NR, Kayano AM, Moreira-Dill LS, Sobrinho JC, Zanchi FB, Fuly AL, Fernandes CF, et al. Camelid Single-Domain Antibodies (VHHs) against Crotoxin: A Basis for Developing Modular Building Blocks for the Enhancement of Treatment or Diagnosis of Crotalic Envenoming. Toxins. 2018; 10(4):142. https://doi.org/10.3390/toxins10040142
Chicago/Turabian StyleLuiz, Marcos B., Soraya S. Pereira, Nidiane D. R. Prado, Naan R. Gonçalves, Anderson M. Kayano, Leandro S. Moreira-Dill, Juliana C. Sobrinho, Fernando B. Zanchi, André L. Fuly, Cleberson F. Fernandes, and et al. 2018. "Camelid Single-Domain Antibodies (VHHs) against Crotoxin: A Basis for Developing Modular Building Blocks for the Enhancement of Treatment or Diagnosis of Crotalic Envenoming" Toxins 10, no. 4: 142. https://doi.org/10.3390/toxins10040142
APA StyleLuiz, M. B., Pereira, S. S., Prado, N. D. R., Gonçalves, N. R., Kayano, A. M., Moreira-Dill, L. S., Sobrinho, J. C., Zanchi, F. B., Fuly, A. L., Fernandes, C. F., Zuliani, J. P., Soares, A. M., Stabeli, R. G., & Fernandes, C. F. C. (2018). Camelid Single-Domain Antibodies (VHHs) against Crotoxin: A Basis for Developing Modular Building Blocks for the Enhancement of Treatment or Diagnosis of Crotalic Envenoming. Toxins, 10(4), 142. https://doi.org/10.3390/toxins10040142