Beauvericin and Enniatins: In Vitro Intestinal Effects


Abstract
:1. Introduction
2. Beauvericin
2.1. In Vitro Effects of BEA on Human Intestinal Models
2.1.1. Cell Models
2.1.2. Simulated Intestinal Environment
2.2. In Vitro Effects of BEA on Species-Specific Intestinal Models
Cell Models
3. Enniatins
3.1. In Vitro Effects of ENNs on Human Intestinal Models
3.1.1. Cell Models
3.1.2. Simulated Intestinal Environment
3.2. In Vitro Effects of ENNs on Species-Specific Intestinal Models
3.2.1. Cell Models
3.2.2. In Vitro Rumen Models
4. Occurrence of Beauvericin and Enniatins in the Food Chain Products
5. Legislation
6. Conclusions and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Jestoi, M. Emerging fusarium-mycotoxins fusaproliferin, beauvericin, enniatins, and moniliformin: A review. Crit. Rev. Food Sci. Nutr. 2008, 48, 21–49. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Guo, C.; Yu, S.; Zhu, L.; Wang, Y.; Hu, H.; Deng, J. Progress in Mycotoxins Affecting Intestinal Mucosal Barrier Function. Int. J. Mol. Sci. 2019, 20, 2777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woloshuk, C.P.; Shim, W.B. Aflatoxins, fumonisins, and trichothecenes: A convergence of knowledge. Fems. Microbiol. Rev. 2013, 37, 94–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Čolović, R.; Puvača, N.; Cheli, F.; Avantaggiato, G.; Greco, D.; Đuragić, O.; Kos, J.; Pinotti, L. Decontamination of Mycotoxin-Contaminated Feedstuffs and Compound Feed. Toxins 2019, 11, 617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schatzmayr, G.; Streit, E. Global occurrence of mycotoxins in the food and feed chain: Facts and figures. World Mycotoxin J. 2013, 6, 213–222. [Google Scholar] [CrossRef]
- Zwartsen, A.; de Korte, T.; Nacken, P.; de Lange, D.W.; Westerink, R.H.S.; Hondebrink, L. Cardiotoxicity screening of illicit drugs and new psychoactive substances (NPS) in human iPSC-derived cardiomyocytes using microelectrode array (MEA) recordings. J. Mol. Cell Cardiol. 2019, 136, 102–112. [Google Scholar] [CrossRef]
- Vaclavikova, M.; Malachova, A.; Veprikova, Z.; Dzuman, Z.; Zachariasova, M.; Hajslova, J. ‘Emerging’ mycotoxins in cereals processing chains: Changes of enniatins during beer and bread making. Food Chem. 2013, 136, 750–757. [Google Scholar] [CrossRef]
- Singh, J.; Mehta, A. Rapid and sensitive detection of mycotoxins by advanced and emerging analytical methods: A review. Food Sci. Nutr. 2020, 8, 2183–2204. [Google Scholar] [CrossRef]
- Tolosa, J.; Rodríguez-Carrasco, Y.; Ferrer, E.; Mañes, J. Identification and Quantification of Enniatins and Beauvericin in Animal Feeds and Their Ingredients by LC-QTRAP/MS/MS. Metabolites 2019, 9, 33. [Google Scholar] [CrossRef] [Green Version]
- van der Fels-Klerx, H.J.; Olesen, J.E.; Naustvoll, L.J.; Friocourt, Y.; Mengelers, M.J.; Christensen, J.H. Climate change impacts on natural toxins in food production systems, exemplified by deoxynivalenol in wheat and diarrhetic shellfish toxins. Food Addit Contam. Part A Chem. Anal. Control Expo Risk Assess 2012, 29, 1647–1659. [Google Scholar] [CrossRef] [PubMed]
- Liew, W.-P.-P.; Mohd-Redzwan, S. Mycotoxin: Its Impact on Gut Health and Microbiota. Front. Cell. Infect. Microbiol. 2018, 8, 60. [Google Scholar] [CrossRef] [Green Version]
- Medina, A.; Akbar, A.; Baazeem, A.; Rodriguez, A.; Magan, N. Climate change, food security and mycotoxins: Do we know enough? Fungal Biol. Rev. 2017, 31, 143–154. [Google Scholar] [CrossRef] [Green Version]
- Fraeyman, S.; Croubels, S.; Devreese, M.; Antonissen, G. Emerging Fusarium and Alternaria Mycotoxins: Occurrence, Toxicity and Toxicokinetics. Toxins 2017, 9, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caloni, F.; Fossati, P.; Anadón, A.; Bertero, A. Beauvericin: The beauty and the beast. Environ. Toxicol. Pharmacol. 2020, 75, 103349. [Google Scholar] [CrossRef]
- EFSA. Scientific opinion on the risks to human and animal health related to the presence of beuvericin and enniatins in food and feed. EFSA J. 2014, 12, 3802. [Google Scholar] [CrossRef]
- Broom, L. Mycotoxins and the intestine. Anim. Nutr. 2015, 1, 262–265. [Google Scholar] [CrossRef]
- Barac, A. Mycotoxins and Human Disease. In Clinically Relevant Mycoses: A Practical Approach; Presterl, E., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 213–225. [Google Scholar]
- Zain, M.E. Impact of mycotoxins on humans and animals. J. Saudi Chem. Soc. 2011, 15, 129–144. [Google Scholar] [CrossRef] [Green Version]
- Alassane-Kpembi, I.; Pinton, P.; Oswald, I.P. Effects of Mycotoxins on the Intestine. Toxins 2019, 11, 159. [Google Scholar] [CrossRef] [Green Version]
- Robert, H.; Payros, D.; Pinton, P.; Théodorou, V.; Mercier-Bonin, M.; Oswald, I.P. Impact of mycotoxins on the intestine: Are mucus and microbiota new targets? J. Toxicol. Env. Health B Crit. Rev. 2017, 20, 249–275. [Google Scholar] [CrossRef]
- Bertero, A.; Augustyniak, J.; Buzanska, L.; Caloni, F. Species-specific models in toxicology: In vitro epithelial barriers. Env. Toxicol. Pharm. 2019, 70, 103203. [Google Scholar] [CrossRef] [PubMed]
- Hamill, R.L.; Higgens, C.E.; Boaz, H.E.; Gorman, M. The structure op beauvericin, a new depsipeptide antibiotic toxic to artemia salina. Tetrahedron Lett. 1969, 10, 4255–4258. [Google Scholar] [CrossRef]
- Luangsa-ard, J.J.; Berkaew, P.; Ridkaew, R.; Hywel-Jones, N.L.; Isaka, M. A beauvericin hot spot in the genus Isaria. Mycol. Res. 2009, 113, 1389–1395. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Xu, W.; Zhang, J.; Xu, J.; Li, F. Natural Occurrence of Beauvericin and Enniatins in Corn- and Wheat-Based Samples Harvested in 2017 Collected from Shandong Province, China. Toxins 2018, 11, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jajić, I.; Dudaš, T.; Krstović, S.; Krska, R.; Sulyok, M.; Bagi, F.; Savić, Z.; Guljaš, D.; Stankov, A. Emerging Fusarium Mycotoxins Fusaproliferin, Beauvericin, Enniatins, and Moniliformin in Serbian Maize. Toxins 2019, 11, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svingen, T.; Lund Hansen, N.; Taxvig, C.; Vinggaard, A.M.; Jensen, U.; Have Rasmussen, P. Enniatin B and beauvericin are common in Danish cereals and show high hepatotoxicity on a high-content imaging platform. Environ. Toxicol. 2017, 32, 1658–1664. [Google Scholar] [CrossRef] [PubMed]
- Zinedine, A.; Meca, G.; Mañes, J.; Font, G. Further data on the occurrence of Fusarium emerging mycotoxins enniatins (A, A1, B, B1), fusaproliferin and beauvericin in raw cereals commercialized in Morocco. Food Control 2011, 22, 1–5. [Google Scholar] [CrossRef]
- Abdallah, M.F.; Girgin, G.; Baydar, T.; Krska, R.; Sulyok, M. Occurrence of multiple mycotoxins and other fungal metabolites in animal feed and maize samples from Egypt using LC-MS/MS. J. Sci. Food Agric. 2017, 97, 4419–4428. [Google Scholar] [CrossRef]
- Beccari, G.; Colasante, V.; Tini, F.; Senatore, M.T.; Prodi, A.; Sulyok, M.; Covarelli, L. Causal agents of Fusarium head blight of durum wheat (Triticum durum Desf.) in central Italy and their in vitro biosynthesis of secondary metabolites. Food Microbiol. 2018, 70, 17–27. [Google Scholar] [CrossRef]
- Streit, E.; Schwab, C.; Sulyok, M.; Naehrer, K.; Krska, R.; Schatzmayr, G. Multi-mycotoxin screening reveals the occurrence of 139 different secondary metabolites in feed and feed ingredients. Toxins 2013, 5, 504–523. [Google Scholar] [CrossRef] [Green Version]
- Mallebrera, B.; Prosperini, A.; Font, G.; Ruiz, M.J. In vitro mechanisms of Beauvericin toxicity: A review. Food Chem. Toxicol. 2018, 111, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Patocka, J.; Nepovimova, E.; Kuca, K. A Review on the Synthesis and Bioactivity Aspects of Beauvericin, a Fusarium Mycotoxin. Front. Pharm. 2018, 9, 1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolosa, J.; Font, G.; Mañes, J.; Ferrer, E. Nuts and dried fruits: Natural occurrence of emerging Fusarium mycotoxins. Food Control 2013, 33, 215–220. [Google Scholar] [CrossRef]
- Jestoi, M.; Rokka, M.; Järvenpää, E.; Peltonen, K. Determination of Fusarium mycotoxins beauvericin and enniatins (A, A1, B, B1) in eggs of laying hens using liquid chromatography–tandem mass spectrometry (LC–MS/MS). Food Chem. 2009, 115, 1120–1127. [Google Scholar] [CrossRef]
- Prosperini, A.; Juan-García, A.; Font, G.; Ruiz, M.J. Beauvericin-induced cytotoxicity via ROS production and mitochondrial damage in Caco-2 cells. Toxicol. Lett. 2013, 222, 204–211. [Google Scholar] [CrossRef]
- Fernández-Blanco, C.; Frizzell, C.; Shannon, M.; Ruiz, M.-J.; Connolly, L. An in vitro investigation on the cytotoxic and nuclear receptor transcriptional activity of the mycotoxins fumonisin B1 and beauvericin. Toxicol. Lett. 2016, 257, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Dornetshuber, R.; Heffeter, P.; Lemmens-Gruber, R.; Elbling, L.; Marko, D.; Micksche, M.; Berger, W. Oxidative stress and DNA interactions are not involved in Enniatin- and Beauvericin-mediated apoptosis induction. Mol. Nutr. Food Res. 2009, 53, 1112–1122. [Google Scholar] [CrossRef]
- Zouaoui, N.; Mallebrera, B.; Berrada, H.; Abid-Essefi, S.; Bacha, H.; Ruiz, M.J. Cytotoxic effects induced by patulin, sterigmatocystin and beauvericin on CHO-K1 cells. Food Chem. Toxicol. 2016, 89, 92–103. [Google Scholar] [CrossRef]
- Ferrer, E.; Juan-García, A.; Font, G.; Ruiz, M.J. Reactive oxygen species induced by beauvericin, patulin and zearalenone in CHO-K1 cells. Toxicol. Vitr. 2009, 23, 1504–1509. [Google Scholar] [CrossRef]
- Mallebrera, B.; Juan-Garcia, A.; Font, G.; Ruiz, M.J. Mechanisms of beauvericin toxicity and antioxidant cellular defense. Toxicol. Lett. 2016, 246, 28–34. [Google Scholar] [CrossRef]
- Ruiz, M.J.; Franzova, P.; Juan-García, A.; Font, G. Toxicological interactions between the mycotoxins beauvericin, deoxynivalenol and T-2 toxin in CHO-K1 cells in vitro. Toxicon 2011, 58, 315–326. [Google Scholar] [CrossRef]
- Ficheux, A.S.; Sibiril, Y.; Parent-Massin, D. Effects of beauvericin, enniatin b and moniliformin on human dendritic cells and macrophages: An in vitro study. Toxicon 2013, 71, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jow, G.-M.; Chou, C.-J.; Chen, B.-F.; Tsai, J.-H. Beauvericin induces cytotoxic effects in human acute lymphoblastic leukemia cells through cytochrome c release, caspase 3 activation: The causative role of calcium. Cancer Lett. 2004, 216, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Manyes, L.; Escrivá, L.; Ruiz, M.J.; Juan-García, A. Beauvericin and enniatin B effects on a human lymphoblastoid Jurkat T-cell model. Food Chem. Toxicol. 2018, 115, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Escrivá, L.; Jennen, D.; Caiment, F.; Manyes, L. Transcriptomic study of the toxic mechanism triggered by beauvericin in Jurkat cells. Toxicol. Lett. 2018, 284, 213–221. [Google Scholar] [CrossRef]
- Ivanova, L.; Skjerve, E.; Eriksen, G.S.; Uhlig, S. Cytotoxicity of enniatins A, A1, B, B1, B2 and B3 from Fusarium avenaceum. Toxicon 2006, 47, 868–876. [Google Scholar] [CrossRef]
- Juan-García, A.; Ruiz, M.-J.; Font, G.; Manyes, L. Enniatin A1, enniatin B1 and beauvericin on HepG2: Evaluation of toxic effects. Food Chem. Toxicol. 2015, 84, 188–196. [Google Scholar] [CrossRef]
- Klarić, M.Š.; Rumora, L.; Ljubanović, D.; Pepeljnjak, S. Cytotoxicity and apoptosis induced by fumonisin B1, beauvericin and ochratoxin A in porcine kidney PK15 cells: Effects of individual and combined treatment. Arch. Toxicol. 2008, 82, 247–255. [Google Scholar] [CrossRef]
- Fraeyman, S.; Meyer, E.; Devreese, M.; Antonissen, G.; Demeyere, K.; Haesebrouck, F.; Croubels, S. Comparative in vitro cytotoxicity of the emerging Fusarium mycotoxins beauvericin and enniatins to porcine intestinal epithelial cells. Food Chem. Toxicol. 2018, 121, 566–572. [Google Scholar] [CrossRef]
- Meca, G.; Sospedra, I.; Soriano, J.M.; Ritieni, A.; Moretti, A.; Mañes, J. Antibacterial effect of the bioactive compound beauvericin produced by Fusarium proliferatum on solid medium of wheat. Toxicon 2010, 56, 349–354. [Google Scholar] [CrossRef]
- Dzoyem, J.P.; Melong, R.; Tsamo, A.T.; Maffo, T.; Kapche, D.G.W.F.; Ngadjui, B.T.; McGaw, L.J.; Eloff, J.N. Cytotoxicity, antioxidant and antibacterial activity of four compounds produced by an endophytic fungus Epicoccum nigrum associated with Entada abyssinica. Rev. Bras. Farmacogn. 2017, 27, 251–253. [Google Scholar] [CrossRef]
- Zhang, H.; Ruan, C.; Bai, X.; Zhang, M.; Zhu, S.; Jiang, Y. Isolation and Identification of the Antimicrobial Agent Beauvericin from the Endophytic Fusarium oxysporum 5-19 with NMR and ESI-MS/MS. Biomed. Res. Int. 2016, 2016, 1084670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sood, S.; Sandhu, S.; Mukherjee, T. Pharmacological and Therapeutic Potential of Beauvericin: A Short Review. J. Proteom. Bioinform. 2017, 10, 18–23. [Google Scholar] [CrossRef]
- Nilanonta, C.; Isaka, M.; Kittakoop, P.; Palittapongarnpim, P.; Kamchonwongpaisan, S.; Pittayakhajonwut, D.; Tanticharoen, M.; Thebtaranonth, Y. Antimycobacterial and antiplasmodial cyclodepsipeptides from the insect pathogenic fungus Paecilomyces tenuipes BCC 1614. Planta Med. 2000, 66, 756–758. [Google Scholar] [CrossRef] [PubMed]
- Shin, C.G.; An, D.G.; Song, H.H.; Lee, C. Beauvericin and enniatins H, I and MK1688 are new potent inhibitors of human immunodeficiency virus type-1 integrase. J. Antibiot. 2009, 62, 687–690. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yan, K.; Zhang, Y.; Huang, R.; Bian, J.; Zheng, C.; Sun, H.; Chen, Z.; Sun, N.; An, R.; et al. High-throughput synergy screening identifies microbial metabolites as combination agents for the treatment of fungal infections. Proc. Natl. Acad. Sci. USA 2007, 104, 4606–4611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, Y.; Liu, M.; Zhang, Y.; Liu, X.; Huang, R.; Song, F.; Dai, H.; Ren, B.; Sun, N.; Pei, G.; et al. Beauvericin counteracted multi-drug resistant Candida albicans by blocking ABC transporters. Synth. Syst. Biotechnol. 2016, 1, 158–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grove, J.F.; Pople, M. The insecticidal activity of beauvericin and the enniatin complex. Mycopathologia 1980, 70, 103–105. [Google Scholar] [CrossRef]
- Gupta, S.; Krasnoff, S.B.; Underwood, N.L.; Renwick, J.A.A.; Roberts, D.W. Isolation of beauvericin as an insect toxin from Fusarium semitectum and Fusarium moniliforme var. subglutinans. Mycopathologia 1991, 115, 185–189. [Google Scholar] [CrossRef]
- Leland, J.E.; McGuire, M.R.; Grace, J.A.; Jaronski, S.T.; Ulloa, M.; Park, Y.-H.; Plattner, R.D. Strain selection of a fungal entomopathogen, Beauveria bassiana, for control of plant bugs (Lygus spp.) (Heteroptera: Miridae). Biol. Control 2005, 35, 104–114. [Google Scholar] [CrossRef]
- Ganassi, S.; Moretti, A.; Maria Bonvicini Pagliai, A.; Logrieco, A.; Agnese Sabatini, M. Effects of beauvericin on Schizaphis graminum (Aphididae). J. Invertebr. Pathol. 2002, 80, 90–96. [Google Scholar] [CrossRef]
- Fornelli, F.; Minervini, F.; Logrieco, A. Cytotoxicity of fungal metabolites to lepidopteran (Spodoptera frugiperda) cell line (SF-9). J. Invertebr. Pathol. 2004, 85, 74–79. [Google Scholar] [CrossRef]
- Shimada, A.; Fujioka, S.; Koshino, H.; Kimura, Y. Nematicidal activity of beauvericin produced by the fungus Fusarium bulbicola. Z. Nat. C J. Biosci. 2010, 65, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Paciolla, C.; Dipierro, N.; Mulè, G.; Logrieco, A.; Dipierro, S. The mycotoxins beauvericin and T-2 induce cell death and alteration to the ascorbate metabolism in tomato protoplasts. Physiol. Mol. Plant Pathol. 2004, 65, 49–56. [Google Scholar] [CrossRef]
- García-Herranz, V.; Valdehita, A.; Navas, J.M.; Fernández-Cruz, M.L. Cytotoxicity against fish and mammalian cell lines and endocrine activity of the mycotoxins beauvericin, deoxynivalenol and ochratoxin-A. Food Chem. Toxicol. 2019, 127, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Albonico, M.; Schutz, L.F.; Caloni, F.; Cortinovis, C.; Spicer, L.J. In vitro effects of the Fusarium mycotoxins fumonisin B1 and beauvericin on bovine granulosa cell proliferation and steroid production. Toxicon 2017, 128, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Zhan, J.; Burns, A.M.; Liu, M.X.; Faeth, S.H.; Gunatilaka, A.A.L. Search for Cell Motility and Angiogenesis Inhibitors with Potential Anticancer Activity: Beauvericin and Other Constituents of Two Endophytic Strains of Fusarium oxysporum. J. Nat. Prod. 2007, 70, 227–232. [Google Scholar] [CrossRef] [Green Version]
- Heilos, D.; Rodríguez-Carrasco, Y.; Englinger, B.; Timelthaler, G.; van Schoonhoven, S.; Sulyok, M.; Boecker, S.; Süssmuth, R.D.; Heffeter, P.; Lemmens-Gruber, R.; et al. The Natural Fungal Metabolite Beauvericin Exerts Anticancer Activity In Vivo: A Pre-Clinical Pilot Study. Toxins 2017, 9, 258. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Patocka, J.; Kuca, K. Beauvericin, a Fusarium Mycotoxin: Anticancer Activity, Mechanisms, and Human Exposure Risk Assessment. Mini Rev. Med. Chem. 2019, 19, 206–214. [Google Scholar] [CrossRef]
- Lu, C.L.; Lin, H.I.; Chen, B.F.; Jow, G.M. Beauvericin-induced cell apoptosis through the mitogen-activated protein kinase pathway in human nonsmall cell lung cancer A549 cells. J. Toxicol. Sci. 2016, 41, 429–437. [Google Scholar] [CrossRef] [Green Version]
- Mallebrera, B.; Font, G.; Ruiz, M.J. Disturbance of antioxidant capacity produced by beauvericin in CHO-K1 cells. Toxicol. Lett. 2014, 226, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Celik, M.; Aksoy, H.; Yilmaz, S. Evaluation of beauvericin genotoxicity with the chromosomal aberrations, sister-chromatid exchanges and micronucleus assays. Ecotoxicol Environ. Saf. 2010, 73, 1553–1557. [Google Scholar] [CrossRef] [PubMed]
- Mastrorocco, A.; Martino, N.A.; Marzano, G.; Lacalandra, G.M.; Ciani, E.; Roelen, B.A.J.; Dell’Aquila, M.E.; Minervini, F. The mycotoxin beauvericin induces oocyte mitochondrial dysfunction and affects embryo development in the juvenile sheep. Mol. Reprod. Dev. 2019, 86, 1430–1443. [Google Scholar] [CrossRef]
- Tomoda, H.; Huang, X.H.; Cao, J.; Nishida, H.; Nagao, R.; Okuda, S.; Tanaka, H.; Omura, S.; Arai, H.; Inoue, K. Inhibition of acyl-CoA: Cholesterol acyltransferase activity by cyclodepsipeptide antibiotics. J. Antibiot. 1992, 45, 1626–1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dornetshuber, R.; Heffeter, P.; Sulyok, M.; Schumacher, R.; Chiba, P.; Kopp, S.; Koellensperger, G.; Micksche, M.; Lemmens-Gruber, R.; Berger, W. Interactions between ABC-transport proteins and the secondary Fusarium metabolites enniatin and beauvericin. Mol. Nutr. Food Res. 2009, 53, 904–920. [Google Scholar] [CrossRef] [PubMed]
- Mei, L.; Zhang, L.; Dai, R. An inhibition study of beauvericin on human and rat cytochrome P450 enzymes and its pharmacokinetics in rats. J. Enzym. Inhib. Med. Chem. 2009, 24, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Tran, V.N.; Viktorova, J.; Augustynkova, K.; Jelenova, N.; Dobiasova, S.; Rehorova, K.; Fenclova, M.; Stranska-Zachariasova, M.; Vitek, L.; Hajslova, J.; et al. In Silico and In Vitro Studies of Mycotoxins and Their Cocktails; Their Toxicity and Its Mitigation by Silibinin Pre-Treatment. Toxins 2020, 12, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prosperini, A.; Meca, G.; Fau-Font, G.; Font, G.; Fau-Ruiz, M.J.; Ruiz, M.J. Study of the cytotoxic activity of beauvericin and fusaproliferin and bioavailability in vitro on Caco-2 cells. Food Chem. Toxicol. 2012, 50, 5. [Google Scholar] [CrossRef]
- Font, G.; Prosperini, A.; Ruiz, M.J. Cytotoxicity, biaccessibility and transport by Caco-2 cells of enniatins and beauvericin. Toxicol. Lett. 2011, 205, S159. [Google Scholar] [CrossRef]
- Olleik, H.; Nicoletti, C.; Lafond, M.; Courvoisier-Dezord, E.; Xue, P.; Hijazi, A.; Baydoun, E.; Perrier, J.; Maresca, M. Comparative Structure-Activity Analysis of the Antimicrobial Activity, Cytotoxicity, and Mechanism of Action of the Fungal Cyclohexadepsipeptides Enniatins and Beauvericin. Toxins 2019, 11, 514. [Google Scholar] [CrossRef] [Green Version]
- Salim, A.; Nadri, S.; Hosseini, M.-J.; Rokni-Zadeh, H.; Mohseni, M. Protective effect of probiotic Lactobacillus acidophilus against the toxicity of beauvericin mycotoxin on the Caco-2 cell line. Toxicon 2020, 185, 184–187. [Google Scholar] [CrossRef]
- Albonico, M.; Cortinovis, C.; Caloni, F. In vitro toxicity of beauvericin alone and combined with fumonisin B1 or deoxynivalenol on Caco-2 cells. Int. J. Health Anim. Sci. Food Saf. 2016, 3. [Google Scholar] [CrossRef]
- Gordon, S.; Daneshian, M.; Bouwstra, J.; Caloni, F.; Constant, S.; Davies, D.E.; Dandekar, G.; Guzman, C.A.; Fabian, E.; Haltner, E.; et al. Non-animal models of epithelial barriers (skin, intestine and lung) in research, industrial applications and regulatory toxicology. Altex 2015, 32, 327–378. [Google Scholar] [CrossRef] [PubMed]
- Artursson, P.; Karlsson, J. Correlation between oral drug absorption in humans and apparent drug permeability coefficients in human intestinal epithelial (Caco-2) cells. Biochem. Biophys. Res. Commun. 1991, 175, 880–885. [Google Scholar] [CrossRef]
- Naseer, O.; Khan, J.A.; Khan, M.S.; Omer, M.O.; Chishti, G.A.; Sohail, M.L.; Saleem, M.U. Comparative Efficacy of Silymarin and Choline Chloride (Liver Tonics) in Preventing the Effects of Aflatoxin B1 in Bovine Calves. Pol. J. Vet. Sci. 2016, 19, 545–551. [Google Scholar] [CrossRef] [Green Version]
- Palumbo, R.; Crisci, A.; Venâncio, A.; Cortiñas Abrahantes, J.; Dorne, J.L.; Battilani, P.; Toscano, P. Occurrence and Co-Occurrence of Mycotoxins in Cereal-Based Feed and Food. Microorganisms 2020, 8, 74. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.-C.; Madec, S.; Coton, E.; Hymery, N. Natural Co-Occurrence of Mycotoxins in Foods and Feeds and Their in vitro Combined Toxicological Effects. Toxins 2016, 8, 94. [Google Scholar] [CrossRef]
- Castlebury, L.A. Use of a bioassay to evaluate the toxicity of beauvericin to bacteria. World J. Microbiol. Biotechnol. 1999, 15, 119–121. [Google Scholar] [CrossRef]
- Clemente, J.C.; Ursell, L.K.; Parfrey, L.W.; Knight, R. The impact of the gut microbiota on human health: An integrative view. Cell 2012, 148, 1258–1270. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Yao, M.; Lv, L.; Ling, Z.; Li, L. The Human Microbiota in Health and Disease. Engineering 2017, 3, 71–82. [Google Scholar] [CrossRef]
- Meca, G.; Ritieni, A.; Mañes, J. Reduction in vitro of the minor Fusarium mycotoxin beauvericin employing different strains of probiotic bacteria. Food Control 2012, 28, 435–440. [Google Scholar] [CrossRef] [Green Version]
- Caloni, F.; Stammati, A.L.; Raimondi, F.; De Angelis, I. In vitro study with Caco-2 cells on fumonisin B1: Aminopentol intestinal passage and role of P-glycoprotein. Vet. Res. Commun. 2005, 29 (Suppl. S2), 285–287. [Google Scholar] [CrossRef]
- Meca, G.; Meneghelli, G.; Ritieni, A.; Mañes, J.; Font, G. Influence of different soluble dietary fibers on the bioaccessibility of the minor Fusarium mycotoxin beauvericin. Food Chem. Toxicol. 2012, 50, 1362–1368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzini, M.; Rodriguez-Estrada, M.T.; Meca, G.; Mañes, J. Reduction of beauvericin and enniatins bioaccessibility by prebiotic compounds, evaluated in static and dynamic simulated gastrointestinal digestion. Food Control 2015, 47, 203–211. [Google Scholar] [CrossRef]
- Mallebrera, B.; Meca, G.; Manyes, L.; Mañes, J.; Font, G. Influence of pro- and prebiotics on gastric, duodenal and colonic bioaccessibility of the mycotoxin beauvericin. J. Food Compos. Anal. 2013, 32, 141–149. [Google Scholar] [CrossRef]
- Ferrer, M.; Manyes, L.; Mañes, J.; Meca, G. Influence of prebiotics, probiotics and protein ingredients on mycotoxin bioaccessibility. Food Funct. 2015, 6, 987–994. [Google Scholar] [CrossRef]
- Bertero, A.; Spicer, L.J.; Caloni, F. Fusarium mycotoxins and in vitro species-specific approach with porcine intestinal and brain in vitro barriers: A review. Food Chem. Toxicol. 2018, 121, 666–675. [Google Scholar] [CrossRef]
- Zakrzewski, S.S.; Richter, J.F.; Krug, S.M.; Jebautzke, B.; Lee, I.F.; Rieger, J.; Sachtleben, M.; Bondzio, A.; Schulzke, J.D.; Fromm, M.; et al. Improved cell line IPEC-J2, characterized as a model for porcine jejunal epithelium. PLoS ONE 2013, 8, e79643. [Google Scholar] [CrossRef] [Green Version]
- Nossol, C.; Barta-Böszörményi, A.; Kahlert, S.; Zuschratter, W.; Faber-Zuschratter, H.; Reinhardt, N.; Ponsuksili, S.; Wimmers, K.; Diesing, A.-K.; Rothkötter, H.-J. Comparing Two Intestinal Porcine Epithelial Cell Lines (IPECs): Morphological Differentiation, Function and Metabolism. PLoS ONE 2015, 10, e0132323. [Google Scholar] [CrossRef] [Green Version]
- Novak, B.; Rainer, V.; Sulyok, M.; Haltrich, D.; Schatzmayr, G.; Mayer, E. Twenty-Eight Fungal Secondary Metabolites Detected in Pig Feed Samples: Their Occurrence, Relevance and Cytotoxic Effects In Vitro. Toxins 2019, 11, 537. [Google Scholar] [CrossRef] [Green Version]
- Khoshal, A.K.; Novak, B.; Martin, P.G.P.; Jenkins, T.; Neves, M.; Schatzmayr, G.; Oswald, I.P.; Pinton, P. Co-occurrence of DON and Emerging Mycotoxins in Worldwide Finished Pig Feed and Their Combined Toxicity in Intestinal Cells. Toxins 2019, 11, 727. [Google Scholar] [CrossRef] [Green Version]
- Springler, A.; Vrubel, G.J.; Mayer, E.; Schatzmayr, G.; Novak, B. Effect of Fusarium-Derived Metabolites on the Barrier Integrity of Differentiated Intestinal Porcine Epithelial Cells (IPEC-J2). Toxins 2016, 8, 345. [Google Scholar] [CrossRef]
- Prosperini, A.; Berrada, H.; Ruiz, M.J.; Caloni, F.; Coccini, T.; Spicer, L.J.; Perego, M.C.; Lafranconi, A. A Review of the Mycotoxin Enniatin B. Front. Public Health 2017, 5, 304. [Google Scholar] [CrossRef] [PubMed]
- Pallarés, N.; Righetti, L.; Generotti, S.; Cavanna, D.; Ferrer, E.; Dall’Asta, C.; Suman, M. Investigating the in vitro catabolic fate of Enniatin B in a human gastrointestinal and colonic model. Food Chem. Toxicol. 2020, 137, 111166. [Google Scholar] [CrossRef]
- Gruber-Dorninger, C.; Novak, B.; Nagl, V.; Berthiller, F. Emerging Mycotoxins: Beyond Traditionally Determined Food Contaminants. J. Agric. Food Chem. 2017, 65, 7052–7070. [Google Scholar] [CrossRef] [PubMed]
- Gautier, C.; Pinson-Gadais, L.; Richard-Forget, F. Fusarium Mycotoxins Enniatins: An Updated Review of Their Occurrence, the Producing Fusarium Species, and the Abiotic Determinants of Their Accumulation in Crop Harvests. J. Agric. Food Chem. 2020, 68, 4788–4798. [Google Scholar] [CrossRef] [PubMed]
- Gromadzka, K.; Górna, K.; Chełkowski, J.; Waskiewicz, A. Mycotoxins and related Fusarium species in preharvest maize ear rot in Poland. Plantsoil Environ. 2016, 62, 348–354. [Google Scholar] [CrossRef] [Green Version]
- Nazari, F.; Sulyok, M.; Kobarfard, F.; Yazdanpanah, H.; Krska, R. Evaluation of Emerging Fusarium mycotoxins beauvericin, Enniatins, Fusaproliferin and Moniliformin in Domestic Rice in Iran. Iran J. Pharm. Res. 2015, 14, 505–512. [Google Scholar] [PubMed]
- Tittlemier, S.A.; Blagden, R.; Chan, J.; Gaba, D.; McKendry, T.; Pleskach, K.; Roscoe, M. Fusarium and Alternaria mycotoxins present in Canadian wheat and durum harvest samples. Can. J. Plant Pathol. 2019, 41, 403–414. [Google Scholar] [CrossRef]
- Tan, D.C.; Flematti, G.R.; Ghisalberti, E.L.; Sivasithamparam, K.; Barbetti, M.J. Toxigenicity of enniatins from Western Australian Fusarium species to brine shrimp (Artemia franciscana). Toxicon 2011, 57, 817–825. [Google Scholar] [CrossRef] [PubMed]
- Tolosa, J.; Font, G.; Mañes, J.; Ferrer, E. Mitigation of enniatins in edible fish tissues by thermal processes and identification of degradation products. Food Chem. Toxicol. 2017, 101, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, L.; Egge-Jacobsen, W.M.; Solhaug, A.; Thoen, E.; Fæste, C.K. Lysosomes as a possible target of enniatin B-induced toxicity in Caco-2 cells. Chem. Res. Toxicol. 2012, 25, 1662–1674. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, M.; Jestoi, M.; Anthoni, M.; Welling, A.; Loivamaa, I.; Hallikainen, V.; Kankainen, M.; Lysøe, E.; Koivisto, P.; Peltonen, K. Fusarium mycotoxin enniatin B: Cytotoxic effects and changes in gene expression profile. Toxicol. Vitr. 2016, 34, 309–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juan-García, A.; Manyes, L.; Ruiz, M.-J.; Font, G. Involvement of enniatins-induced cytotoxicity in human HepG2 cells. Toxicol. Lett. 2013, 218, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Prosperini, A.; Juan-Garcia, A.; Font, G.; Ruiz, M.J. Reactive oxygen species involvement in apoptosis and mitochondrial damage in Caco-2 cells induced by enniatins A, A(1), B and B(1). Toxicol. Lett. 2013, 222, 36–44. [Google Scholar] [CrossRef]
- Gammelsrud, A.; Solhaug, A.; Dendelé, B.; Sandberg, W.J.; Ivanova, L.; Kocbach Bølling, A.; Lagadic-Gossmann, D.; Refsnes, M.; Becher, R.; Eriksen, G.; et al. Enniatin B-induced cell death and inflammatory responses in RAW 267.4 murine macrophages. Toxicol. Appl. Pharmacol. 2012, 261, 74–87. [Google Scholar] [CrossRef]
- Behm, C.; Degen, G.H.; Föllmann, W. The Fusarium toxin enniatin B exerts no genotoxic activity, but pronounced cytotoxicity in vitro. Mol. Nutr. Food Res. 2009, 53, 423–430. [Google Scholar] [CrossRef]
- Krug, I.; Behrens, M.; Esselen, M.; Humpf, H.U. Transport of enniatin B and enniatin B1 across the blood-brain barrier and hints for neurotoxic effects in cerebral cells. PLoS ONE 2018, 13, e0197406. [Google Scholar] [CrossRef]
- Meca, G.; Font, G.; Ruiz, M.J. Comparative cytotoxicity study of enniatins A, A1, A2, B, B1, B4 and J3 on Caco-2 cells, Hep-G2 and HT-29. Food Chem. Toxicol. 2011, 49, 2464–2469. [Google Scholar] [CrossRef]
- Wätjen, W.; Debbab, A.; Hohlfeld, A.; Chovolou, Y.; Kampkötter, A.; Edrada, R.A.; Ebel, R.; Hakiki, A.; Mosaddak, M.; Totzke, F.; et al. Enniatins A1, B and B1 from an endophytic strain of Fusarium tricinctum induce apoptotic cell death in H4IIE hepatoma cells accompanied by inhibition of ERK phosphorylation. Mol. Nutr. Food Res. 2009, 53, 431–440. [Google Scholar] [CrossRef]
- Prosperini, A.; Font, G.; Ruiz, M.J. Interaction effects of Fusarium enniatins (A, A1, B and B1) combinations on in vitro cytotoxicity of Caco-2 cells. Toxicol. Vitr. 2014, 28, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Dornetshuber, R.; Heffeter, P.; Kamyar, M.R.; Peterbauer, T.; Berger, W.; Lemmens-Gruber, R. Enniatin exerts p53-dependent cytostatic and p53-independent cytotoxic activities against human cancer cells. Chem. Res. Toxicol. 2007, 20, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Kalayou, S.; Ndossi, D.; Frizzell, C.; Groseth, P.K.; Connolly, L.; Sørlie, M.; Verhaegen, S.; Ropstad, E. An investigation of the endocrine disrupting potential of enniatin B using in vitro bioassays. Toxicol. Lett. 2015, 233, 84–94. [Google Scholar] [CrossRef]
- Meca, G.; Sospedra, I.; Valero, M.A.; Mañes, J.; Font, G.; Ruiz, M.J. Antibacterial activity of the enniatin B, produced by Fusarium tricinctum in liquid culture, and cytotoxic effects on Caco-2 cells. Toxicol. Mech. Methods 2011, 21, 503–512. [Google Scholar] [CrossRef]
- Föllmann, W.; Behm, C.; Degen, G.H. The emerging Fusarium toxin enniatin B: In-vitro studies on its genotoxic potential and cytotoxicity in V79 cells in relation to other mycotoxins. Mycotoxin Res. 2009, 25, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Roig, M.; Meca, G.; Marín, R.; Ferrer, E.; Mañes, J. Antibacterial activity of the emerging Fusarium mycotoxins enniatins A, A1, A2, B, B1, and B4 on probiotic microorganisms. Toxicon 2014, 85, 1–4. [Google Scholar] [CrossRef]
- Sebastià, N.; Meca, G.; Soriano, J.M.; Mañes, J. Antibacterial effects of enniatins J(1) and J(3) on pathogenic and lactic acid bacteria. Food Chem. Toxicol. 2011, 49, 2710–2717. [Google Scholar] [CrossRef]
- Meca, G.; Soriano, J.M.; Gaspari, A.; Ritieni, A.; Moretti, A.; Mañes, J. Antifungal effects of the bioactive compounds enniatins A, A(1), B, B(1). Toxicon 2010, 56, 480–485. [Google Scholar] [CrossRef]
- Jeschke, P.; Harder, A.; Etzel, W.; Schindler, M.; Thielking, G. Synthesis and anthelmintic activity of cyclohexadepsipeptides with cyclohexylmethyl side chains. Bioorg. Med. Chem. Lett. 2007, 17, 3690–3695. [Google Scholar] [CrossRef]
- Burmeister, H.R.; Plattner, R.D. Enniatin production by Fusarium tricinctum and its effect on germinating wheat seeds. Phytopathology 1987, 77, 1483–1487. [Google Scholar] [CrossRef]
- Herrmann, M.; Zocher, R.; Haese, A. Enniatin production by fusarium strains and its effect on potato tuber tissue. Appl. Environ. Microbiol. 1996, 62, 393. [Google Scholar] [CrossRef] [Green Version]
- Hershenhorn, J.; Park, S.H.; Stierle, A.; Strobel, G.A. Fusarium avenaceum as a novel pathogen of spotted knapweed and its phytotoxins, acetamido-butenolide and enniatin B. Plant Sci. 1992, 86, 155–160. [Google Scholar] [CrossRef]
- Zonno, M.C.; Vurro, M. Effect of fungal toxins on germination of Striga hermonthica seeds. Weed Res. 1999, 39, 15–20. [Google Scholar] [CrossRef]
- Dornetshuber-Fleiss, R.; Heilos, D.; Mohr, T.; Richter, L.; Süssmuth, R.D.; Zlesak, M.; Novicky, A.; Heffeter, P.; Lemmens-Gruber, R.; Berger, W. The naturally born fusariotoxin enniatin B and sorafenib exert synergistic activity against cervical cancer in vitro and in vivo. Biochem Pharm. 2015, 93, 318–331. [Google Scholar] [CrossRef] [Green Version]
- Kamyar, M.; Rawnduzi, P.; Studenik, C.R.; Kouri, K.; Lemmens-Gruber, R. Investigation of the electrophysiological properties of enniatins. Arch. Biochem. Biophys. 2004, 429, 215–223. [Google Scholar] [CrossRef]
- Sy-Cordero, A.A.; Pearce, C.J.; Oberlies, N.H. Revisiting the enniatins: A review of their isolation, biosynthesis, structure determination and biological activities. J. Antibiot. 2012, 65, 541–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonshin, A.A.; Teplova, V.V.; Andersson, M.A.; Salkinoja-Salonen, M.S. The Fusarium mycotoxins enniatins and beauvericin cause mitochondrial dysfunction by affecting the mitochondrial volume regulation, oxidative phosphorylation and ion homeostasis. Toxicology 2010, 276, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Mereish, K.A.; Solow, R.; Bunner, D.L.; Fajer, A.B. Interaction of cyclic peptides and depsipeptides with calmodulin. Pept. Res. 1990, 3, 233–237. [Google Scholar]
- Ivanova, L.; Uhlig, S.; Eriksen, G.; Johannessen, L. Enniatin B 1 is a substrate of intestinal P-glycoprotein, multidrug resistance-associated protein 2 and breast cancer resistance protein. World Mycotoxin J. 2010, 3, 271–281. [Google Scholar] [CrossRef]
- Meca, G.; Mañes, J.; Font, G.; Ruiz, M.-J. Study of the potential toxicity of commercial crispy breads by evaluation of bioaccessibility and bioavailability of minor Fusarium mycotoxins. Food Chem. Toxicol. 2012, 50, 288–294. [Google Scholar] [CrossRef]
- Meca, G.; Mañes, J.; Font, G.; Ruiz, M.J. Study of the potential toxicity of enniatins A, A1, B, B1 by evaluation of duodenal and colonic bioavailability applying an in vitro method by Caco-2 cells. Toxicon 2012, 59, 1–11. [Google Scholar] [CrossRef]
- Fernández-García, E.; Carvajal-Lérida, I.; Pérez-Gálvez, A. In vitro bioaccessibility assessment as a prediction tool of nutritional efficiency. Nutr. Res. 2009, 29, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Meca, G.; Ruiz, M.J.; Soriano, J.M.; Ritieni, A.; Moretti, A.; Font, G.; Mañes, J. Isolation and purification of enniatins A, A1, B, B1, produced by Fusarium tricinctum in solid culture, and cytotoxicity effects on Caco-2 cells. Toxicon 2010, 56, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Meca, G.; Soriano, J.M.; Font, G.; Mañes, J.; Ruiz, M.J. Production, purification, and mass spectrometry characterization of the cyclohexadepsipeptide enniatin J3 and study of the cytoxicity on differentiated and undifferentiated Caco-2 cells. Toxicol. Environ. Chem. 2011, 93, 383–395. [Google Scholar] [CrossRef]
- Vejdovszky, K.; Warth, B.; Sulyok, M.; Marko, D. Non-synergistic cytotoxic effects of Fusarium and Alternaria toxin combinations in Caco-2 cells. Toxicol. Lett. 2016, 241, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Blanco, C.; Font, G.; Ruiz, M.-J. Interaction effects of enniatin B, deoxinivalenol and alternariol in Caco-2 cells. Toxicol. Lett. 2016, 241, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Meca, G.; Giri, A.; Sagratini, G.; Vittori, S.; Font, G.; Mañes, J. The soluble dietary fiber inulin can influence the bioaccessibility of enniatins. Food Funct 2012, 3, 853–858. [Google Scholar] [CrossRef]
- Prosperini, A.; Meca, G.; Font, G.; Ruiz, M.-J. Bioaccessibility of Enniatins A, A1, B, and B1 in Different Commercial Breakfast Cereals, Cookies, and Breads of Spain. J. Agric. Food Chem. 2013, 61, 456–461. [Google Scholar] [CrossRef]
- Aoudia, N.; Tangni, E.K.; Larondelle, Y. Distribution of ochratoxin A in plasma and tissues of rats fed a naturally contaminated diet amended with micronized wheat fibres: Effectiveness of mycotoxin sequestering activity. Food Chem. Toxicol. 2008, 46, 871–878. [Google Scholar] [CrossRef]
- Serrano, A.B.; Meca, G.; Font, G.; Ferrer, E. Risk assessment of beauvericin, enniatins and fusaproliferin present in follow-up infant formula by in vitro evaluation of the duodenal and colonic bioaccessibility. Food Control 2014, 42, 234–241. [Google Scholar] [CrossRef]
- Versantvoort, C.H.M.; Oomen, A.G.; Van de Kamp, E.; Rompelberg, C.J.M.; Sips, A.J.A.M. Applicability of an in vitro digestion model in assessing the bioaccessibility of mycotoxins from food. Food Chem. Toxicol. 2005, 43, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Kolf-Clauw, M.; Sassahara, M.; Lucioli, J.; Rubira-Gerez, J.; Alassane-Kpembi, I.; Lyazhri, F.; Borin, C.; Oswald, I.P. The emerging mycotoxin, enniatin B1, down-modulates the gastrointestinal toxicity of T-2 toxin in vitro on intestinal epithelial cells and ex vivo on intestinal explants. Arch. Toxicol. 2013, 87, 2233–2241. [Google Scholar] [CrossRef] [PubMed]
- Reisinger, N.; Schürer-Waldheim, S.; Mayer, E.; Debevere, S.; Antonissen, G.; Sulyok, M.; Nagl, V. Mycotoxin Occurrence in Maize Silage-A Neglected Risk for Bovine Gut Health? Toxins 2019, 11, 577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertero, A.; Moretti, A.; Spicer, L.J.; Caloni, F. Fusarium Molds and Mycotoxins: Potential Species-Specific Effects. Toxins 2018, 10, 244. [Google Scholar] [CrossRef] [Green Version]
- Fink-Gremmels, J. The role of mycotoxins in the health and performance of dairy cows. Vet. J. 2008, 176, 84–92. [Google Scholar] [CrossRef]
- Gallo, A.; Giuberti, G.; Frisvad, J.C.; Bertuzzi, T.; Nielsen, K.F. Review on Mycotoxin Issues in Ruminants: Occurrence in Forages, Effects of Mycotoxin Ingestion on Health Status and Animal Performance and Practical Strategies to Counteract Their Negative Effects. Toxins 2015, 7, 3057–3111. [Google Scholar] [CrossRef] [PubMed]
- Upadhaya, S.D.; Park, M.A.; Ha, J.K. Mycotoxins and Their Biotransformation in the Rumen: A Review. Asian-Australas J. Anim. Sci. 2010, 23, 1250–1260. [Google Scholar] [CrossRef]
- Debevere, S.; Cools, A.; Baere, S.; Haesaert, G.; Rychlik, M.; Croubels, S.; Fievez, V. In Vitro Rumen Simulations Show a Reduced Disappearance of Deoxynivalenol, Nivalenol and Enniatin B at Conditions of Rumen Acidosis and Lower Microbial Activity. Toxins 2020, 12, 101. [Google Scholar] [CrossRef] [Green Version]
- Debevere, S.; Schatzmayr, D.; Reisinger, N.; Aleschko, M.; Haesaert, G.; Rychlik, M.; Croubels, S.; Fievez, V. Evaluation of the Efficacy of Mycotoxin Modifiers and Mycotoxin Binders by Using an In Vitro Rumen Model as a First Screening Tool. Toxins 2020, 12, 405. [Google Scholar] [CrossRef]
- Moreira, G.M.; Nicolli, C.P.; Gomes, L.B.; Ogoshi, C.; Scheuermann, K.K.; Silva-Lobo, V.L.; Schurt, D.A.; Ritieni, A.; Moretti, A.; Pfenning, L.H.; et al. Nationwide survey reveals high diversity of Fusarium species and related mycotoxins in Brazilian rice: 2014 and 2015 harvests. Food Control 2020, 113, 107171. [Google Scholar] [CrossRef]
- Stanciu, O.; Juan, C.; Miere, D.; Dumitrescu, A.; Bodoki, E.; Loghin, F.; Mañes, J. Climatic conditions influence emerging mycotoxin presence in wheat grown in Romania–A 2-year survey. Crop Prot. 2017, 100, 124–133. [Google Scholar] [CrossRef]
- Decleer, M.; Landschoot, S.; De Saeger, S.; Rajkovic, A.; Audenaert, K. Impact of fungicides and weather on cyclodepsipeptide-producing Fusarium spp. and beauvericin and enniatin levels in wheat grains. J. Sci. Food Agric. 2019, 99, 253–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nácher-Mestre, J.; Beltrán, E.; Strachan, F.; Dick, J.R.; Pérez-Sánchez, J.; Berntssen, M.H.G.; Tocher, D.R. No transfer of the non-regulated mycotoxins, beauvericin and enniatins, from feeds to farmed fish reared on plant-based diets. Food Chem. 2020, 323, 126773. [Google Scholar] [CrossRef] [PubMed]
- Kolawole, O.; Graham, A.; Donaldson, C.; Owens, B.; Abia, W.A.; Meneely, J.; Alcorn, M.J.; Connolly, L.; Elliott, C.T. Low Doses of Mycotoxin Mixtures below EU Regulatory Limits Can Negatively Affect the Performance of Broiler Chickens: A Longitudinal Study. Toxins 2020, 12, 433. [Google Scholar] [CrossRef]
- Braun, D.; Ezekiel, C.N.; Marko, D.; Warth, B. Exposure to Mycotoxin-Mixtures via Breast Milk: An Ultra-Sensitive LC-MS/MS Biomonitoring Approach. Front Chem. 2020, 8, 423. [Google Scholar] [CrossRef]
- Polišenská, I.; Jirsa, O.; Vaculová, K.; Pospíchalová, M.; Wawroszova, S.; Frydrych, J. Fusarium Mycotoxins in Two Hulless Oat and Barley Cultivars Used for Food Purposes. Foods 2020, 9, 1037. [Google Scholar] [CrossRef]
- Bryła, M.; Waśkiewicz, A.; Podolska, G.; Szymczyk, K.; Jędrzejczak, R.; Damaziak, K.; Sułek, A. Occurrence of 26 Mycotoxins in the Grain of Cereals Cultivated in Poland. Toxins 2016, 8, 160. [Google Scholar] [CrossRef] [Green Version]
- Panasiuk, L.; Jedziniak, P.; Pietruszka, K.; Piatkowska, M.; Bocian, L. Frequency and levels of regulated and emerging mycotoxins in silage in Poland. Mycotoxin Res. 2019, 35, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Arroyo-Manzanares, N.; Rodríguez-Estévez, V.; Arenas-Fernández, P.; García-Campaña, A.M.; Gámiz-Gracia, L. Occurrence of Mycotoxins in Swine Feeding from Spain. Toxins 2019, 11, 342. [Google Scholar] [CrossRef] [Green Version]
- Juan, C.; Oueslati, S.; Mañes, J.; Berrada, H. Multimycotoxin Determination in Tunisian Farm Animal Feed. J. Food Sci. 2019, 84, 3885–3893. [Google Scholar] [CrossRef]
- Braun, D.; Schernhammer, E.; Marko, D.; Warth, B. Longitudinal assessment of mycotoxin co-exposures in exclusively breastfed infants. Environ. Int. 2020, 142, 105845. [Google Scholar] [CrossRef] [PubMed]
- EEC. Council Regulation (EEC) No 315/93 of 8 February 1993 laying down Community procedures for contaminants in food. Off. J. Eur. Communities 1993, 37, 1–3. [Google Scholar]
- EC. Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Communities 2006, 364, 5–24. [Google Scholar]
- EC. Commission Regulation (EU) No 165/2010 of 26 February 2010 amending Regulation (EC) No 1881/2006 setting maximum levels for certain contaminants in foodstuffs as regards aflatoxins. Off. J. Eur. Union 2010, 50, 8–12. [Google Scholar]
- Siegel, D.; Babuscio, T. Mycotoxin management in the European cereal trading sector. Food Control 2011, 22, 1145–1153. [Google Scholar] [CrossRef]
- Wu, F. A tale of two commodities: How EU mycotoxin regulations have affected u.s. tree nut industries. World Mycotoxin J. 2008, 1, 95–102. [Google Scholar] [CrossRef]
- EC. Commission Regulation (EC) No 401/2006 of 23 February 2006 laying down the methods of sampling and analysis for the official control of the levels of mycotoxins in foodstuffs. Off. J. Eur. Union 2006, 70, 12–34. [Google Scholar]
- EC. Commission Regulation (EC) No 152/2009 of 27 January 2009 laying down the methods of sampling and analysis for the official control of feed. Off. J. Eur. Union 2009, 54, 1–130. [Google Scholar]


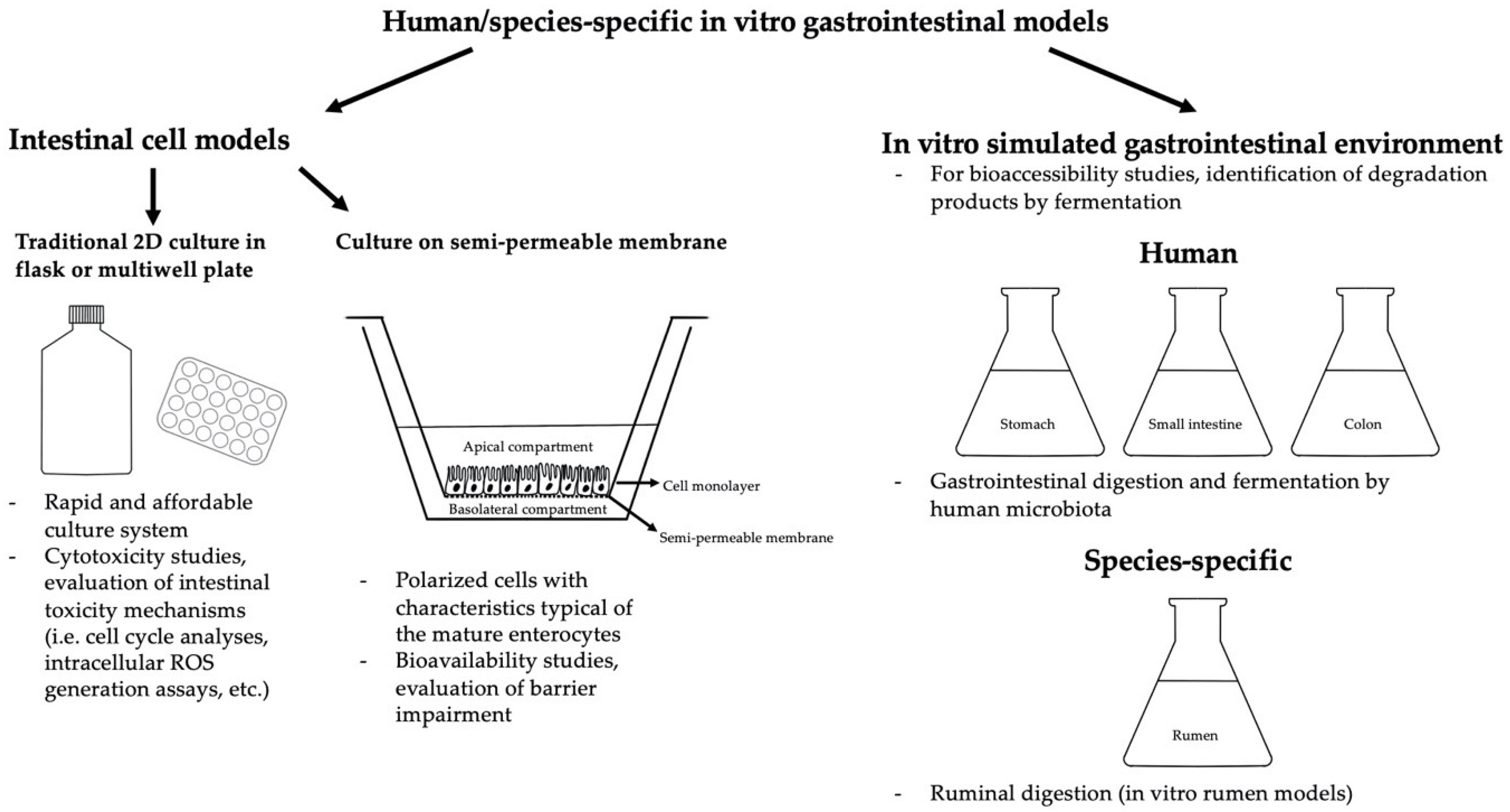{kind=link}
| Models/Cells | Concentration and Exposure Time | Method/Assay | Effects | Ref. |
|---|---|---|---|---|
| ACD/Percepta (in silico model) | BEA | In silico toxicity analysis |
| [78] |
| Caco-2 cells | 0–10 μM BEA; 24 and 48 h | HCA multiparameter assay | Cytotoxicity ((BEA) ≥ 1 μM) | [37] |
| Caco-2 cells | 0–100 μM BEA; 48 h | Resazurin-based in vitro toxicity assay | Cytotoxicity IC50 = 3.9 ± 0.7 μM) | [81] |
| Caco-2 cells | 0–30 μM BEA; 24 and 48 h | MTT assay | Cytotoxicity (IC50 at 24 h = 20.62 ± 6.9 μM; IC50 at 48 h = 12.75 ± 4.8 μM) | [79] |
| Caco-2 cells | 0–50 nM BEA; 72 h | Resazurin-based in vitro toxicity assay | No cytotoxicity | [78] |
| Caco-2 cells | BEA; 24, 48 and 72 h | MTT assay | Cytotoxicity | [80] |
| Caco-2 cells | 1–20 μM BEA; 24–48–72 h | MTT assay | Cytotoxicity: IC50 = 4.87 ± 0.42 (24 h), 4.07 ± 0.38 (48 h), 3.16 ± 0.45 (72 h) μM | [82] |
| Caco-2 cells | 3.125–25 μM BEA; 24–48–72 h | MTT and NR assays | MTT assay: IC50 = 3.2 (72 h)–20.6 (24 h) μM NR assay: IC50 = 1.9 (72 h)–8.8 (24 h) μM | [36] |
| Caco-2 cells | 1.5 and 3.0 μM BEA; 4 h | LC analysis | Bioavailability: 54.3% (transport profile with 1.5 μM BEA) and 50.1% (3 μM BEA) | [79] |
| Coculture Caco-2 and RAW 264.7 cells | Noncytotoxic doses of BEA + silibinin; 4 h | U-HPLC-MS | Transepithelial transport of BEA: Ap = 10.40 ± 3.89% of the initial concentration; Caco-2 cells = 10.36 ± 0.98%; Bl = 2.31 ± 0.24%; Raw 264.7 cells = 0.57 ± 0.03 | [78] |
| Caco-2 cells | 1.5 μM BEA; 24 h | TEER evaluation | No barrier impairment | [83] |
| Caco-2 cells | 1.5 μM BEA + 1.5 μM AFB1; 24 h | TEER evaluation | TEER decrease (at 1 and 2 h of exposure) | [83] |
| Caco-2 cells | 1.5 μM BEA; 24 h | IL-8 determination | No effect on IL-8 release | [83] |
| Caco-2 cells | 1.5 μM BEA + 3.5 μM DON; 24 h | IL-8 determination | Increased IL-8 production | [83] |
| Caco-2 cells | 1.5 and 3.0 μM BEA; 0–120 min | DCFH-DA assay | Increased ROS production (up to 2-fold higher than control) | [36] |
| Caco-2 cells | 1.5 and 3.0 μM BEA; 24 h | Lipid peroxidation assay | Increased MDA production (120% with 1.5 μM BEA and 207% with 3.0 μM BEA) | [36] |
| Caco-2 cells | 1.5 and 3.0 μM BEA; 24 h | Reduced and oxidized glutathione (GSH, GSSG) determination (fluorometric method) |
| [36] |
| Caco-2 cells | 1.5 and 3.0 μM BEA; 24, 48 and 72 h | Tetramethylrhodamine methyl ester (TMRM) method + flow cytometry | Loss of mitochondrial membrane potential (from 2% to 95% with 1.5 μM BEA and from 10% to 80% with 3.0 μM BEA) | [36] |
| Caco-2 cells | 1.5, 3.0 and 12.0 μM BEA; 24 h | Alkaline comet assay | No increase in the TM parameter with 1.5 and 3 μM BEA; significant (p ≤ 0.000) increase in the TM with 12.0 μM BEA | [36] |
| Caco-2 cells | 1.5 and 3.0 μM BEA; 24, 48 and 72 h | Cell cycle analysis via propidium iodide (PI) staining |
| [36] |
| Caco-2 cells | 5 μM BEA + 2.5–5 × 105 CFU/mL L. acidophilus; 12–24–48–72 h | MTT assay | At 12 and 24 h cell viability in the presence of L. acidophilus and BEA is higher (p < 0.05) than with BEA alone | [82] |
| Probiotic bacterial strains typical of mammalian intestinal tract | 5 mg/L BEA + 108 CFU/mL; 4–48 h | LC-MS/MS |
| [92] |
| Panel of bacterial strains typical of mammalian intestinal tract | 0.1–25 μg BEA | Agar diffusion assay | Inhibition of B. pumilus, B. cereus, B. mycoides, B. sphaericus, P. alvei, P. azotofixans, P. macquariensis, P. pulvifaciens and P. validus. BEA also inhibited anaerobes: E. biforme, P. anaerobius, P. productus, B. adolescentis and C. perfringens | [89] |
| Panel of Gram-positive and Gram-negative bacteria | 0–100 μM BEA; 18–24 h | Antimicrobial activity assay | Antibacterial activity vs. Gram-positive bacteria ((BEA) > 6 and ≤ 12.5 μM) and mycobacteria ((BEA) = 25 μM) | [81] |
| HT-29 cells | 0–30 μM BEA; 24 and 48 h | MTT assay | Cytotoxicity (IC50 at 24 h = 15.00 ± 6.9 μM; IC50 at 48 h = 9.75 ± 4.4 μM) | [79] |
| N87 (human gastric cell line) | 0–100 μM; 48 h | Resazurin-based in vitro toxicity assay | Cytotoxicity IC50 = 27.5 ± 0.7 μM) | [81] |
| Models/Cells | Concentration and Exposure Time | Method/Assay | Effects | Ref. |
|---|---|---|---|---|
| Simulated gastrointestinal environment | 5 and 25 mg/kg BEA in the model solution and in wheat crispy bread with different natural binding compounds (dietary fibers, 1–5% w/w) | LC-MS | Bioaccessibility in the model solution: 31.8% (duodenal digestion)–54.0% (samples that underwent also colonic fermentation) Bioaccessibility in wheat crispy bread: 1.9% (duodenal digestion)–27.0% (samples that underwent also colonic fermentation) | [94] |
| Pro- and prebiotics in a simulated gastrointestinal environment | Model solution: 10 mg/L BEA + 2 × 106 CFU and 10 mg/L BEA + 1–5 g dietary fibers | LC-MS |
| [96] |
| Pro- and prebiotics in dynamic simulated gastrointestinal environment | BEA in 20 g of wheat crispy bread produced with 300 g of wheat flour added with 106 conidia/mL of F. tricinctum, F. culmorum, G. zeae and fermented for 30 days + 1–5% w/w prebiotics or 1 × 105 UFC/mL probiotics in the simulated saliva | LC-MS/MS | Probiotics (highest reduction 10.1 and 6.6%, gastric and duodenal bioaccessibility, respectively, ctr 15.8 and 11.3%) and prebiotics (highest reduction 15.3 and 12.0%, gastric and duodenal bioaccessibility, respectively, ctr 28.4 and 19.6%) caused a reduction of BEA bioaccessibility | [97] |
| Static and dynamic simulated gastrointestinal environment | BEA in 10 g (static model) or 100 g (dynamic model) of wheat crispy bread produced with 300 g of durum wheat added with 106 conidia/mL of F. tricinctum and fermented for 30 days, mixed with 1–5% w/w inulin and fructooligosaccharides (FOS) | LC-MS/MS |
| [95] |
| Models/Cells | Concentration and Exposure Time | Method/Assay | Effects | Ref. |
|---|---|---|---|---|
| Proliferating IPEC-J2 cells | 0–10 μM BEA; 24 h | Cytotoxicity assay (flow cytometry + Annexin-V-FITC and PI staining) | 5 μM BEA: 82% viable, 15% early apoptotic, 3% apoptotic/necrotic cells; 10 μM BEA complete disruption | [50] |
| Differentiated IPEC-J2 cells | 0–10 μM BEA; 24 h | Cytotoxicity assay (flow cytometry + Annexin-V-FITC and PI staining) | 5 μM BEA no effect; 10 μM BEA: 47% viable, 27% early apoptotic, 27% apoptotic/necrotic cells | [50] |
| IPEC-J2 cells | 1.5–10 μM BEA; 24, 48 and 72 h | TEER evaluation | 5 μM BEA significantly (p < 0.05) reduced TEER starting from 24 h of exposure; 80% reduction after 72 h of exposure to 10 μM BEA | [103] |
| IPEC-J2 cells | 1.5–10 μM BEA; 24, 48 and 72 h | NR assay | No cytotoxicity | [103] |
| IPEC-J2 cells | 2.5 μM BEA + 1.5 or 3 μM DON; 24, 48 and 72 h | TEER evaluation | No effect on barrier integrity | [103] |
| IPEC-J2 cells | 2.5 μM BEA + 1.5 or 3 μM DON; 24, 48 and 72 h | NR assay | No cytotoxicity | [103] |
| IPEC-J2 cells | 0–20 μM BEA; 48 h | Sulforhodamine B (SRB) assay | Cytotoxicity: absolute IC50 = 2.43 μM; relative IC50 = 2.24 μM | [101] |
| IPEC-1 cells | BEA; 48 h | CellTiter-Glo® Luminescent Cell Viability Assay | Cytotoxicity: IC50 = 4.3 ± 1.8 μM (classified as highly toxic) | [102] |
| Models/Cells | Concentration and Exposure Time | Method/Assay | Effects | Ref. |
|---|---|---|---|---|
| Caco-2 cells | 0–100 μM ENN; 48 h | Resazurin-based in vitro toxicity assay | Cytotoxicity IC50 range = (1.1 ± 0.2 μM (ENN A))–(4.6 ± 1.3 μM (ENN B)) | [81] |
| Caco-2 cells | 0.1–10 μM ENN; 72 h | MTT assay | IC75 = 1.38 ± 0.07 μM; IC50 = 1.99 ± 0.09 μM; IC25 = 2.63 ± 0.21 μM | [123] |
| Caco-2 cells | ENN; 24, 48 and 72 h | MTT assay | Cytotoxicity (3- to 4-fold higher than BEA) | [80] |
| Caco-2 cells | 1–25 μM ENN B; 24 h | Cell counting | Viability: 85 ± 7% after exposure to 1 μM ENN B; 50 ± 5% after exposure to 25 μM ENN B | [113] |
| Caco-2 cells (undifferentiated and differentiated) | 0.6–30 μM ENN B; 24 and 48 h | MTT assay |
| [125] |
| Caco-2 cells | Up to 437.2 μM ENN B; max 48 h | Combined bioassays for cytotoxicity (AB –metabolic activity, LDH—cell membrane integrity and NR—lysosomal activity |
| [113] |
| Caco-2 cells (undifferentiated and differentiated) | ENN J3 | Cytotoxicity assay | No cytotoxic effects at any of the concentrations tested | [145] |
| Caco-2 cells | 0.6–30 μM ENN B, B1, A, A1; 24 h | MTT assay |
| [144] |
| Caco-2 cells | 0–30 μM ENN A, A1, B, B1; 24 h | MTT assay | IC50 A1 = 12.3 μM; IC50 B1 = 19.5 μM. No IC50 were obtained for ENN A and B at the concentrations tested | [141] |
| Caco-2 cells | 0.6-30 μM ENN A, A1, A2, B, B1, B4 and J3; 24 and 48 h | MTT assay | At 48 h (μM): IC50 A = 9.3 ± 0.6; IC50 A1 = 2.7 ± 0.8; IC50 A2 = 2.6 ± 0.7; IC50 B = no values obtained in the range of concentrations tested; IC50 B1 = 11.5 ± 5.3; IC50 B4 = 4.5 ± 2.9 and IC50 J3= no values obtained in the range of concentrations tested | [120] |
| Caco-2 cells | 0–50 nM ENNs (A, A1, B, B1); 72 h | Resazurin-based in vitro toxicity assay | No cytotoxicity | [78] |
| Caco-2 cells | 0.9–15 μM ENN A1, B, B1 and 0.45–7.5 μM ENN A; 24, 48, 72 h | MTT and NR assay | MTT assay, at 72 h (μM): IC50 A = 1.6 ± 0.8; IC50 A1 = 1.3 ± 0.6; IC50 B = 11.7 ± 2.4; IC50 B1 = 2.8 ± 1.1. NR assay, at 72 h (μM): IC50 A = 0.46 ± 0.1; IC50 A1 = 0.46 ± 0.1; IC50 B = 1.4 ± 0.2; IC50 B1 = 0.8 ± 0.3 | [116] |
| Caco-2 cells | 0.9–15.0 μM ENN A, A1, B, B1 alone and combined; 24 h | MTT assay |
| [122] |
| Caco-2 cells | 0.312–10 μM ENN B (alone), 0.312–5 μM in coexposures; 24, 48 and 72 h | MTT assay |
| [147] |
| Caco-2 cells | Binary mixture of ENN B + other Fusarium and Alternaria mycotoxins (0–250 μM); 24 h | Cell proliferation assay WST-1 |
| [146] |
| Caco-2 cells | 1–25 μM ENN B; 24 h | Double staining with PI/Hoechst 33342 and fluorescence microscopy |
| [113] |
| Caco-2 cells | 1.5 and 3 μM ENN A, A1, B and B1; 24, 48 and 72 h | Flow cytometry analysis of apoptosis and necrosis (V-FITC/PI double staining) | Apoptotic effects observed after 24 and 48 h of exposure; necrotic effects observed for ENN A and A1 after 24 h, and for all mycotoxins at 72 h | [116] |
| Caco-2 cells | ENN B1 | Absorption profile |
| [140] |
| Caco-2 cells | 1.5 and 3.0 μM (ENN A, A1, B and B1); 1–4 h | LC-DAD and LC-MS |
| [142] |
| Caco-2 cells | ENNs in the duodenal fluid from the simulated gastrointestinal digestion of 3 g of wheat crispy bread spiked with ENN A, A1, B and B1 at 1.5 and 3.0 μmol/g (1.5 and 3.0 μM) | LC-DAD |
| [141] |
| Caco-2 cells grown with bacterial strains (107–108 CFU/mL) typical of the intestinal tract | 1.5 and 3.0 μM (ENN A, A1, B and B1); 48 h | LC-MS |
| [142] |
| Caco-2 cells | 1–25 μM ENN B; 24 h | PI staining and flow cytometry | Increase of the cells in the G2/M phase (25 μM ENN B for 24 h = 31 ± 1.3% cells in the G2/M phase; control = 23 ± 1.0%) | [113] |
| Caco-2 cells | 1–25 μM ENN B; 24 h | Red fluorescent LysoTracker Red DND-99 staining |
| [113] |
| Caco-2 cells | 5 and 10 μM ENN B; 3 and 24 h | Lipophilic cationic probe + flow cytometry | Dose-dependent decrease in the FL2/FL1 ratio after exposure to 5 μM and 10 μM ENN B for 24 h (indicator of change in the membrane potential and MOMP) | [113] |
| Caco-2 cells | 1–25 μM ENN B; 3 and 24 h | Dihydroethidium (DHE) (oxidation- sensitive fluorescent probe for ROS detection) | Increased ROS production already with 5 μM ENN B for 3 h; 2.4-fold increase after 24 h | [113] |
| Caco-2 cells | 1.5 and 3 μM ENN A, A1, B and B1; up to 120 min | ROS generation assay (H2-DCFDA probe) |
| [116] |
| Caco-2 cells | 1.5 and 3 μM ENN A, A1, B and B1; 24 h | Lipid peroxidation assay (TBARS method) | Increased MDA production: at 3 μM, increase of 111% (ENN A), 58% (ENNA1), 48% (ENN B), 59% (ENN B1) | [116] |
| Caco-2 cells | 1.5 and 3 μM ENN A, A1, B and B1; 24, 48 and 72 h | Cell cycle analysis (PI staining) by flow cytometry | Alteration (time- and concentration-dependent) of the % of cells in SubG0/G1, G0/G1 and G2/M phases for all ENNs tested | [116] |
| n | 1.5 and 3 μM ENN A, A1, B and B1; 24, 48 and 72 h | Detection of mitochondrial membrane potential by tetramethyl rhodamine methyl ester (TMRM) method | Dose- and time-dependent decrease in TMRM fluorescence intensity. At 72 h reduction, % of TMRM intensity ranged from 91.0% (ENN B) to 98.7% (ENN A) | [116] |
| Caco-2 cells | 1.5 and 3 μM ENN A, A1, B and B1; 24 h | Alkaline comet assay | ENN A (1.5 and 3.0 μM) and 3.0 μM ENN A1 and B1 induced a significant increase in the TM parameter; 1.5 μM ENN A1 and ENN B did not cause any DNA damage | [116] |
| Coculture Caco-2 and RAW 264.7 cells | Noncytotoxic doses of ENNs (A, A1, B, B1) + silibinin; 4 h | U-HPLC-MS | Transepithelial transport of ENNs (ranges): Ap = (9.52 ± 3.02% (ENN A1))–(24.08 ± 3.20% (ENN B)) of the initial concentration; Caco-2 cells = (0.51 ± 0.03% (ENN B1))–(10.76 ± 0.43% (ENN A)); Bl = (13.06 ± 0.45% (ENN A1))–(28.16 ± 3.37% (ENN B1)); Raw 264.7 cells = (0.22 ± 0.03% (ENN A1))–(0.60 ± 0.10% (ENN B1)) | [78] |
| Panel of Gram-positive and Gram-negative bacteria | 0–100 μM ENNs; 18–24 h | Antimicrobial activity assay | Antibacterial activity vs. Gram-positive bacteria (range: 3.12 μM (ENN A)–>100 μM (ENN B)) and mycobacteria (range: 3.12 μM (ENN A1)–100 μM (ENN B)) | [81] |
| Bacterial strains normal pathogens of the intestinal tract | 0.2–2000 μg ENN B; 48 h | Disk diffusion assay |
| [125] |
| Probiotic bacteria, Saccharomyces cerevisiae strains and Bacillus subtilis strains | 0.2–20,000 ng ENN A, A1, A2, B, B1 and B4; 24 h | Disk diffusion assay |
| [127] |
| HCT116 cells | 0–10 μM ENN; 24 h | 3H-thymidine incorporation | p53-dependent cytostatic and p53-independent cytotoxic activities | [123] |
| HT29 | 0.6–30 μM ENN A, A1, A2, B, B1, B4 and J3; 24 and 48 h | MTT assay | At 48 h (μM): IC50 A = 8.2 ± 1.8; IC50 A1 = 1.4 ± 0.7; IC50 A2 = no values obtained in the range of concentrations tested; IC50 B = 2.8 ± 0.9; IC50 B1 = 3.7 ± 0.7; IC50 B4 = 15.0 ± 4.0 and IC50 J3 = no values obtained in the range of concentrations tested | [120] |
| N87 (human gastric cell line) | 0–100 μM ENN; 48 h | Resazurin-based in vitro toxicity assay | Cytotoxicity IC50 range = (0.003 ± 0.002 μM (ENN A1))–(1.7 ± 0.1 μM (ENN B)) | [81] |
| ACD/Percepta (in silico model) | ENNs (A, A1, B, B1) | In silico toxicity analysis |
| [78] |
| Models/Cells | Concentration and Exposure Time | Method/Assay | Effects | Ref. |
|---|---|---|---|---|
| Simulated gastrointestinal environment | ENN A, A1, B, B1 spiked (1.5 and 3.0 μmol/g) in wheat crispy bread (sample: 3 g) | LC-DAD | Duodenal bioaccessibility (1.5 and 3.0 μmol/g) ENN A = 84.6 ± 2.5% and 87.3 ± 2.9%; ENN A1 = 72.6 ± 1.8% and 70.0 ± 1.7%; ENN B = 68.6 ± 2.9% and 73.3 ± 1.5%; ENN B1 = 74.0 ± 1.6; 74.0 ± 1.9% | [141] |
| Simulated gastrointestinal environment | ENN A, A1, B, B1 added (1.5 and 3.0 μmol/g) to breakfast cereals, cookies and breads (sample: 3 g) | LC-DAD | Lowest bioaccessibility values found in wheat bran with fibers (1.5 μM ENN A = 50.1 ± 3.1%; 1.5 μM ENN A1 = 40.4 ± 1.9%; 1.5 μM ENN B = 43.9 ± 3.4%; 1.5 μM ENN B1 = 46.3 ± 3.1%); highest values in white loaf bread (1.5 μM ENN A = 79.9 ± 2.8%; 1.5 μM ENN A1 = 64.2 ± 2.4%; 1.5 μM ENN B = 69.8 ± 2.9%; 1.5 μM ENN B1 = 73.6 ± 2.2%) | [149] |
| Simulated gastrointestinal environment | ENN A, A1, B, B1 spiked (1.5 and 3.0 μmol/g) in 3 g of wheat crispy bread with 0, 1, 5 or 10% inulin | LC-DAD | Bioaccessibility of ENNs in crispy bread without inulin: 69% (1.5 μM ENN B) and 87% (3.0 μM ENN A); bioaccessibility with 1% inulin: 65% (3 μM ENN A)–83% (1.5 μM ENN A); bioaccessibility with 5% inulin: 58% (3.0 μM ENN A1)–74% (1.5 μM ENN B1); bioaccessibility with 10% inulin 51% (3 μM ENN A)–74% (1.5 μM ENN B1) | [148] |
| Simulated gastrointestinal environment | ENNs (A, A1, B and B1) in follow-up infant formula | LC-DAD | Colonic + duodenal bioaccessibility, range: ENN A = 1.63 ± 0.01% (just one sample found positive); ENN A1 = (4.36 ± 0.61%)–(60.53 ± 2.64%); ENN B1 = (1.49 ± 0.04%)–(8.43 ± 0.64%). No samples found positive for ENN B | [151] |
| Static and dynamic simulated gastrointestinal environment | ENNs in 10 g (static model) or 100 g (dynamic model) of wheat crispy bread produced with 300 g of durum wheat added with 106 conidia/mL of F. tricinctum and fermented for 30 days, mixed with 1–5% w/w inulin and fructooligosaccharides (FOS) | LC-MS/MS |
| [95] |
| Pro- and prebiotics in dynamic simulated gastrointestinal environment | ENNs (A, A1, B and B1) in 20 g of wheat crispy bread produced with 300 g of wheat flour added with 106 conidia/mL of F. tricinctum, F. culmorum, G. zeae and fermented for 30 days + 1–5% w/w prebiotics or 1 × 105 UFC/mL probiotics in the simulated saliva | LC-MS/MS | Probiotics (highest reduction 27.8 and 21.2%, gastric and duodenal bioaccessibility, respectively, ctr 39.6 and 33.4%) and prebiotics/food ingredients (highest reduction 20.3 and 17.0%, gastric and duodenal bioaccessibility, respectively, ctr 39.6 and 33.4%) caused a reduction of ENNs bioaccessibility | [97] |
| Simulated gastrointestinal environment | 500 μg/L ENN B (final concentration in the digestion solution) | Targeted and untargeted UHPLC-MS/MS |
| [105] |
| Models/Cells | Concentration and Exposure Time | Method/Assay | Effects | Ref. |
|---|---|---|---|---|
| IPEC-1 cells | 0–100 μM ENN alone and in coexposure with 0–100 nM T2; 48 h | CellTiter- Glo® Luminescent Cell Viability Assay | IC50 ENN = 15.80 μM; IC50 T2 = 9.35 nM; IC50 T2 + ENN (1:1000) = 14.41 μM | [153] |
| IPEC-1 cells | ENN A1, B, B1; 48 h | CellTiter-Glo® Luminescent Cell Viability Assay | IC50 ENN A1 = 1.6 ± 0.3 μM (classified as highly toxic); IC50 ENN B = 4.4 ± 0.9 μM (highly toxic); IC50 ENN B1 = 13.5 ± 2.5 μM (moderately toxic) | [102] |
| Proliferating IPEC-J2 cells | 0–100 μM ENN A, A1, B, B1; 24 h | Cytotoxicity assay (flow cytometry + Annexin-V-FITC and PI staining) | 5 μM ENNs: no effects; ENN A most cytotoxic (10 μM exposure caused a reduction of viable cells to 30%); 10 μM ENN A1 and ENN B1: 86% and 93% viable cells, respectively while 25 μM ENN A1 and ENN B1: complete disruption and 25% viable cells, respectively; 25 μM ENN B: 92% viable cells 5 μM ENN A and ENN A1: 4.1% and 3.5% early apoptosis and 2.7% and 2.4% late apoptosis/necrosis, respectively | [50] |
| Differentiated IPEC-J2 cells | 0–100 μM ENN A, A1, B, B1; 24 h | Cytotoxicity assay (flow cytometry + Annexin-V-FITC and PI staining) | 5 μM ENNs no effect; ENN A most cytotoxic (10 μM exposure caused a reduction of viable cells to 36%); 25 μM ENN A1 and ENN B1: complete disruption and 36% viable cells, respectively; 25 μM ENN B: 89% viable cells | [50] |
| IPEC-J2 cells | 0–20 μM ENN A, A1, B, B1; 48 h | Sulforhodamine B (SRB) assay | Cytotoxicity: IC50 ENN A = 3.40 μM; IC50 ENN A1 = 4.15 μM; IC50 ENN B = 3.25 μM; IC50 ENN B1 = 3.67 μM | [101] |
| IPEC-J2 cells | 0–10 μM ENNs (A, A1, B and B1); 24, 48 and 72 h | TEER evaluation | ENN A: TEER reduction after 72 h exposure to 5 μM; ENN A1: TEER reduction starting from 24 h exposure to 10 μM; ENN B: TEER reduction after 48 h and 72 h of exposure to 2.5 and 5 μM; ENN B1: TEER reduction starting from 48 h of exposure to 5 μM; | [103] |
| IPEC-J2 cells | 1.5–3 μM ENNs (A, A1, B and B1) in absence or presence of 1.5-3 μM DON; 24, 48 and 72 h | TEER evaluation | ENN A + A1 + B + B1: TEER reduction starting from 24 h exposure to 1.5 μM; ENN A + A1 + B + B1 + DON: TEER reduction starting from 24 h exposure to 1.5 μM | [103] |
| IPEC-J2 cells | 0–10 μM ENNs (A, A1, B and B1); 72 h | NR assay | No cytotoxicity | [103] |
| IPEC-J2 cells | 1.5–3 μM ENNs (A, A1, B and B1) in absence or presence of 1.5–3 μM DON; 72 h | NR assay | No cytotoxicity | [103] |
| Calf small intestinal epithelial cells B | 0–200 μM ENN B; 48 h | NR assay and WST-1 assay | IC50 ENN B = 4.0–6.7 μM | [154] |
| Models/Cells | Concentration and Exposure Time | Method/Assay | Effects | Ref. |
|---|---|---|---|---|
| In vitro rumen model | 50 mg of maize silage spiked with 1 mg/Kg ENN B; up to 48 h | UPLC system coupled to Xevo® TQ-S MS/MS system | ENN B degradation up to 72% (48 h of incubation) | [159] |
| In vitro rumen model | 50 mg of maize silage spiked with 1 mg/Kg ENN B + 3 g/kg Binder 1, 2 and 3; up to 48 h | UPLC system coupled to Xevo® TQ-S MS/MS system | Binder 1 (clay minerals and yeast derivatives): adsorbed ENN B by 24%; Binder 2 (bentonite, leonardite, plant extracts, epoxidase): adsorbed ENN B by 28%; Binder 3 (bentonite and sepiolite): adsorbed ENN B by 22% | [160] |
| Source | Tested Samples | BEA | ENN A | ENN A1 | ENN B | ENN B1 | Country | References |
|---|---|---|---|---|---|---|---|---|
| Rice Flour | 93 | 810.1 | <LOQ | <LOQ | <LOQ | 2.4 | Brazil | [161] |
| Rice Husk | 93 | 110.4 | <LOQ | <LOQ | 2.6 | 1.2 | Brazil | [161] |
| Wheat | 97 | 9.1 | 139.8 | 356.0 | 814.6 | 510.0 | Romania | [162] |
| Wheat | 140 | 13.5 | 15.6 | 165.0 | 2168 | 776.7 | Belgium | [163] |
| Barley | 10 | ND | ND | ND | 1.3 | ND | Spain | [10] |
| Rice bran | 4 | 64.8 | ND | ND | ND | ND | Spain | [10] |
| Corn pulp | 4 | 37.8 | ND | ND | 2.2 | ND | Spain | [10] |
| Barley hulless | 12 | 423 | 13 | 87 | 592 | 281 | Czech Republic | [167] |
| Oats hulless | 12 | 35 | ND | ND | 55 | 15 | Czech Republic | [167] |
| Barley | 56 | 130 | 39 | 140 | 2100 | 520 | Denmark | [27] |
| Oat | 11 | 110 | <10 | 39 | 470 | 120 | Denmark | [27] |
| Wheat | 33 | <15 | <10 | 60 | 1600 | 290 | Denmark | [27] |
| Rye | 10 | <15 | 100 | 100 | 3900 | 860 | Denmark | [27] |
| Spring barley | 8 | ND | 3 | 45 | 301 | 240 | Poland | [168] |
| Winter barley | 16 | ND | ND | 16 | 253 | 81 | Poland | [168] |
| Oats | 4 | ND | ND | 11 | 162 | 67 | Poland | [168] |
| Triticale | 20 | ND | 135 | 882 | 3328 | 1347 | Poland | [168] |
| Maize | 73 | 136 | 17.1 | 27.4 | 1.52 | 16.3 | Serbia | [26] |
| Sugar beet pulp | 1 | 3.0 | ND | ND | ND | ND | Spain | [10] |
| Silage (maize, grass) | 120 | 228 | 9.14 | 51.2 | 101 | 57.2 | Poland | [169] |
| Formula feed | ||||||||
| Pig | 1141 | 413 | 307 | 549 | 1514 | 1846 | Austria | [101] |
| Pig | 228 | 747 | 64.9 | 140 | 1222 | 247 | Spain | [170] |
| Poultry | 78 | 474.9 | 34.7 | 32.1 | 2190.2 | 396.0 | UK | [165] |
| Bovine | 8 | 51.4 | ND | 10.7 | 41.6 | 20.2 | Spain | [10] |
| Ovine | 13 | 129.6 | ND | 13.1 | 89.5 | 28.8 | Spain | [10] |
| Caprine | 1 | 23.2 | ND | 8.5 | 23.9 | 15 | Spain | [10] |
| Horses | 3 | 29.8 | ND | 10.1 | 43.8 | 15.5 | Spain | [10] |
| Porcine | 4 | 14.6 | ND | 11.9 | 55.1 | 24.0 | Spain | [10] |
| Poultry | 11 | 23.8 | ND | 11.9 | 51.1 | 23.1 | Spain | [10] |
| Rabbits | 2 | 13.5 | ND | 11.8 | 50.3 | 23.6 | Spain | [10] |
| Dogs | 3 | 40.5 | ND | ND | 24.8 | 10.1 | Spain | [10] |
| Cats | 3 | ND | ND | ND | 6.7 | 8.9 | Spain | [10] |
| Chicken | 43 | 29.3 | 12.6 | 5 | 39.8 | 15.7 | Tunisia | [171] |
| Cattle | 35 | 5.7 | 0.9 | 4.3 | 21.9 | ND | Tunisia | [171] |
| Rabbit | 12 | 2.3 | ND | 7.9 | 9.7 | 26.6 | Tunisia | [171] |
| Sheep | 16 | ND | 4.3 | 20.5 | 21.7 | 12.8 | Tunisia | [171] |
| Horse | 16 | 2.1 | ND | ND | 2.4 | 1.5 | Tunisia | [171] |
| Human | ||||||||
| Breast milk | 87 | 0.0017 | <LOQ | <LOQ | 0.0086 | 0.0019 | Austria | [172] |
| Fish | ||||||||
| Sea bass (Dicentrarchus labrax) | 10 | ND or <LOQ | ND or <LOQ | 6.9 | 12.8 | 31.5 | Spain (from aquaculture) | [112] |
| Sea bream (Sparus aurata) | 10 | ND or <LOQ | ND or <LOQ | 7.5 | 21.6 | 19 | Greece (from aquaculture) | [112] |
| Atlantic salmon (Salmo salar) | 10 | ND or <LOQ | ND or <LOQ | 29 | 103 | 94 | Norway (from aquaculture) | [112] |
| Rainbow trout (Oncorhynchus mykiss) | 10 | ND or <LOQ | ND or <LOQ | ND | 3.6 | 2.9 | Spain (from aquaculture) | [112] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertero, A.; Fossati, P.; Tedesco, D.E.A.; Caloni, F. Beauvericin and Enniatins: In Vitro Intestinal Effects. Toxins 2020, 12, 686. https://doi.org/10.3390/toxins12110686
Bertero A, Fossati P, Tedesco DEA, Caloni F. Beauvericin and Enniatins: In Vitro Intestinal Effects. Toxins. 2020; 12(11):686. https://doi.org/10.3390/toxins12110686
Chicago/Turabian StyleBertero, Alessia, Paola Fossati, Doriana Eurosia Angela Tedesco, and Francesca Caloni. 2020. "Beauvericin and Enniatins: In Vitro Intestinal Effects" Toxins 12, no. 11: 686. https://doi.org/10.3390/toxins12110686
APA StyleBertero, A., Fossati, P., Tedesco, D. E. A., & Caloni, F. (2020). Beauvericin and Enniatins: In Vitro Intestinal Effects. Toxins, 12(11), 686. https://doi.org/10.3390/toxins12110686