Structural and Functional Investigation and Pharmacological Mechanism of Trichosanthin, a Type 1 Ribosome-Inactivating Protein


Abstract
:1. Introduction
2. The Structural Investigation of TCS
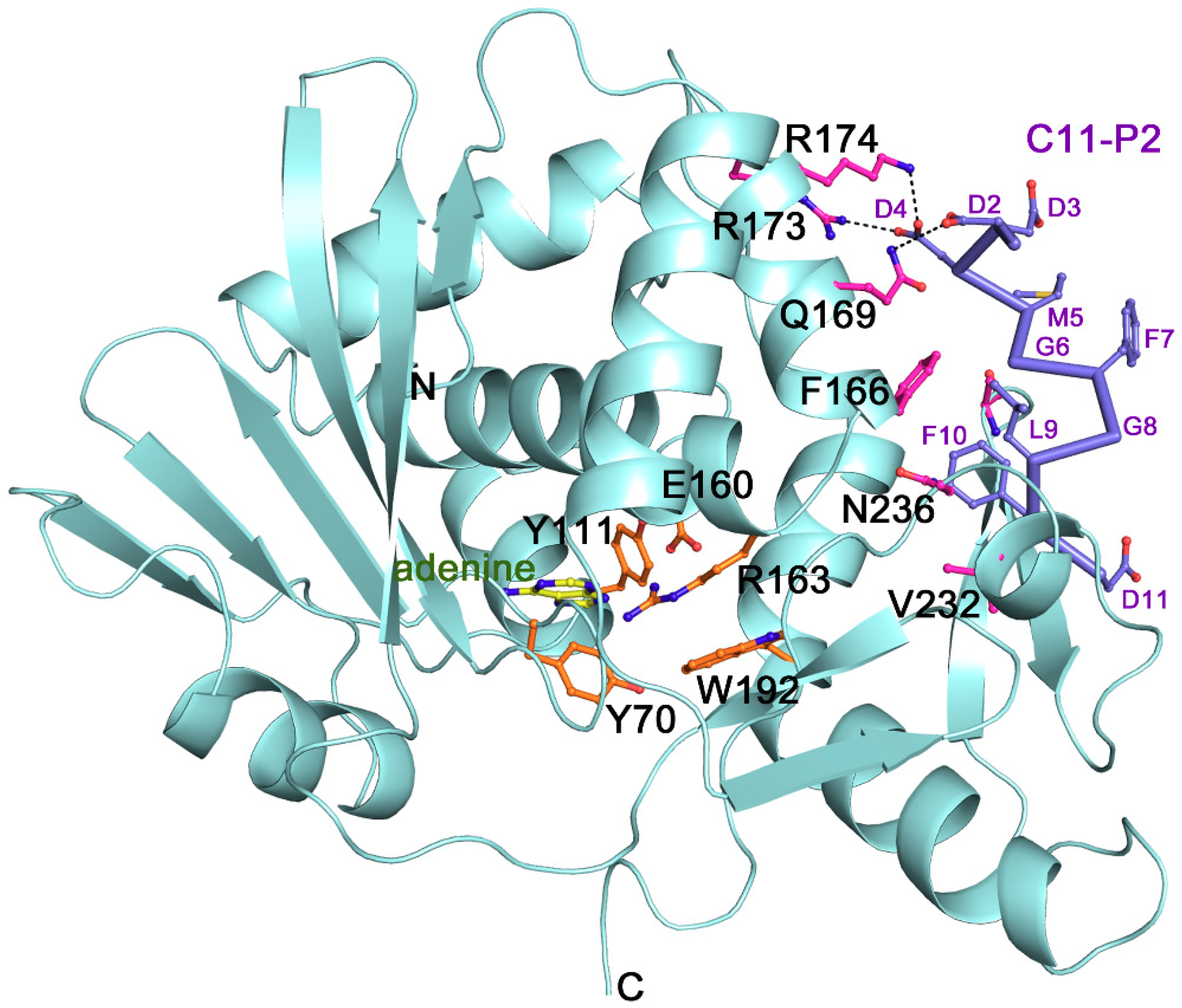
2.1. The Structural Feature and Ribosomal Interaction Mode of TCS
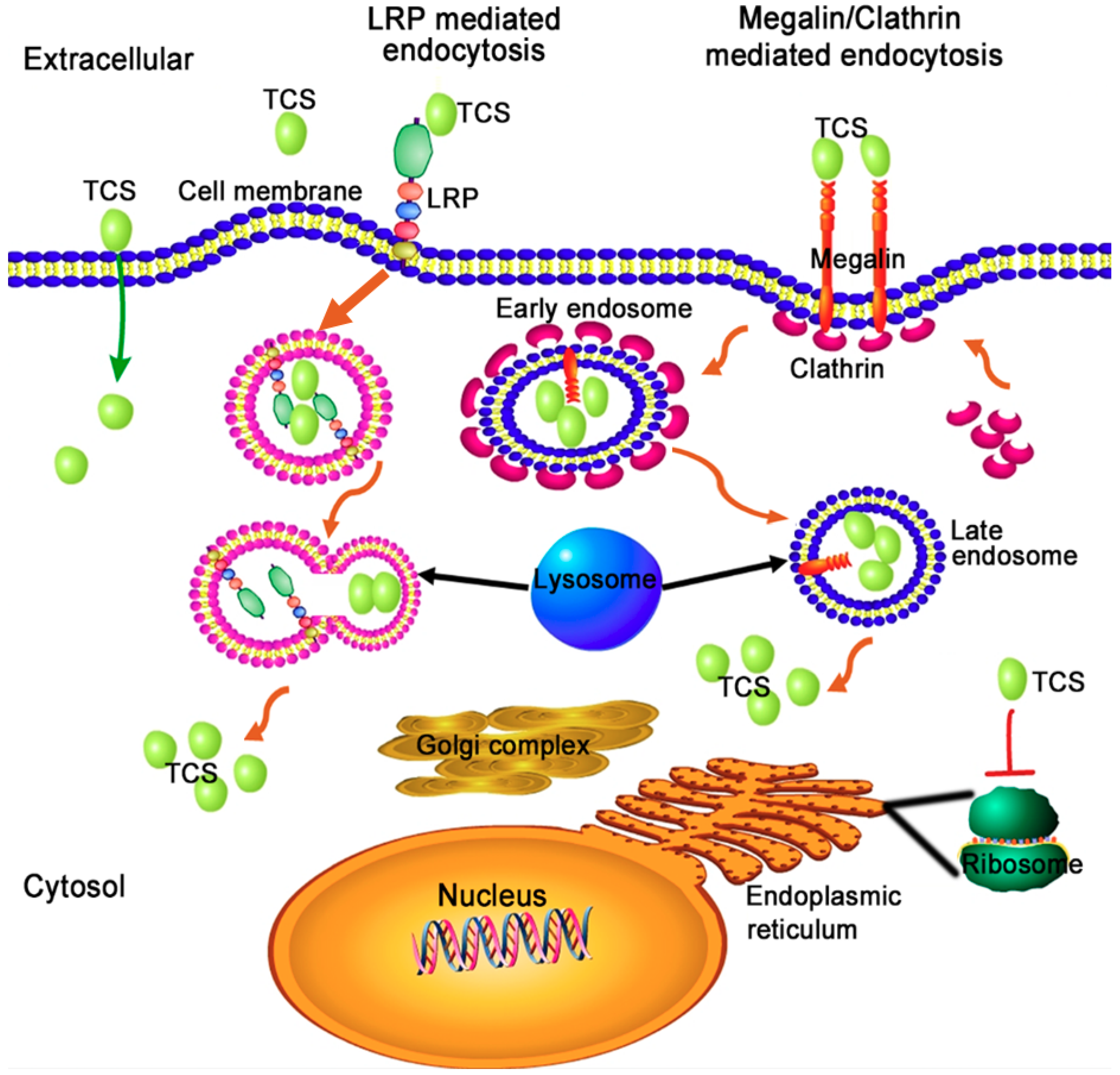
2.2. The Possible Pell-Entry Pathway of TCS
3. TCS Possesses Multiple Pharmacological Properties
3.1. Anti-Viral Activity of TCS
3.1.1. Anti-HIV-1 Activity
3.1.2. Anti-Hepatitis B Virus Activity
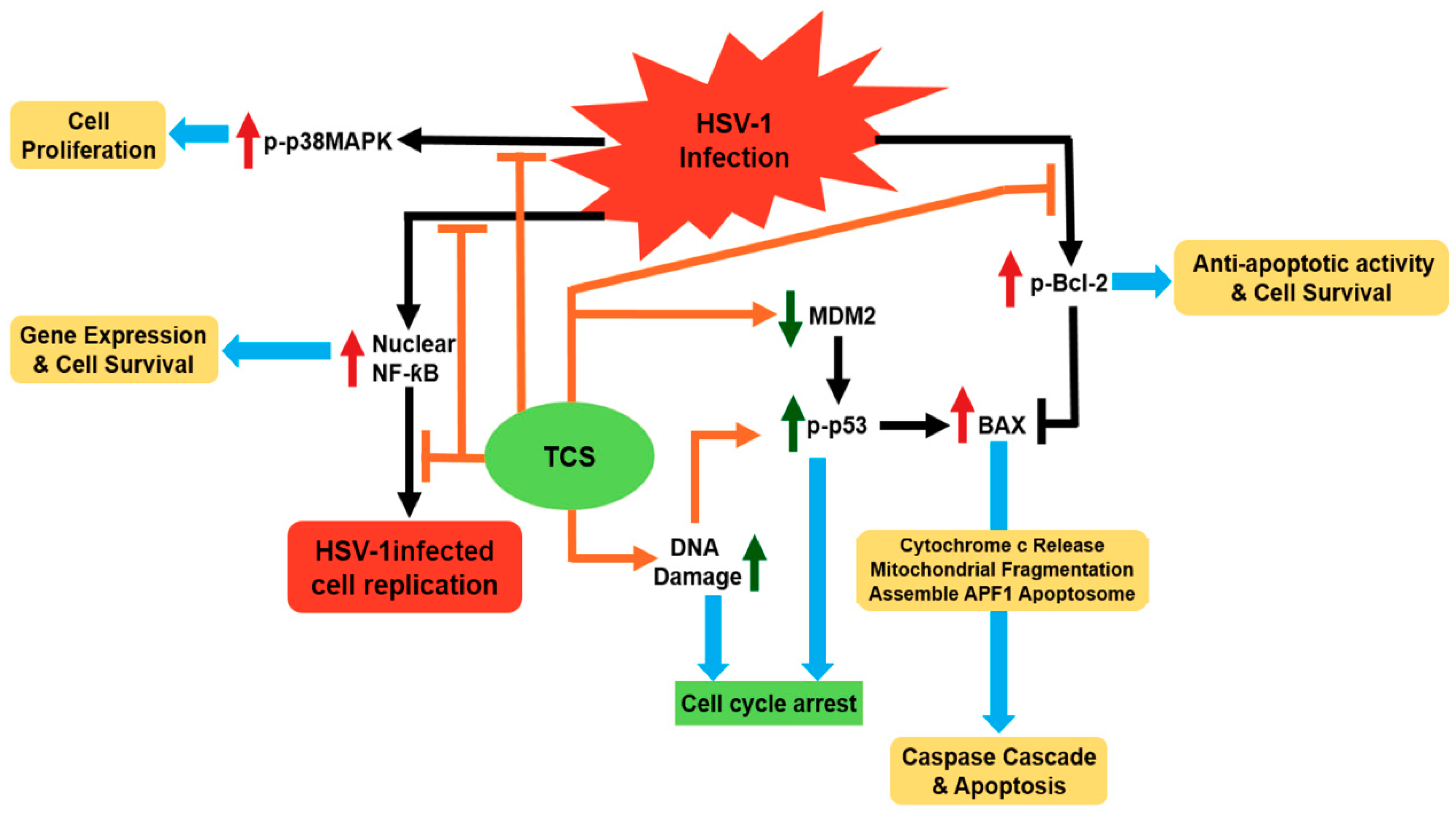
3.1.3. TCS has Protective Effect against Herpes Simplex Virus in Animal Model
3.2. The Anti-Tumor Activities of TCS
3.2.1. TCS Inhibits Various Tumor Cells
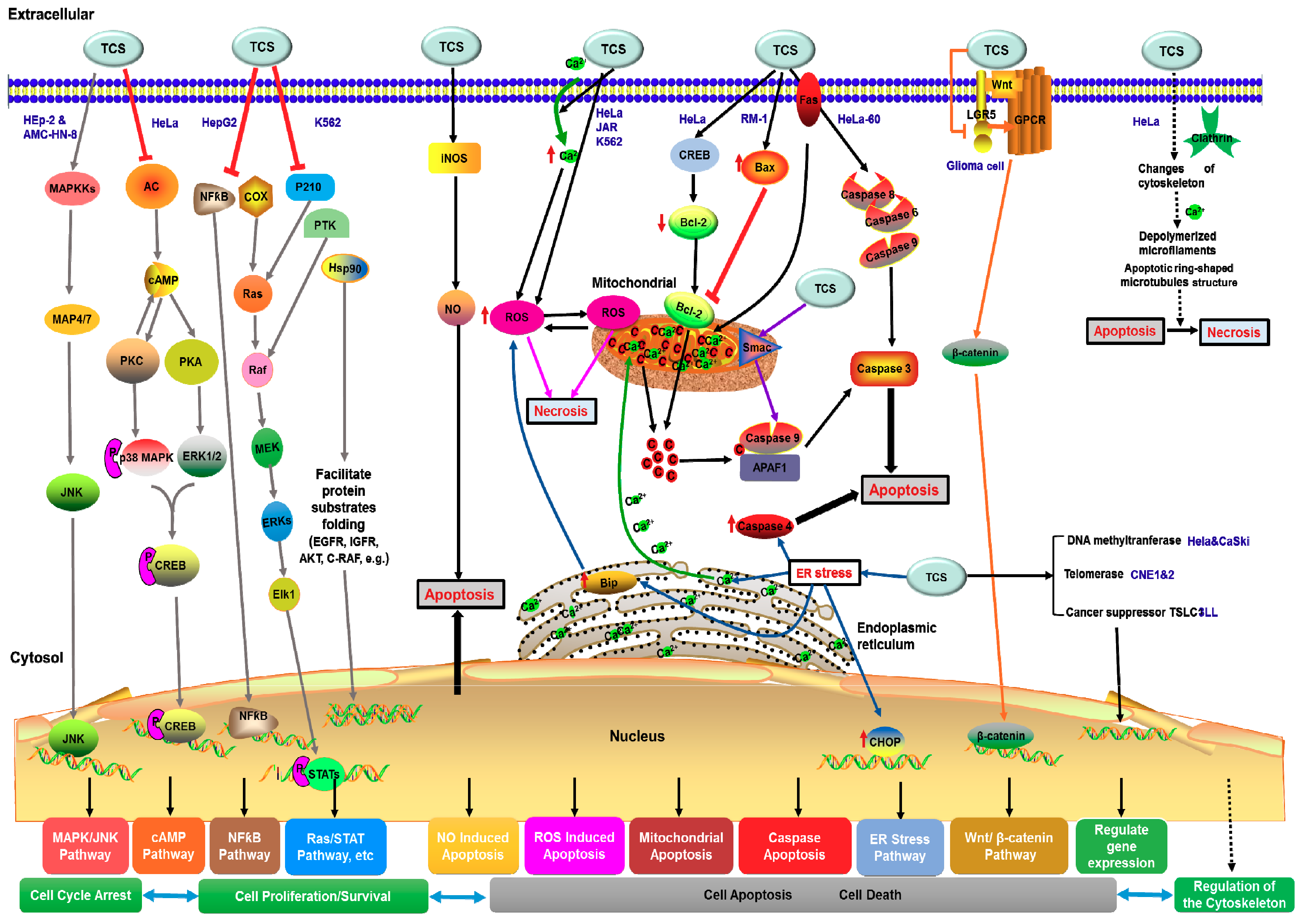
3.2.2. The Possible Anti-Tumor Mechanism of TCS
Apoptosis Related Anti-Tumor Mechanism
Autophagy-Related Anti-Tumor Mechanism
3.3. The Immunomodulatory Activity of TCS
4. TCS-Derivatives Are Promising Therapeutic Agents
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Maraganore, J.M.; Joseph, M.; Bailey, M.C. Purification and characterization of trichosanthin. Homology to the ricin A chain and implications as to mechanism of abortifacient activity. J. Biol. Chem. 1987, 262, 11628–11633. [Google Scholar] [PubMed]
- Byers, V.S.; Levin, A.S.; Waites, L.A.; Starrett, B.A.; Mayer, R.A.; Clegg, J.A.; Price, M.R.; Robins, R.A.; Delaney, M.; Baldwin, R.W. A phase I/II study of trichosanthin treatment of HIV disease. AIDS 1990, 4, 1189–1196. [Google Scholar] [CrossRef] [PubMed]
- Byers, V.S.; Baldwin, P.W. Trichosanthin treatment of HIV disease. AIDS 1991, 5, 1150–1151. [Google Scholar] [PubMed]
- Tsao, S.W.; Ng, T.B.; Yeung, H.W. Toxicities of trichosanthin and alpha-momorcharin, abortifacient proteins from Chinese medicinal plants, on cultured tumor cell lines. Toxicon 1990, 28, 1183–1192. [Google Scholar] [CrossRef]
- de Virgilio, M.; Lombardi, A.; Caliandro, R.; Fabbrini, M.S. Ribosome-inactivating proteins: From plant defense to tumor attack. Toxins 2010, 2, 2699–2737. [Google Scholar] [CrossRef] [PubMed]
- Pan, K.Z.; Lin, Y.J.; Zhou, K.J.; Fu, Z.J.; Chen, M.H.; Huang, D.R.; Huang, D.H. The crystal and molecular structure of trichosanthin at 2.6 A resolution. Sci. China B 1993, 36, 1069–1081. [Google Scholar] [PubMed]
- Shaw, P.C.; Chan, W.L.; Yeung, H.W.; Ng, T.B. Minireview: Trichosanthin—A protein with multiple pharmacological properties. Life Sci. 1994, 55, 253–262. [Google Scholar] [CrossRef]
- Wong, K.B.; Ke, Y.B.; Dong, Y.C.; Li, X.B.; Guo, Y.W.; Yeung, H.W.; Shaw, P.C. Structure/function relationship study of Gln156, Glu160 and Glu189 in the active site of trichosanthin. Eur. J. Biochem. 1994, 221, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Fang, E.F.; Ng, T.B.; Shaw, P.C.; Wong, R.N. Recent progress in medicinal investigations on trichosanthin and other ribosome inactivating proteins from the plant genus trichosanthes. Curr. Med. Chem. 2011, 18, 4410–4417. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, X.; Li, J.C. Possible mechanisms of trichosanthin-induced apoptosis of tumor cells. Anat. Rec. 2010, 293, 986–992. [Google Scholar] [CrossRef] [PubMed]
- Collins, E.J.; Robertus, J.D.; LoPresti, M.; Stone, K.L.; Williams, K.R.; Wu, P.; Hwang, K.; Piatak, M. Primary amino acid sequence of alpha-trichosanthin and molecular models for abrin A-chain and alpha-trichosanthin. J. Biol. Chem. 1990, 265, 8665–8669. [Google Scholar] [PubMed]
- Zhang, J.S.; Liu, W.Y. The mechanism of action of trichosanthin on eukaryotic ribosomes—RNA N-glycosidase activity of the cytotoxin. Nucleic Acids Res. 1992, 20, 1271–1275. [Google Scholar] [CrossRef] [PubMed]
- Endo, Y. Mechanism of action of ricin and related toxins on the inactivation of eukaryotic ribosomes. Cancer Treat Res. 1988, 37, 75–89. [Google Scholar] [PubMed]
- Peumans, W.J.; Hao, Q.; Van Damme, E.J. Ribosome-inactivating proteins from plants: More than RNA N-glycosidases? FASEB J. 2001, 15, 1493–1506. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, P.; Gear, R.W.; Green, P.G.; Levine, J.D. IB4-saporin attenuates acute and eliminates chronic muscle pain in the rat. Exp. Neurol. 2012, 233, 859–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilabert-Oriol, R.; Weng, A.; Mallinckrodt, B.; Melzig, M.F.; Fuchs, H.; Thakur, M. Immunotoxins constructed with ribosome-inactivating proteins and their enhancers: A lethal cocktail with tumor specific efficacy. Curr. Pharm. Des. 2014, 20, 6584–6643. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, N.; Tyagi, M.; Pachauri, M.; Ghosh, P.C. Potential therapeutic applications of plant toxin-ricin in cancer: Challenges and advances. Tumour Biol. 2015, 36, 8239–8246. [Google Scholar] [CrossRef] [PubMed]
- Byers, V.S.; Levin, A.S.; Malvino, A.; Waites, L.; Robins, R.A.; Baldwin, R.W. A phase II study of effect of addition of trichosanthin to zidovudine in patients with HIV disease and failing antiretroviral agents. AIDS Res. Hum. Retrovirus 1994, 10, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Bi, L.Q.; Liu, J.W.; Song, Y. The effect of trichosanthin on immunoregulatory T lymphocytes. Chin. J. Integr. Traditional West. Med. 1994, 14, 18–20. [Google Scholar]
- Gong, Q.; Deng, D.; Ding, J.; Wang, C.; Bian, Z.; Ye, Z.; Xu, J. Trichosanthin, an extract of Trichosanthes kirilowii, effectively prevents acute rejection of major histocompatibility complex-mismatched mouse skin allograft. Transpl. Proc. 2008, 40, 3714–3718. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, M.; Jin, H.; Tang, Y.; Wu, A.; Xu, Q.; Huang, Y. Prodrug-like, PEGylated protein toxin trichosanthin for reversal of chemoresistance. Mol. Pharm. 2017, 14, 1429–1438. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Nie, H.L.; Tam, S.C.; Huang, H.; Zheng, Y.T. Anti-HIV-1 property of trichosanthin correlates with its ribosome inactivating activity. FEBS Lett. 2002, 531, 295–298. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.F.; Huang, W.G.; Chen, F.Y.; Shan, J.L. Protective effects of trichosanthin in Herpes simplex virus-1 encephalitis in mice. Chin. J. Cont. Pediat. 2006, 8, 239–241. [Google Scholar]
- Wen, D.; Wang, J.; Yan, H.; Chen, J.; Xia, K.; Liu, J.; Zhang, A. Effect of radix trichosanthis and trichosanthin on Hepatitis B virus in HepG2.2.15 cells. J. Nanosci. Nanotechnol. 2015, 15, 2094–2098. [Google Scholar] [CrossRef] [PubMed]
- Chou, K.Y.; Chan, M.; Bias, W.B. Differential expression of the down-regulatory function of CD8 cells in trichosanthin-induced immunosuppression and its genetic control in humans. Eur. J. Immunogenet. 1996, 23, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Jiao, Z.; Pan, J.; Hong, J.; Tao, J.; Li, N.; Zhou, Y.; Zhang, J.; Chou, K.Y. Immune suppression via IL-4/IL-10-secreting t cells: A nontoxic property of anti-HIV agent trichosanthin. Clin. Immunol. 2007, 122, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.H.; Ji, Y.Y.; Yeh, M. The kinetics of IL-4 and IFN-gamma gene expression in mice after trichosansin immunization. Cell Res. 1998, 8, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.L.; Su, H.; Chen, Y.; Wang, J.; Xu, G.L. A role for trichosanthin in the expansion of CD4CD25 regulatory T cells. Scand. J. Immunol. 2010, 71, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Liu, S.; Tang, Y.; Jin, S.; Wang, Y. Studies on crystal structures, active-centre geometry and depurinating mechanism of two ribosome-inactivating proteins. Biochem. J. 1995, 309 Pt 1, 285–298. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Wu, S.; Li, H.G.; Li, J.H.; Wong, R.N.; Shi, Q.L.; Dong, Y.C. Role of Tyr70 in the N-glycosidase activity of neo-trichosanthin. Toxicon 1999, 37, 961–972. [Google Scholar] [CrossRef]
- Ding, Y.; Too, H.; Wang, Z.; Liu, Y.; Bartlam, M.; Dong, Y.; Wong, K.; Shaw, P.; Rao, Z. The structural basis of Trp192 and the C-terminal region in trichosanthin for activity and conformational stability. Protein Eng. 2003, 16, 351–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, S.H.; Hung, F.S.; Chan, D.S.; Shaw, P.C. Trichosanthin interacts with acidic ribosomal proteins P0 and P1 and mitotic checkpoint protein MAD2B. Eur. J. Biochem. 2001, 268, 2107–2112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tchorzewski, M. The acidic ribosomal P proteins. Int. J. Biochem. Cell Biol. 2002, 34, 911–915. [Google Scholar] [CrossRef]
- Wahl, M.C.; Moller, W. Structure and function of the acidic ribosomal stalk proteins. Curr. Protein Pept. Sci. 2002, 3, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo, P.; Reboud, J.P. The puzzling lateral flexible stalk of the ribosome. Biol. Cell. 2003, 95, 179–193. [Google Scholar] [CrossRef] [Green Version]
- Wilson, D.N.; Nierhaus, K.H. Ribosomal proteins in the spotlight. Crit. Rev. Biochem. Mol. Biol. 2005, 40, 243–267. [Google Scholar] [CrossRef] [PubMed]
- Nomura, N.; Honda, T.; Baba, K.; Naganuma, T.; Tanzawa, T.; Arisaka, F.; Noda, M.; Uchiyama, S.; Tanaka, I.; Yao, M.; et al. Archaeal ribosomal stalk protein interacts with translation factors in a nucleotide-independent manner via its conserved C terminus. Proc. Natl. Acad. Sci. USA 2012, 109, 3748–3753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, D.S.B.; Chu, L.O.; Lee, K.M.; Too, P.H.M.; Ma, K.W.; Sze, K.H.; Zhu, G.; Shaw, P.C.; Wong, K.B. Interaction between trichosanthin, a ribosome inactivating protein, and the ribosomal stalk protein P2 by chemical shift perturbation and mutagenesis analyses. Nucleic Acids Res. 2007, 35, 1660–1672. [Google Scholar] [CrossRef] [PubMed]
- McCluskey, A.J.; Poon, G.M.K.; Bolewska-Pedyczak, E.; Srikumar, T.; Jeram, S.M.; Raught, B.; Gariepy, J. The catalytic subunit of Shiga-like toxin 1 interacts with ribosomal stalk proteins and is inhibited by their conserved C-terminal domain. J. Mol. Biol. 2008, 378, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.H.; Mak, A.N.S.; Shaw, P.C.; Sze, K.H. Solution structure of an active mutant of maize ribosome-inactivating protein (MOD) and its interaction with the ribosomal stalk protein P2. J. Mol. Biol. 2010, 395, 897–907. [Google Scholar] [CrossRef] [PubMed]
- McCluskey, A.J.; Bolewska-Pedyczak, E.; Jarvik, N.; Chen, G.; Sidhu, S.S.; Gariepy, J. Charged and hydrophobic surfaces on the A chain of Shiga-like toxin 1 recognize the C-terminal domain of ribosomal stalk proteins. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Too, P.H.; Ma, M.K.; Mak, A.N.; Wong, Y.T.; Tung, C.K.; Zhu, G.; Au, S.W.; Wong, K.B.; Shaw, P.C. The C-terminal fragment of the ribosomal P protein complexed to trichosanthin reveals the interaction between the ribosome-inactivating protein and the ribosome. Nucleic Acids Res. 2009, 37, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.M.; Yu, C.W.; Chan, D.S.; Chiu, T.Y.; Zhu, G.; Sze, K.H.; Shaw, P.C.; Wong, K.B. Solution structure of the dimerization domain of ribosomal protein P2 provides insights for the structural organization of eukaryotic stalk. Nucleic Acids Res. 2010, 38, 5206–5216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.M.; Yu, C.W.; Chiu, T.Y.; Sze, K.H.; Shaw, P.C.; Wong, K.B. Solution structure of the dimerization domain of the eukaryotic stalk P1/P2 complex reveals the structural organization of eukaryotic stalk complex. Nucleic Acids Res. 2012, 40, 3172–3182. [Google Scholar] [CrossRef] [PubMed]
- Grela, P.; Helgstrand, M.; Krokowski, D.; Boguszewska, A.; Svergun, D.; Liljas, A.; Bernado, P.; Grankowski, N.; Akke, M.; Tchorzewski, M. Structural characterization of the ribosomal P1A-P2B protein dimer by small-angle X-ray scattering and NMR spectroscopy. Biochemistry 2007, 46, 1988–1998. [Google Scholar] [CrossRef] [PubMed]
- Diaconu, M.; Kothe, U.; Schlunzen, F.; Fischer, N.; Harms, J.M.; Tonevitsky, A.G.; Stark, H.; Rodnina, M.V.; Wahl, M.C. Structural basis for the function of the ribosomal L7/12 stalk in factor binding and gtpase activation. Cell 2005, 121, 991–1004. [Google Scholar] [CrossRef] [PubMed]
- Naganuma, T.; Nomura, N.; Yao, M.; Mochizuki, M.; Uchiumi, T.; Tanaka, I. Structural basis for translation factor recruitment to the eukaryotic/archaeal ribosomes. J. Biol. Chem. 2010, 285, 4747–4756. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.M.; Yusa, K.; Chu, L.O.; Yu, C.W.H.; Oono, M.; Miyoshi, T.; Ito, K.; Shaw, P.C.; Wong, K.B.; Uchiumi, T. Solution structure of human P1·P2 heterodimer provides insights into the role of eukaryotic stalk in recruiting the ribosome-inactivating protein trichosanthin to the ribosome. Nucleic Acids Res. 2013, 41, 8776–8787. [Google Scholar] [CrossRef] [PubMed]
- Bargis-Surgey, P.; Lavergne, J.P.; Gonzalo, P.; Vard, C.; Filhol-Cochet, O.; Reboud, J.P. Interaction of elongation factor eEF-2 with ribosomal P proteins. Eur. J. Biochem. 1999, 262, 606–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolan, R.D.; Grasmuk, H.; Drews, J. The binding of tritiated elongation-factors 1 and 2 to ribosomes from Krebs II mouse ascites-tumore cells. The influence of various antibiotics and toxins. Eur. J. Biochem. 1976, 64, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Honda, T.; Suzuki, T.; Miyoshi, T.; Murakami, R.; Yao, M.; Uchiumi, T. Molecular insights into the interaction of the ribosomal stalk protein with elongation factor 1alpha. Nucleic Acids Res. 2014, 42, 14042–14052. [Google Scholar] [CrossRef] [PubMed]
- Choi, A.K.; Wong, E.C.; Lee, K.M.; Wong, K.B. Structures of eukaryotic ribosomal stalk proteins and its complex with trichosanthin, and their implications in recruiting ribosome-inactivating proteins to the ribosomes. Toxins 2015, 7, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Shaw, P.C.; Lee, K.M.; Wong, K.B. Recent advances in trichosanthin, a ribosome-inactivating protein with multiple pharmacological properties. Toxicon 2005, 45, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.L.; Shaw, P.C.; Tam, S.C.; Jacobsen, C.; Gliemann, J.; Nielsen, M.S. Trichosanthin interacts with and enters cells via LDL receptor family members. Biochem. Biophys. Res. Commun 2000, 270, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.Y.; Huang, H.; Tam, S.C. Receptor-mediated endocytosis of trichosanthin in choriocarcinoma cells. Toxicology 2003, 186, 191–203. [Google Scholar] [CrossRef]
- Xia, X.F.; Zhang, F.; Shaw, P.C.; Sui, S.F. Trichosanthin induces leakage and membrane fusion of liposome. IUBMB Life 2003, 55, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Lu, Y.J.; Shaw, P.C.; Sui, S.F. Change in ph-dependent membrane insertion characteristics of trichosanthin caused by deletion of its last seven C-terminal amino acid residues. Biochemistry 2003, 68, 436–445. [Google Scholar] [PubMed]
- Zhang, C.Y.; Gong, Y.X.; Ma, H.; An, C.C.; Chen, D.Y. Trichosanthin induced Calcium-dependent generation of reactive oxygen species in human choriocarcinoma cells. Analyst 2000, 125, 1539–1542. [Google Scholar] [PubMed]
- Zhang, C.; Gong, Y.; Ma, H.; An, C.; Chen, D.; Chen, Z.L. Reactive oxygen species involved in trichosanthin-induced apoptosis of human choriocarcinoma cells. Biochem. J. 2001, 355, 653–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, E.F.; Zhang, C.Z.; Zhang, L.; Wong, J.H.; Chan, Y.S.; Pan, W.L.; Dan, X.L.; Yin, C.M.; Cho, C.H.; Ng, T.B. Trichosanthin inhibits breast cancer cell proliferation in both cell lines and nude mice by promotion of apoptosis. PLoS ONE 2012, 7, e41592. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Sun, Y.; Cai, Y.; Sha, O.; Jiang, W. Trichosanthin reduces the viability of SUDHL2 cells via the activation of the extrinsic and intrinsic apoptotic pathways. Mol. Med. Rep. 2016, 13, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.L.; Zheng, Y.T.; Huang, H.; Tam, S.C. Relationship between trichosanthin cytotoxicity and its intracellular concentration. Toxicology 2002, 177, 245–251. [Google Scholar] [CrossRef]
- Tang, Y.; Liang, J.; Wu, A.; Chen, Y.; Zhao, P.; Lin, T.; Zhang, M.; Xu, Q.; Wang, J.; Huang, Y. Co-delivery of trichosanthin and albendazole by nano-self-assembly for overcoming tumor multidrug-resistance and metastasis. ACS Appl. Mater. Interface 2017, 9, 26648–26664. [Google Scholar] [CrossRef] [PubMed]
- McGrath, M.S.; Santulli, S.; Gaston, I. Effects of GLQ223 on HIV replication in human monocyte/macrophages chronically infected in vitro with HIV. AIDS Res. Hum. Retroviruses 1990, 6, 1039–1043. [Google Scholar] [CrossRef] [PubMed]
- Kahn, J.O.; Gorelick, K.J.; Gatti, G.; Arri, C.J.; Lifson, J.D.; Gambertoglio, J.G.; Bostrom, A.; Williams, R. Safety, activity, and pharmacokinetics of GLQ223 in patients with aids and aids-related complex. Antimicrob. Agents Chemother. 1994, 38, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Tam, S.C.; Huang, H.; Ouyang, D.Y.; Wang, Y.Y.; Zheng, Y.T. Site-directed PEGylation of trichosanthin retained its anti-HIV activity with reduced potency in vitro. Biochem. Biophys. Res. Commun. 2004, 317, 965–971. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Nie, H.L.; Huang, H.; Tam, S.C.; Zheng, Y.T. Independency of anti-HIV-1 activity from ribosome-inactivating activity of trichosanthin. Biochem. Biophys. Res. Commun. 2003, 302, 89–94. [Google Scholar] [CrossRef]
- Zhao, W.; Feng, D.; Sun, S.; Han, T.; Sui, S. The anti-viral protein of trichosanthin penetrates into human immunodeficiency virus type 1. Acta Biochim. Biophys. Sin. (Shanghai) 2010, 42, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.L.; Zhang, F.; Feng, D.; Wu, J.; Chen, S.; Sui, S.F. A novel sorting strategy of trichosanthin for hijacking human immunodeficiency virus type 1. Biochem. Biophys. Res. Commun. 2009, 384, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.L.; Feng, D.; Wu, J.; Sui, S.F. Trichosanthin inhibits integration of human immunodeficiency virus type 1 through depurinating the long-terminal repeats. Mol. Biol. Rep. 2010, 37, 2093–2098. [Google Scholar] [CrossRef] [PubMed]
- Wutzler, P. Antiviral therapy of Herpes simplex and Varicella-zoster virus infections. Intervirology 1997, 40, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Hook, E.W., 3rd; Cannon, R.O.; Nahmias, A.J.; Lee, F.F.; Campbell, C.H., Jr.; Glasser, D.; Quinn, T.C. Herpes simplex virus infection as a risk factor for human immunodeficiency virus infection in heterosexuals. J. Infect. Dis. 1992, 165, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.T.; Chan, W.L.; Chan, P.; Huang, H.; Tam, S.C. Enhancement of the anti-herpetic effect of trichosanthin by acyclovir and interferon. FEBS Lett. 2001, 496, 139–142. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Chan, H.; Wang, Y.Y.; Ouyang, D.Y.; Zheng, Y.T.; Tam, S.C. Trichosanthin suppresses the elevation of P38 MAPK, and Bcl-2 induced by HSV-1 infection in vero cells. Life Sci. 2006, 79, 1287–1292. [Google Scholar] [CrossRef] [PubMed]
- He, D.X.; Tam, S.C. Trichosanthin affects HSV-1 replication in Hep-2 cells. Biochem. Biophys. Res. Commun. 2010, 402, 670–675. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Zheng, Y.; Tam, S. The anti-herpetic activity of trichosanthin via the nuclear factor-kappab and p53 pathways. Life Sci. 2012, 90, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.T.; Zhang, W.F.; Ben, K.L.; Wang, J.H. In vitro immunotoxicity and cytotoxicity of trichosanthin against human normal immunocytes and leukemia-lymphoma cells. Immunopharmacol. Immunotoxicol. 1995, 17, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Ru, Q.H.; Luo, G.A.; Liao, J.J.; Liu, Y. Capillary electrophoretic determination of apoptosis of hela cells induced by trichosanthin. J. Chromatogr. A 2000, 894, 165–170. [Google Scholar] [CrossRef]
- Peng, P.; Huang, L.; Wang, Y.; You, C.; Cao, W.; Song, H.; Tan, H.; Wu, Y. Effect of recombinant trichosanthin on proliferation of human cevical cancer caski cells. Chin. J. Chin. Mater. Med. 2011, 36, 2539–2542. [Google Scholar]
- Jiao, Y.; Liu, W. Low-density lipoprotein receptor-related protein 1 is an essential receptor for trichosanthin in 2 choriocarcinoma cell lines. Biochem. Biophys. Res. Commun. 2010, 391, 1579–1584. [Google Scholar] [CrossRef] [PubMed]
- Dou, C.M.; Li, J.C. Effect of extracts of trichosanthes root tubers on Hepa-H cells and HeLa cells. World J. Gastroenterol. 2004, 10, 2091–2094. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Gao, D.F.; Yan, G.L.; Fan, J.M. Induced apoptotic action of recombinant trichosanthin in human stomach adenocarcinoma MCG803 cells. Mol. Biol. Rep. 2009, 36, 1559–1564. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xia, X.; Ke, Y.; Nie, H.; Smith, M.A.; Zhu, X. Trichosanthin induced apoptosis in HL-60 cells via mitochondrial and endoplasmic reticulum stress signaling pathways. Biochim. Biophys. Acta 2007, 1770, 1169–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Xia, X.; Nie, H.; Smith, M.A.; Zhu, X. PKC inhibition is involved in trichosanthin-induced apoptosis in human chronic myeloid leukemia cell line K562. Biochim. Biophys. Acta 2007, 1770, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Li, C.T.; Lin, C.H.; Kao, T.Y.; Wu, M.F.; Yeh, C.S.; Yeh, K.T.; Ko, J.L. The mechanisms of action of tianhua on antitumor activity in lung cancer cells. Pharm. Biol. 2010, 48, 1302–1309. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Xiong, S.; Zheng, Y.; Luo, F.; Jiang, P.; Chu, Y. Trichosanthin enhances anti-tumor immune response in a murine lewis lung cancer model by boosting the interaction between TSLC1 and CRTAM. Cell. Mol. Immunol. 2011, 8, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.; Ou, H.; Wang, R.; Liu, W.; Mao, Y.; Tang, A. Effect of trichosanthin on apoptosis and telomerase activity of nasopharyngeal carcinomas in nude mice. J. BUON 2013, 18, 675–682. [Google Scholar] [PubMed]
- Li, J.; Li, H.; Zhang, Z.; Wang, N.; Zhang, Y. The anti-cancerous activity of recombinant trichosanthin on prostate cancer cell PC3. Biol. Res. 2016, 49. [Google Scholar] [CrossRef] [PubMed]
- Bi, L.; Li, H.; Zhang, Y. Effect of trichosanthin of cell cycle and apoptosis of murine melanoma cells. Zhongguo Zhong Xi Yi Jie He Za Zhi 1998, 18, 35–37. [Google Scholar] [PubMed]
- Miao, J.; Jiang, Y.; Wang, D.; Zhou, J.; Fan, C.; Jiao, F.; Liu, B.; Zhang, J.; Wang, Y.; Zhang, Q. Trichosanthin suppresses the proliferation of glioma cells by inhibiting lGR5 expression and the Wnt/beta-catenin signaling pathway. Oncol. Rep. 2015, 34, 2845–2852. [Google Scholar] [CrossRef] [PubMed]
- Sha, O.; Niu, J.; Ng, T.B.; Cho, E.Y.; Fu, X.; Jiang, W. Anti-tumor action of trichosanthin, a type 1 ribosome-inactivating protein, employed in traditional Chinese medicine: A mini review. Cancer Chemother. Pharmacol. 2013, 71, 1387–1393. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Mei, Y.; Wang, Y.; Chen, C.; Tu, J.; Xiao, B.; Xu, L. Trichosanthin inhibits antigen-specific T cell expansion through nitric oxide-mediated apoptosis pathway. Cell. Immunol. 2005, 234, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Chen, L.L.; Yan, H.; Li, J.C. Trichosanthin suppresses hela cell proliferation through inhibition of the PKC/MAPK signaling pathway. Cell Biol. Toxicol. 2009, 25, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Bai, T.; Shen, S.; Li, L.; Ding, H.; Wang, P. Increase of cytosolic calcium induced by trichosanthin suppresses CAMP/PKC levels through the inhibition of adenylyl cyclase activity in hela cells. Mol. Biol. Rep. 2011, 38, 2863–2868. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Song, H.; Hu, H.; Cui, L.; You, C.; Huang, L. Trichosanthin inhibits DNA methyltransferase and restores methylation-silenced gene expression in human cervical cancer cells. Mol. Med. Rep. 2012, 6, 872–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, L.; Song, J.; Wu, L.; Huang, L.; Wang, Y.; Huang, Y.; Yu, H.; Huang, Y.; You, C.C.; Ye, J. Smac is another pathway in the anti-tumour activity of trichosanthin and reverses trichosanthin resistance in CaSki cervical cancer cells. Biomed. Pharmacother. 2015, 69, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Chen, B.; Zhou, J.; Zhou, L.; Li, Q.; Liu, F.; Chou, K.Y.; Tao, L.; Lu, L.M. Low concentrations of trichosanthin induce apoptosis and cell cycle arrest via C-jun N-terminal protein kinase/mitogen-activated protein kinase activation. Mol. Med. Rep. 2015, 11, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Xu, J.; Zhang, C. Creb, a possible upstream regulator of Bcl-2 in trichosanthin-induced hela cell apoptosis. Mol. Biol. Rep. 2010, 37, 1891–1896. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.S.; Wu, L.P.; Li, K.H.; Liu, Y.P.; Xiang, R.; Zhang, S.B.; Zhu, L.Y.; Zhang, L.Y. Involvement of nuclear factor kappab (NF-kappab) in the downregulation of cyclooxygenase-2 (COX-2) by genistein in gastric cancer cells. J. Int. Med. Res. 2011, 39, 2141–2150. [Google Scholar] [CrossRef] [PubMed]
- Venkateswararao, E.; Anh Hle, T.; Sharma, V.K.; Lee, K.C.; Sharma, N.; Kim, Y.; Jung, S.H. Study on anti-proliferative effect of benzoxathiole derivatives through inactivation of NF-kappab in human cancer cells. Bioorg. Med. Chem. Lett. 2012, 22, 4523–4527. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Xu, J.; Huang, X.; Wu, L.; Wen, C.; Hu, Y.; Su, Y.; Chen, Y.; Zhang, Z. Trichosanthin down-regulated p210Bcr-Abl and enhanced imatinib-induced growth arrest in chronic myelogenous leukemia cell line K562. Cancer Chemother. Pharmacol. 2007, 60, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Shan, S.D.; Yuan, T.; Gui, Y.P.; Cao, C.H.; Zhang, J.F. Mechanism of trichosanthin inducing apoptosis of mouse prostatic cancer RM-1 cells in vitro. Zhong Yao Cai 2009, 32, 239–242. [Google Scholar] [PubMed]
- Wang, P.; Li, J.C. Trichosanthin-induced specific changes of cytoskeleton configuration were associated with the decreased expression level of actin and tubulin genes in apoptotic HeLa cells. Life Sci. 2007, 81, 1130–1140. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Huang, Q.; Huang, S.; Mai, W.; Zhong, X. Trichosanthin-induced autophagy in gastric cancer cell MKN-45 is dependent on reactive oxygen species (ROS) and Nf-kappaB/p53 pathway. J. Pharmacol. Sci. 2016, 131, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, S.; Surendranath, K.; Bora, N.; Surolia, A.; Karande, A.A. Ribosome inactivating proteins and apoptosis. FEBS Lett. 2005, 579, 1324–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, M.K.; Sharma, R.S.; Mishra, V. Induction of apoptosis by ribosome inactivating proteins: Importance of N-glycosidase activity. Appl. Biochem. Biotechnol. 2012, 166, 1552–1561. [Google Scholar] [CrossRef] [PubMed]
- Leung, K.N.; Yeung, H.W.; Leung, S.O. The immunomodulatory and antitumor activities of trichosanthin-an abortifacient protein isolated from tian-hua-fen (Trichosanthes kirilowii). Asian Pac. J. Allergy Immunol. 1986, 4, 111–120. [Google Scholar] [PubMed]
- Zhao, S.; Wang, Y.; Wei, H. Trichosanthin induced Th2 polarization status. Cell. Mol. Immunol. 2006, 3, 297–301. [Google Scholar] [PubMed]
- Wang, Q.C.; Ying, W.B.; Xie, H.; Zhang, Z.C.; Yang, Z.H.; Ling, L.Q. Trichosanthin-monoclonal antibody conjugate specifically cytotoxic to human hepatoma cells in vitro. Cancer Res. 1991, 51, 3353–3355. [Google Scholar] [PubMed]
- Shin, M.C.; Zhang, J.; Min, K.A.; He, H.; David, A.E.; Huang, Y.; Yang, V.C. PTD-modified attempts for enhanced toxin-based cancer therapy: An in vivo proof-of-concept study. Pharm Res. 2015, 32, 2690–2703. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Li, Z.; Jiao, Z.; Gu, P.; Zhou, Y.; Lu, L.; Chou, K.Y. A trichosanthin-derived peptide suppresses type 1 immune responses by TLR2-dependent activation of CD8 (+) CD 28 (−) Tregs. Clin. Immunol. 2014, 153, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zeng, M.; Chi, H.; Shen, J.; Ng, T.B.; Jin, G.; Lu, D.; Fan, X.; Xiong, B.; Xiao, Z.; et al. Trichosanthin increases granzyme B penetration into tumor cells by upregulation of CI-MPR on the cell surface. Oncotarget 2017, 8, 26460–26470. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Yang, X.Z.; Cao, X.W.; Zhang, T.Z.; Wang, F.J.; Zhao, J. A novel trichosanthin fusion protein with increased cytotoxicity to tumor cells. Biotechnol. Lett. 2017, 39, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.Z.; Li, P.F.; Li, Y.Z.; Luo, F.; Guo, C.; Lin, B.; Cao, X.W.; Zhao, J.; Wang, F.J. Enhanced anti-tumor activity of trichosanthin after combination with a human-derived cell-penetrating peptide, and a possible mechanism of activity. Fitoterapia 2016, 112, 183–190. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System | Tumor Type | Tested Cell Line | Tested Model |
|---|---|---|---|
| Female reproductive | Breast cancer | MCF-7, BT-474 and MDA-MB-231 [60] | Nude mice [60] |
| Cervical cancer | HeLa [78] and Caski cells [79] | - | |
| Choriocarcinoma | JAR [59] and BeWo [80] | - | |
| Immune | Lymphoma | SU-DHL-2 cells [61] | - |
| Digestive | Colon cancer | CT-26 [81] | - |
| Hepatoma | HepA-H cells [81] | - | |
| Gastric cancer | MCG803 [82] | - | |
| Blood | Leukemia | HL-60 [83] and K562 [84] | - |
| Respiratory | Lung cancer | A549 cells [85] and 3LL [86] | Nude mouse [85] Lewis rat murine models [86] |
| Nasopharyngeal cancer | CNE1 and CNE2 [87] | - | |
| Male reproductive | Prostate cancer | RM-1 [88] | - |
| Integumentary | Melanoma | B16 [89] | - |
| Nervous | Glioma | U87 and U251 [90] | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, W.-W.; Wong, K.-B.; Shaw, P.-C. Structural and Functional Investigation and Pharmacological Mechanism of Trichosanthin, a Type 1 Ribosome-Inactivating Protein. Toxins 2018, 10, 335. https://doi.org/10.3390/toxins10080335
Shi W-W, Wong K-B, Shaw P-C. Structural and Functional Investigation and Pharmacological Mechanism of Trichosanthin, a Type 1 Ribosome-Inactivating Protein. Toxins. 2018; 10(8):335. https://doi.org/10.3390/toxins10080335
Chicago/Turabian StyleShi, Wei-Wei, Kam-Bo Wong, and Pang-Chui Shaw. 2018. "Structural and Functional Investigation and Pharmacological Mechanism of Trichosanthin, a Type 1 Ribosome-Inactivating Protein" Toxins 10, no. 8: 335. https://doi.org/10.3390/toxins10080335
APA StyleShi, W. -W., Wong, K. -B., & Shaw, P. -C. (2018). Structural and Functional Investigation and Pharmacological Mechanism of Trichosanthin, a Type 1 Ribosome-Inactivating Protein. Toxins, 10(8), 335. https://doi.org/10.3390/toxins10080335