Phylogeny and Toxin Profile of Freshwater Pufferfish (Genus Pao) Collected from 2 Different Regions in Cambodia

 , , and
, , and
Abstract
:1. Introduction
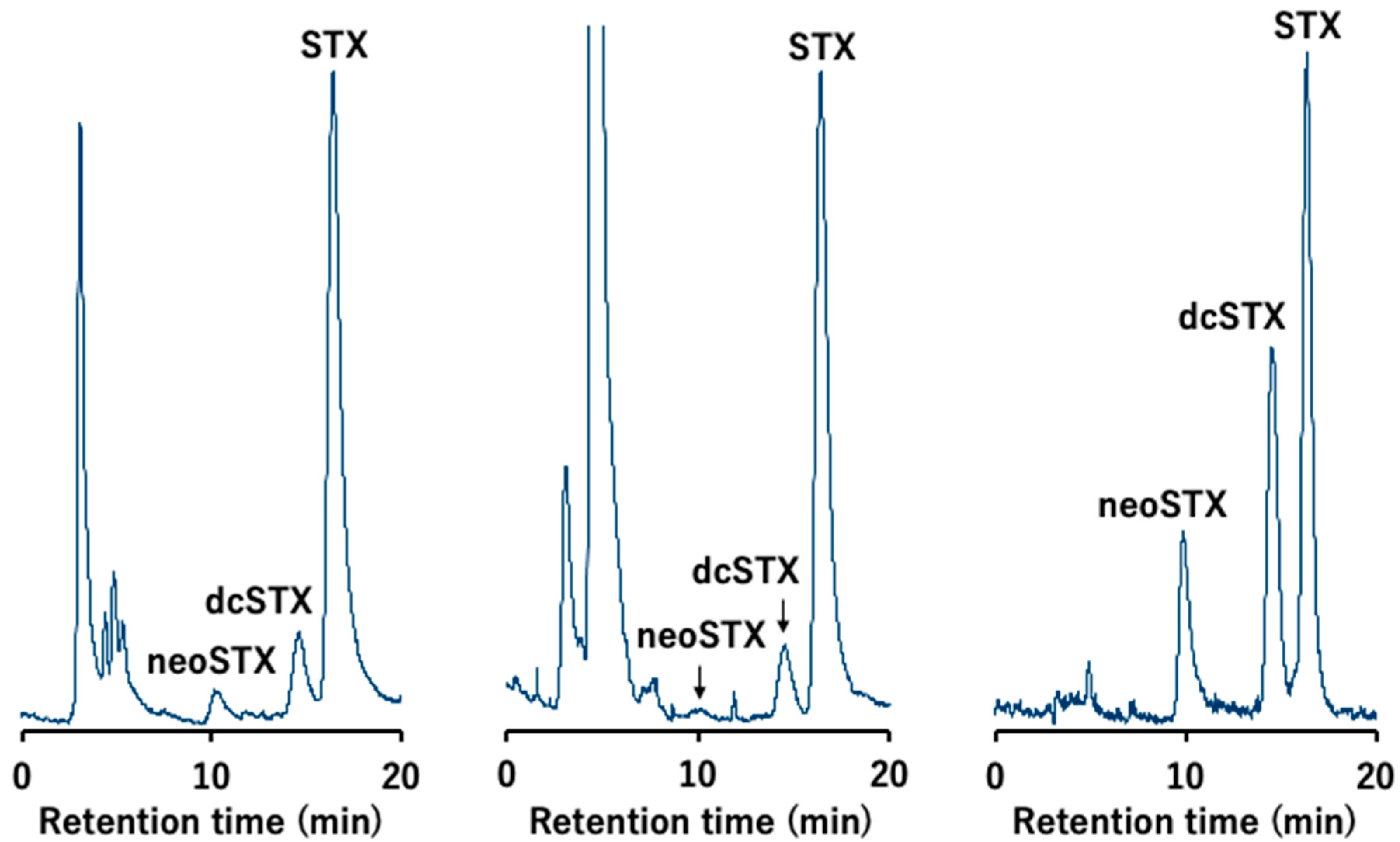
2. Results
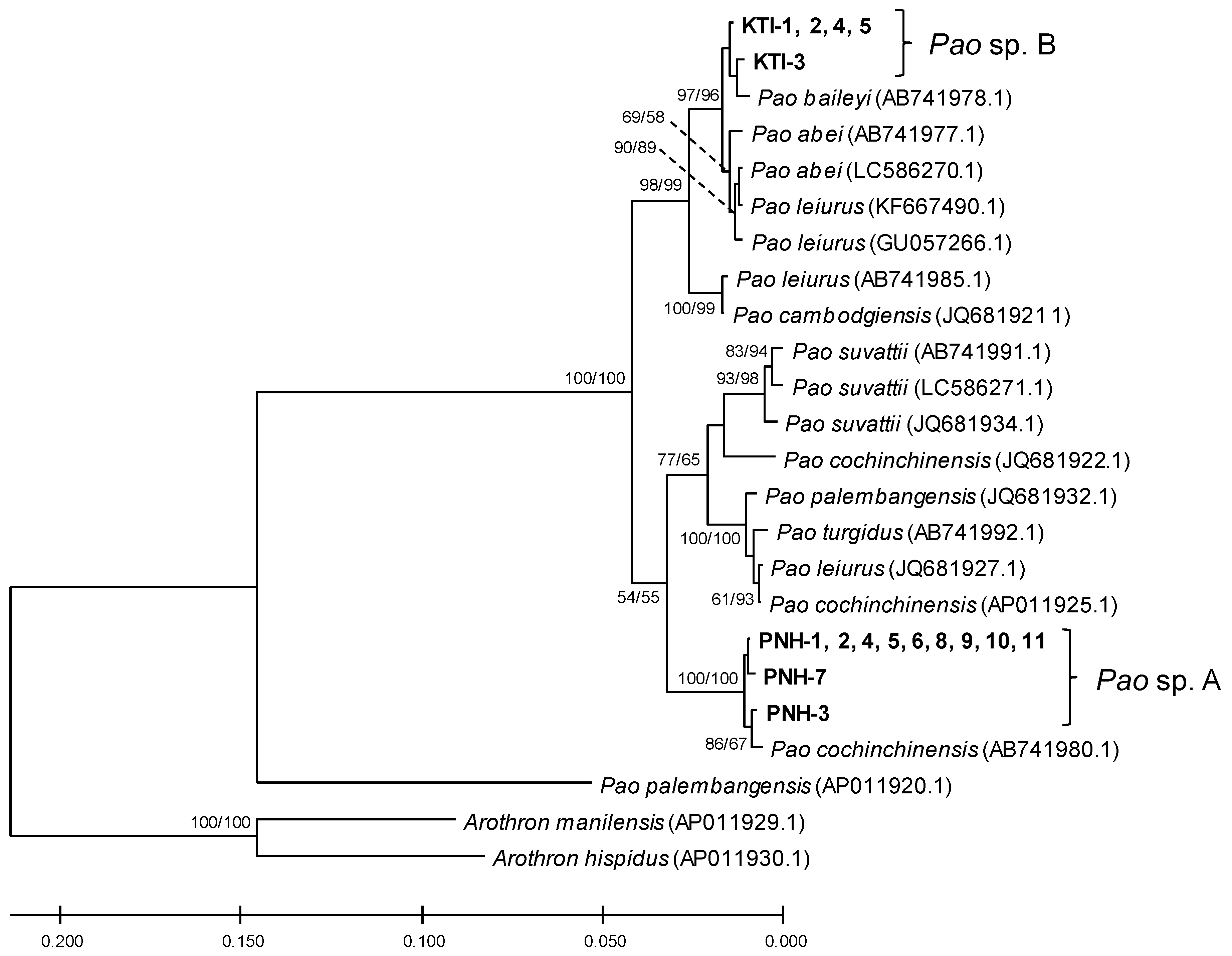
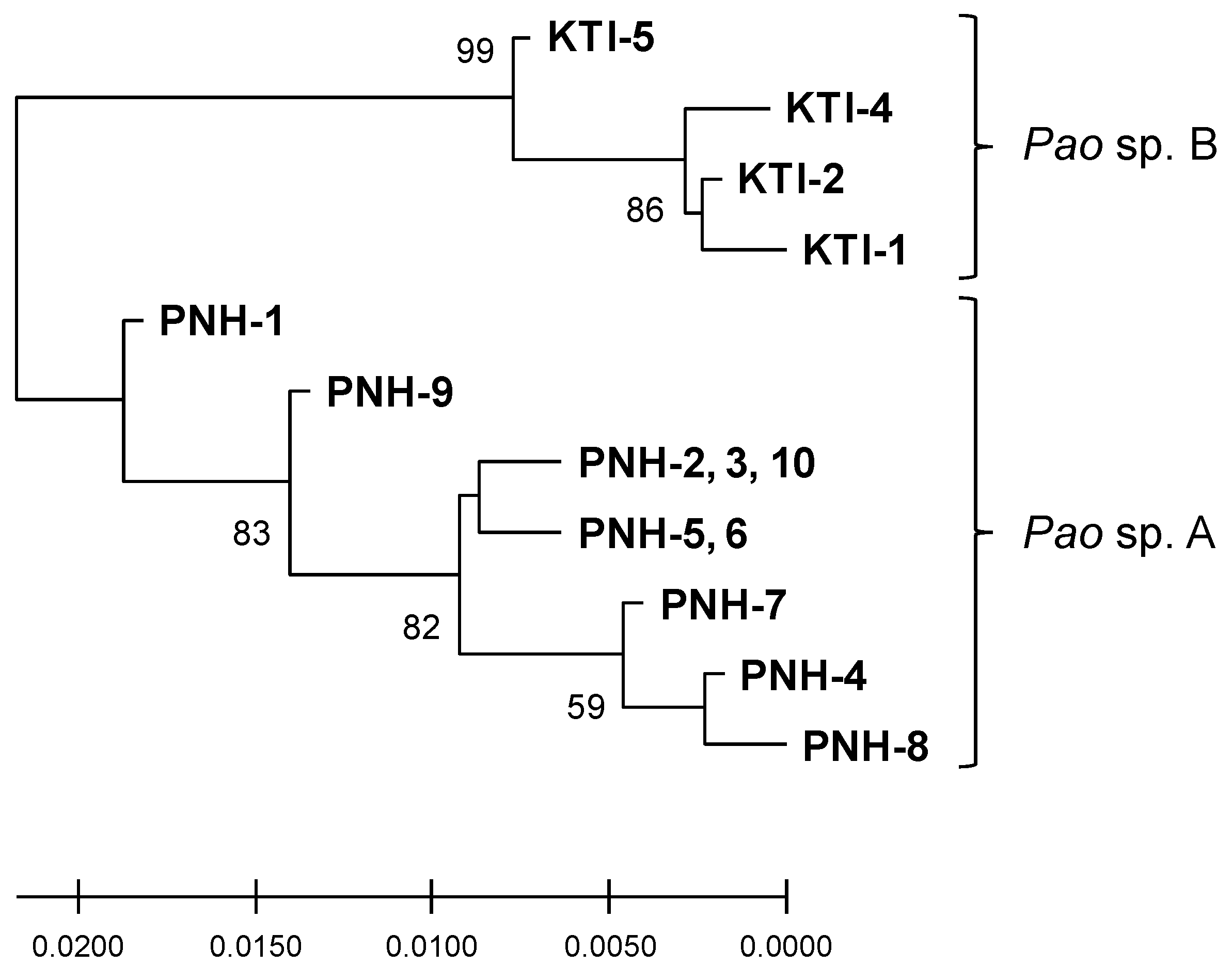
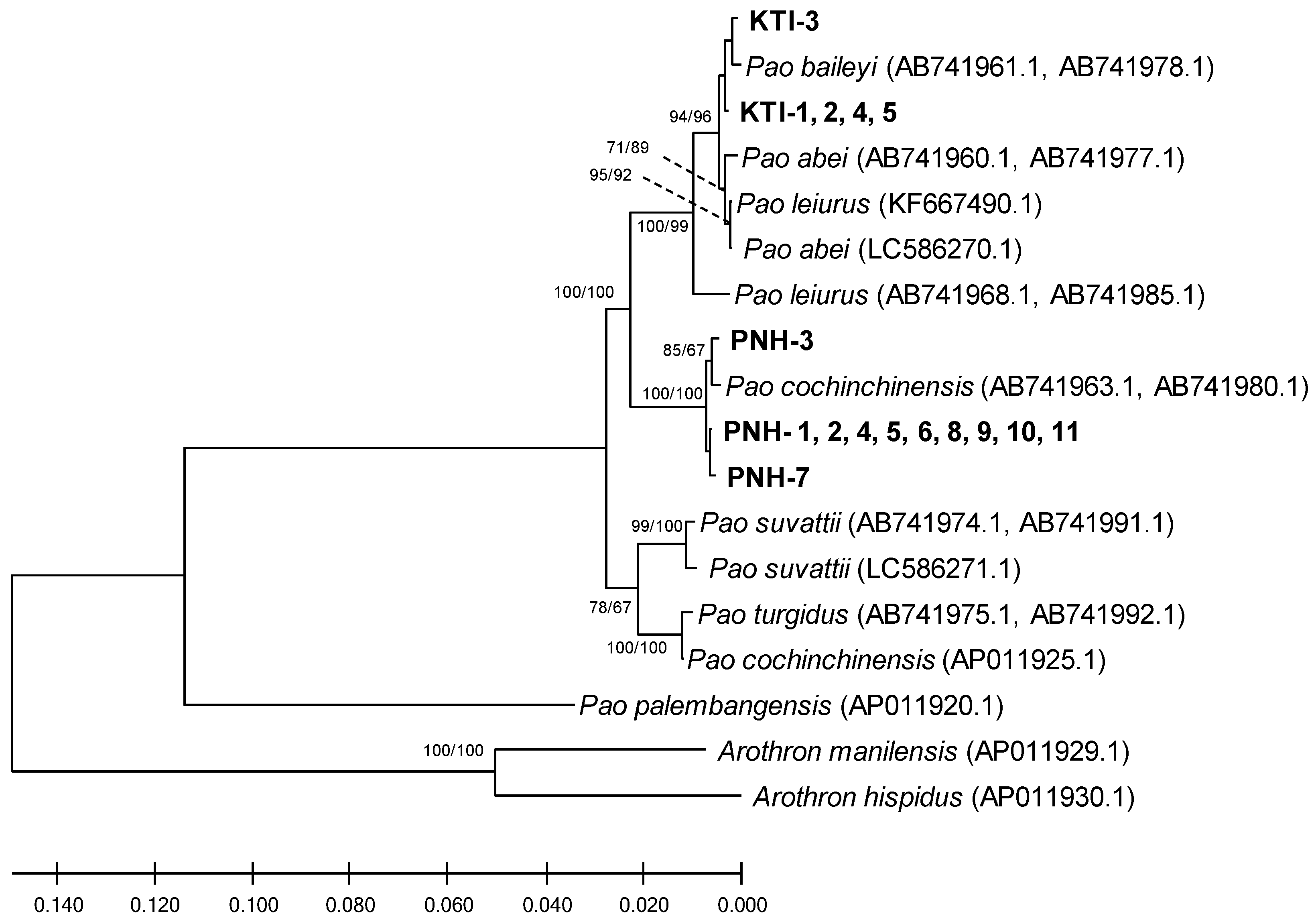
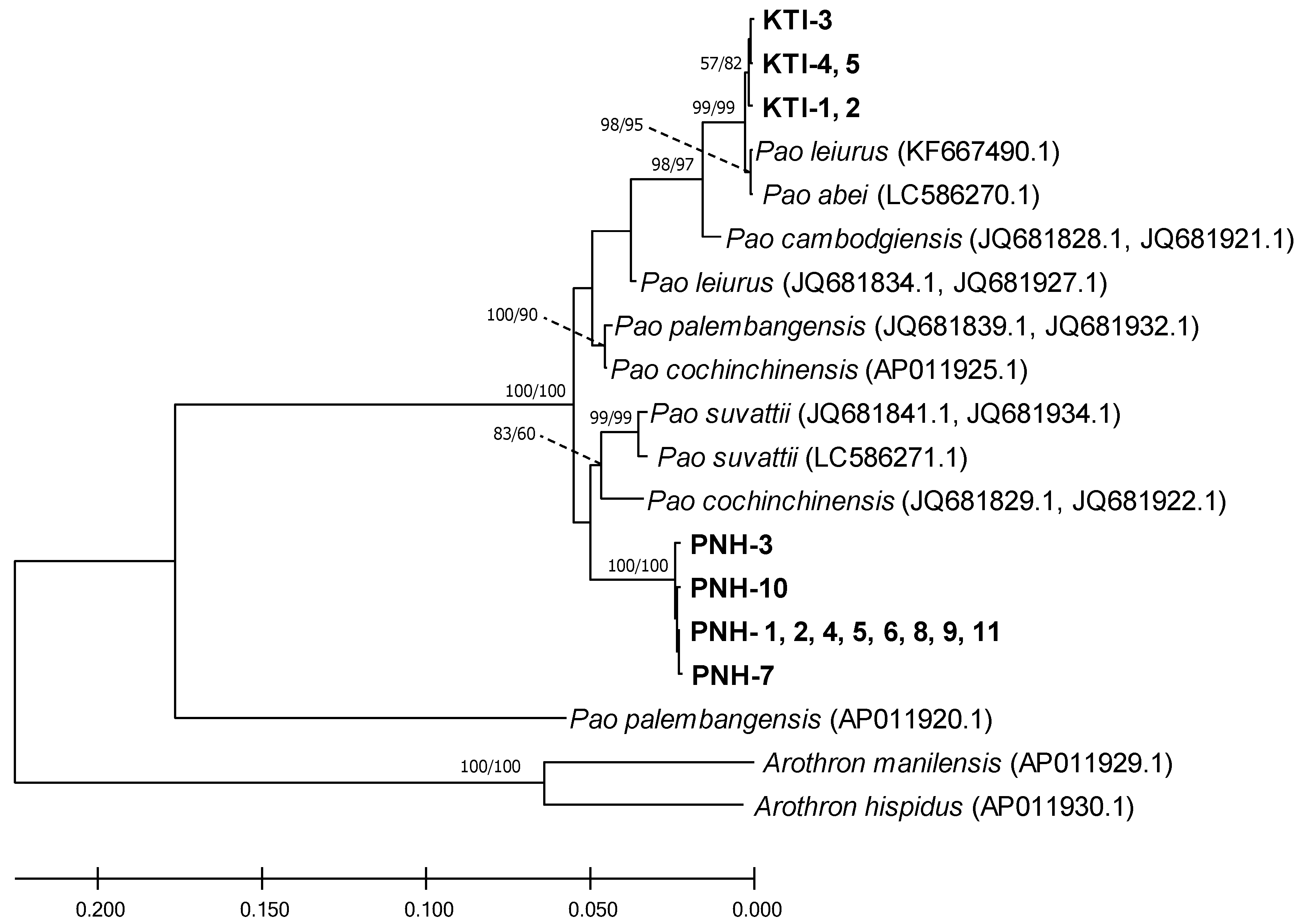
2.1. Phylogeny
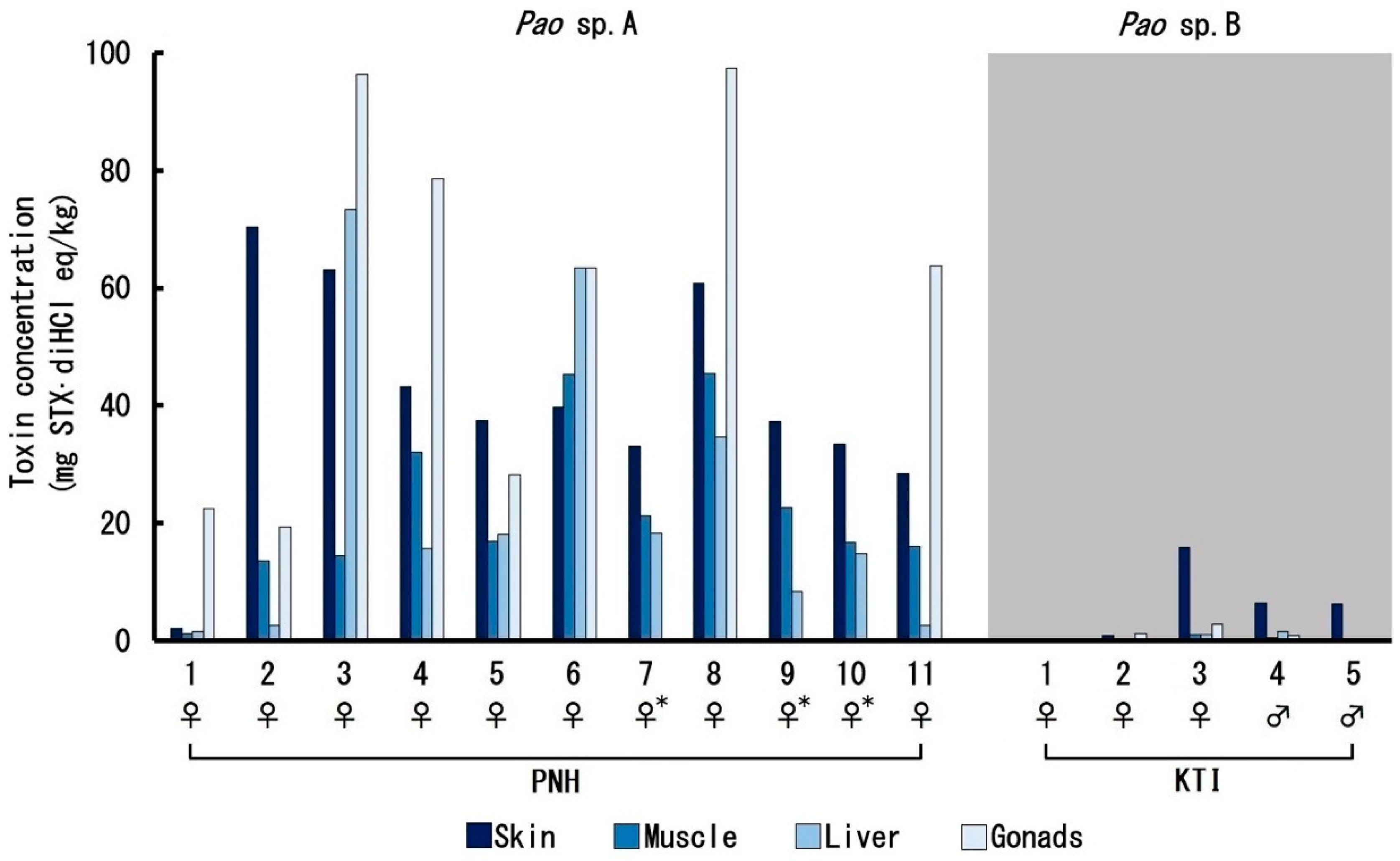
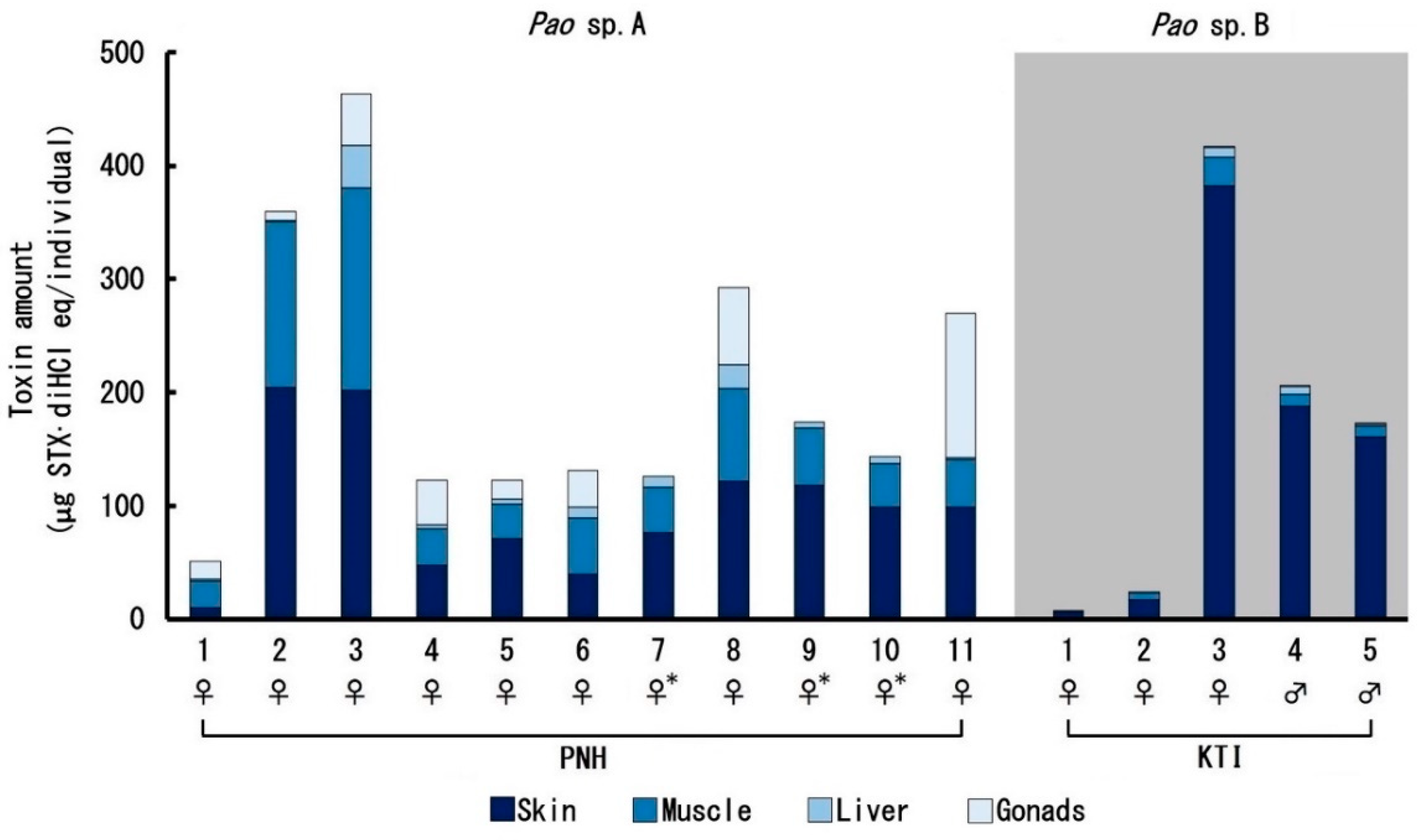
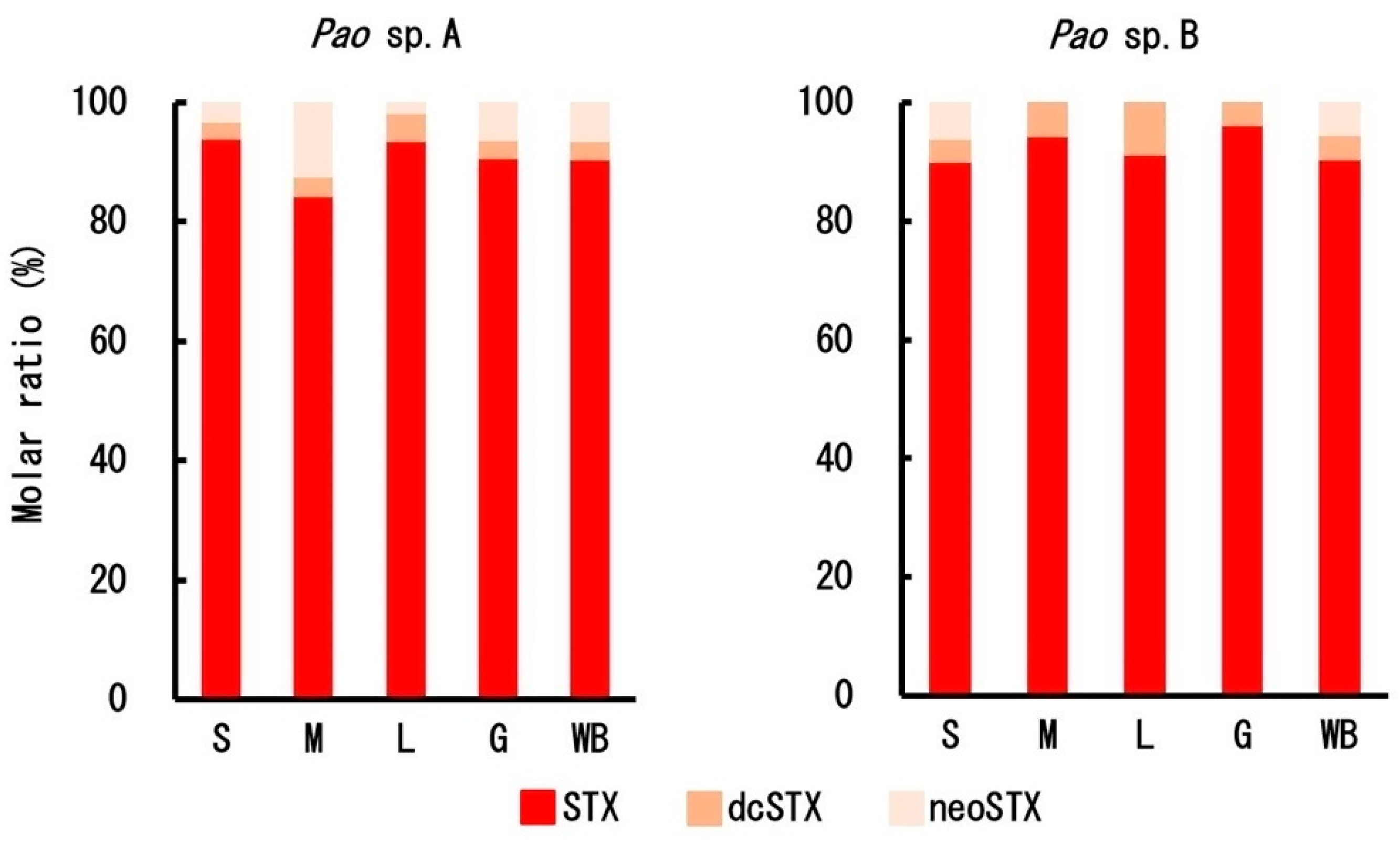
2.2. Toxin Profile
3. Discussion
4. Materials and Methods
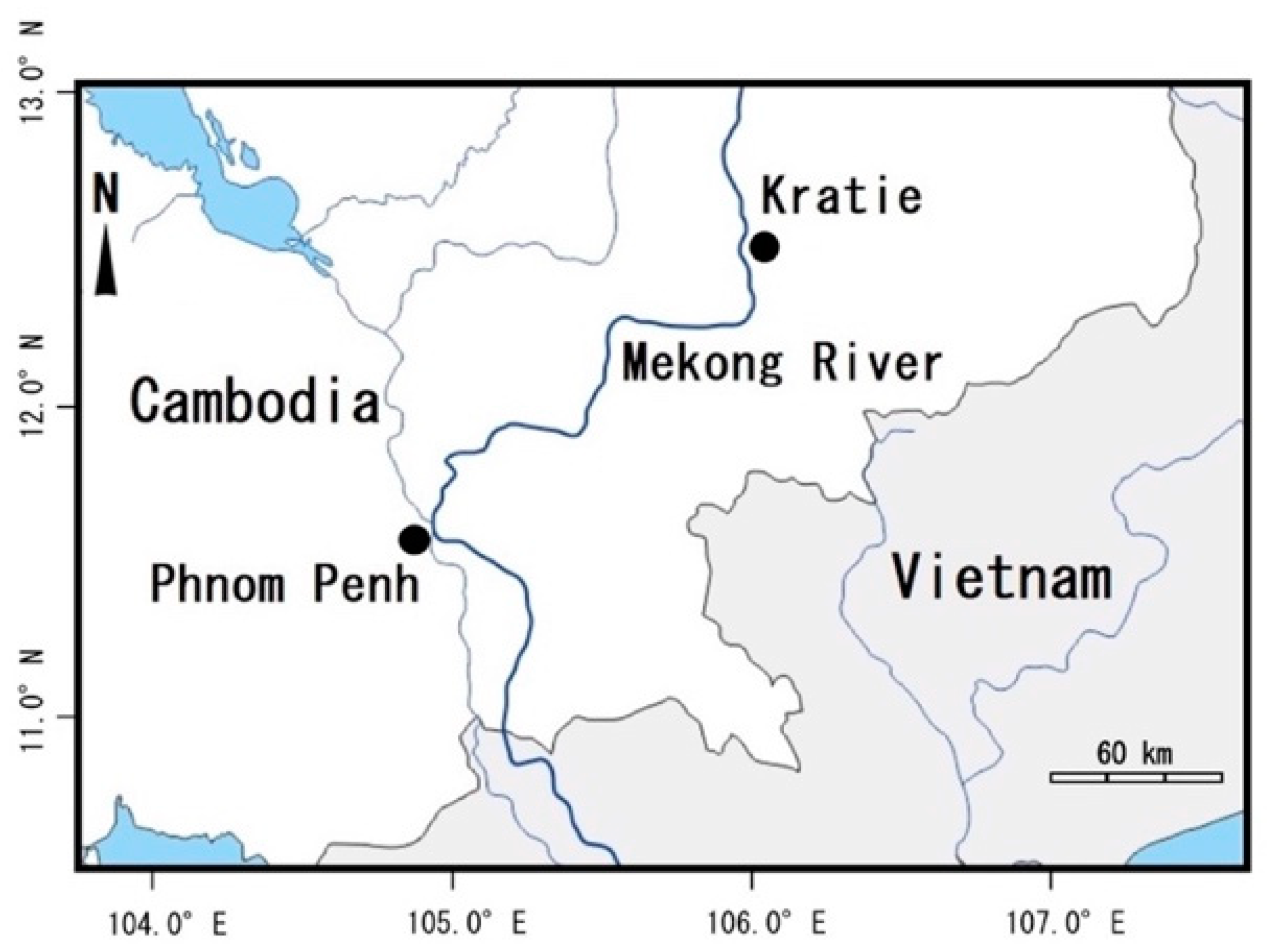
4.1. Pufferfish Specimens
4.2. DNA Extraction, PCR Amplification, and Sequencing
4.3. Phylogenetic Reconstructions
4.4. Toxin Quantification
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Sample | Sex | TL (cm) 1 | BW (g) 2 | Toxin Concentration (mg STXdi·HCL eq/kg) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Skin | Muscle | Liver 3 | Ovary 3,4 | Testis | |||||
| Pao sp. A | PNH-1 | ♀ | 9.7 | 26.1 | 2.1 | 1.2 | 1.5 | 22.5 | |
| 2 | ♀ | 8.0 | 15.1 | 70.5 | 13.7 | 2.7 | 19.3 | ||
| 3 | ♀ | 9.1 | 22.7 | 63.0 | 14.4 | 73.4 | 96.3 | ||
| 4 | ♀ | 7.0 | 14.4 | 43.1 | 32.1 | 15.8 | 78.5 | ||
| 5 | ♀ | 8.0 | 15.9 | 37.5 | 17.0 | 18.1 | 28.2 | ||
| 6 | ♀ | 6.5 | 12.8 | 39.7 | 45.3 | 63.4 | 63.5 | ||
| 7 | ♀ | 7.5 | 21.1 | 33.2 | 21.2 | 18.3 | – | ||
| 8 | ♀ | 7.5 | 22.2 | 60.8 | 45.4 | 34.7 | 97.3 | ||
| 9 | ♀ | 7.6 | 17.2 | 37.2 | 22.7 | 8.3 | – | ||
| 10 | ♀ | 8.0 | 17.0 | 33.4 | 16.7 | 14.8 | – | ||
| 11 | ♀ | 8.2 | 22.0 | 28.4 | 16.0 | 2.7 | 63.7 | ||
| Mean ± SD | 7.9 ± 0.9 | 18.8 ± 4.2 | 40.8 ± 18.9 | 22.3 ± 13.6 | 23.1 ± 24.4 | 58.7 ± 31.9 | |||
| Pao sp. B | KTI-1 | ♀ | 15.0 | 85.0 | 0.3 | 0.1 | 0 | 0 | |
| 2 | ♀ | 16.0 | 90.0 | 0.8 | 0.3 | 0 | 1.2 | ||
| 3 | ♀ | 15.7 | 108 | 15.8 | 1.1 | 1.0 | 2.8 | ||
| 4 | ♂ | 15.5 | 105 | 6.5 | 0.5 | 1.5 | 0.9 | ||
| 5 | ♂ | 16.0 | 109 | 6.3 | 0.4 | 0.2 | 0.3 | ||
| Mean ± SD | 15.6 ± 0.4 | 99.4 ± 10.5 | 6.0 ± 6.2 | 0.5 ± 0.4 | 0.6 ± 0.7 | 1.3 ± 1.4 | 0.6 | ||


| Sample | 7 | 23 | 24 | 33 | 138 | 169 | 193 | 225 | 228 | 231 | 232 | 286 | 367 | 429 | 432 | 448 | 499 | 505 | 524 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PNH-1 | A | T | G | C | C | T | C | C | T | G | C | G | A | G | C | C | T | T | A |
| PNH-2 | . | . | . | Y 1 | . | . | . | . | W 1 | R 1 | M 1 | . | . | . | S 1 | Y 1 | . | . | . |
| PNH-3 | . | C | . | Y | . | . | . | . | W | R | M | . | . | . | S | Y | . | . | . |
| PNH-4 | . | Y | . | Y | M | . | . | . | W | R | M | . | . | . | S | . | . | . | R |
| PNH-5 | . | . | . | Y | . | . | . | M | W | R | M | . | . | . | S | . | . | . | . |
| PNH-6 | . | . | . | Y | . | . | . | M | W | R | M | . | . | . | S | . | . | . | . |
| PNH-7 | . | Y | . | Y | M | . | . | . | W | R | M | . | . | . | S | . | . | . | G |
| PNH-8 | nd 1 | nd | nd | nd | M | . | . | M | W | R | M | . | . | . | S | . | . | . | R |
| PNH-9 | nd | nd | nd | nd | . | . | . | . | . | R | M | . | . | . | . | . | . | . | . |
| PNH-10 | nd | nd | nd | nd | . | . | . | . | W | R | M | . | . | . | S | Y | . | . | . |
| PNH-11 | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | . | . | . | . | . | . | . | . |
| KTI-1 | nd | C | . | . | . | G | Y | . | . | . | . | A | W | K 1 | G | . | C | G | G |
| KTI-2 | G | C | . | . | . | G | Y | . | . | . | . | A | W | T | G | . | C | G | G |
| KTI-3 | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd |
| KTI-4 | G | C | R | . | . | G | Y | . | . | . | . | R | W | T | G | . | C | G | G |
| KTI-5 | nd | C | R | . | . | G | . | . | . | . | . | A | . | T | G | . | C | G | G |

References
- Noguchi, T.; Arakawa, O. Tetrodotoxin—Distribution and accumulation in aquatic organisms, and cases of human intoxication. Mar. Drugs 2008, 6, 220–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmud, Y.; Yamamori, K.; Noguchi, T. Occurrence of TTX in a brackish water puffer ‘midorifugu’, Tetraodon nigroviridis, collected from Thailand. J. Food Hyg. Soc. Jpn. 1999, 40, 363–367. [Google Scholar] [CrossRef] [Green Version]
- Mahmud, Y.; Yamamori, K.; Noguchi, T. Toxicity and tetrodotoxin as the toxic principle of a brackish water puffer, Tetraodon steindachneri, collected from Thailand. J. Food Hyg. Soc. Jpn. 1999, 40, 391–395. [Google Scholar] [CrossRef]
- Kungsuwan, A.; Arakawa, O.; Promdet, M.; Onoue, Y. Occurrence of paralytic shellfish poisons in Thai freshwater puffers. Toxicon 1997, 35, 1341–1346. [Google Scholar] [CrossRef]
- Sato, S.; Kodama, M.; Ogata, T.; Saitanu, K.; Furuya, M.; Hirayama, K.; Kamimura, K. Saxitoxin as a toxic principle of a freshwater puffer, Tetraodon fangi, in Thailand. Toxicon 1997, 35, 137–140. [Google Scholar] [CrossRef]
- Zaman, L.; Arakawa, O.; Shimosu, A.; Onoue, Y. Occurrence of paralytic shellfish poison in Bangladeshi freshwater puffers. Toxicon 1997, 35, 423–431. [Google Scholar] [CrossRef]
- Ngy, L.; Tada, K.; Yu, C.F.; Takatani, T.; Arakawa, O. Occurrence of paralytic shellfish toxins in Cambodian Mekong pufferfish Tetraodon turgidus: Selective toxin accumulation in the skin. Toxicon 2008, 51, 280–288. [Google Scholar] [CrossRef] [Green Version]
- Landsberg, J.H.; Hall, S.; Johannessen, J.N.; White, K.D.; Conrad, S.M. Saxitoxin puffer fish poisoning in the United States, with the first report of Pyrodinium bahamense as the putative toxin source. Environ. Health Perspect. 2006, 114, 1502–1507. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.; Ogata, T.; Borja, V.; Gonzales, C.; Fukuyo, Y.; Kodama, M. Frequent occurrence of paralytic shellfish poisoning toxins as dominant toxins in marine puffer from tropical water. Toxicon 2000, 38, 1101–1109. [Google Scholar] [CrossRef]
- Nakashima, K.; Arakawa, O.; Taniyama, S.; Nonaka, M.; Takatani, T.; Yamamori, K.; Fuchi, Y.; Noguchi, T. Occurrence of saxitoxins as a major toxin in the ovary of a marine puffer Arothron firmamentum. Toxicon 2004, 43, 207–212. [Google Scholar] [CrossRef]
- Barrientos, R.G.; Hernández-Mora, G.; Alegre, F.; Field, T.; Flewelling, L.; McGrath, S.; Deeds, J.; Chacón, Y.S.; Arrieta, K.R.; Vargas, E.C.; et al. Saxitoxin poisoning in green turtles (Chelonia mydas) linked to scavenging on mass mortality of Caribbean sharpnose puffer fish (Canthigaster rostrata-Tetraodontidae). Front. Vet. Sci. 2019, 6, 466. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Sonoyama, T.; Yamada, M.; Gao, W.; Tatsuno, R.; Takatani, T.; Arakawa, O. Co-occurrence of tetrodotoxin and saxitoxins and their intra-body distribution in the pufferfish Canthigaster valentini. Toxins 2020, 12, 436. [Google Scholar] [CrossRef]
- Kodama, M.; Ogata, T.; Noguchi, T.; Maruyama, J.; Hashimoto, K. Occurrence of saxitoxin and other toxins in the liver of pufferfish Takifugu Pardalis. Toxicon 1983, 21, 897–900. [Google Scholar] [CrossRef]
- Nakamura, M.; Oshima, Y.; Yasumoto, T. Occurrence of saxitoxin in puffer fish. Toxicon 1984, 22, 381–385. [Google Scholar] [CrossRef]
- Jang, J.; Yotsu-Yamashita, M. Distribution of tetrodotoxin, saxitoxin, and their analogs among tissues of the puffer fish Fugu pardalis. Toxicon 2006, 48, 980–987. [Google Scholar] [CrossRef]
- Zhang, X.; Zong, J.; Chen, S.; Li, M.; Lu, Y.; Wang, R.; Xu, H. Accumulation and elimination of tetrodotoxin in the pufferfish Takifugu obscurus by dietary administration of the wild toxic gastropod Nassarius semiplicata. Toxins 2020, 12, 278. [Google Scholar] [CrossRef]
- Honda, S.; Arakawa, O.; Takatani, T.; Tachibana, K.; Yagi, M.; Tanigawa, A.; Noguchi, T. Toxification of cultured puffer fish Takifugu rubripes by feeding on tetrodotoxin-containing diet. Nippon Suisan Gakkaishi 2005, 71, 815–820. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, K.; Murakami, Y.; Emoto, Y.; Ngy, L.; Taniyama, S.; Yagi, M.; Takatani, T.; Arakawa, O. Transfer profile of intramuscularly administered tetrodotoxin to non-toxic cultured specimens of the pufferfish Takifugu rubripes. Toxicon 2009, 53, 99–103. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Araki, T.; Tatsuno, R.; Nina, S.; Ikeda, K.; Hamasaki, M.; Sakakura, Y.; Takatani, T.; Arakawa, O. Transfer profile of intramuscularly administered tetrodotoxin to artificial hybrid specimens of pufferfish, Takifugu rubripes and Takifugu niphobles. Toxicon 2011, 58, 565–569. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Araki, T.; Tatsuno, R.; Nina, S.; Ikeda, K.; Takatani, T.; Arakawa, O. Transfer profile of orally and intramuscularly administered tetrodotoxin to artificial hybrid specimens of the pufferfish Takifugu rubripes and Takifugu porphyreus. J. Food Hyg. Soc. Jpn. 2012, 55, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Tatsuno, R.; Shikina, M.; Shirai, Y.; Wang, J.; Soyano, K.; Nishihara, G.N.; Takatani, T.; Arakawa, O. Change in the transfer profile of orally administered tetrodotoxin to non-toxic cultured pufferfish Takifugu rubripes depending of its development stage. Toxicon 2013, 65, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Tatsuno, R.; Gao, W.; Ibi, K.; Mine, T.; Okita, K.; Nishihara, G.N.; Takatani, T.; Arakawa, O. Profile differences in tetrodotoxin transfer to skin and liver in the pufferfish Takifugu rubripes. Toxicon 2017, 130, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Yotsu-Yamashita, M.; Sugimoto, A.; Terakawa, T.; Shoji, Y.; Miyazawa, T.; Yasumoto, T. Purification, characterization, and cDNA cloning of a novel soluble saxitoxin and tetrodotoxin binding protein from plasma of the puffer fish, Fugu pardalis. Eur. J. Biochem. 2001, 268, 5937–5946. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Okoshi, N.; Watanabe, K.; Araki, N.; Yamaki, H.; Shoji, Y.; Terakawa, T. Localization of pufferfish saxitoxin and tetrodotoxin binding protein (PSPBP) in the tissues of the pufferfish, Takifugu pardalis, analyzed by immunohistochemical staining. Toxicon 2013, 72, 23–28. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Yamaki, H.; Okoshi, N.; Araki, N. Distribution of homologous proteins to puffer fish saxitoxin and tetrodotoxin binding protein in the plasma of puffer fish and among the tissues of Fugu pardalis examined by Western blot analysis. Toxicon 2010, 55, 1119–1124. [Google Scholar] [CrossRef]
- Tatsuno, R.; Yamaguchi, K.; Takatani, T.; Arakawa, O. RT-PCR- and MALDI-TOF mass spectrometry-based identification and discrimination of isoforms homologous to pufferfish saxitoxin- and tetrodotoxin-binding protein in the plasma of non-toxic cultured pufferfish (Takifugu rubripes). Biosci. Biotechnol. Biochem. 2013, 77, 208–212. [Google Scholar] [CrossRef] [Green Version]
- Yotsu-Yamashita, M.; Nagaoka, Y.; Muramoto, K.; Cho, Y.; Konoki, K. Pufferfish saxitoxin and tetrodotoxin binding protein (PSPBP) analogues in the blood plasma of the pufferfish Arothron nigropunctatus, A. hispidus, A. manilensis, and Chelonodon patoca. Mar. Drugs 2018, 16, 224. [Google Scholar] [CrossRef] [Green Version]
- Cusick, K.D.; Sayler, G.S. An overview on the marine neurotoxin, saxitoxin: Genetics, molecular targets, methods of detection and ecological functions. Mar. Drugs 2013, 11, 991–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, W.; Kanahara, Y.; Yamada, M.; Tatsuno, R.; Yoshikawa, H.; Doi, H.; Takatani, T.; Arakawa, O. Contrasting toxin selectivity between the marine pufferfish Takifugu pardalis and the freshwater pufferfish Pao suvattii. Toxins 2019, 11, 470. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization & Food and Agriculture Organization of the United Nations. Toxicity Equivalence Factors for Marine Biotoxins Associated with Bivalve Molluscs; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Oba, Y.; Shimasaki, Y.; Oshima, Y.; Satone, H.; Kitano, T.; Nakao, M.; Kawabata, S.; Honjo, T. Purification and characterization of tributyltin-binding protein type 2 from plasma of Japanese flounder, Paralichthys olivaceus. J. Biochem. 2007, 142, 229–238. [Google Scholar] [CrossRef]
- Hashiguchi, Y.; Lee, J.M.; Shiraishi, M.; Komatsu, S.; Miki, S.; Shimasaki, Y.; Mochioka, N.; Kusakabe, T.; Oshima, Y. Characterization and evolutionary analysis of tributyltin-binding protein and pufferfish saxitoxin and tetrodotoxin-binding protein genes in toxic and nontoxic pufferfishes. J. Evol. Biol. 2015, 28, 1103–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oshima, Y. Post-column derivatization HPLC method for analysis of PSP. J. AOAC Inter. 1995, 78, 528–532. [Google Scholar] [CrossRef]
- Gao, W.; Kanahara, Y.; Tatsuno, R.; Soyano, K.; Nishihara, G.N.; Urata, C.; Takatani, T.; Arakawa, O. Maturation-associated changes in internal distribution and intra-ovarian microdistribution of tetrodotoxin in the pufferfish Takifugu pardalis. Fish. Sci. 2018, 84, 723–732. [Google Scholar] [CrossRef]
- Igarashi, Y.; Doi, H.; Yamanoue, Y.; Kinoshita, S.; Ishibashi, T.; Ushio, H.; Asakawa, S.; Nishida, M.; Watabe, S. Molecular phylogenetic relationship of Tetraodon pufferfish based on mitochondrial DNA analysis. Fish. Sci. 2013, 79, 243–250. [Google Scholar] [CrossRef]
- Takahashi, H.; Toyoda, A.; Yamazaki, T.; Narita, S.; Mashiko, T.; Yamazaki, Y. Asymmetric hybridization and introgression between sibling species of the pufferfish Takifugu that have undergone explosive speciation. Mar. Biol. 2017, 164, 90. [Google Scholar] [CrossRef]
- Yamasaki, Y.Y.; Nishida, M.; Suzuki, T.; Mukai, T.; Watanabe, K. Phylogeny, hybridization, and life history evolution of Rhinogobius gobies in Japan, inferred from multiple nuclear gene sequences. Mol. Phylogenetics Evol. 2015, 90, 20–33. [Google Scholar] [CrossRef] [Green Version]
- Quayle, D.B. Paralytic shellfish poisoning in British Columbia. Fish. Res. Bd. Can. Bull. 1969, 158, 1–68. [Google Scholar]
- Etheridge, S.M. Paralytic shellfish poisoning: Seafood safety and human health perspective. Toxicon 2010, 56, 108–122. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, K.; Emoto, Y.; Tatsuno, R.; Wang, J.-J.; Ngy, L.; Taniyama, S.; Takatani, T.; Arakawa, O. Maturation-associated change in toxicity of the pufferfish Takifugu poecilonotus. Toxicon 2010, 55, 289–297. [Google Scholar] [CrossRef] [Green Version]
- Zaman, L.; Arakawa, O.; Shimosu, A.; Shida, Y.; Onoue, Y. Occurrence of a methyl derivative of saxitoxin in Bangladeshi freshwater puffers. Toxicon 1988, 36, 627–630. [Google Scholar] [CrossRef]
- Palumbi, S.; Martin, A.; Romaro, S.; McMillan, W.O.; Stice, L.; Grabowski, G. The Simple Fool’s Guide to PCR. Version 2.0; University of Hawaii: Honolulu, HI, USA, 2002; pp. 1–45. [Google Scholar]
- Ivanova, N.V.; Zemlak, T.S.; Hanner, R.H.; Hebert, P.D.N. Universal primer cocktails for fish DNA barcoding. Mol. Ecol. Notes 2007, 7, 544–548. [Google Scholar] [CrossRef]
- Chen, W.J.; Bonillo, C.; Lecointre, G. Repeatability of clades as a criterion of reliability: A case study for molecular phylogeny of Acanthomorpha (Teleostei) with larger number of taxa. Mol. Phylogenetics Evol. 2003, 26, 262–288. [Google Scholar] [CrossRef]
- Santini, F.; Nguyen, M.T.T.; Sorenson, L.; Waltzek, T.B.; Lynch Alfaro, J.W.; Eastman, J.M.; Alfaro, M.E. Do habitat shifts drive diversification in teleost fishes? An example from the pufferfishes (Tetraodontidae). J. Evol. Biol. 2013, 26, 1003–1018. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanabe, A.S. Kakusan4 and Aminosan: Two programs for comparing nonpartitioned, proportional and separate models for combined molecular phylogenetic analyses of multilocus sequence data. Mol. Ecol. Resour. 2011, 11, 914–921. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Felsenstein, J. PHYLIP (Phylogeny Interference Package) Version 3.6. 2005. Available online: https://evolution.genetics.washington.edu/phylip.html (accessed on 20 August 2020).
- Price, M.N. Fast Tree-Comparison Tools. Available online: http://www.microbesonline.org/fasttree/treecmp.html (accessed on 20 August 2020).
- Arakawa, O.; Noguchi, T.; Shida, Y.; Onoue, Y. Occurrence of carbamoyl-N-hydroxy derivatives of saxitoxin and neosaxitoxin in a xanthid crab Zosimus aeneus. Toxicon 1994, 32, 175–183. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, H.; Yamada, A.; Goto, Y.; Horn, L.; Ngy, L.; Wada, M.; Doi, H.; Lee, J.S.; Takatani, T.; Arakawa, O. Phylogeny and Toxin Profile of Freshwater Pufferfish (Genus Pao) Collected from 2 Different Regions in Cambodia. Toxins 2020, 12, 689. https://doi.org/10.3390/toxins12110689
Zhu H, Yamada A, Goto Y, Horn L, Ngy L, Wada M, Doi H, Lee JS, Takatani T, Arakawa O. Phylogeny and Toxin Profile of Freshwater Pufferfish (Genus Pao) Collected from 2 Different Regions in Cambodia. Toxins. 2020; 12(11):689. https://doi.org/10.3390/toxins12110689
Chicago/Turabian StyleZhu, Hongchen, Akinori Yamada, Yui Goto, Linan Horn, Laymithuna Ngy, Minoru Wada, Hiroyuki Doi, Jong Soo Lee, Tomohiro Takatani, and Osamu Arakawa. 2020. "Phylogeny and Toxin Profile of Freshwater Pufferfish (Genus Pao) Collected from 2 Different Regions in Cambodia" Toxins 12, no. 11: 689. https://doi.org/10.3390/toxins12110689
APA StyleZhu, H., Yamada, A., Goto, Y., Horn, L., Ngy, L., Wada, M., Doi, H., Lee, J. S., Takatani, T., & Arakawa, O. (2020). Phylogeny and Toxin Profile of Freshwater Pufferfish (Genus Pao) Collected from 2 Different Regions in Cambodia. Toxins, 12(11), 689. https://doi.org/10.3390/toxins12110689