Involvement of Osteocytes in the Action of Pasteurella multocida Toxin


 ,
,  and
and 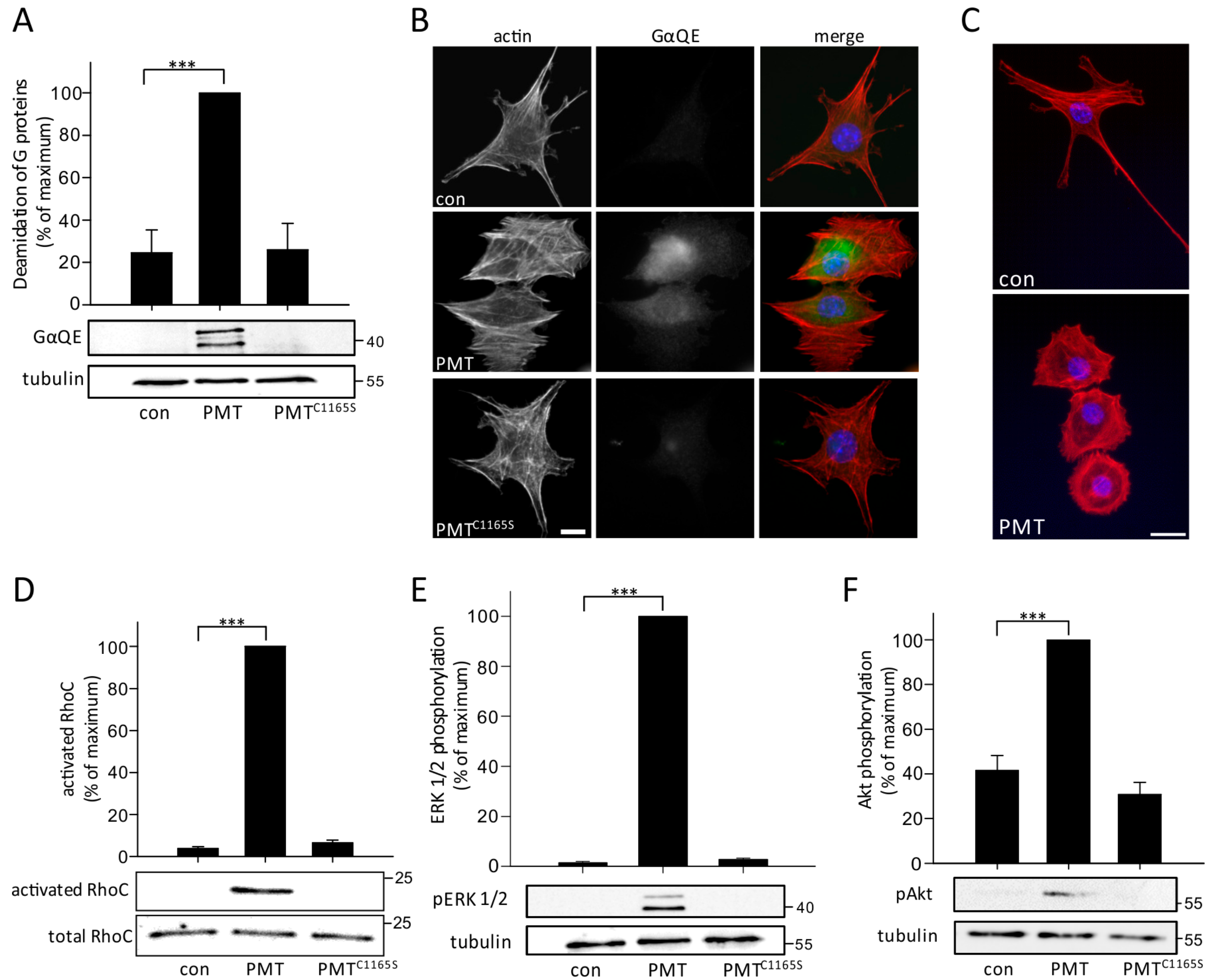{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. PMT Activates G Proteins and Downstream Signaling Pathways in MLO-Y4 Cells
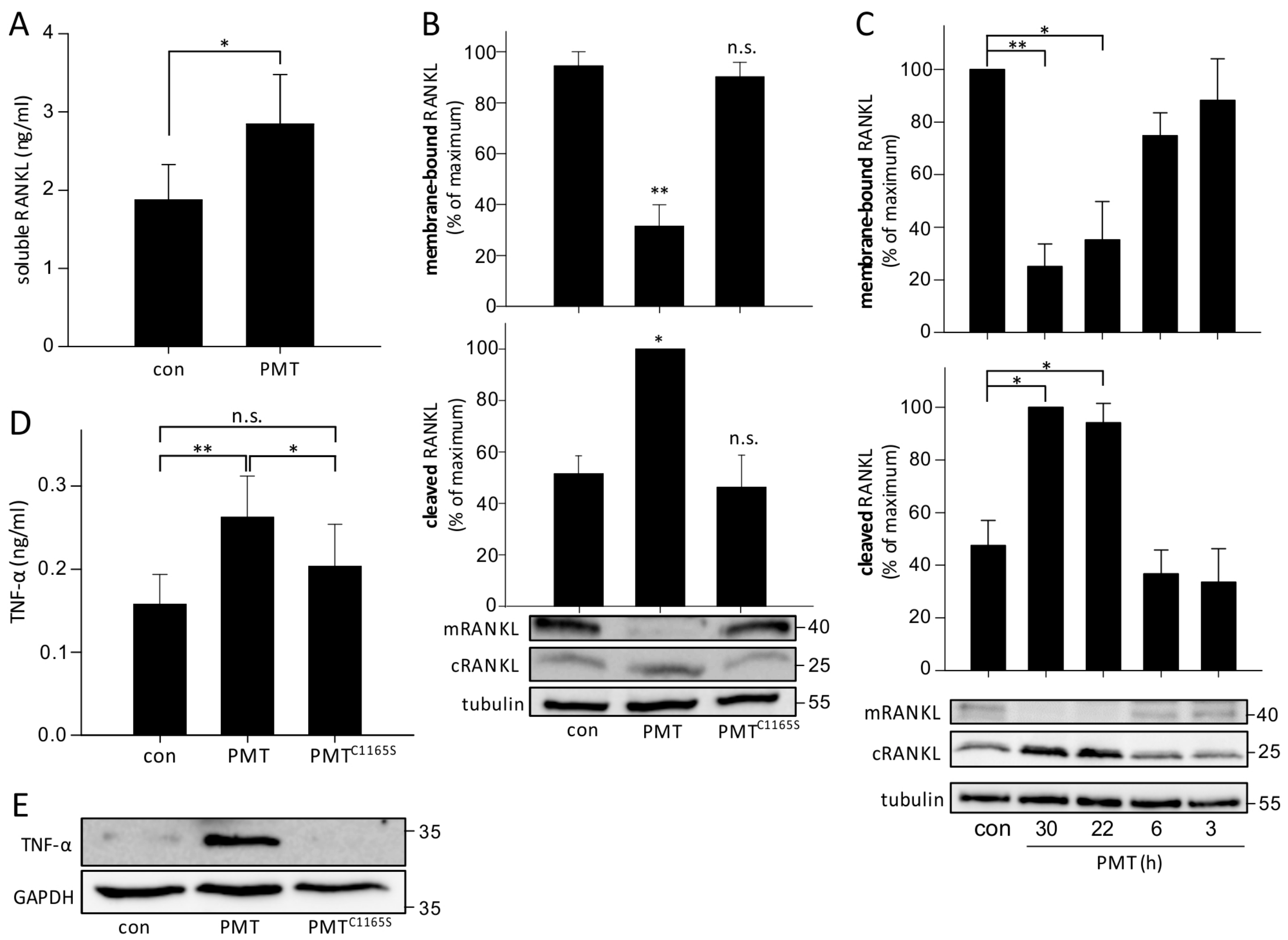
2.2. PMT Influences RANKL and TNF-α Expression
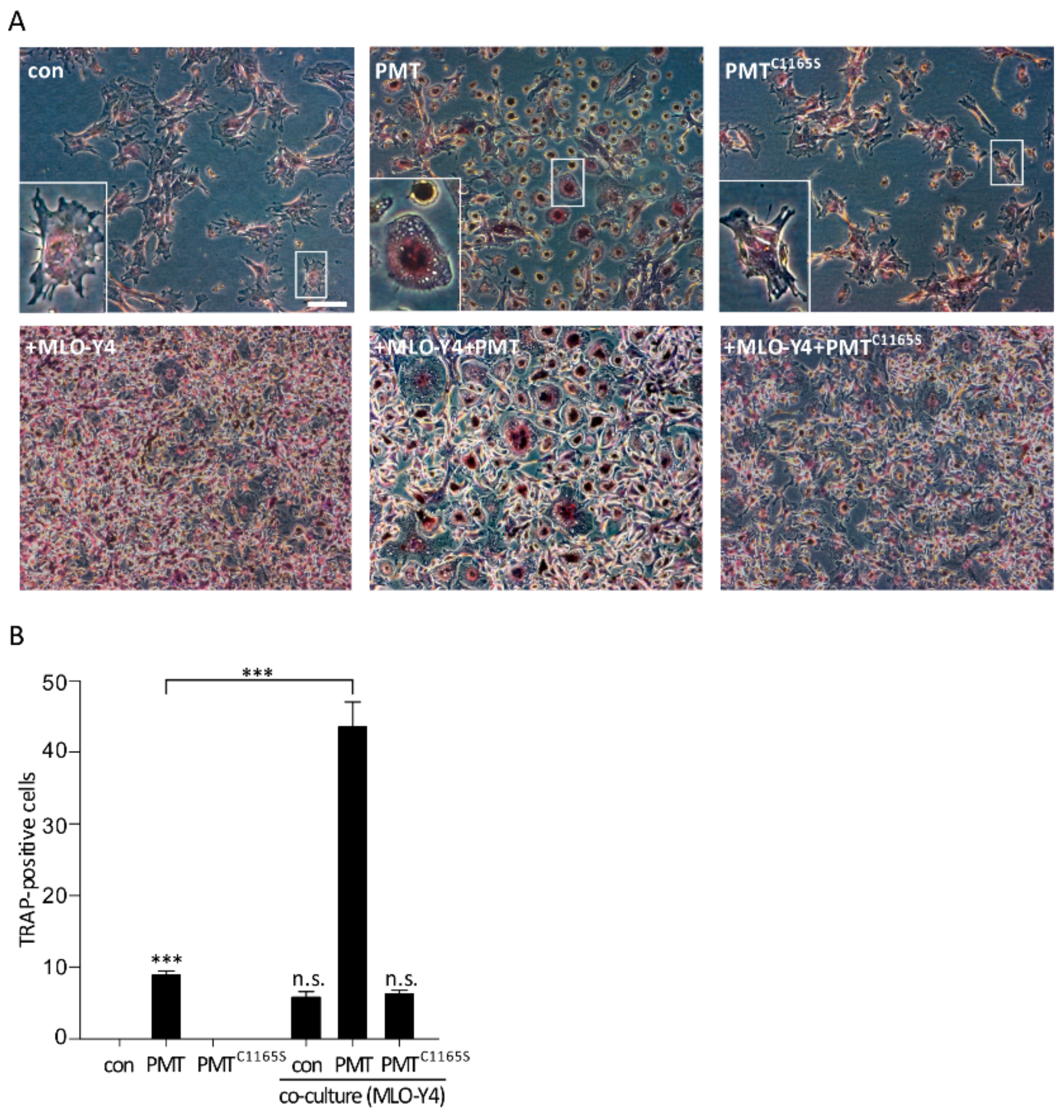
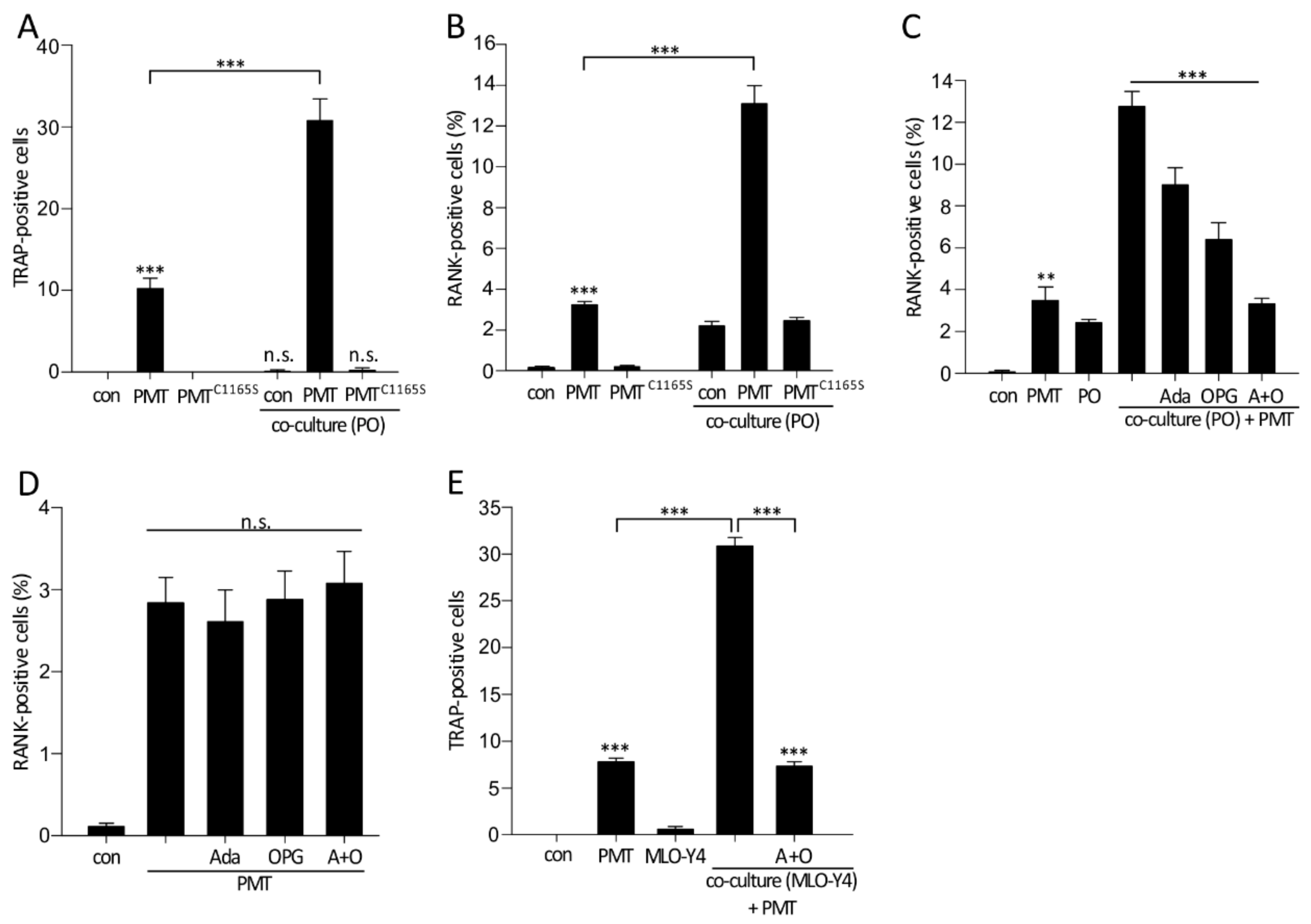
2.3. Increased Osteoclastogenesis in a Co-Culture Model of MLO-Y4 and Osteoclast Precursor Cells
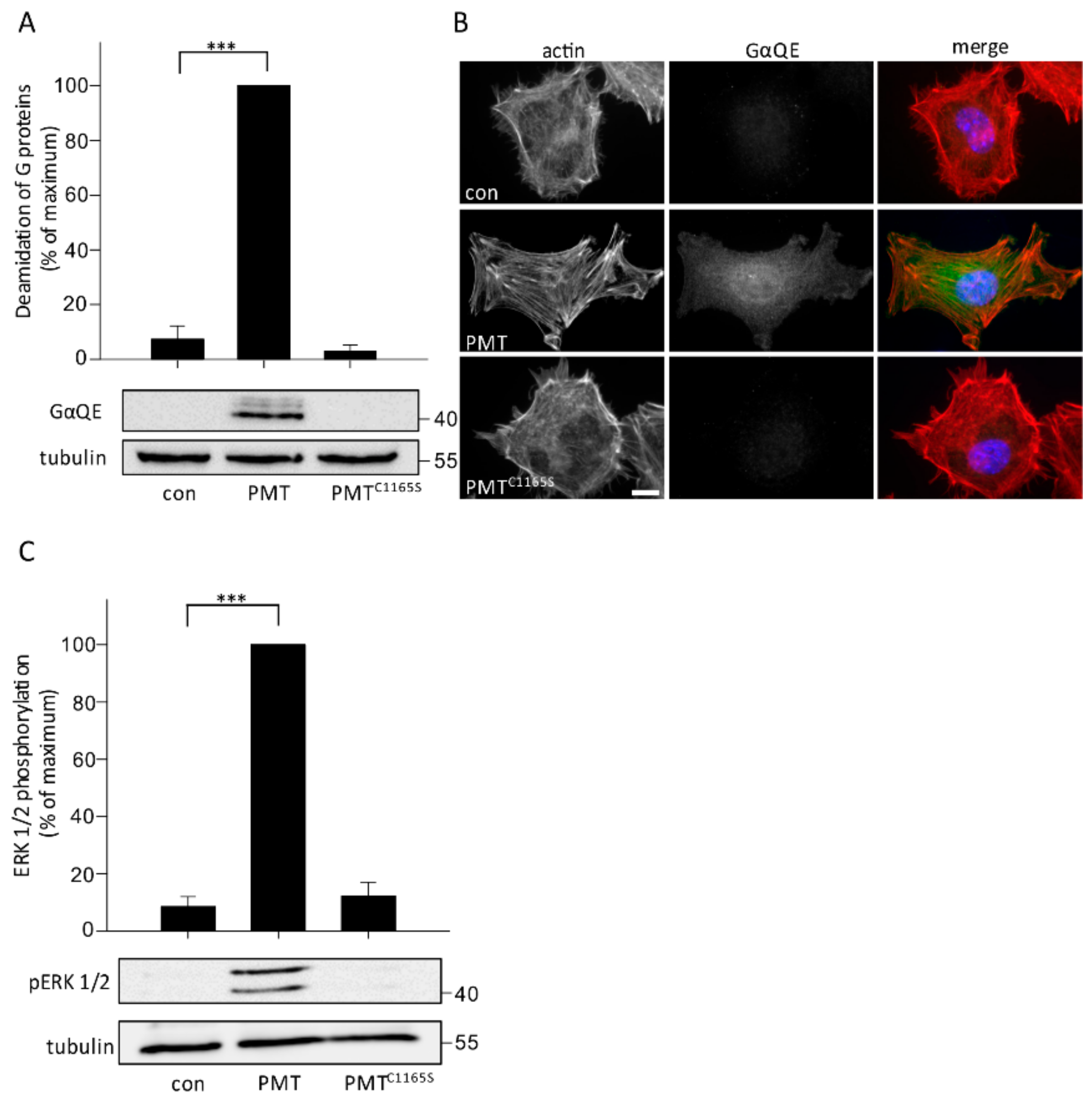
2.4. Primary Osteocytes Are Susceptible to PMT
2.5. Effects of PMT on the Differentiation of Osteoclasts in a Co-Culture Model
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Isolation of Primary Osteocytes and BMC
4.2. FACS Measurements
4.3. Immunofluorescence
4.4. Immunoblot Analysis
4.5. Rhotekin Pulldown Assay
4.6. ELISA
4.7. Cell Viability Assay
4.8. Measurement of The Caspase-3/7 Activity
4.9. TRAP Staining
4.10. Statistics
4.11. Ethics Statement
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Harper, M.; Boyce, J.D.; Adler, B. Pasteurella Multocida pathogenesis: 125 years after pasteur. FEMS Microbiol. Lett. 2006, 265, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Horiguchi, Y. Swine atrophic rhinitis caused by Pasteurella Multocida toxim and Bordetella dermonecrotic toxin. In Pasteurella multocida; Springer: Berlin/Heidelberg, Germany, 2012; pp. 113–129. [Google Scholar]
- Mullan, P.B.; Lax, A.J. Pasteurella Multocida toxin stimulates bone resorption by osteoclasts via interaction with osteoblasts. Calcif. Tissue Int. 1998, 63, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Sterner-Kock, A.; Lanske, B.; Uberschar, S.; Atkinson, M.J. Effects of the Pasteurella Multocida toxin on osteoblastic cells in vitro. Vet. Pathol. 1995, 32, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Orth, J.H.; Preuss, I.; Fester, I.; Schlosser, A.; Wilson, B.A.; Aktories, K. Pasteurella Multocida toxin activation of heterotrimeric g proteins by deamidation. Proc. Natl. Acad. Sci. USA 2009, 106, 7179–7184. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.A.; Ho, M. Recent insights into Pasteurella Multocida toxin and other g-protein-modulating bacterial toxins. Future Microbiol. 2010, 5, 1185–1201. [Google Scholar] [CrossRef] [PubMed]
- Orth, J.H.; Aktories, K. Molecular biology of Pasteurella Multocida toxin. In Pasteurella multocida; Springer: Berlin/Heidelberg, Germany, 2012; pp. 73–92. [Google Scholar]
- Wilson, B.A.; Ho, M. Pasteurella Multocida toxin interaction with host cells: Entry and cellular effects. In Pasteurella m ultocida; Springer: Berlin/Heidelberg, Germany, 2012; pp. 93–111. [Google Scholar]
- Strack, J.; Heni, H.; Gilsbach, R.; Hein, L.; Aktories, K.; Orth, J.H. Noncanonical g-protein-dependent modulation of osteoclast differentiation and bone resorption mediated by Pasteurella Multocida toxin. mBio. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Siegert, P.; Schmidt, G.; Papatheodorou, P.; Wieland, T.; Aktories, K.; Orth, J.H. Pasteurella Multocida toxin prevents osteoblast differentiation by transactivation of the map-kinase cascade via the galpha(q/11)- P63rhogef-Rhoa axis. PLoS. Pathog. 2013, 9, e1003385. [Google Scholar] [CrossRef] [PubMed]
- Massey, H.M.; Flanagan, A.M. Human osteoclasts derive from CD14-positive monocytes. Br. J. Haematol. 1999, 106, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, T.; Hayashi, M.; Fukunaga, T.; Kurata, K.; Oh-Hora, M.; Feng, J.Q.; Bonewald, L.F.; Kodama, T.; Wutz, A.; Wagner, E.F.; et al. Evidence for osteocyte regulation of bone homeostasis through rankl expression. Nat. Med. 2011, 17, 1231–1234. [Google Scholar] [CrossRef] [PubMed]
- Sila-Asna, M.; Bunyaratvej, A.; Maeda, S.; Kitaguchi, H.; Bunyaratavej, N. Osteoblast differentiation and bone formation gene expression in strontium-inducing bone marrow mesenchymal stem cell. Kobe. J. Med. Sci. 2007, 53, 25–35. [Google Scholar] [PubMed]
- Atkins, G.J.; Findlay, D.M. Osteocyte regulation of bone mineral: A little give and take. Osteoporos. Int. 2012, 23, 2067–2079. [Google Scholar] [CrossRef] [PubMed]
- Schaffler, M.B.; Cheung, W.Y.; Majeska, R.; Kennedy, O. Osteocytes: Master orchestrators of bone. Calcif. Tissue Int. 2014, 94, 5–24. [Google Scholar] [CrossRef] [PubMed]
- Prideaux, M.; Findlay, D.M.; Atkins, G.J. Osteocytes: The master cells in bone remodelling. Curr. Opin. Pharmacol. 2016, 28, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Bonewald, L.F.; Kneissel, M.; Johnson, M. Preface: The osteocyte. Bone 2013, 54, 181. [Google Scholar] [CrossRef] [PubMed]
- Al-Dujaili, S.A.; Lau, E.; Al-Dujaili, H.; Tsang, K.; Guenther, A.; You, L. Apoptotic osteocytes regulate osteoclast precursor recruitment and differentiation in vitro. J. Cell Biochem. 2011, 112, 2412–2423. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Senda, T.; Kubo, K.Y. The osteocyte plays multiple roles in bone remodeling and mineral homeostasis. Med. Mol. Morphol. 2015, 48, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Dallas, S.L.; Prideaux, M.; Bonewald, L.F. The osteocyte: An endocrine cell … and more. Endocr. Rev. 2013, 34, 658–690. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, T.; Hayashi, M.; Takayanagi, H. New insights into osteoclastogenic signaling mechanisms. Trends Endocrinol. Metab. 2012, 23, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Onal, M.; Jilka, R.L.; Weinstein, R.S.; Manolagas, S.C.; O’Brien, C.A. Matrix-embedded cells control osteoclast formation. Nat. Med. 2011, 17, 1235–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, J.; Piemontese, M.; Onal, M.; Campbell, J.; Goellner, J.J.; Dusevich, V.; Bonewald, L.; Manolagas, S.C.; O’Brien, C.A. Osteocytes, not osteoblasts or lining cells, are the main source of the rankl required for osteoclast formation in remodeling bone. PLoS ONE 2015, 10, e0138189. [Google Scholar] [CrossRef] [PubMed]
- You, L.; Temiyasathit, S.; Lee, P.; Kim, C.H.; Tummala, P.; Yao, W.; Kingery, W.; Malone, A.M.; Kwon, R.Y.; Jacobs, C.R. Osteocytes as mechanosensors in the inhibition of bone resorption due to mechanical loading. Bone 2008, 42, 172–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, Y.; Windle, J.J.; Koop, B.A.; Mundy, G.R.; Bonewald, L.F. Establishment of an osteocyte-like cell line, mlo-y4. J. Bone Miner. Res. 1997, 12, 2014–2023. [Google Scholar] [CrossRef] [PubMed]
- Bonewald, L.F. Establishment and characterization of an osteocyte-like cell line, mlo-y4. J. Bone Miner. Metab. 1999, 17, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Kamitani, S.; Ao, S.; Toshima, H.; Tachibana, T.; Hashimoto, M.; Kitadokoro, K.; Fukui-Miyazaki, A.; Abe, H.; Horiguchi, Y. Enzymatic actions of Pasteurella multocida toxin detected by monoclonal antibodies recognizing the deamidated alpha subunit of the heterotrimeric gtpase g(q). FEBS J. 2011, 278, 2702–2712. [Google Scholar] [CrossRef] [PubMed]
- Orth, J.H.; Aktories, K. Pasteurella multocida toxin activates various heterotrimeric g proteins by deamidation. Toxins 2010, 2, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Orth, J.H.; Fester, I.; Siegert, P.; Weise, M.; Lanner, U.; Kamitani, S.; Tachibana, T.; Wilson, B.A.; Schlosser, A.; Horiguchi, Y.; et al. Substrate specificity of Pasteurella multocida toxin for alpha subunits of heterotrimeric g proteins. FASEB J. 2013, 27, 832–842. [Google Scholar] [CrossRef] [PubMed]
- Knowles, H.J.; Athanasou, N.A. Canonical and non-canonical pathways of osteoclast formation. Histol. Histopathol. 2009, 24, 337–346. [Google Scholar] [PubMed]
- O’Brien, C.A.; Nakashima, T.; Takayanagi, H. Osteocyte control of osteoclastogenesis. Bone 2013, 54, 258–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kogianni, G.; Mann, V.; Noble, B.S. Apoptotic bodies convey activity capable of initiating osteoclastogenesis and localized bone destruction. J. Bone Miner. Res. 2008, 23, 915–927. [Google Scholar] [CrossRef] [PubMed]
- Azuma, Y.; Kaji, K.; Katogi, R.; Takeshita, S.; Kudo, A. Tumor necrosis factor-alpha induces differentiation of and bone resorption by osteoclasts. J. Biol. Chem. 2000, 275, 4858–4864. [Google Scholar] [CrossRef] [PubMed]
- Komine, M.; Kukita, A.; Kukita, T.; Ogata, Y.; Hotokebuchi, T.; Kohashi, O. Tumor necrosis factor-alpha cooperates with receptor activator of nuclear factor kappab ligand in generation of osteoclasts in stromal cell-depleted rat bone marrow cell culture. Bone 2001, 28, 474–483. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Heulsmann, A.; Tondravi, M.M.; Mukherjee, A.; Abu-Amer, Y. Tumor necrosis factor-alpha (tnf) stimulates rankl-induced osteoclastogenesis via coupling of tnf type 1 receptor and rank signaling pathways. J. Biol. Chem. 2001, 276, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Orth, J.H.; Lang, S.; Taniguchi, M.; Aktories, K. Pasteurella multocida toxin-induced activation of rhoa is mediated via two families of gα proteins, gαq and gα12/13. J. Biol. Chem. 2005, 280, 36701–36707. [Google Scholar] [CrossRef] [PubMed]
- Sabokbar, A.; Mahoney, D.J.; Hemingway, F.; Athanasou, N.A. Non-canonical (rankl-independent) pathways of osteoclast differentiation and their role in musculoskeletal diseases. Clin. Rev. Allergy Immunol. 2016, 51, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Crockett, J.C.; Rogers, M.J.; Coxon, F.P.; Hocking, L.J.; Helfrich, M.H. Bone remodelling at a glance. J. Cell Sci. 2011, 124, 991–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takayanagi, H. Osteoimmunology: Shared mechanisms and crosstalk between the immune and bone systems. Nat. Rev. Immunol. 2007, 7, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Kleyer, A.; Schett, G. Arthritis and bone loss: A hen and egg story. Curr. Opin. Rheumatol. 2014, 26, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Ritchlin, C.T.; Haas-Smith, S.A.; Li, P.; Hicks, D.G.; Schwarz, E.M. Mechanisms of tnf-alpha- and rankl-mediated osteoclastogenesis and bone resorption in psoriatic arthritis. J. Clin. Investig. 2003, 111, 821–831. [Google Scholar] [CrossRef]
- Chakraborty, S.; Kloos, B.; Harre, U.; Schett, G.; Kubatzky, K.F. Pasteurella multocida toxin triggers rankl-independent osteoclastogenesis. Front. Immunol. 2017, 8, 185. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, D.; Heeg, K.; Kubatzky, K.F. Pasteurella multocida toxin-stimulated osteoclast differentiation is b cell dependent. Infect. Immun. 2011, 79, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Stern, A.R.; Stern, M.M.; Van Dyke, M.E.; Jahn, K.; Prideaux, M.; Bonewald, L.F. Isolation and culture of primary osteocytes from the long bones of skeletally mature and aged mice. Biotechnology 2012, 52, 361–373. [Google Scholar] [CrossRef]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heni, H.; Ebner, J.K.; Schmidt, G.; Aktories, K.; Orth, J.H.C. Involvement of Osteocytes in the Action of Pasteurella multocida Toxin. Toxins 2018, 10, 328. https://doi.org/10.3390/toxins10080328
Heni H, Ebner JK, Schmidt G, Aktories K, Orth JHC. Involvement of Osteocytes in the Action of Pasteurella multocida Toxin. Toxins. 2018; 10(8):328. https://doi.org/10.3390/toxins10080328
Chicago/Turabian StyleHeni, Hannah, Julia K. Ebner, Gudula Schmidt, Klaus Aktories, and Joachim H. C. Orth. 2018. "Involvement of Osteocytes in the Action of Pasteurella multocida Toxin" Toxins 10, no. 8: 328. https://doi.org/10.3390/toxins10080328
APA StyleHeni, H., Ebner, J. K., Schmidt, G., Aktories, K., & Orth, J. H. C. (2018). Involvement of Osteocytes in the Action of Pasteurella multocida Toxin. Toxins, 10(8), 328. https://doi.org/10.3390/toxins10080328