Insights into the Voltage Regulation Mechanism of the Pore-Forming Toxin Lysenin


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussions
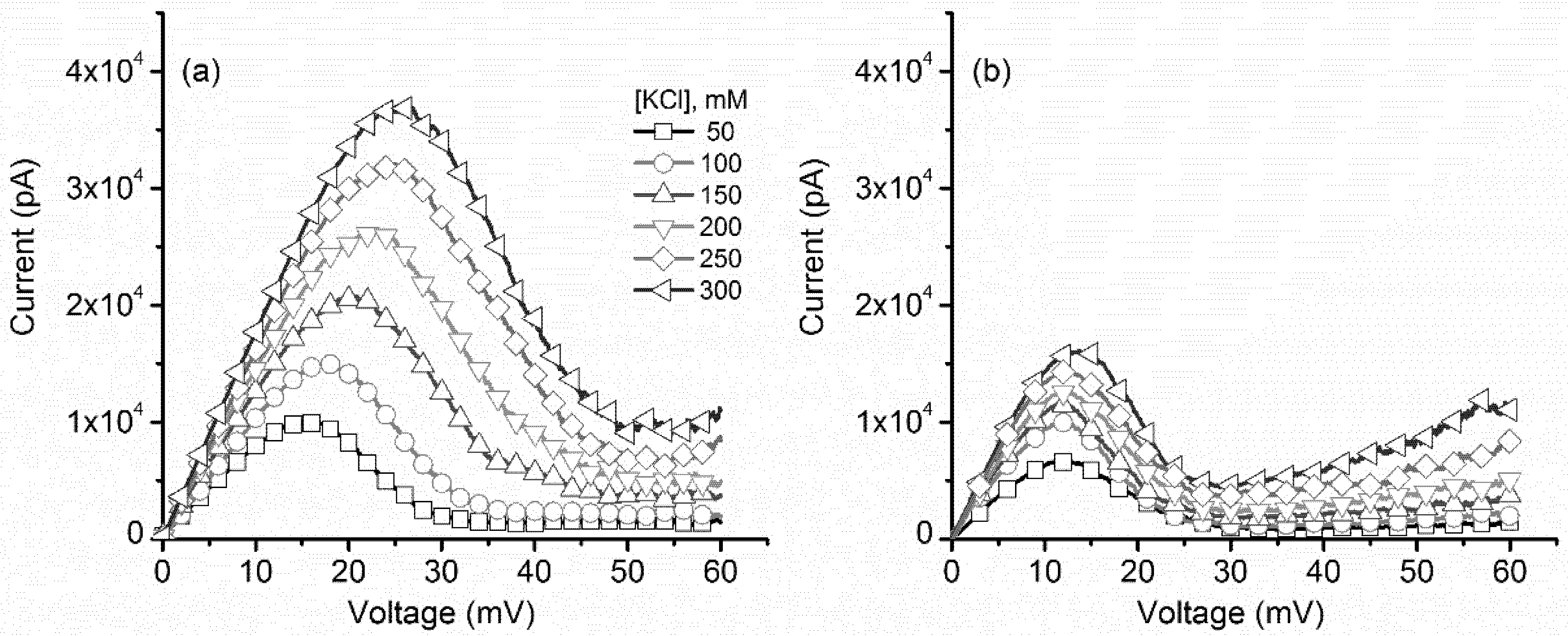
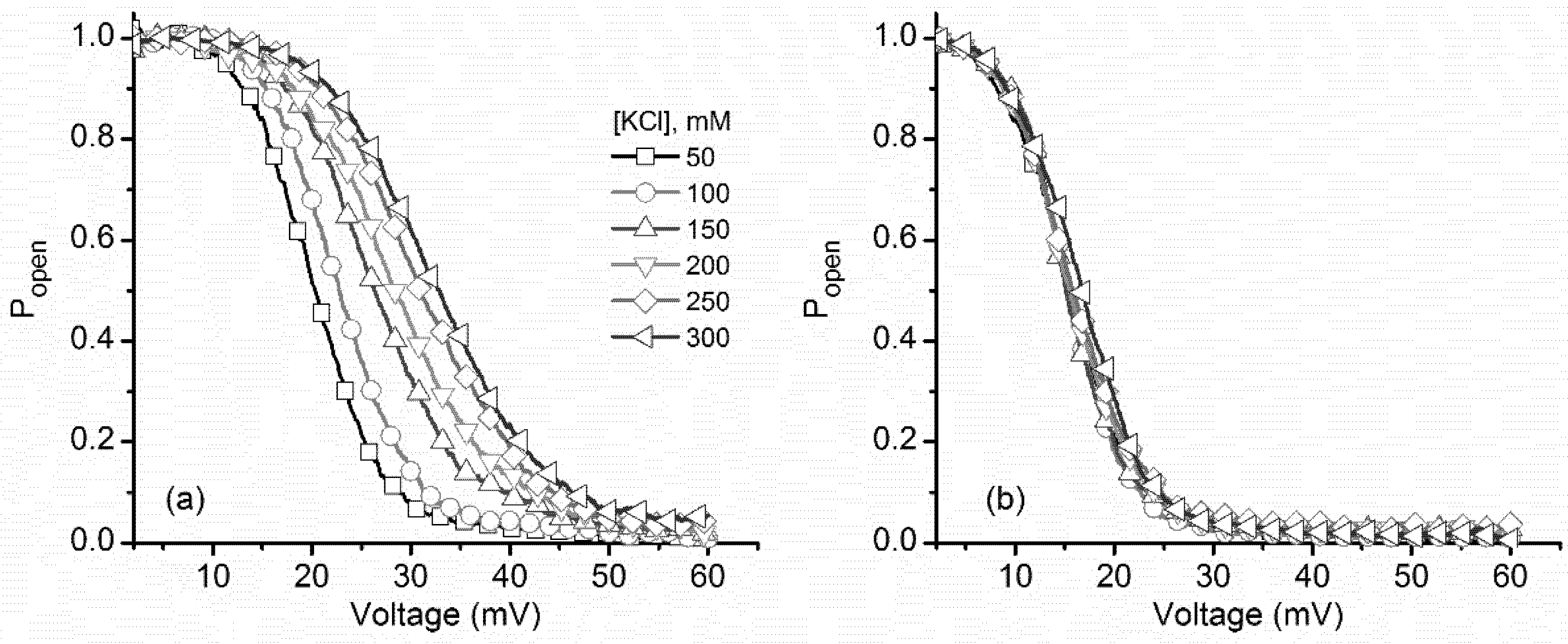
2.1. Monovalent Metal Cations Modulate the Voltage-Induced Gating of Lysenin Channels During Inactivation, while Minimally Influencing the Reactivation Pathway
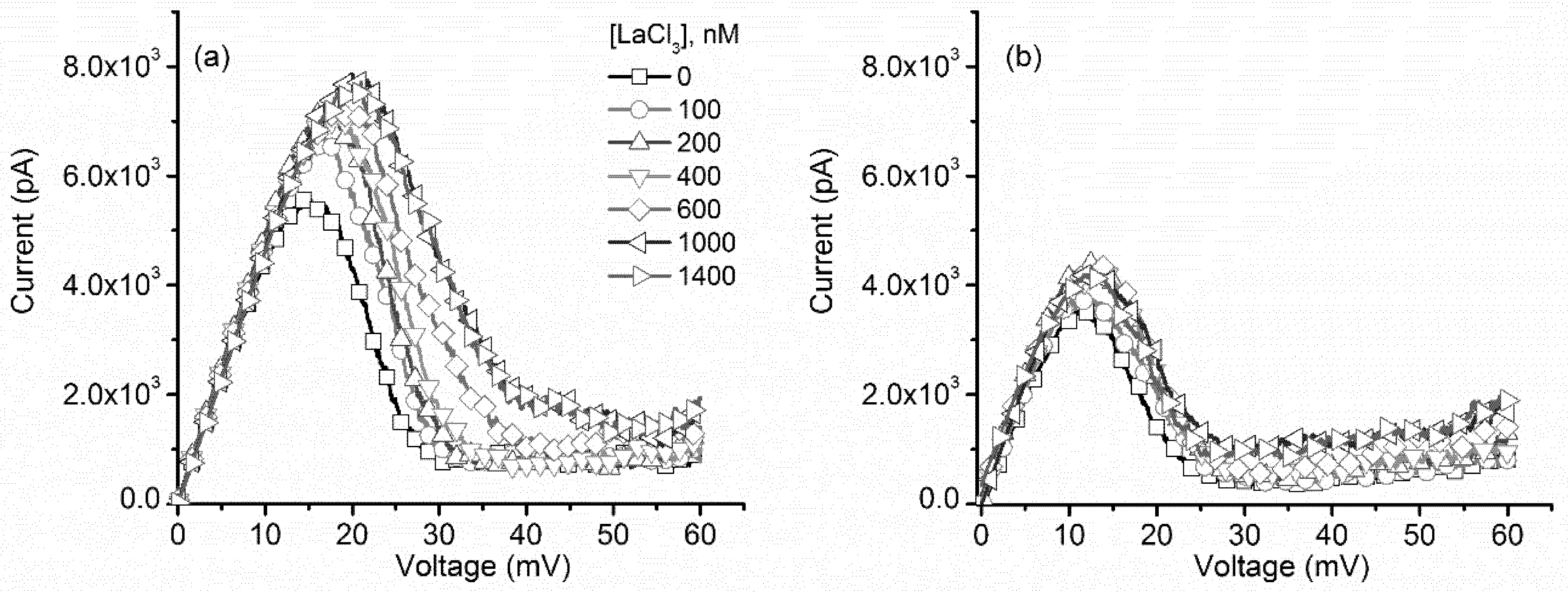
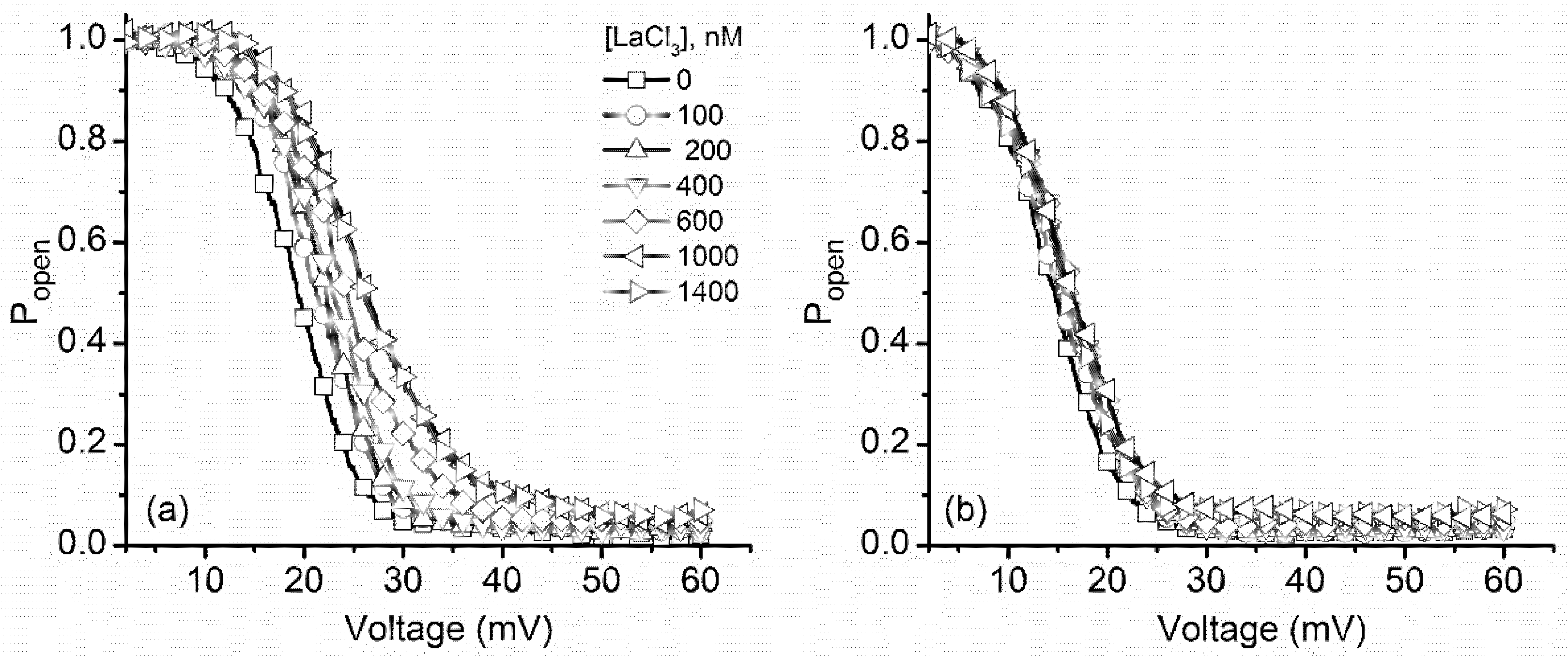
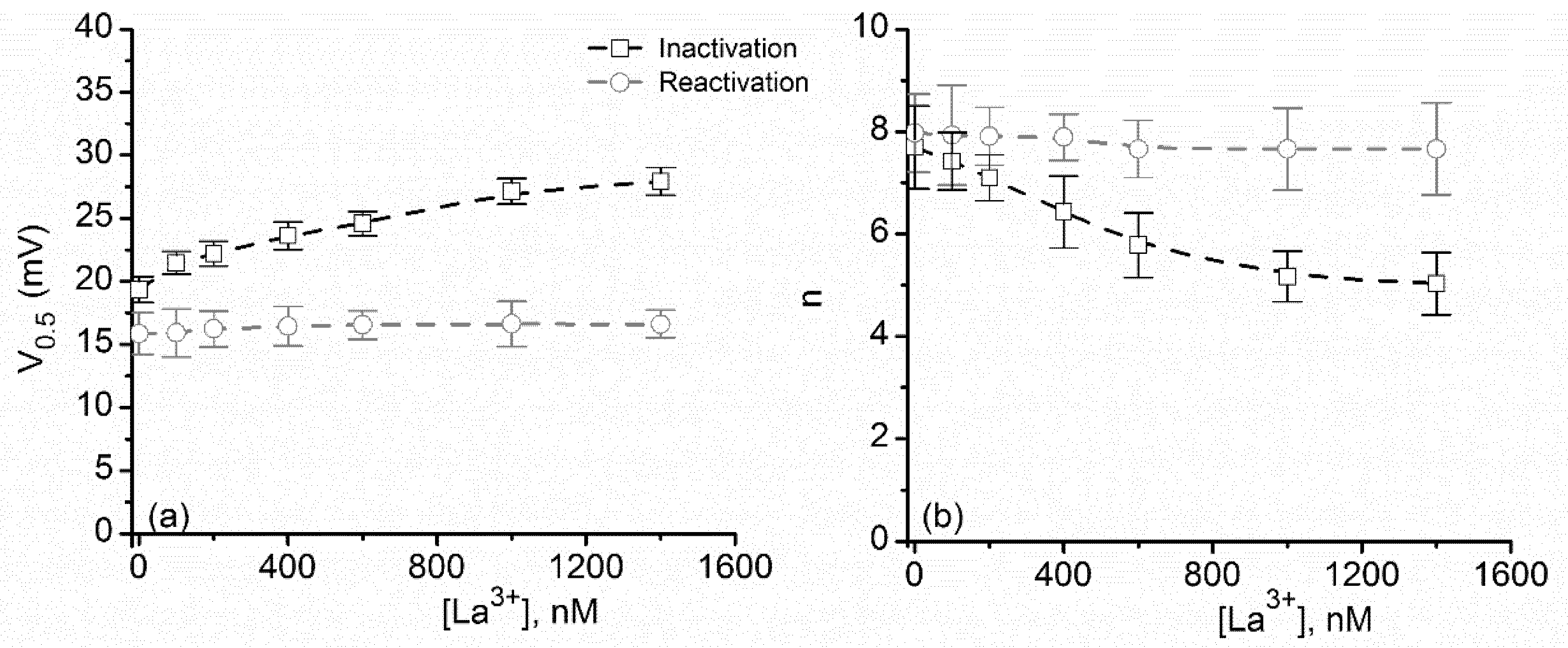
2.2. Multivalent Metal Cations Influence the Voltage Regulation of Lysenin Channels Similarly to Monovalent Ions
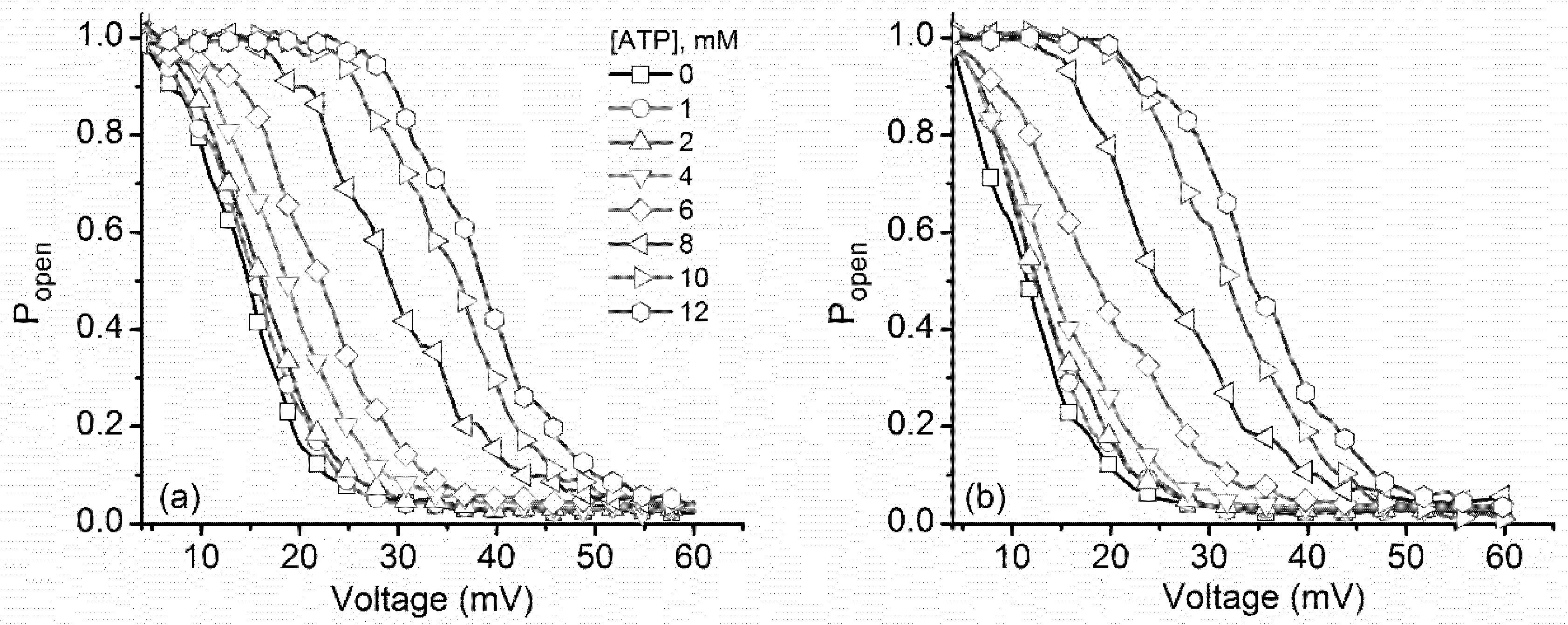
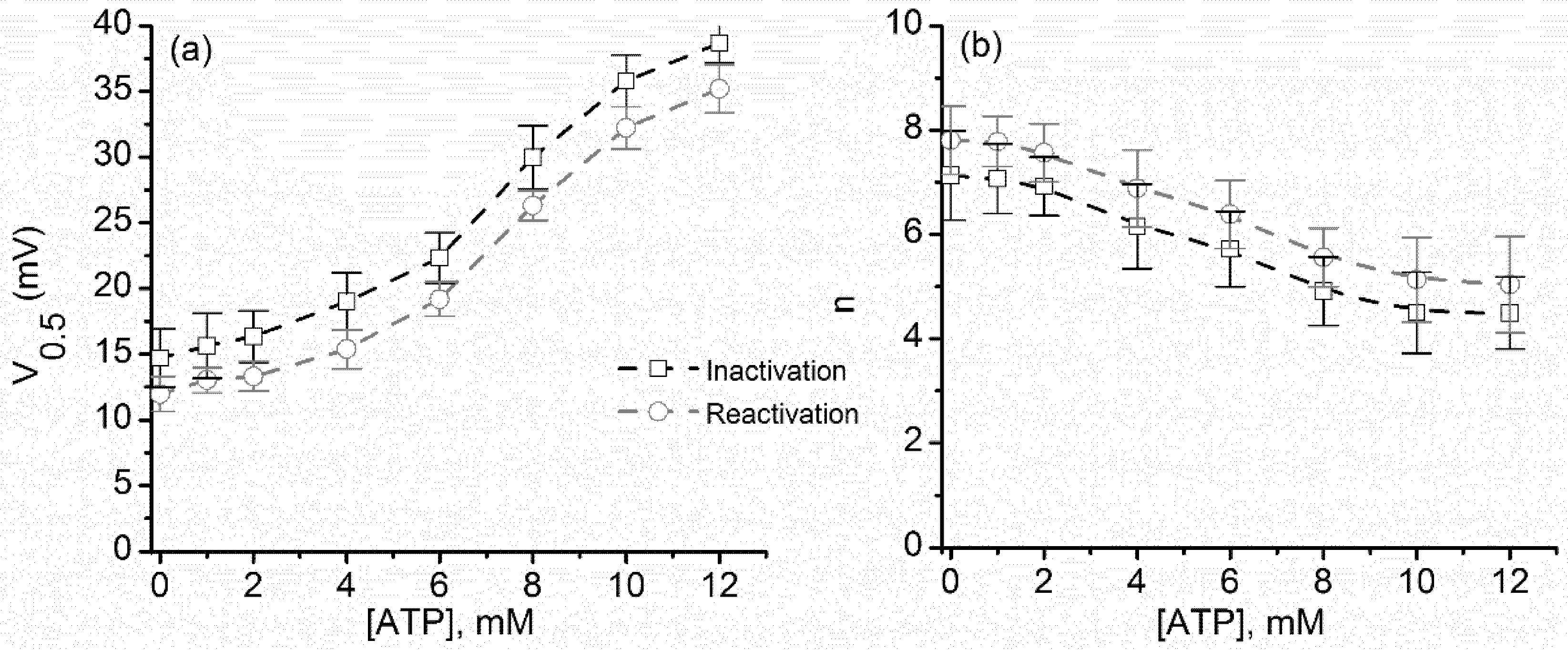
2.3. ATP Binding to Lysenin Channels Modulates the Voltage-Induced Gating and Affects Both the Inactivation and Reactivation Pathways
3. Materials and Methods
3.1. Bilayer Lipid Formation, Channel Insertion, and Ionic Addition
3.2. Data Collection, Analysis, and Mathematical Modelling
Author Contributions
Funding
Conflicts of Interest
References
- Bruhn, H.; Winkelmann, J.; Andersen, C.; Andra, J.; Leippe, M. Dissection of the mechanisms of cytolytic and antibacterial activity of lysenin, a defense protein of the annelid Eisenia fetida. Dev. Comp. Immunol. 2006, 30, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowska, K.; Hordejuk, R.; Szymczyk, P.; Kulma, M.; Abdel-Shakor, A.-B.; Plucienniczak, A.; Dolowy, K.; Szewczyk, A.; Sobota, A. Lysenin-His, a sphingomyelin-recognizing toxin, requires tryptophan 20 for cation-selective channel assembly but not for membrane binding. Mol. Membr. Biol. 2007, 24, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Shakor, A.-B.A.; Czurylo, E.A.; Sobota, A. Lysenin, a unique sphingomyelin-binding protein. FEBS Lett. 2003, 542, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Shogomori, H.; Kobayashi, T. Lysenin: A sphingomyelin specific pore-forming toxin. Biochim. Biophys. Acta 2008, 1780, 612–618. [Google Scholar] [CrossRef] [PubMed]
- Yamaji-Hasegawa, A.; Makino, A.; Baba, T.; Senoh, Y.; Kimura-Suda, H.; Sato, S.B.; Terada, N.; Ohno, S.; Kiyokawa, E.; Umeda, M.; et al. Oligomerization and pore formation of a sphingomyelin-specific toxin, lysenin. J. Biol. Chem. 2003, 278, 22762–22770. [Google Scholar] [CrossRef] [PubMed]
- Fologea, D.; Krueger, E.; Lee, R.; Naglak, M.; Mazur, Y.; Henry, R.; Salamo, G. Controlled gating of lysenin pores. Biophys. Chem. 2010, 146, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Ide, T.; Aoki, T.; Takeuchi, Y.; Yanagida, T. Lysenin forms a voltage-dependent channel in artificial lipid bilayer membranes. Biochem. Biophys. Res. Commun. 2006, 346, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, N.; Kobayashi, T. Visualization of lipid membrane reorganization induced by a pore-forming toxin using high-speed atomic force microscopy. ACS Nano 2015, 9, 7960–7967. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, N.; Yamada, T.; Greimel, P.; Uchihashi, T.; Ando, T.; Kobayashi, T. Real-time visualization of assembling of a sphingomyelin-specific toxin on planar lipid membranes. Biophys. J. 2013, 105, 1397–1405. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, N.; Yamaji-Hasegawa, A.; Hullin-Matsuda, F.; Kobayashi, T. Molecular mechanisms of action of sphingomyelin-specific pore-forming toxin, lysenin. Semin. Cell Dev. Biol. 2018, 73, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Fologea, D.; Al Faori, R.; Krueger, E.; Mazur, Y.I.; Kern, M.; Williams, M.; Mortazavi, A.; Henry, R.; Salamo, G.J. Potential analytical applications of lysenin channels for detection of multivalent ions. Anal. Bioanal. Chem. 2011, 401, 1871–1879. [Google Scholar] [CrossRef] [PubMed]
- Fologea, D.; Krueger, E.; Al Faori, R.; Lee, R.; Mazur, Y.I.; Henry, R.; Arnold, M.; Salamo, G.J. Multivalent ions control the transport through lysenin channels. Biophys. Chem. 2010, 152, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Bryant, S.; Shrestha, N.; Carnig, P.; Kosydar, S.; Belzeski, P.; Hanna, C.; Fologea, D. Purinergic control of lysenin’s transport and voltage-gating properties. Purinerg. Signal. 2016, 12, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Krueger, E.; Al Faouri, R.; Fologea, D.; Henry, R.; Straub, D.; Salamo, G. A model for the hysteresis observed in gating of lysenin channels. Biophys. Chem. 2013, 184, 126–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krueger, E.; Bryant, S.; Shrestha, N.; Clark, T.; Hanna, C.; Pink, D.; Fologea, D. Intramembrane congestion effects on lysenin channel voltage-induced gating. Eur. Biophys. J. 2016, 45, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Bainbridge, G.; Gokce, I.; Lakey, J.H. Voltage gating is a fundamental feature of porin and toxin β-barrel membrane channels. FEBS Lett. 1998, 431, 305–308. [Google Scholar] [CrossRef] [Green Version]
- Fologea, D.; Krueger, E.; Mazur, Y.I.; Stith, C.; Okuyama, Y.; Henry, R.; Salamo, G.J. Bi-stability, hysteresis, and memory of voltage-gated lysenin channels. Biochim. Biophys. Acta-Biomembr. 2011, 1808, 2933–2939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pustovoit, M.A.; Berezhkovskii, A.M.; Bezrukov, S.M. Analytical theory of hysteresis in ion channels: Two state model. J. Chem. Phys. 2006, 125, 194907. [Google Scholar] [CrossRef] [PubMed]
- Bokori-Brown, M.; Martin, T.G.; Naylor, C.E.; Basak, A.K.; Titball, R.W.; Savva, C.G. Cryo-EM structure of lysenin pore elucidates membrane insertion by an aerolysin family protein. Nat. Commun. 2016, 7, 11293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Colibus, L.; Sonnen, A.F.; Morris, K.J.; Siebert, C.A.; Abrusci, P.; Plitzko, J.; Hodnik, V.; Leippe, M.; Volpi, E.; Anderluh, G.; et al. Structures of lysenin reveal a shared evolutionary origin for pore-forming proteins and its mode of sphingomyelin recognition. Structure 2012, 20, 1498–1507. [Google Scholar] [CrossRef] [PubMed]
- Podobnik, M.; Savory, P.; Rojko, N.; Kisovec, M.; Wood, N.; Hambley, R.; Pugh, J.; Wallace, E.J.; McNeill, L.; Bruce, M.; et al. Crystal structure of an invertebrate cytolysin pore reveals unique properties and mechanism of assembly. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Andersen, O.S.; Ingolfsson, H.I.; Lundbaek, J.A. Ion channels. Wiley Encycl. Chem. Biol. 2008, 2, 419–436. [Google Scholar] [CrossRef]
- Bezanilla, F. The voltage sensor in voltage-dependent ion channels. Physiol. Rev. 2000, 80, 555–592. [Google Scholar] [CrossRef] [PubMed]
- Bezanilla, F. Voltage-gated ion channels. IEEE T. Nanobiosci. 2005, 4, 34–48. [Google Scholar] [CrossRef]
- Bezanilla, F. How membrane proteins sense voltage. Nat. Rev. Mol. Cell Biol. 2008, 9, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Bezanilla, F. Ion channels: From conductance to structure. Neuron 2008, 60, 456–468. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, C.; Morera, F.J.; Rosenmann, E.; Alvarez, O.; Latorre, R. S3b amino acid residues do not shuttle across the bilayer in voltage-dependent shaker K+ channels. Proc. Nat. Acad. Sci. USA 2005, 102, 5020–5025. [Google Scholar] [CrossRef] [PubMed]
- Swartz, K.J. Sensing voltage across lipid membranes. Nature 2008, 456, 891–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryant, S.L.; Eixenberger, J.E.; Rossland, S.; Apsley, H.; Hoffmann, C.; Shrestha, N.; McHugh, M.; Punnoose, A.; Fologea, D. ZnO nanoparticles modulate the ionic transport and voltage regulation of lysenin nanochannels. J. Nanobiotechnol. 2017, 15, 90. [Google Scholar] [CrossRef] [PubMed]
- Islas, L.D.; Sigworth, F.J. Electrostatics and the gating pore of Shaker potassium channels. J. Gen. Physiol. 2001, 117, 69–89. [Google Scholar] [CrossRef] [PubMed]
- Rappaport, S.M.; Teijido, O.; Hoogerheide, D.P.; Rostovtseva, T.K.; Berezhkovskii, A.M.; Bezrukov, S.M. Conductance hysteresis in the voltage-dependent anion channel. Eur. Biophys. J. 2015, 44, 465–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.P.; Shin, H.G.; Szep, S.; Lu, Z. Physical determinants of strong voltage sensitivity of K+ channel block. Nat. Struct. Mol. Biol. 2009, 16, 1252–1258. [Google Scholar] [CrossRef] [PubMed]
- Correa, A.M.; Bezanilla, F.; Latorre, R. Gating kinetics of batrachotoxin-modified Na+ channels in the squid giant axon-voltage and temperature effects. Biophys. J. 1992, 61, 1332–1352. [Google Scholar] [CrossRef]
- Block, B.M.; Stacey, W.C.; Jones, S.W. Surface charge and lanthanum block of calcium current in bullfrog sympathetic neurons. Biophys. J. 1998, 74, 2278–2284. [Google Scholar] [CrossRef]
- Bostrom, M.; Ninham, B.W. Energy of an ion crossing a low dielectric membrane: The role of dispersion self-free energy. Biophys. Chem. 2005, 114, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Cherstvy, A.G. Electrostatic screening and energy barriers of ions in low-dielectric membranes. J. Phys. Chem. B 2006, 110, 14503–14506. [Google Scholar] [CrossRef] [PubMed]
- Glaeser, R.M.; Jap, B.K. The Born energy problem in bacteriorhodopsin. Biophys. J. 1984, 45, 95–97. [Google Scholar] [CrossRef]
- Parsegian, A. Energy of an ion crossing a low dielectric membrane: Solutions to four relevant electrostatic problems. Nature 1969, 221, 844–846. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.X.; Lee, A.; Chen, J.Y.; Ruta, V.; Cadene, M.; Chait, B.T.; MacKinnon, R. X-ray structure of a voltage-dependent K+ channel. Nature 2003, 423, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.X.; Ruta, V.; Chen, J.Y.; Lee, A.; MacKinnon, R. The principle of gating charge movement in a voltage-dependent K+ channel. Nature 2003, 423, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.G. Ion channels: A paddle in oil. Nature 2006, 444, 697. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, D.; Jiang, Q.X.; MacKinnon, R. Phospholipids and the origin of cationic gating charges in voltage sensors. Nature 2006, 444, 775–779. [Google Scholar] [CrossRef] [PubMed]
- Sigworth, F.J. Structural biology: Life’s transistors. Nature 2003, 423, 21–22. [Google Scholar] [CrossRef] [PubMed]
- Debye, V.P.; Huckel, E. Zur theorie der elektrolyte. Phys. Chem. 1923, 24, 185–206. [Google Scholar]
- Solomon, T. The definition and unit of ionic strength. J. Chem. Educ. 2001, 78, 1691–1692. [Google Scholar] [CrossRef]
- Latorre, R.; Vargas, G.; Orta, G.; Brauchi, S. Voltage and temperature gating of thermoTRP channels. In TRP Ion Channel Function in Sensory Transduction and Cellular Signaling Cascades; Liedte, W.B., Heller, S., Eds.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2007; Chapter 21; pp. 287–302. [Google Scholar]
- Tao, X.; Lee, A.; Limapichat, W.; Dougherty, D.A.; MacKinnon, R. A gating charge transfer center in voltage sensors. Science 2010, 328, 67–73. [Google Scholar] [CrossRef] [PubMed]









© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bryant, S.L.; Clark, T.; Thomas, C.A.; Ware, K.S.; Bogard, A.; Calzacorta, C.; Prather, D.; Fologea, D. Insights into the Voltage Regulation Mechanism of the Pore-Forming Toxin Lysenin. Toxins 2018, 10, 334. https://doi.org/10.3390/toxins10080334
Bryant SL, Clark T, Thomas CA, Ware KS, Bogard A, Calzacorta C, Prather D, Fologea D. Insights into the Voltage Regulation Mechanism of the Pore-Forming Toxin Lysenin. Toxins. 2018; 10(8):334. https://doi.org/10.3390/toxins10080334
Chicago/Turabian StyleBryant, Sheenah Lynn, Tyler Clark, Christopher Alex Thomas, Kaitlyn Summer Ware, Andrew Bogard, Colleen Calzacorta, Daniel Prather, and Daniel Fologea. 2018. "Insights into the Voltage Regulation Mechanism of the Pore-Forming Toxin Lysenin" Toxins 10, no. 8: 334. https://doi.org/10.3390/toxins10080334
APA StyleBryant, S. L., Clark, T., Thomas, C. A., Ware, K. S., Bogard, A., Calzacorta, C., Prather, D., & Fologea, D. (2018). Insights into the Voltage Regulation Mechanism of the Pore-Forming Toxin Lysenin. Toxins, 10(8), 334. https://doi.org/10.3390/toxins10080334