Antivenom Production against Bothrops jararaca and Bothrops erythromelas Snake Venoms Using Cross-Linked Chitosan Nanoparticles as an Immunoadjuvant

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
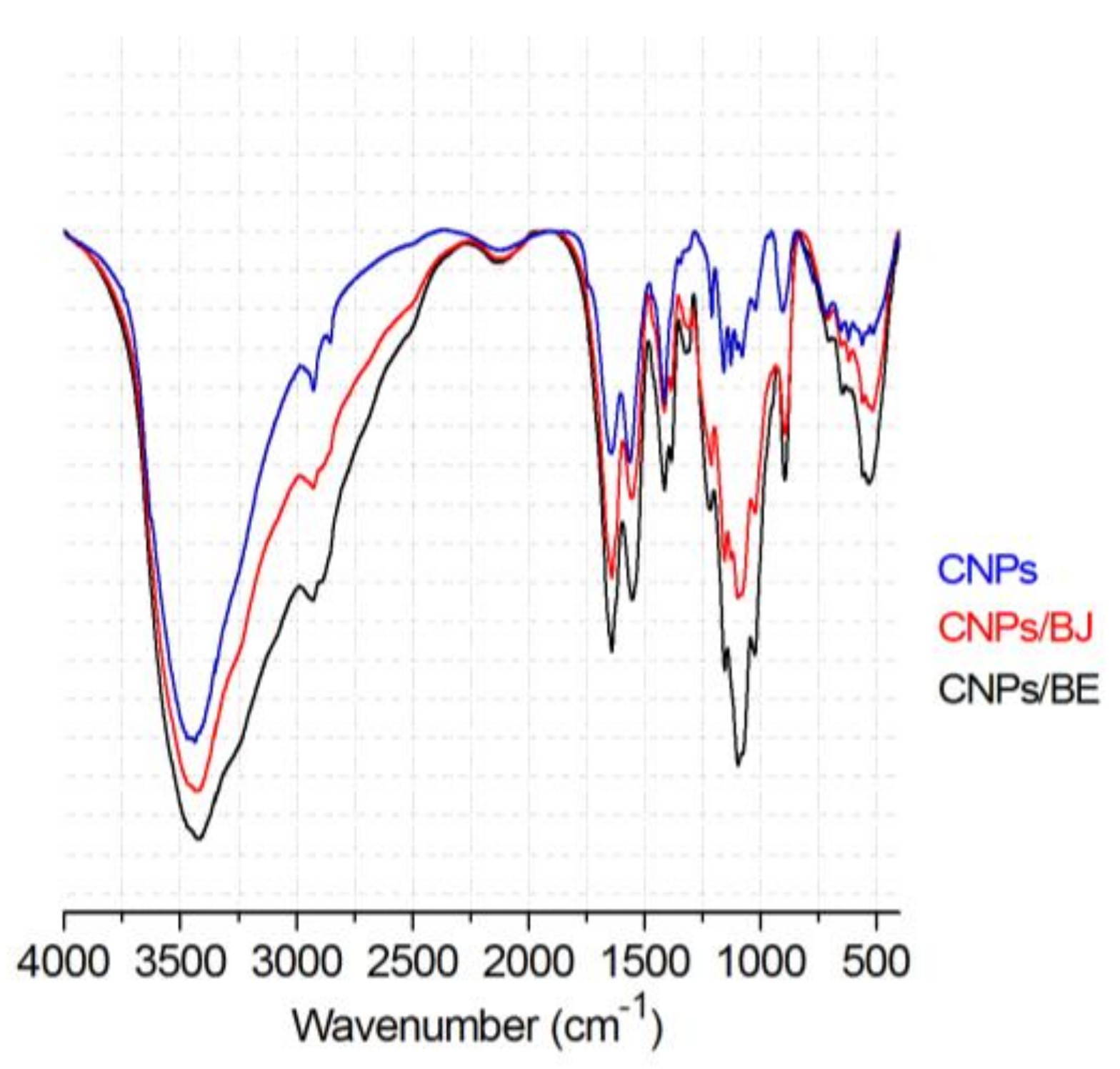
2.1. Preparation of Chitosan Nanoparticles
2.2. Protein Loading Efficiency of the Bothrops jararaca and Bothrops erythromelas Venoms
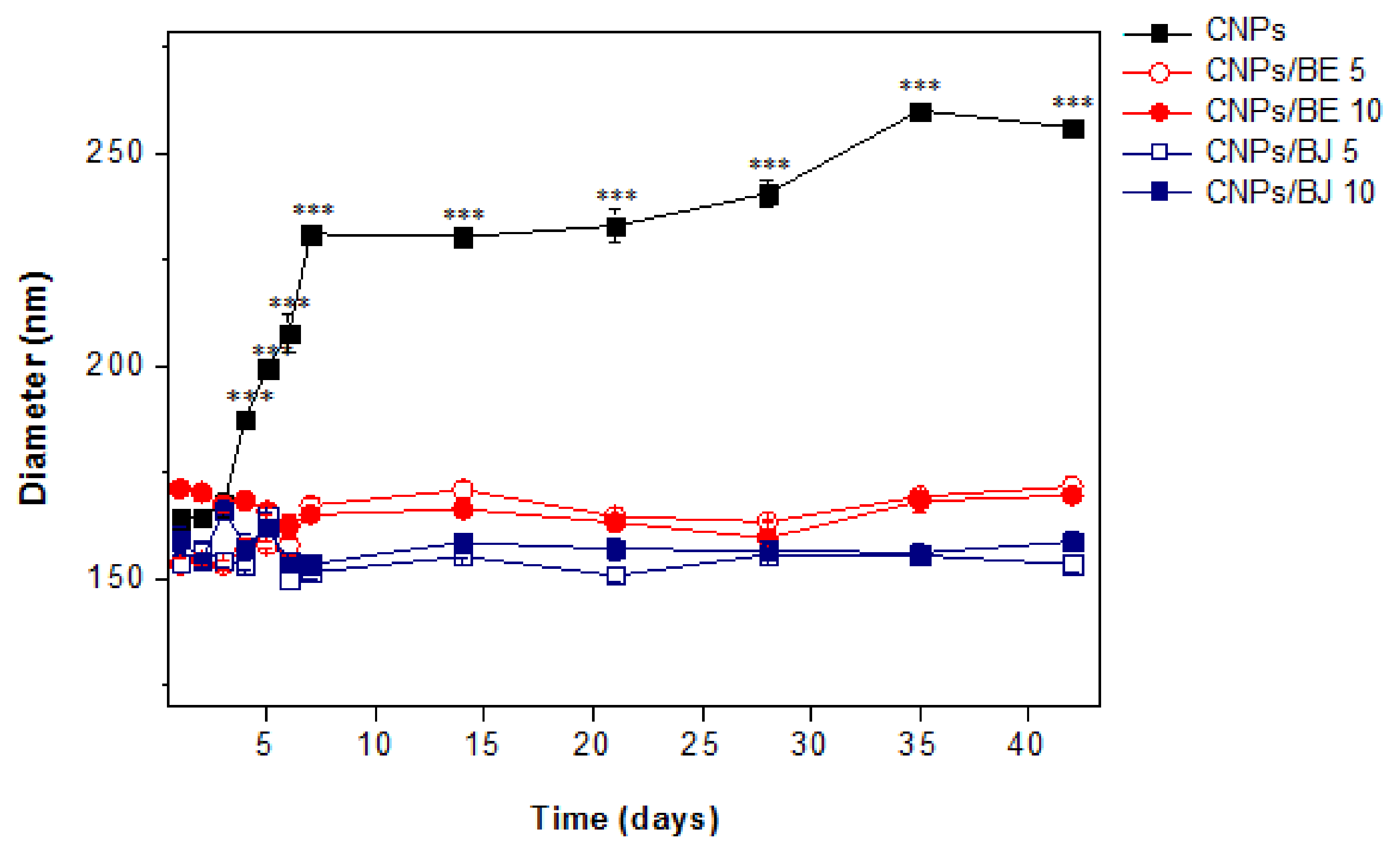
2.3. Stability Assay
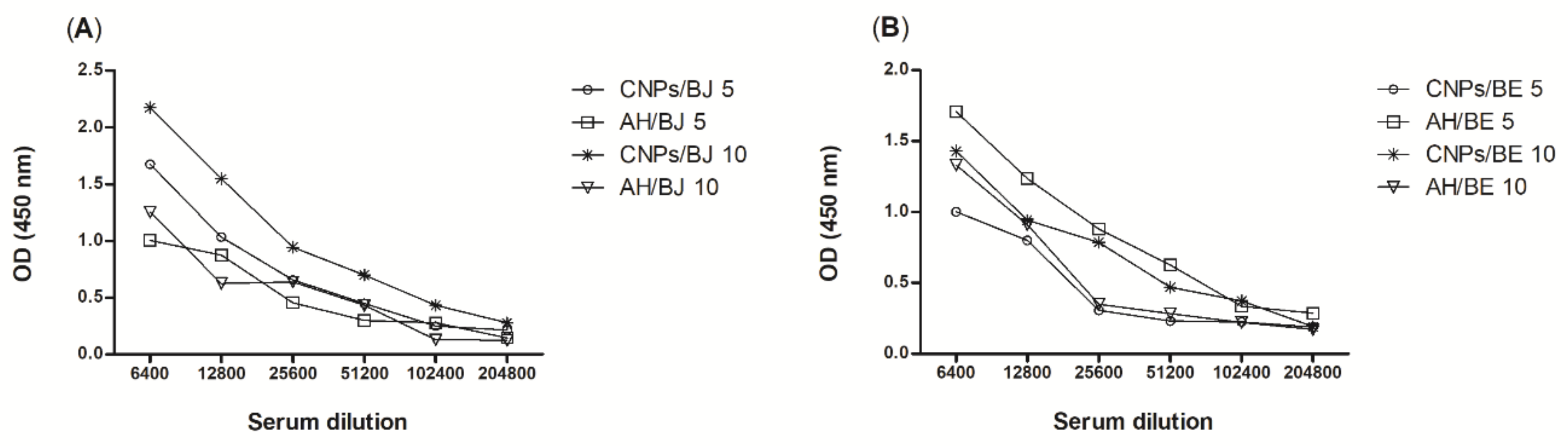
2.4. Antibody Titres
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cross-Linked Chitosan Nanoparticles
4.3. Loading of the B. jararaca and B. erythromelas Venoms into Cross-Linked Chitosan Nanoparticles
4.4. Protein-Loading Efficiency Assay
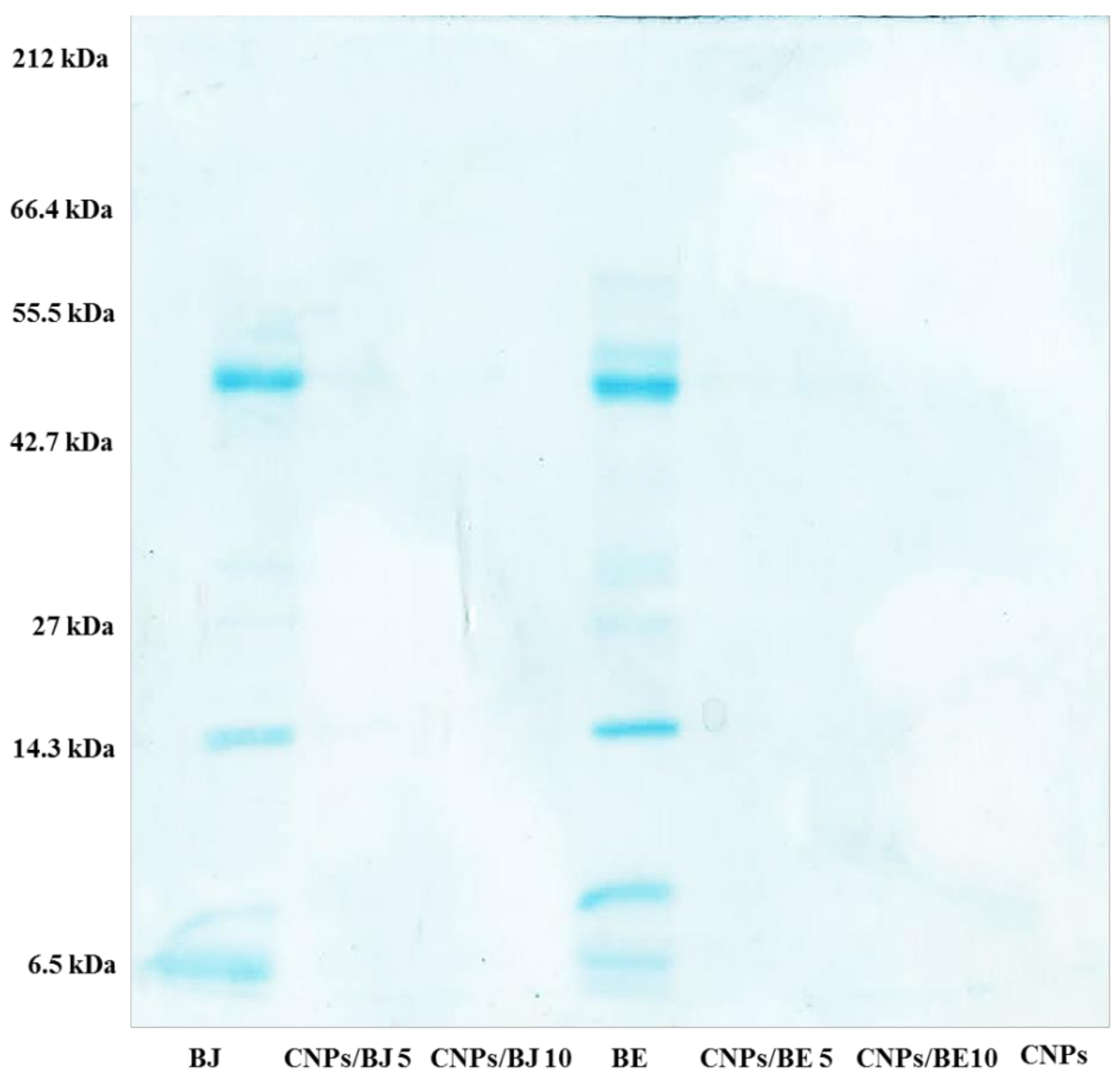
4.5. Electrophoresis
4.6. Physicochemical Properties of the Nanoparticles
4.7. Stability Assay
4.8. Animals
4.9. Immunization
4.10. Serum Production
4.11. Serum Antibody Responses
4.12. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dharmadasa, R.M.; Akalanka, G.C.; Muthukumarana, P.R.M.; Wijesekara, R.G.S. Ethnopharmacological survey on medicinal plants used in snakebite treatments in Western and Sabaragamuwa provinces in Sri Lanka. J. Ethnopharmacol. 2016, 179, 110–127. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.M.; Lomonte, B.; Sanz, L.; Calvete, J.J.; Pla, D. Immunological profile of antivenoms: Preclinical analysis of the efficacy of a polyspecific antivenom through antivenomics and neutralization assays. J. Proteom. 2014, 105, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Karabuva, S.; Vrkić, I.; Brizić, I.; Ivić, I.; Lukšić, B. Venomous snakebites in children in southern Croatia. Toxicon 2016, 112, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Kasturiratne, A.; Wickremasinghe, A.R.; De Silva, N.; Gunawardena, N.K.; Pathmeswaran, A.; Premaratna, R.; Savioli, L.; Lalloo, D.G.; De Silva, H.J. The global burden of snakebite: A literature analysis and modelling based on regional estimates of envenoming and deaths. PLoS Med. 2008, 5, 1591–1604. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves-Machado, L.; Pla, D.; Sanz, L.; Jorge, R.J.B.; Leitão-De-Araújo, M.; Alves, M.L.M.; Alvares, D.J.; De Miranda, J.; Nowatzki, J.; de Morais-Zani, K.; et al. Combined venomics, venom gland transcriptomics, bioactivities, and antivenomics of two Bothrops jararaca populations from geographic isolated regions within the Brazilian Atlantic rainforest. J. Proteom. 2016, 135, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Fenwick, A.M.; Greene, H.W.; Parkinson, C.L. The serpent and the egg: Unidirectional evolution of reproductive mode in vipers? J. Zool. Syst. Evol. Res. 2012, 50, 59–66. [Google Scholar] [CrossRef]
- Oliveira, F.N.; Brito, M.T.; de Morais, I.C.O.; Fook, S.M.L.; de Albuquerque, H.N. Accidents caused by Bothrops and Bothropoides in the State of Paraiba: Epidemiological and clinical aspects. Rev. Soc. Bras. Med. Trop. 2010, 43, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Lira-da-silva, R.M.; Mise, Y.F.; Casais-e-silva, L.L.; Ulloa, J.; Hamdan, B.; Brazil, T.K. Serpentes de importância médica do nordeste do Brasil. Gazeta Médica da Bahia 2009, 79, 7–20. [Google Scholar]
- Delafontaine, M.; Villas-Boas, I.M.; Mathieu, L.; Josset, P.; Blomet, J.; Tambourgi, D.V. Enzymatic and pro-inflammatory activities of Bothrops lanceolatus venom: Relevance for envenomation. Toxins (Basel) 2017, 9, 244. [Google Scholar] [CrossRef] [PubMed]
- Jorge, R.J.B.; Monteiro, H.S.A.; Gonçalves-Machado, L.; Guarnieri, M.C.; Ximenes, R.M.; Borges-Nojosa, D.M.; Luna, K.P.d.O.; Zingali, R.B.; Corrêa-Netto, C.; Gutiérrez, J.M.; et al. Venomics and antivenomics of Bothrops erythromelas from five geographic populations within the Caatinga ecoregion of northeastern Brazil. J. Proteom. 2015, 114, 93–114. [Google Scholar] [CrossRef] [PubMed]
- Zychar, B.C.; Dale, C.S.; Demarchi, D.S.; Gonçalves, L.R.C. Contribution of metalloproteases, serine proteases and phospholipases A2 to the inflammatory reaction induced by Bothrops jararaca crude venom in mice. Toxicon 2010, 55, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.M.; Lomonte, B. Efectos Locales en el Envenenamiento Ofídico en America Latina. In Animais Peçonhentos No Brasil; Sarvier: Sao Paulo, Brazil, 2003; pp. 310–323. [Google Scholar] [CrossRef]
- Freitas-de-sousa, L.A.; Colombini, M.; Lopes-Ferreira, M.; Serrano, S.M.T.; Moura-da-silva, A.M. Insights into the mechanisms involved in strong hemorrhage and dermonecrosis induced by atroxlysin-Ia, a PI-class snake venom metalloproteinase. Toxins (Basel) 2017, 9, 239. [Google Scholar] [CrossRef] [PubMed]
- Bjarnason, J.B.; Fox, J.W. Hemorrhagic Metalloproteinases from Snake-Venoms. Pharmacol. Ther. 1994, 62, 325–372. [Google Scholar] [CrossRef]
- Serrano, S.M.T.; Maroun, R.C. Snake venom serine proteinases: Sequence homology vs. substrate specificity, a paradox to be solved. Toxicon 2005, 45, 1115–1132. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, F.B.; Gomes, M.S.R.; de Souza, D.L.N.; Gimenes, S.N.C.; Castanheira, L.E.; Borges, M.H.; Rodrigues, R.S.; Yoneyama, K.A.G.; Brandeburgo, M.I.H.; Rodrigues, V.M. Molecular cloning and pharmacological properties of an acidic PLA2 from Bothrops pauloensis snake venom. Toxins (Basel) 2013, 5, 2403–2419. [Google Scholar] [CrossRef] [PubMed]
- Oguiura, N.; Kapronezai, J.; Ribeiro, T.; Rocha, M.M.T.; Medeiros, C.R.; Marcelino, J.R.; Prezoto, B.C. An alternative micromethod to access the procoagulant activity of Bothrops jararaca venom and the efficacy of antivenom. Toxicon 2014, 90, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Espino-Solis, G.P.; Riaño-Umbarila, L.; Becerril, B.; Possani, L.D. Antidotes against venomous animals: State of the art and prospectives. J. Proteom. 2009, 72, 183–199. [Google Scholar] [CrossRef] [PubMed]
- Alvarenga, L.M.; Zahid, M.; Tommaso, A.D.; Juste, M.O.; Aubrey, N.; Billiald, P.; Muzard, J. Engineering venom’s toxin-neutralizing antibody fragments and its therapeutic potential. Toxins (Basel) 2014, 6, 2541–2567. [Google Scholar] [CrossRef] [PubMed]
- Faioli, C.N.; Domingos, T.F.S.; de Oliveira, E.C.; Sanchez, E.F.; Ribeiro, S.; Muricy, G.; Fuly, A.L. Appraisal of antiophidic potential of marine sponges against Bothrops jararaca and Lachesis muta venom. Toxins (Basel) 2013, 5, 1799–1813. [Google Scholar] [CrossRef] [PubMed]
- Rocha Soares, K.S.; Fonseca, J.L.C.; Bitencourt, M.A.O.; Santos, K.S.C.R.; Silva-Júnior, A.A.; Fernandes-Pedrosa, M.F. Serum production against Tityus serrulatus scorpion venom using cross-linked chitosan nanoparticles as immunoadjuvant. Toxicon 2012, 60, 1349–1354. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.K.; Siber, G.R. Adjuvants for human vaccines-current status, problems and future prospects. Vaccine 1995, 13, 1263–1276. [Google Scholar] [CrossRef]
- Li, X.; Aldayel, A.M.; Cui, Z. Aluminum hydroxide nanoparticles show a stronger vaccine adjuvant activity than traditional aluminum hydroxide microparticles. J. Control. Release 2014, 173, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Gherardi, R.K. Lessons from macrophagic myofasciitis: Towards definition of a vaccine adjuvant-related syndrome. Rev. Neurol. (Paris) 2003, 159, 162–164. [Google Scholar] [CrossRef] [PubMed]
- Petrovsky, N.; Aguilar, J.C. Special Feature Vaccine adjuvants: Current state and future trends. Immunol. Cell Biol. 2004, 82, 488–496. [Google Scholar] [CrossRef] [PubMed]
- prasanth Koppolu, B.; Smith, S.G.; Ravindranathan, S.; Jayanthi, S.; Suresh Kumar, T.K.; Zaharoff, D.A. Controlling chitosan-based encapsulation for protein and vaccine delivery. Biomaterials 2014, 35, 4382–4389. [Google Scholar] [CrossRef] [PubMed]
- Zaharoff, D.A.; Rogers, C.J.; Hance, K.W.; Schlom, J.; Greiner, J.W. Chitosan solution enhances the immunoadjuvant properties of GM-CSF. Vaccine 2007, 25, 8673–8686. [Google Scholar] [CrossRef] [PubMed]
- Bergfors, E.; Trollfors, B.; Inerot, A. Unexpectedly high incidence of persistent itching nodules and delayed hypersensitivity to aluminium in children after the use of adsorbed vaccines from a single manufacturer. Vaccine 2003, 22, 64–69. [Google Scholar] [CrossRef]
- Bansal, V.; Kumar, M.; Dalela, M.; Brahmne, H.G.; Singh, H. Evaluation of synergistic effect of biodegradable polymeric nanoparticles and aluminum based adjuvant for improving vaccine efficacy. Int. J. Pharm. 2014, 471, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, J.C.; Rodriguez, E.G. Vaccine adjuvants revisited. Vaccine 2007, 25, 3752–3762. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.K.; Rost, B.E.; Relyveld, E.; Siber, G.R. Adjuvant Properties of Aluminum and Calcium Compounds. Pharm Biotechnol. 2000, 6, 229–248. [Google Scholar] [CrossRef]
- Nait Mohamed, F.A.; Laraba-Djebari, F. Development and characterization of a new carrier for vaccine delivery based on calcium-alginate nanoparticles: Safe immunoprotective approach against scorpion envenoming. Vaccine 2016, 34, 2692–2699. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Mohanty, S. Nanoparticle formulation having ability to control the release of protein for drug delivery application. Mater. Sci. Eng. C 2017, 70, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Du, Y. Effect of molecular structure of chitosan on protein delivery properties of chitosan nanoparticles. Int. J. Pharm. 2003, 250, 215–226. [Google Scholar] [CrossRef]
- Janes, K.A.; Calvo, P.; Alonso, M.J. Polysaccharide colloidal particles as delivery systems for macromolecules. Adv. Drug Deliv. Rev. 2001, 47, 83–97. [Google Scholar] [CrossRef]
- Mohammadpourdounighi, N.; Behfar, A.; Ezabadi, A.; Zolfagharian, H.; Heydari, M. Preparation of chitosan nanoparticles containing Naja naja oxiana snake venom, Nanomedicine Nanotechnology. Biol. Med. 2010, 6. [Google Scholar] [CrossRef]
- McClements, D.J. Encapsulation, protection, and delivery of bioactive proteins and peptides using nanoparticle and microparticle systems: A review. Adv. Colloid Interface Sci. 2018, 253, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Calvo, P.; Remunan-Lopez, C. Novel hydrophilic chitosan-polyethylene oxide nanoparticles as protein carriers. J. Appl. Polym. Sci. 1997, 63, 125–132. [Google Scholar] [CrossRef]
- Sinha, R. Nanotechnology in cancer therapeutics: bioconjugated nanoparticles for drug delivery. Mol. Cancer Ther. 2006, 5, 1909–1917. [Google Scholar] [CrossRef] [PubMed]
- Scherließ, R.; Buske, S.; Young, K.; Weber, B.; Rades, T.; Hook, S. In vivo evaluation of chitosan as an adjuvant in subcutaneous vaccine formulations. Vaccine 2013, 31, 4812–4819. [Google Scholar] [CrossRef] [PubMed]
- Rampino, A.; Borgogna, M.; Blasi, P.; Bellich, B.; Cesàro, A. Chitosan nanoparticles: Preparation, size evolution and stability. Int. J. Pharm. 2013, 455, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Borges, O.; Silva, M.; de Sousa, A.; Borchard, G.; Junginger, H.E.; Cordeiro-da-Silva, A. Alginate coated chitosan nanoparticles are an effective subcutaneous adjuvant for hepatitis B surface antigen. Int. Immunopharmacol. 2008, 8, 1773–1780. [Google Scholar] [CrossRef] [PubMed]
- Ramesh Kumar, D.; Saravana Kumar, P.; Gandhi, M.R.; Al-Dhabi, N.A.; Paulraj, M.G.; Ignacimuthu, S. Delivery of chitosan/dsRNA nanoparticles for silencing of wing development vestigial (vg) gene in Aedes aegypti mosquitoes. Int. J. Biol. Macromol. 2016, 86, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Xu, Z.; Jiang, X.; Hu, C.; Zou, X. Preparation and antibacterial activity of chitosan nanoparticles. Carbohydr. Res. 2004, 339, 2693–2700. [Google Scholar] [CrossRef] [PubMed]
- Anandhakumar, S.; Krishnamoorthy, G.; Ramkumar, K.M.; Raichur, A.M. Preparation of collagen peptide functionalized chitosan nanoparticles by ionic gelation method: An effective carrier system for encapsulation and release of doxorubicin for cancer drug delivery. Mater. Sci. Eng. C 2017, 70, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Tavares, I.S.; Caroni, A.L.P.F.; Neto, A.A.D.; Pereira, M.R.; Fonseca, J.L.C. Surface charging and dimensions of chitosan coacervated nanoparticles. Colloids Surf. B Biointerfaces 2012, 90, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Gan, Q.; Wang, T.; Cochrane, C.; McCarron, P. Modulation of surface charge, particle size and morphological properties of chitosan-TPP nanoparticles intended for gene delivery. Colloids Surf. B Biointerfaces 2005, 44, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Zhou, N.J.; Gong, Y.F.; Zhou, X.J.; Chen, J.; Hu, S.J.; Lu, N.H.; Hou, X.H. Th immune response induced by H. pylori vaccine with chitosan as adjuvant and its relation to immune protection. World J. Gastroenterol. 2007, 13, 1547–1553. [Google Scholar] [CrossRef] [PubMed]
- Zhiming, H.; Sinha, R.; McNeela, E.A.; Borrow, R.; Giemza, R.; Cosgrove, C.; Heath, P.T.; Mills, K.H.G.; Rappuoli, R.; Griffin, G.E.; et al. Induction of protective serum meningococcal bactericidal and diphtheria-neutralizing antibodies and mucosal immunoglobulin A in volunteers by nasal insufflations of the Neisseria meningitidis serogroup C polysaccharide-CRM197 conjugate vaccine mixed with. Infect. Immun. 2005, 73, 8256–8265. [Google Scholar] [CrossRef]
- Prego, C.; Paolicelli, P.; Díaz, B.; Vicente, S.; Sánchez, A.; González-Fernández, Á.; Alonso, M.J. Chitosan-based nanoparticles for improving immunization against hepatitis B infection. Vaccine 2010, 28, 2607–2614. [Google Scholar] [CrossRef] [PubMed]
- Gan, Q.; Wang, T. Chitosan nanoparticle as protein delivery carrier-Systematic examination of fabrication conditions for efficient loading and release. Colloids Surf. B Biointerfaces 2007, 59, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Li, Y.J.; Zhao, H.Y.; Zheng, J.M.; Xu, H.; Wei, G.; Hao, J.S.; De Cui, F. Bioadhesive polysaccharide in protein delivery system: Chitosan nanoparticles improve the intestinal absorption of insulin in vivo. Int. J. Pharm. 2002, 249, 139–147. [Google Scholar] [CrossRef]
- Antoniou, J.; Liu, F.; Majeed, H.; Qi, J.; Yokoyama, W.; Zhong, F. Aspects Physicochemical and morphological properties of size-controlled chitosan—Tripolyphosphate nanoparticles. Colloids Surf. A Physicochem. Eng. 2015, 465, 137–146. [Google Scholar] [CrossRef]
- Rocha Soares, K.S.; AOliveira, R.; Daniele-Silva, A.; Glaucia-Silva, F.; Caroni, A.L.P.; Fernandes-Pedrosa, M.F.; da Silva-Júnior, A.A. Self-assembled scorpion venom proteins cross-linked chitosan nanoparticles for use in the immunotherapy. J. Mol. Liq. 2017, 241, 540–548. [Google Scholar] [CrossRef]
- Papadimitriou, S.; Bikiaris, D.; Avgoustakis, K.; Karavas, E.; Georgarakis, M. Chitosan nanoparticles loaded with dorzolamide and pramipexole. Carbohydr. Polym. 2008, 73, 44–54. [Google Scholar] [CrossRef]
- Rodrigues, S.; Da Costa, A.M.R.; Grenha, A. Chitosan/carrageenan nanoparticles: Effect of cross-linking with tripolyphosphate and charge ratios. Carbohydr. Polym. 2012, 89, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Hamidi, M.; Azadi, A.; Rafiei, P. Hydrogel nanoparticles in drug delivery. Adv. Drug Deliv. Rev. 2008, 60, 1638–1649. [Google Scholar] [CrossRef] [PubMed]
- Costa, E.M.; Silva, S.; Vicente, S.; Neto, C.; Castro, P.M.; Veiga, M.; Madureira, R.; Tavaria, F.; Pintado, M.M. Chitosan nanoparticles as alternative anti-staphylococci agents: Bactericidal, antibiofilm and antiadhesive effects. Mater. Sci. Eng. C 2017, 79, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Gomathi, T.; Sudha, P.N.; Florence, J.A.K.; Venkatesan, J.; Anil, S. Fabrication of letrozole formulation using chitosan nanoparticles through ionic gelation method. Int. J. Biol. Macromol. 2016, 104, 1820–1832. [Google Scholar] [CrossRef] [PubMed]
- Mesquita, P.C.; dos Santos-Silva, E.; Streck, L.; Damasceno, I.Z.; Maia, A.M.S.; Fernandes-Pedrosa, M.F.; da Silva-Júnior, A.A. Cationic functionalized biocompatible polylactide nanoparticles for slow release of proteins, Colloids Surfaces A Physicochem. Eng. Asp. 2017, 513, 442–451. [Google Scholar] [CrossRef]
- Silva, M.C.; Silva, T.L.; Silva, M.V.; Mota, C.M.; Santiago, F.M.; Fonseca, K.C.; Oliveira, F.; Mineo, T.W.P.; Mineo, J.R. Interaction between TNF and BmooMP-Alpha-I, a Zinc Metalloprotease Derived from Bothrops moojeni Snake Venom, Promotes Direct Proteolysis of This Cytokine: Molecular Modeling and Docking at a Glance. Toxins (Basel) 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Leong, K.W.; Mao, H.Q.; Truong-Le, V.L.; Roy, K.; Walsh, S.M.; August, J.T. DNA-polycation nanospheres as non-viral gene delivery vehicles. J. Control. Release 1998, 53, 183–193. [Google Scholar] [CrossRef]
- Son, S.; Kim, W.J. Biodegradable nanoparticles modified by branched polyethylenimine for plasmid DNA delivery. Biomaterials 2010, 31, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-W.; Chen, Y.-H.; Chen, J.-W.; Tsai, P.-J.; Huang, I.-H. Immunization with Recombinant TcdB-Encapsulated Nanocomplex Induces Protection against Clostridium difficile Challenge in a Mouse Model. Front. Microbiol. 2017, 8, 1411. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227. [Google Scholar] [CrossRef]
- Morrissey, J.H. Silver stains for proteins in polyacrylamide gels: A modified procedure with enhanced sensitivity. Anal. Biochem. 1981, 117, 307–310. [Google Scholar] [CrossRef]
- Bhattacharjee, S. DLS and zeta potential—What they are and what they are not? J. Control. Release 2016, 235, 337–351. [Google Scholar] [CrossRef] [PubMed]
- Nemen, D.; Lemos-Senna, E. Preparação e caracterização de suspensões coloidais de nanocarreadores lipídicos contendo resveratrol destinados à administração cutânea. Química Nova 2011, 34, 408–413. [Google Scholar] [CrossRef]
- Pedrosa, M.D.; de Azevedo, I.D.; Gonçalves-de-Andrade, R.M.; van den Berg, C.W.; Ramos, C.R.; Ho, P.L.; Tambourgi, D.V. Molecular cloning and expression of a functional dermonecrotic and haemolytic factor from Loxosceles laeta venom. Biochem. Biophys. Res. Commun. 2002, 298, 638–645. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Size (nm) | Zeta Potential (mV) | PdI | Encapsulation Efficiency (%) |
|---|---|---|---|---|
| CNPs | 159.6 ± 2.2 | 24.50 ± 3.64 | 0.272 ± 0.01 | - |
| CNPs/BJ 5 | 179.3 ± 9.4 ** | 23.03 ± 3.09 | 0.188 ± 0.08 | 76.7 |
| CNPs/BJ 10 | 174.7 ± 5.0 ** | 24.91 ± 2.91 | 0.203 ± 0.07 | 67.7 |
| CNPs/BJ 15 | 187.3 ± 7.5 *** | 31.40 ± 3.86 | 0.185 ± 0.07 | 74.0 |
| CNPs/BE 5 | 189.4 ± 1.0 *** | 20.71 ± 2.93 | 0.305 ± 0.05 | 97.2 |
| CNPs/BE 10 | 160.0 ± 2.3 | 19.00 ± 2.76 | 0.302 ± 0.01 | 87.6 |
| CNPs/BE 15 | 200.3 ± 5.6 *** | 27.21 ± 1.50 | 0.328 ± 0.05 | 88.1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soares, K.S.R.; Gláucia-Silva, F.; Daniele-Silva, A.; Torres-Rêgo, M.; Araújo, N.K.d.; Menezes, Y.A.S.d.; Damasceno, I.Z.; Tambourgi, D.V.; Da Silva-Júnior, A.A.; Fernandes-Pedrosa, M.D.F. Antivenom Production against Bothrops jararaca and Bothrops erythromelas Snake Venoms Using Cross-Linked Chitosan Nanoparticles as an Immunoadjuvant. Toxins 2018, 10, 158. https://doi.org/10.3390/toxins10040158
Soares KSR, Gláucia-Silva F, Daniele-Silva A, Torres-Rêgo M, Araújo NKd, Menezes YASd, Damasceno IZ, Tambourgi DV, Da Silva-Júnior AA, Fernandes-Pedrosa MDF. Antivenom Production against Bothrops jararaca and Bothrops erythromelas Snake Venoms Using Cross-Linked Chitosan Nanoparticles as an Immunoadjuvant. Toxins. 2018; 10(4):158. https://doi.org/10.3390/toxins10040158
Chicago/Turabian StyleSoares, Karla Samara Rocha, Fiamma Gláucia-Silva, Alessandra Daniele-Silva, Manoela Torres-Rêgo, Nathália Kelly de Araújo, Yamara Arruda Silva de Menezes, Igor Zumba Damasceno, Denise Vilarinho Tambourgi, Arnóbio Antônio Da Silva-Júnior, and Matheus De Freitas Fernandes-Pedrosa. 2018. "Antivenom Production against Bothrops jararaca and Bothrops erythromelas Snake Venoms Using Cross-Linked Chitosan Nanoparticles as an Immunoadjuvant" Toxins 10, no. 4: 158. https://doi.org/10.3390/toxins10040158
APA StyleSoares, K. S. R., Gláucia-Silva, F., Daniele-Silva, A., Torres-Rêgo, M., Araújo, N. K. d., Menezes, Y. A. S. d., Damasceno, I. Z., Tambourgi, D. V., Da Silva-Júnior, A. A., & Fernandes-Pedrosa, M. D. F. (2018). Antivenom Production against Bothrops jararaca and Bothrops erythromelas Snake Venoms Using Cross-Linked Chitosan Nanoparticles as an Immunoadjuvant. Toxins, 10(4), 158. https://doi.org/10.3390/toxins10040158