Tetanus Toxin Synthesis is Under the Control of A Complex Network of Regulatory Genes in Clostridium tetani

 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Genomic Analysis of Regulatory Genes in C. tetani
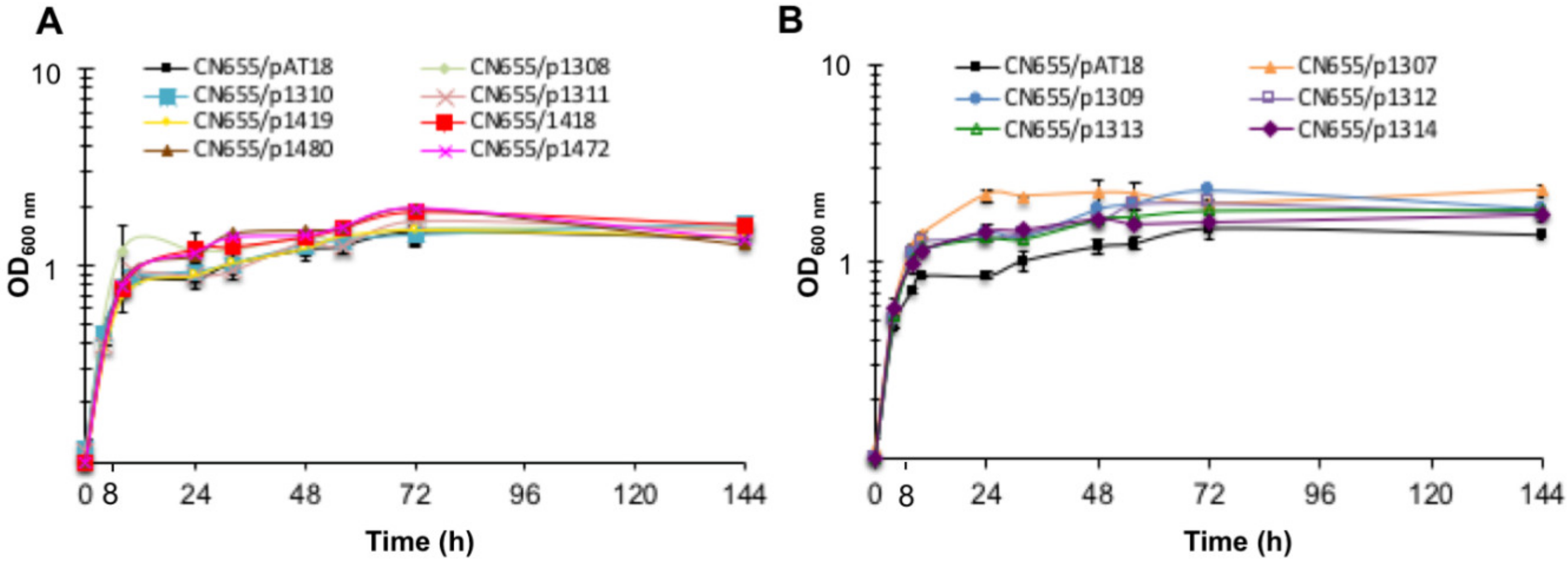
2.2. Construction of TCS Anti-Sense Strains
2.3. TeNT Synthesis is Altered in Five TCS Anti-Sense Strains and in the codY Anti-Sense Strain
2.4. Five TCSs and codY Control Tent and/or Tetr at the Transcriptional Level
2.5. The CTC_RS07315 and CTC_RS04785 Response Regulators as well as Cody Bind to the Tent Promoter
2.6. Inorganic Phosphate (Pi) Influences TeNT Production
2.7. Carbonate Stimulates TeNT Synthesis
2.8. The TCS CTC_RS05745/CTC_RS05750 is Involved in Bacterial Cell Wall Organization
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Construction of Vectors Encoding Anti-Sense mRNA for the Different Two-Component Systems (TCS) and Others Regulators
4.3. Tetanus Toxin Assay
4.4. Total RNA Extraction, Reverse Transcription and Quantitative Real-Time PCR Assay
4.5. Expression and Purification of Recombinant Proteins
4.6. Electrophoretic Mobility Shift Assay (EMSA)
4.7. Electron Microscopy
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Popoff, M.R.; Mazuet, C.; Poulain, B. Botulism and tetanus. In The Prokaryotes: Human Microbiology, 4th ed.; Springer: Berlin/Heidelberg, Germany, 2013; Volume 5, pp. 247–290. [Google Scholar]
- Bercsenyi, K.; Schmieg, N.; Bryson, J.B.; Wallace, M.; Caccin, P.; Golding, M.; Zanotti, G.; Greensmith, L.; Nischt, R.; Schiavo, G. Tetanus toxin entry. Nidogens are therapeutic targets for the prevention of tetanus. Science 2014, 346, 1118–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossetto, O.; Scorzeto, M.; Megighian, A.; Montecucco, C. Tetanus neurotoxin. Toxicon 2013, 66, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Schmieg, N.; Berscenyi, K.; Schiavo, G. Uptake and transport of clostridial neurotoxins. In The Comprehensive Sourcebook of Bacterial Protein Toxins, 4th ed.; Alouf, J., Ladant, D., Popoff, M.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 337–360. [Google Scholar]
- WHO. Tetanus vaccines: WHO position paper—February 2017. Wkly. Epidemiol. Rec. 2017, 92, 53–76. [Google Scholar]
- Licona-Cassani, C.; Steen, J.A.; Zaragoza, N.E.; Moonen, G.; Moutafis, G.; Hodson, M.P.; Power, J.; Nielsen, L.K.; Marcellin, E. Tetanus toxin production is triggered by the transition from amino acid consumption to peptides. Anaerobe 2016, 41, 113–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laird, W.J.; Aaronson, W.; Silver, R.P.; Habig, W.H.; Hardegree, M.C. Plasmid-associated toxigenicity in Clostridium tetani. J. Infect. Dis. 1980, 142, 623. [Google Scholar] [CrossRef] [PubMed]
- Finn, C.W.; Silver, R.P.; Habig, W.H.; Hardegree, M.C.; Zon, G.; Garon, C.F. The structural gene for tetanus neurotoxin is on a plasmid. Science 1984, 224, 881–884. [Google Scholar] [CrossRef]
- Brüggemann, H.; Bäumer, S.; Fricke, W.F.; Wiezr, A.; Liesagang, H.; Decker, I.; Herzberg, C.; Martinez-Arias, R.; Henne, A.; Gottschalk, G. The genome sequence of Clostridium tetani, the causative agent of tetanus disease. Proc. Natl. Acad. Sci. USA 2003, 100, 1316–1321. [Google Scholar] [CrossRef] [Green Version]
- Bruggemann, H.; Brzuszkiewicz, E.; Chapeton-Montes, D.; Plourde, L.; Speck, D.; Popoff, M.R. Genomics of Clostridium tetani. Res. Microbiol. 2015, 166, 326–331. [Google Scholar] [CrossRef]
- Chapeton-Montes, D.; Plourde, L.; Bouchier, C.; Ma, L.; Diancourt, L.; Criscuolo, A.; Popoff, M.R.; Bruggemann, H. The population structure of Clostridium tetani deduced from its pan-genome. Sci. Rep. 2019, 9, 11220. [Google Scholar] [CrossRef] [Green Version]
- Fournier, P.E.; Levy, P.Y.; Million, M.; Croce, O.; Blanc-Tailleur, C.; Brouqui, P.; Raoult, D. Genome of a chronic osteitis-causing Clostridium tetani. New Microbes New Infect. 2014, 2, 25–26. [Google Scholar] [CrossRef] [Green Version]
- Marvaud, J.C.; Eisel, U.; Binz, T.; Niemann, H.; Popoff, M.R. tetR is a positive regulator of the tetanus toxin gene in Clostridium tetani and is homologous to botR. Infect. Immun. 1998, 66, 5698–5702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marvaud, J.C.; Gibert, M.; Inoue, K.; Fujinaga, V.; Oguma, K.; Popoff, M.R. botR is a positive regulator of botulinum neurotoxin and associated non toxic protein genes in Clostridium botulinum A. Mol. Microbiol. 1998, 29, 1009–1018. [Google Scholar] [CrossRef] [PubMed]
- Dupuy, B.; Raffestin, S.; Matamouros, S.; Mani, N.; Popoff, M.R.; Sonenshein, A.L. Regulation of toxin and bacteriocin gene expression in Clostridium by interchangeable RNA polymerase sigma factors. Mol. Microbiol. 2006, 60, 1044–1057. [Google Scholar] [CrossRef] [PubMed]
- Raffestin, S.; Dupuy, B.; Marvaud, J.C.; Popoff, M.R. BotR/A and TetR are alternative RNA polymerase sigma factors controlling the expression of the neurotoxin and associated protein genes in Clostridium botulinum type A and Clostridium tetani. Mol. Microbiol. 2005, 55, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Brinsmade, S.R. CodY, a master integrator of metabolism and virulence in Gram-positive bacteria. Curr. Genet. 2017, 63, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Mitrophanov, A.Y.; Groisman, E.A. Signal integration in bacterial two-component regulatory systems. Genes Dev. 2008, 22, 2601–2611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenz, L.; Francois, P.; Whiteson, K.; Wolz, C.; Linder, P.; Schrenzel, J. The CodY pleiotropic repressor controls virulence in gram-positive pathogens. FEMS Immunol. Med. Microbiol. 2011, 62, 123–139. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Dahlsten, E.; Korkeala, H.; Lindström, M. Positive regulation of botulinum neurotoxin gene expression by CodY in Clostridium botulinum ATCC 3502. Appl. Environ. Microbiol. 2014, 80, 7651–7658. [Google Scholar] [CrossRef] [Green Version]
- Mascher, G.; Mertaoja, A.; Korkeala, H.; Lindstrom, M. Neurotoxin synthesis is positively regulated by the sporulation transcription factor Spo0A in Clostridium botulinum type E. Environ. Microbiol. 2017, 19, 4287–4300. [Google Scholar] [CrossRef]
- Kirk, D.G.; Palonen, E.; Korkeala, H.; Lindstrom, M. Evaluation of normalization reference genes for RT-qPCR analysis of spo0A and four sporulation sigma factor genes in Clostridium botulinum Group I strain ATCC 3502. Anaerobe 2014, 26, 14–19. [Google Scholar] [CrossRef]
- Wörner, K.; Szurmant, H.; Chiang, C.; Hoch, J.A. Phosphorylation and functional analysis of the sporulation initiation factor Spo0A from Clostridium botulinum. Mol. Microbiol. 2006, 59, 1000–1012. [Google Scholar] [CrossRef] [PubMed]
- Willing, S.E.; Richards, E.J.; Sempere, L.; Dale, A.G.; Cutting, S.M.; Fairweather, N.F. Increased toxin expression in a Clostridium difficile mfd mutant. BMC Microbiol. 2015, 15, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connan, C.; Brueggemann, H.; Mazuet, C.; Raffestin, S.; Cayet, N.; Popoff, M.R. Two-component systems are involved in the regulation of botulinum neurotoxin synthesis in Clostridium botulinum type A strain Hall. PLoS ONE 2012, 7, e41848. [Google Scholar] [CrossRef]
- Zhang, Z.; Korkeala, H.; Dahlsten, E.; Sahala, E.; Heap, J.T.; Minton, N.P.; Lindstrom, M. Two-component signal transduction system CBO0787/CBO0786 represses transcription from botulinum neurotoxin promoters in Clostridium botulinum ATCC 3502. PLoS Pathog. 2013, 9, e1003252. [Google Scholar] [CrossRef] [Green Version]
- Artin, I.; Carter, A.T.; Holst, E.; Lovenklev, M.; Mason, D.R.; Peck, M.W.; Radstrom, P. Effects of carbon dioxide on neurotoxin gene expression in nonproteolytic Clostridium botulinum Type E. Appl. Environ. Microbiol. 2008, 74, 2391–2397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artin, I.; Mason, D.R.; Pin, C.; Schelin, J.; Peck, M.W.; Holst, E.; Radstrom, P.; Carter, A.T. Effects of carbon dioxide on growth of proteolytic Clostridium botulinum, its ability to produce neurotoxin, and its transcriptome. Appl. Environ. Microbiol. 2010, 76, 1168–1172. [Google Scholar] [CrossRef] [Green Version]
- Lawson, P.A.; Rainey, F.A. Proposal to restrict the genus Clostridium Prazmowski to Clostridium butyricum and related species. Int. J. Syst. Evol. Microbiol. 2016, 66, 1009–1016. [Google Scholar] [CrossRef]
- Fratelli, F.; Siquini, T.J.; de Abreu, M.E.; Higashi, H.G.; Converti, A.; de Carvalho, J.C. Fed-batch production of tetanus toxin by Clostridium tetani. Biotechnol. Prog. 2010, 26, 88–92. [Google Scholar] [CrossRef]
- Fratelli, F.; Siquini, T.J.; Prado, S.M.; Higashi, H.G.; Converti, A.; de Carvalho, J.C. Effect of medium composition on the production of tetanus toxin by Clostridium tetani. Biotechnol. Prog. 2005, 21, 756–761. [Google Scholar] [CrossRef]
- Porfirio, Z.; Prado, S.M.; Vancetto, M.D.C.; Fratelli, F.; Alves, E.W.; Raw, I.; Fernandes, B.L.; Camargo, A.C.M.; Lebrun, I. Specific peptides of casein pancreatic digestion enhance the production of tetanus toxin. J. Appl. Microbiol. 1997, 83, 678–684. [Google Scholar] [CrossRef] [Green Version]
- Carter, G.P.; Cheung, J.K.; Larcombe, S.; Lyras, D. Regulation of toxin production in the pathogenic clostridia. Mol. Microbiol. 2014, 91, 221–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connan, C.; Deneve, C.; Mazuet, C.; Popoff, M.R. Regulation of toxin synthesis in Clostridium botulinum and Clostridium tetani. Toxicon 2013, 75, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Brüggemann, H.; Gottschalk, G. Insights in metabolism and toxin production from the complete genome sequence of Clostridium tetani. Anaerobe 2004, 10, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Mellanby, J. The effect of glutamate on toxin production by Clostridium tetani. J. Gen. Microbiol. 1968, 54, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Ohtani, K.; Yuan, Y.; Hassan, S.; Wang, R.; Wang, Y.; Shimizu, T. Virulence gene regulation by the agr system in Clostridium perfringens. J. Bacteriol. 2009, 191, 3919–3927. [Google Scholar] [CrossRef] [Green Version]
- Ohtani, K.; Shimizu, T. Regulation of toxin gene expression in Clostridium perfringens. Res. Microbiol. 2015, 166, 280–289. [Google Scholar] [CrossRef]
- Ohtani, K.; Shimizu, T. Regulation of Toxin Production in Clostridium perfringens. Toxins 2016, 8, 207. [Google Scholar] [CrossRef]
- Feng, L.; Zhu, J.; Chang, H.; Gao, X.; Gao, C.; Wei, X.; Yuan, F.; Bei, W. The CodY regulator is essential for virulence in Streptococcus suis serotype 2. Sci. Rep. 2016, 6, 21241. [Google Scholar] [CrossRef]
- Lemos, J.A.; Nascimento, M.M.; Lin, V.K.; Abranches, J.; Burne, R.A. Global regulation by (p)ppGpp and CodY in Streptococcus mutans. J. Bacteriol. 2008, 190, 5291–5299. [Google Scholar] [CrossRef] [Green Version]
- Lindback, T.; Mols, M.; Basset, C.; Granum, P.E.; Kuipers, O.P.; Kovacs, A.T. CodY, a pleiotropic regulator, influences multicellular behaviour and efficient production of virulence factors in Bacillus cereus. Environ. Microbiol. 2012, 14, 2233–2246. [Google Scholar] [CrossRef] [Green Version]
- Lobel, L.; Sigal, N.; Borovok, I.; Ruppin, E.; Herskovits, A.A. Integrative genomic analysis identifies isoleucine and CodY as regulators of Listeria monocytogenes virulence. PLoS Genet. 2012, 8, e1002887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Schaik, W.; Chateau, A.; Dillies, M.A.; Coppee, J.Y.; Sonenshein, A.L.; Fouet, A. The global regulator CodY regulates toxin gene expression in Bacillus anthracis and is required for full virulence. Infect. Immun. 2009, 77, 4437–4445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dineen, S.S.; Villapakkam, A.C.; Nordman, J.T.; Sonenshein, A.L. Repression of Clostridium difficile toxin gene expression by CodY. Mol. Microbiol. 2007, 66, 206–219. [Google Scholar] [CrossRef] [PubMed]
- Martin-Verstraete, I.; Peltier, J.; Dupuy, B. The regulatory networks that control Clostridium difficile toxin synthesis. Toxins 2016, 8, 153. [Google Scholar] [CrossRef] [PubMed]
- Daou, N.; Wang, Y.; Levdikov, V.M.; Nandakumar, M.; Livny, J.; Bouillaut, L.; Blagova, E.; Zhang, K.; Belitsky, B.R.; Rhee, K.; et al. Impact of CodY protein on metabolism, sporulation and virulence in Clostridioides difficile ribotype 027. PLoS ONE 2019, 14, e0206896. [Google Scholar] [CrossRef]
- Kovacs, A.T. Bacterial differentiation via gradual activation of global regulators. Curr. Genet. 2016, 62, 125–128. [Google Scholar] [CrossRef]
- Ravagnani, A.; Jennert, K.C.; Steiner, E.; Grunberg, R.; Jefferies, J.R.; Wilkinson, S.R.; Young, D.I.; Tidswell, E.C.; Brown, D.P.; Youngman, P.; et al. Spo0A directly controls the switch from acid to solvent production in solvent-forming clostridia. Mol. Microbiol. 2000, 37, 1172–1185. [Google Scholar] [CrossRef] [Green Version]
- Mascher, G.; Derman, Y.; Kirk, D.G.; Palonen, E.; Lindstrôm, M.; Korkeala, H. The CLO3403/CLO3404 two-component system of Clostridium botulinum E1 Beluga is important for cold shock response and growth at low temperatures. Appl. Environ. Microbiol. 2014, 80, 399–407. [Google Scholar] [CrossRef] [Green Version]
- Couesnon, A.; Raffestin, S.; Popoff, M.R. Expression of botulinum neurotoxins A and E, and associated non-toxin genes, during the transition phase and stability at high temperature: Analysis by quantitative reverse transcription-PCR. Microbiology 2006, 152, 759–770. [Google Scholar] [CrossRef] [Green Version]
- Anellis, A.; Berkowitz, D.; Kemper, D.; Rowley, D.B. Production of types A and B spores of Clostridium botulinum by the biphasic method: Effect on spore population, radiation resistance, and toxigenicity. Appl. Microbiol. 1972, 23, 734–739. [Google Scholar] [CrossRef] [Green Version]
- Mackin, K.E.; Carter, G.P.; Howarth, P.; Rood, J.I.; Lyras, D. Spo0A differentially regulates toxin production in evolutionarily diverse strains of Clostridium difficile. PLoS ONE 2013, 8, e79666. [Google Scholar] [CrossRef] [PubMed]
- Bouillaut, L.; Dubois, T.; Sonenshein, A.L.; Dupuy, B. Integration of metabolism and virulence in Clostridium difficile. Res. Microbiol. 2015, 166, 375–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philippe, V.A.; Mendez, M.B.; Huang, I.H.; Orsaria, L.M.; Sarker, M.R.; Grau, R.R. Inorganic phosphate induces spore morphogenesis and enterotoxin production in the intestinal pathogen Clostridium perfringens. Infect. Immun. 2006, 74, 3651–3656. [Google Scholar] [CrossRef] [Green Version]
- Paredes-Sabja, D.; Sarker, M.R. Clostridium perfringens sporulation and its relevance to pathogenesis. Future Microbiol. 2009, 4, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Kurt, A.; Ozkan, M.; Ozcengiz, G. Inorganic phosphate has a crucial effect on Cry3Aa delta-endotoxin production. Lett. Appl. Microbiol. 2005, 41, 303–308. [Google Scholar] [CrossRef]
- Chekabab, S.M.; Harel, J.; Dozois, C.M. Interplay between genetic regulation of phosphate homeostasis and bacterial virulence. Virulence 2014, 5, 786–793. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, S.; Somani, V.K.; Bhatnagar, R. Phosphate starvation enhances the pathogenesis of Bacillus anthracis. Int. J. Med. Microbiol. 2015, 305, 523–531. [Google Scholar] [CrossRef]
- Allenby, N.E.; Laing, E.; Bucca, G.; Kierzek, A.M.; Smith, C.P. Diverse control of metabolism and other cellular processes in Streptomyces coelicolor by the PhoP transcription factor: Genome-wide identification of in vivo targets. Nucleic Acids Res. 2012, 40, 9543–9556. [Google Scholar] [CrossRef]
- Baker, J.J.; Johnson, B.K.; Abramovitch, R.B. Slow growth of Mycobacterium tuberculosis at acidic pH is regulated by phoPR and host-associated carbon sources. Mol. Microbiol. 2014, 94, 56–69. [Google Scholar] [CrossRef] [Green Version]
- Lovenklev, M.; Artin, I.; Hagberg, O.; Borch, E.; Holst, E.; Radstrom, P. Quantitative interaction effects of carbon dioxide, sodium chloride, and sodium nitrite on neurotoxin gene expression in nonproteolytic Clostridium botulinum type B. Appl. Environ. Microbiol. 2004, 70, 2928–2934. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, S.; Burman, L.G.; Akerlund, T. Suppression of toxin production in Clostridium difficile VPI10463 by amino acids. Microbiology 1999, 145, 1683–1693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgess, C.; Gasse, F.; Steinglass, R.; Yakubu, A.; Raza, A.A.; Johansen, K. Eliminating maternal and neonatal tetanus and closing the immunity gap. Lancet 2017, 389, 1380–1381. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Recombinant Antisense mRNA Plasmid | Gene Bank Accession Number | Other Gene Name | Role | Familly (RR) | Genetic Environment | Homology | Homologs (RR) in Other Clostridia (Protein Identity > 60%) | Homolog TCS in C. botulinum strain Hall | Homolog TCS in C. botulinum Strain ATCC 3502 | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Old Locus Tag | Locus Tag | Homolog System (RR/SHK) | Protein Identity (RR) | Regulation of Botulism Neurotoxin | Homolog System (RR/SHK) | Protein Identity/Positive | |||||||
| chromosomal localization | |||||||||||||
| CTC_00189 | CTC_RS00820 | RR | OmpR | Periplasmic endopeptidase | ResD, respiration control | C. botulinum, C. cochlearium, C. tetanomorphum, C. scatologenes, C. drakei, C. magnum, C. sporogenes, C. tyrobutyricum, C. acetireducens, C. ljungdahlii, C. novyi, C. pasteurianum, | CLC_3521/CLC_3520 | 81% | None | CBO3543 | 81/87 | ||
| CTC_00191 | CTC_RS00825 | SHK | CBO3542 | 67/73 | |||||||||
| CTC_00392 | CTC_RS01985 | RR | OmpR | ABC Transporter | BacS/BacR regulation of resistance to bacitracin | C. botulinum, C. tetanomorphum, C. sporogenes, C. lundense, C. cavendishii, C. acetobutylicum, C. pasteurianum, C. ljungdahlii | CLC_0331/CLC_0332 | 90% | None | CBO0272 | 90/92 | ||
| CTC_00393 | CTC_RS01990 | SHK | CBO0273 | 86/89 | |||||||||
| CTC_00411 | CTC_RS02080 | phoP | RR | OmpR | hydroxylamne reductase | VirI/VirJ regulation of toxin synthesis | C. botulinum, C. perfringens, C. carboxidivorans, C. pasteurianum, C. lundense, C. novyi, C. Septicum | CLC_0661/CLC_0663 | 65% | Positive | CBO0607 | 64/72 | |
| p1308 | CTC_00412 | CTC_RS02085 | phoR | SHK | CBO0608 | 46/55 | |||||||
| CTC_00455 | CTC_RS02330 | RR | phosphomannomutase/ phosphoglucomutase | putative rRNA methylase | C. tetanomorphum, C. ljungdahlii, C. homopropionicum | CLC_0413 | 55% | unknown | CBO0355 | 54/59 | |||
| CTC_00456 | CTC_RS14405 | SHK | no | ||||||||||
| CTC_00597 | CTC_RS02990 | RR | OmpR | Glycerine-deshydrogenase | BacR regulation of resistance to bacitracin | C. lundense, C. tetanomorphum, C. ljungdahlii, C. argetinense, C. drakei | CLC_2212/CLC_2211 | 45% | None | several | 45/66 | ||
| CTC_00598 | CTC_RS02995 | SHK | no | ||||||||||
| CTC_00628 | CTC_RS03125 | spaR | RR | OmpR | ABC Transporter SpaEFG: export subtiline | SpaR/SpaK regulation of resistance to subtilin | C. botulinum, C. novyi, C. beijerinckii, C. saccharobutylicum, C. butyricum, C. sporogenes | CLC_1615/CLC_1616 | 65% | unknown | CBO1585 | 65/69 | |
| CTC_00629 | CTC_RS03130 | spaK | SHK | CBO1586 | 37/43 | ||||||||
| CTC_00805 | CTC_RS04010 | RR | OmpR | Protein transport | VanR/VanS regulation of resistance to vancomycin | C. botulinum, C. cochlearium, C. tetanomorphum, C. lundens, C. carboxidivorans, C. sporogenes, C. pasteurianum | CLC_0423/CLC_0424 | 76% | None | CBO0365 | 78/82 | ||
| CTC_00806 | CTC_RS04015 | SHK | CBO0366 | 44/50 | |||||||||
| CTC_00848 | CTC_RS04235 | RR | OmpR | Heat shock protein | GtcS/GtcR regulation of antibiotic synthesis (gramicidin) | C. lundense, C. tetanomorphum, C. amylolyticum, C. perfringens | CLC_1640/CLC_1639 | 48% | None | several | 47/55 | ||
| CTC_00849 | CTC_RS04240 | SHK | no | ||||||||||
| CTC_00872 | CTC_RS04365 | RR | OmpR | Efflux-ATPase Copper | Arc, regulation of aerobic/anaerobic respiration | C. botulinum, C. tetanomorphum, C. lundense, C. sporogenes, C. pasteurianum, C. beijerinckii,, C. ljungdahlii | CLC_1088/CLC_1089 | 68% | None | CBO1035 | 66/71 | ||
| CTC_00873 | CTC_RS04370 | SHK | CBO1036 | 55/63 | |||||||||
| CTC_00924 | CTC_RS04645 | SHK | PucR | Zn-dependent protease | regulation of purine catabolism, methyl-accepting chemotaxis protein | No | CBO1773 | 43/50 | |||||
| CTC_00925 | CTC_RS04650 | RR | no | ||||||||||
| p1309 | CTC_00934 | CTC_RS04705 | SHK | AraC | amihydrolase; Pyruvate formate lyase | LytS, autolysis regulation | C. botulinum B str. Eklund 17B, C. novyi | CLC_1627 | 41% | unknown | no | ||
| CTC_00935 | CTC_RS04710 | RR | no | ||||||||||
| CTC_00949 | CTC_RS04780 | virS | SHK | LytR/AlgR | transcriptional regulator, merR family; putative methyl-accepting chemotaxis protein | virulence regulation | C. butyricum, C. septicum | CLC_1105/CLC_1104 | 35% | None | no | ||
| p1310 | CTC_00950 | CTC_RS04785 | virR | RR | CBO1053 | 35/54 | |||||||
| CTC_01130 | CTC_RS05745 | RR | OmpR | ABC Transporteur: phosphates | PhoP/PhoR, regulation of phosphate uptake | C. lundense, C. carboxidivorans, C. ljungdahlii, C. botulinum, C. sporogenes, C. butyricum, C. acetobutylicum, C. butyricum, C. pasteurianum, C. neonatale, C. baratii | CLC_2386/CLC_2385 | 73% | None | CBO2527 | 73/87 | ||
| p1311 | CTC_01131 | CTC_RS05750 | phoR | SHK | CBO2526 | 52/75 | |||||||
| p1312 | CTC_01211 | CTC_RS06180 | tlpA | SHK | LysR | anaerobic sulfite reductase | LytR, autolysis regulation, methyl-accepting chemotaxis protein tlpA | C. cochlearium | CLC_3570 | 35% | unknown | CBO2828 | 36/61 |
| CTC_01212 | CTC_RS06185 | RR | several | 35/57 | |||||||||
| CTC_01420 | CTC_RS07310 | resE | SHK | OmpR | ABC Transporter | YycG/YycF, regulation of cell division | C. lundense, C. cellulovorans, C. amylolyticum, C. botulinum, | CLC_0842/CLC_0843 CB00786/CB00787 * | 58% | None Negative | CBO0787 | 47/65 | |
| p1419 | CTC_01421 | CTC_RS07315 | RR | CBO0786 | 58/76 | ||||||||
| CTC_01481 | CTC_RS07700 | SHK | OmpR | Conserved proteins with transmembrane helices and 4Fe4S motif | FeuQ/FeuP, regulation of iron acquisition | C. novyi, C. yurii | CLC_3521/CLC_3520 | 43% | None | no | |||
| CT_01482 | CTC_RS07705 | RR | several | 43/60 | |||||||||
| CTC_01490 | CTC_RS07755 | RR | OmpR | Heat shock protein HtpG (chaperonne); Membrane protein | unknown | C. lundense, C. tetanomorphum, C. kluyveri, C. pasteurianum, C. carboxidivorans, C. ljungdahlii, C. tyrobutyricum, C. acetobutylicum | CLC_0423/CLC_0424 | 44% | None | several | 44/64 | ||
| CTC_01491 | CTC_RS07760 | SHK | no | ||||||||||
| CTC_01523 | CTC_RS07895 | RR | NarL | Fumarate-reductase soluble flavoprotein | DcuR, regulation of fumarate anaerobic respiration through C4-dicarboxylates | C. cochlearium | CLC_0307/CLC_0306 | 48% | None | CBO0249 | 48/69 | ||
| CTC_01524 | CTC_RS07900 | dpiB | SHK | CBO0248 | 41/60 | ||||||||
| CTC_01804 | CTC_RS09305 | SHK | OmpR | ABC Transporter | BacS/BacR, AB-Bacitracine synthesis and regulation | C. tetanomorphum, C. lundense, C. novyi | CLC_2212/CLC_2211 | 56% | None | CBO2284 | 47/72 | ||
| CTC_01805 | CTC_RS09310 | RR | CBO2285 | 56/79 | |||||||||
| CTC_01818 | CTC_RS09380 | resE | SHK | OmpR | ABC Transporter; RNA polymerase sigma factor | unknown | C. tetanomorphum, C. lundense, C. ljungdahlii, C. cavendishii | CLC_0842/CLC_0843 CB00786/CB00787* | 51% | None Negative | CBO0787 | 38/63 | |
| CTC_01819 | CTC_RS09385 | RR | CBO0786 | 51/71 | |||||||||
| CTC_01848 | CTC_RS14320 | yesM | SHK | NarL/FixJ | Fumarate-reductase | unknown | C. cochlearium | CLC_2236/CLC_2235 | 26% | None | no | ||
| CTC_01849 | CTC_RS09510 | RR | no | ||||||||||
| CTC_01857 | CTC_RS09550 | SHK | XRE | Helicase, oleate hydratase | SinR regulation of entry in stationary phase, to nutrient depletion; Spo0A repressor | C. botulinum CDC_69094, C. magnum, C. beijerinckii | No | CBO0693 | 48/70 | ||||
| CTC_01858 | CTC_RS09555 | sinR | RR | no | |||||||||
| CTC_01905 | CTC_RS09790 | SHK | OmpR | ABC Transporter | BacR; VanR (synthèse et régulation AB) | C. indolis, C. methoxybenzovorans | CLC_0423/CLC_0424 | 45% | None | no | |||
| CTC_01906 | CTC_RS09795 | RR | CBO0365 | 45/66 | |||||||||
| CTC_01918 | CTC_RS09860 | resE | SHK | OmpR | ABC Transporter | BacR; VanR (synthèse et régulation AB) | C. kluyveri, C. uliginosum, C. puniceum, C. scatologenes, C. saccharobutylicum, C. oryzae, C. ljungdahlii, C. lundense, C. drakey, C. botulinum B2 331, C. sporogenes, C. acetobutylicum, C. butyricum | CLC_0842/CLC_0843 CB00786/CB00787* | 42% | None Negative | CBO0787 | 32/53 | |
| CTC_01919 | CTC_RS09865 | RR | CBO0786 | 42/60 | |||||||||
| p1313 | CTC_01951 | CTC_RS10030 | phoR | SHK | OmpR | Heavy metal translocating P-type aTPase | PhoP/PhoR regulation of phosphate uptake | C. cochlearium, C. lundense, C. tetanomorphum, C. intestinale, C. amylolyticum, C. baratii, C. botulinum, C. chauvoei, C. sporogenes, C. perfringens, C. ljungdahlii | CLC_0410/CLC_0411 | 68% | Positive | CBO0353 | 57/78 |
| CTC_01953 | CTC_RS10035 | phoP | RR | CBO0352 | 68/86 | ||||||||
| CTC_01978 | CTC_RS10150 | SHK | LytR/AlgR | carbon starvation protein A CstA | LytS/LytR autolysis regulation | C. cochlearium, C. tetanomorphum, C. lundense | CLC_3250/CLC_3251 | 55% | None | CBO3309 | 51/71 | ||
| p1314 | CTC_01979 | CTC_RS10155 | RR | CBO3308 | 55/78 | ||||||||
| CTC_02155 | CTC_RS11115 | SHK | OmpR | DNA mismatch repair protein hexA | VanS/VanR regulation of resistance to glycopeptides | C. carboxidivorans, C. lundense, C. amylolyticum, C. botulinum, C. oryzae, C. pasteurianum, C. sporogenes, C. novyi, C. neonatale | CLC_1640/CLC_1639 | 77% | None | CBO1612 | 48/68 | ||
| CTC_02156 | CTC_RS11120 | RR | CBO1613 | 77/86 | |||||||||
| CTC_02178 | CTC_RS11240 | SHK | Fis | ethanolamine utilization protein EutA, EutP | EutS/EutR regulation of éthanolamine utilization | C. argentinense, C. drakei, C. lundense, C. tetanomorphum | No | no | |||||
| CTC_02179 | CTC_RS11245 | RR | no | ||||||||||
| CTC_02322 | CTC_RS11915 | RR | Fis | sodium/glutamate symport carrier protein; V-Typ-ATPase-protein | AtoS/AtoC regulation of acetoacetate metabolism | C. cochlearium, C. lundense, C. tetanomorphum | CLC_1882 | 40% | unknown | several | 46/63 | ||
| CTC_02323 | CTC_RS11920 | SHK | no | ||||||||||
| plasmid localization | |||||||||||||
| p1307 | CTC_p22 | CTC_RS13810 | SHK | OmpR | ATP-binding protein | unknown | C. lundense | CLC_1431/CLC_1432 | 56% | None | CBO1395 | 39/60 | |
| CTC_p21 | CTC_RS13805 | RR | CBO1394 | 56/74 | |||||||||
| Isogenic Antisense Strains | Target Gene | S/R | Primer | Nucleotide Sequence (5′--> 3) | Product Length (bp) |
|---|---|---|---|---|---|
| CN655/1307 | CTC_p22 | S | P2020-F | CCGCTGCAGGATAATTTGGGAATGATTATTTTA | 228 |
| P2021-R | GGCCATGGTTAACATATCGTCCATACTC | ||||
| CN655/1308 | CTC_00412 | S | P2022-F | CCGCTGCAGGAGGTGATTGAAAAATAG | 208 |
| P2023-R | GGCCATGGTAAATCTAACATAGTAAATTTATAC | ||||
| CN655/1310 | CTC_00950 | R | P2024-F | CCGCTGCAGGGAGGGTTAAATTATGTATAATG | 236 |
| P2025-R | GGCCATGGGCTACTTCTATACCATTTATTTC | ||||
| CN655/1309 | CTC_00934 | S | P2026-F | CCGCTGCAGCAGGGGGTATTTTTGTGTTAAATAATAGG | 236 |
| P2027-R | GGCCATGGCATTGGCATCGCAACATATGCG | ||||
| CN655/1312 | CTC_01211 | S | P2028-F | CCGCTGCAGGGGGAGACAGTGGTGAAGTTGCG | 223 |
| P2029-R | GGCCATGGGGTTAAAAAATTTTCTTTTATATTTC | ||||
| CN655/1314 | CTC_01979 | R | P2030-F | CCGCTGCAGGAGATGAATTTATGAACAAAAT | 231 |
| P2031-R | GGCCATGGGCTAATTCCATGCCATTTTTAG | ||||
| CN655/1311 | CTC_01131 | S | P2032-F | CCGCTGCAGGGTGGTAAAATGAAAAAAAG | 245 |
| P2033-R | GGCCATGGCCTTATATCACTATCATTA | ||||
| CN655/1313 | CTC_01951 | S | P2034-F | CCGCTGCAGGGAAGGTAGAAAATGAAAAGTATAAAG | 243 |
| P2035-R | GGCCATGGCCACATTATCCATTATATTTTCTTC | ||||
| CN655/1419 | CTC_01421 | R | P2291-F | CCGCTGCAGGGGAGATTTTGTGAACAACATATT | 242 |
| P2292-R | GGCCATGGTCTGATGCCTTTCTTATTTCTTTAC | ||||
| CN655/1418 | CTC_01260 | CodY | P2289-F | CCGCTGCAGGAGGAGTTACAAATGTCATCATTATTA | 232 |
| P2290-R | GGCCATGGACTACCTTGTCTCTTACTGTCTG | ||||
| CN655/1472 | CTC_00222 | Spo0 | P2361-F | CCGCTGCAGGGAGGTATAAAATATATGATA | 225 |
| P2362-R | GGCCATGGTTATTACACTCTTTAAAGGTGAA | ||||
| CN655/1480 | CTC_00194 | mfd | P2359-F | CCGCTGCAGGAGGTGAATTTTATTATGAGAT | 236 |
| P2360-R | GGCCATGGAATATTTTTTGCTTCTATATCG |
| Target Gene | Primer | Nucleotide Sequence (5′-->3) | Product Length (bp) |
|---|---|---|---|
| qRT-PCR | |||
| tent | P1714-F | CCAAGGTGCACAAGGAATTT | 146 |
| P1715-R | CAATGTTTAATGCGGGTCCT | ||
| tetR | P1726-F | GTTGCTCAAATTATTTAAACTTCGAA | 115 |
| P1727-R | GCTATATCACATTCTTTCATATCTTCAAA | ||
| rpoB | P2142-F | TTGAAGAATGTAAAGAGAGAGATGCTAC | 118 |
| P2143-R | GGGAAGTCACCCATAAAGACA | ||
| gyrA | P2146-F | AAGATGATGTAGCAGTAAGTATGGA | 98 |
| P2147-R | CTCTGAAGCCAATGTCCTTTT | ||
| Recombinant protein expression | |||
| CTC_p21 | P2349-F | CGCCGCGGATCCATGTATAAGATATTGATTGTTGAA | 711 |
| P2350-R | CCGCCGGAATTCTTACACCTGAAATAAACGATAGCC | ||
| CTC_01979 | P2351-F | CGCCGCGGATCCATGAACAAAATAAATTGTGTAATAATA | 792 |
| P2352-R | CCGCCGGAATTCTTAAAAATCTAATATGTCCTTTAAGTG | ||
| CTC_01421 | P2353-F | CGCCGCGGATCCGTGAACAACATATTGTTAGTTGAA | 717 |
| P2354-R | CCGCCGGAATTCCTATTTATTAATTTCGTAGTTCCACCT | ||
| codY | P2355-F | CGCGGATCCATGTCATCATTATTAGAGAAG | 801 |
| P2356-R | CCGCCGGAATTCTTACTTAATTTTTTTCAATTCCTC | ||
| CTC_00935 | P2357-F | CGCGGATCCGTGTGTAGAGTAGTGCTT | 759 |
| P2358-R | CCGCCGGAATTCTTATACTTTTTTATTATTCAC | ||
| EMSA | |||
| Ptent | P2365-F | (5’-end labelled biotin) GGTGGCTCCATCATAATAATTGTAT | 359 |
| P2366-R | (5’-end labelled biotin) GGTTTTAGCATTAAAAAAATTAGAACCTA | ||
| PtetR | P2363-F | (5’-end labelled biotin) CAGTATTTTTGAAATGTATAATAATTACTTC | 316 |
| P2364-R | (5’-end labelled biotin) CGGTTCTCTTAATTTAGTAATATCAATAT | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chapeton-Montes, D.; Plourde, L.; Deneve, C.; Garnier, D.; Barbirato, F.; Colombié, V.; Demay, S.; Haustant, G.; Gorgette, O.; Schmitt, C.; et al. Tetanus Toxin Synthesis is Under the Control of A Complex Network of Regulatory Genes in Clostridium tetani. Toxins 2020, 12, 328. https://doi.org/10.3390/toxins12050328
Chapeton-Montes D, Plourde L, Deneve C, Garnier D, Barbirato F, Colombié V, Demay S, Haustant G, Gorgette O, Schmitt C, et al. Tetanus Toxin Synthesis is Under the Control of A Complex Network of Regulatory Genes in Clostridium tetani. Toxins. 2020; 12(5):328. https://doi.org/10.3390/toxins12050328
Chicago/Turabian StyleChapeton-Montes, Diana, Lucile Plourde, Cecile Deneve, Dominique Garnier, Fabien Barbirato, Vincent Colombié, Sandy Demay, Georges Haustant, Olivier Gorgette, Christine Schmitt, and et al. 2020. "Tetanus Toxin Synthesis is Under the Control of A Complex Network of Regulatory Genes in Clostridium tetani" Toxins 12, no. 5: 328. https://doi.org/10.3390/toxins12050328
APA StyleChapeton-Montes, D., Plourde, L., Deneve, C., Garnier, D., Barbirato, F., Colombié, V., Demay, S., Haustant, G., Gorgette, O., Schmitt, C., Thouvenot, C., Brüggemann, H., & Popoff, M. R. (2020). Tetanus Toxin Synthesis is Under the Control of A Complex Network of Regulatory Genes in Clostridium tetani. Toxins, 12(5), 328. https://doi.org/10.3390/toxins12050328