The Cadherin Protein Is Not Involved in Susceptibility to Bacillus thuringiensis Cry1Ab or Cry1Fa Toxins in Spodoptera frugiperda


 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
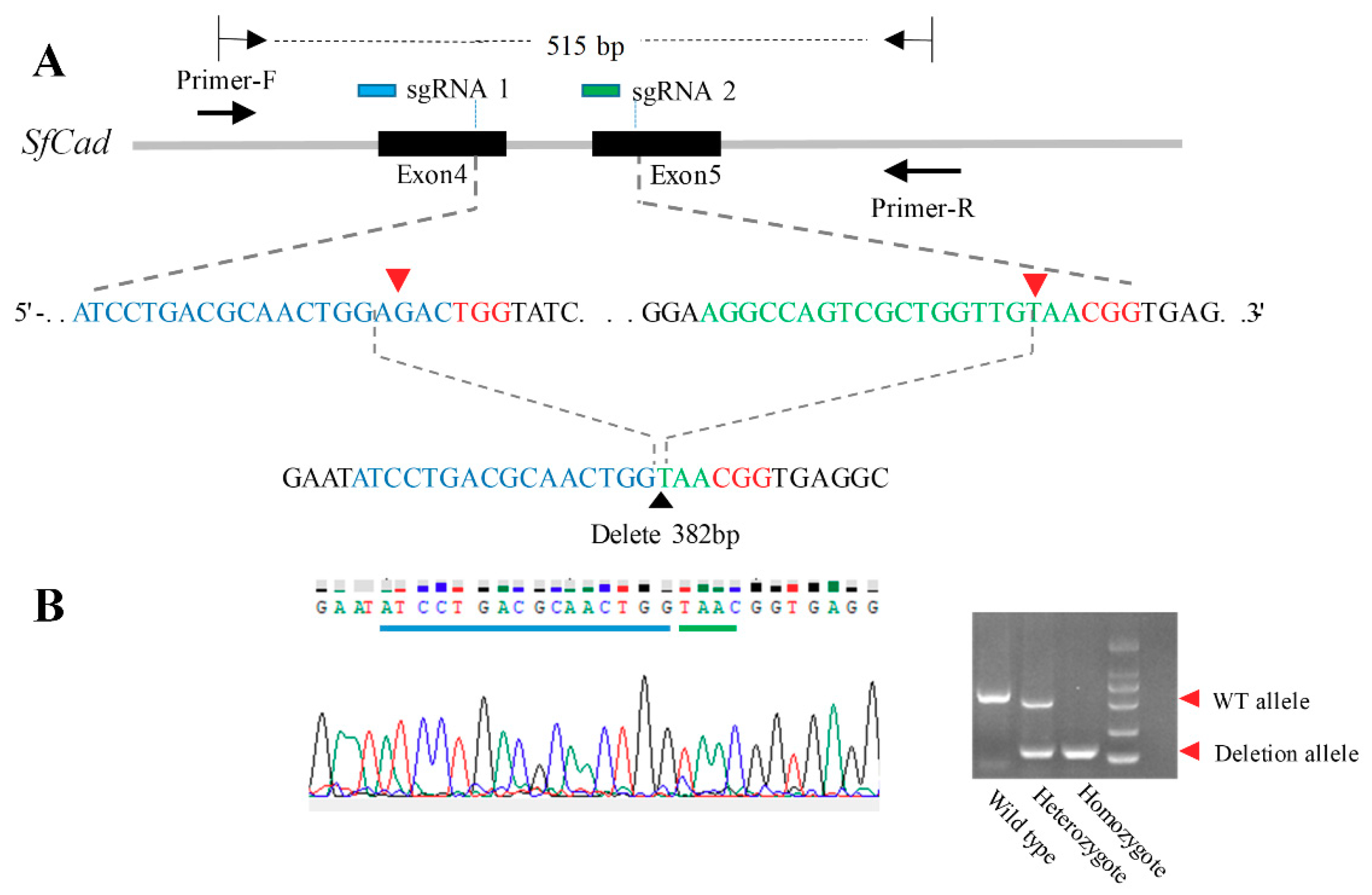
2.1. Construction of SfCad Gene Deleted Mutant by CRISPR/Cas 9 Genome Editing
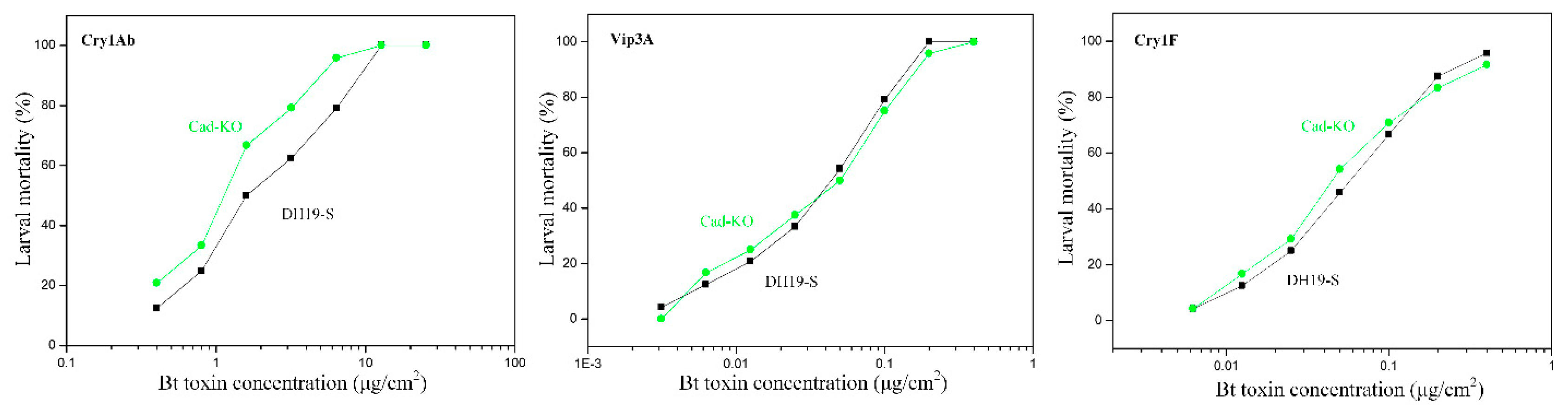
2.2. Susceptibility of Cad-KO Strain to the Bt Toxins
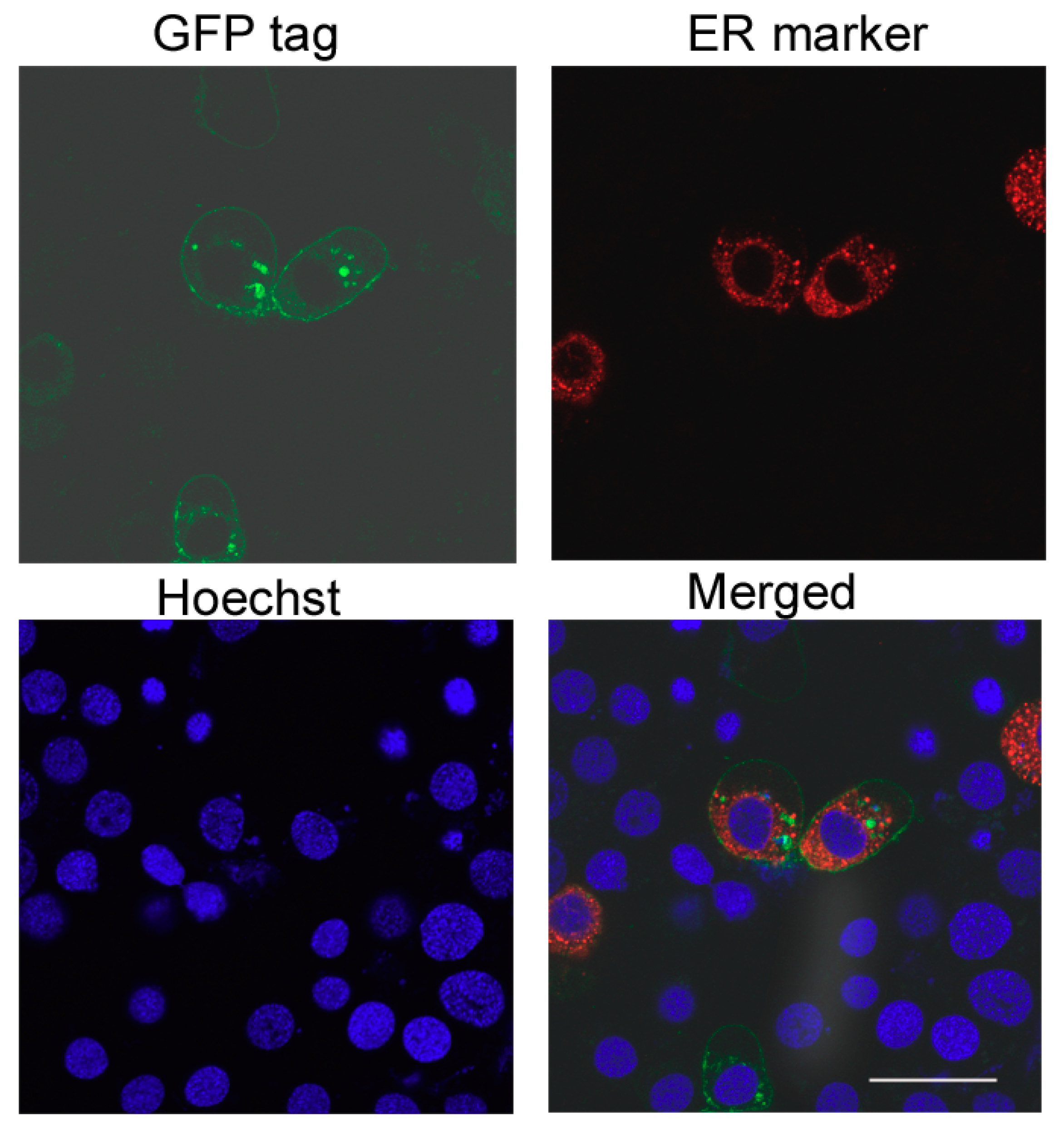
2.3. SfCAD Expression Did Not Increase Susceptibility of Hi5 Insect Cells to Cry1Ab or Cry1Fa Toxins
2.4. Cry1Ac, Cry1Ab, and Cry1Fa Did Not Bind to the Toxin-Binding Region (TBR) of SfCad
3. Discussion
4. Materials and Methods
4.1. S. frugiperda Strain and Insect Cell Cultures
4.2. Preparation of sgRNAs
4.3. Cas 9 Protein
4.4. Egg Collection and Microinjection
4.5. Identification of SfCad Mutations Mediated by CRISPR/Cas 9 System
4.6. Bt Toxins and Bioassay
4.7. Plasmids, Transfection, and Fluorescence Observation
4.8. Cytotoxicity Assay
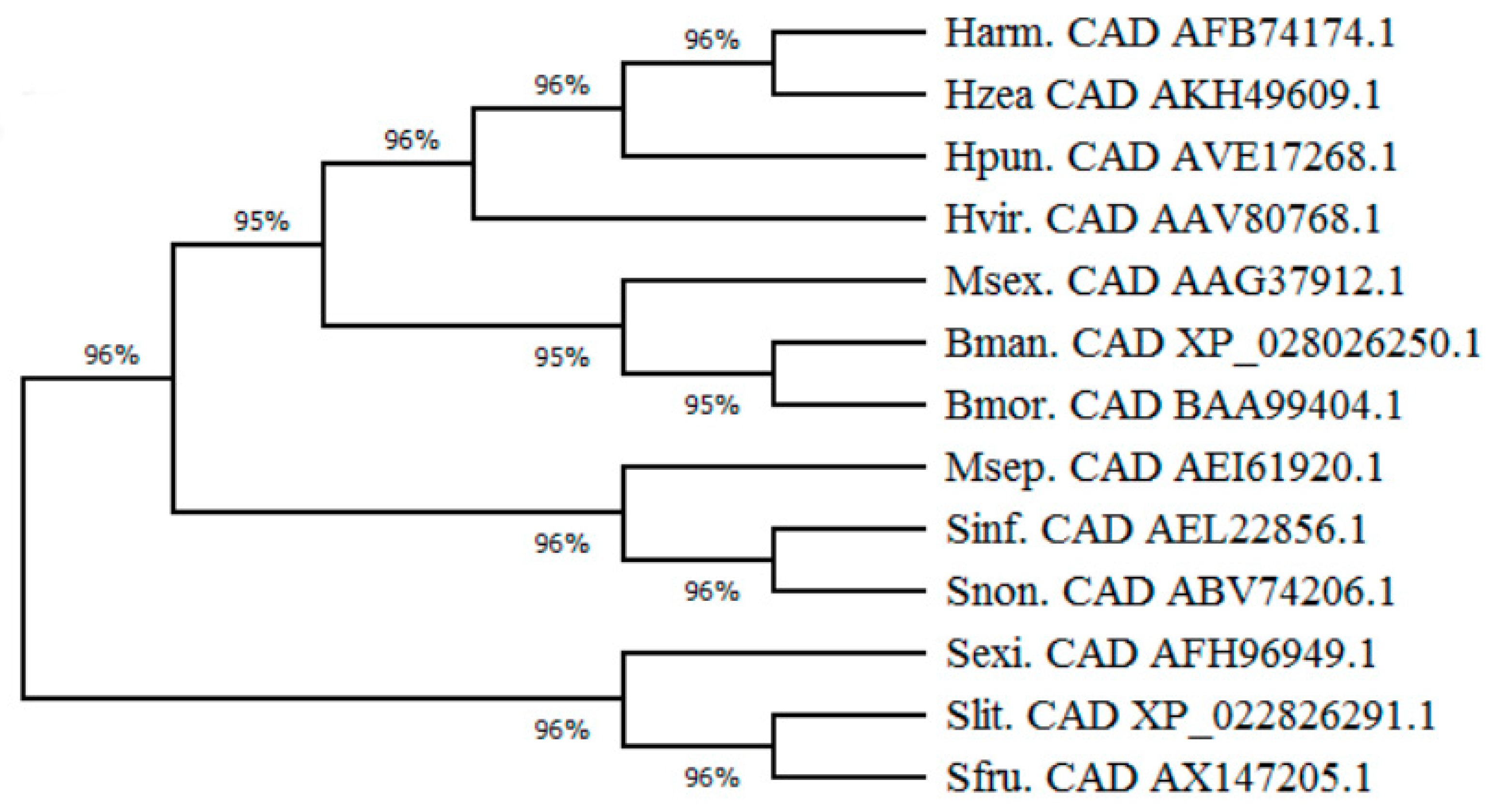
4.9. Construction of a Lepidoptera Cadherin Phylogenetic Tree
4.10. Purification of Proteins Expressed in Bacteria
4.11. Ligand Blot Assays
Author Contributions
Funding
Conflicts of Interest
References
- Wu, K.M.; Lu, Y.H.; Feng, H.Q.; Jiang, Y.Y.; Zhao, J.Z. Suppression of cotton bollworm in multiple crops in China in areas with Bt toxin–containing cotton. Science 2008, 321, 1676–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, S.S.; Cowles, E.A.; Pietrantonio, P.V. The mode of action of Bacillus thuringiensis endotoxins. Annu. Rev. Entomol. 1992, 37, 615–636. [Google Scholar] [CrossRef] [PubMed]
- Bravo, A.; Likitvivatanavong, S.; Gill, S.S.; Soberón, M. Bacillus thuringiensis: A story of a successful bioinsecticide. Insect Biochem. Mol. Biol. 2011, 41, 423–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estruch, J.J.; Warren, G.W.; Mullins, M.A.; Nye, G.J.; Craig, J.A.; Koziel, M.G. Vip3A, a novel Bacillus thuringiensis vegetative insecticidal protein with a wide spectrum of activities against lepidopteran insects. Proc. Natl. Acad. Sci. USA 1996, 93, 5389–5394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.K.; Walters, F.S.; Hart, H.; Palekar, N.; Chen, J.S. The mode of action of the Bacillus thuringiensis vegetative insecticidal protein Vip3A differs from that of Cry1Ab delta-endotoxin. Appl. Environ. Microbiol. 2003, 69, 4648–4657. [Google Scholar] [CrossRef] [Green Version]
- Jiang, K.; Hou, X.; Han, L.; Tan, T.; Cao, Z.; Cai, J. Fibroblast growth factor receptor, a novel receptor for vegetative insecticidal protein Vip3Aa. Toxins 2018, 10, 546. [Google Scholar] [CrossRef] [Green Version]
- Jiang, K.; Hou, X.Y.; Tan, T.T.; Cao, Z.L.; Mei, S.Q.; Yan, B.; Chang, J.; Han, L.; Zhao, D.; Cai, J. Scavenger receptor-C acts as a receptor for Bacillus thuringiensis vegetative insecticidal protein Vip3Aa and mediates the internalization of Vip3Aa via endocytosis. PLoS Pathog. 2018, 14. [Google Scholar] [CrossRef]
- Gahan, L.J.; Gould, F.; Heckel, D.G. Identification of a gene associated with Bt resistance in Heliothis virescens. Science 2001, 293, 857–860. [Google Scholar] [CrossRef]
- Morin, S.; Biggs, R.W.; Sisterson, M.S.; Shriver, L.; Ellers-Kirk, C.; Higginson, D.; Holley, D.; Gahan, L.J.; Heckel, D.G.; Carrière, Y.; et al. Three cadherin alleles associated with resistance to Bacillus thuringiensis in pink bollworm. Proc. Natl. Acad. Sci. USA 2003, 100, 5004–5009. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Yang, Y.; Gao, W.; Guo, J.; Wu, Y.; Wu, Y. Introgression of a disrupted cadherin gene enables susceptible Helicoverpa armigera to obtain resistance to Bacillus thuringiensis toxin Cry1Ac. Bull. Entomol. Res. 2009, 99, 175–181. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, H.; Wang, H.; Zhao, S.; Zuo, Y.; Yang, Y.; Wu, Y. Functional validation of cadherin as a receptor of Bt toxin Cry1Ac in Helicoverpa armigera utilizing the CRISPR/Cas9 system. Insect Biochem. Mol. Biol. 2016, 76, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, J.; Ma, Y.; Wan, P.; Liu, K.; Cong, S.; Xiao, Y.; Xu, D.; Wu, K.; Fabrick, J.A.; et al. Transposon insertion causes cadherin mis-splicing and confers resistance to Bt cotton in pink bollworm from China. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wu, K.; Liang, G.; Guo, Y. Gene cloning and expression of cadherin in midgut of Helicoverpa armigera and its Cry1A binding region. Sci. China C Life Sci. 2005, 48, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhang, J.; Xiao, Y.; Yang, Y.; Liu, C.; Peng, R.; Yang, Y.; Bravo, A.; Soberón, M.; Liu, K. The cadherin Cry1Ac binding-region is necessary for the cooperative effect with ABCC2 transporter enhancing insecticidal activity of Bacillus thuringiensis Cry1Ac toxin. Toxins 2019, 11. [Google Scholar] [CrossRef] [Green Version]
- Fabrick, J.A.; Mathew, L.G.; LeRoy, D.M.; Hull, J.J.; Unnithan, G.C.; Yelich, A.J.; Carrière, Y.; Li, X.; Tabashnik, B.E. Reduced cadherin expression associated with resistance to Bt toxin Cry1Ac in pink bollworm. Pest Manag. Sci. 2020, 76, 67–74. [Google Scholar] [CrossRef]
- Brestschneider, A.; Heckel, D.G.; Pauchet, Y. Three toxins, two receptors, one mechanism: Mode of action of Cry1A toxins from Bacillus thuringiensis in Heliothis virescens. Insect Biochem. Mol. Biol. 2016, 76, 109–117. [Google Scholar] [CrossRef]
- Guo, Z.; Kang, S.; Zhu, X.; Wu, Q.; Wang, S.; Xie, W.; Zhang, Y. The midgut cadherin-like gene is not associated with resistance to Bacillus thuringiensis toxin Cry1Ac in Plutella xylostella (L.). J. Invertebr. Pathol. 2015, 126, 21–30. [Google Scholar] [CrossRef]
- Park, Y.; Herrero, S.; Kim, Y. A single type of cadherin is involved in Bacillus thuringiensis toxicity in Plutella xylostella. Insect Mol. Biol. 2015, 24, 624–633. [Google Scholar] [CrossRef]
- Gao, M.; Dong, S.; Hu, X.; Zhang, X.; Liu, Y.; Zhong, J.; Lu, L.; Wang, Y.; Chen, L.; Liu, X. Roles of midgut cadherin from two moths in different Bacillus thuringiensis action mechanisms: Correlation among toxin binding, cellular toxicity, and synergism. J. Agric. Food Chem. 2019, 67, 13237–13246. [Google Scholar] [CrossRef]
- Hu, X.; Zhang, X.; Zhong, J.; Liu, Y.; Zhang, C.; Xie, Y.; Lin, M.; Xu, C.; Lu, L.; Zhu, Q.; et al. Expression of Cry1Ac toxin-binding region in Plutella xylostella cadherin-like receptor and studying their interaction mode by molecular docking and site-directed mutagenesis. Int. J. Biol. Macromol. 2018, 111, 822–831. [Google Scholar] [CrossRef]
- Ingber, D.A.; Mason, C.E.; Flexner, L. Cry1 Bt susceptibilities of fall armyworm (Lepidoptera: Noctuidae) host strains. J. Econ. Entomol. 2018, 111, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Morsello, S.; Head, G.P.; Sansone, C.; Huang, F.; Gilreath, R.T.; Kerns, D.L. F2 screen, inheritance and cross-resistance of field-derived Vip3A resistance in Spodoptera frugiperda (Lepidoptera: Noctuidae) collected from Louisiana, USA. Pest Manag. Sci. 2018, 74, 1769–1778. [Google Scholar] [CrossRef] [PubMed]
- Vélez, A.M.; Spencer, T.A.; Alves, A.P.; Moellenbeck, D.; Meagher, R.L.; Chirakkal, H.; Siegfried, B.D. Inheritance of Cry1F resistance, cross-resistance and frequency of resistant alleles in Spodoptera frugiperda (Lepidoptera: Noctuidae). Bull. Entomol. Res. 2013, 103, 700–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tay, W.T.; Mahon, R.J.; Heckel, D.G.; Walsh, T.K.; Downes, S.; James, W.J.; Lee, S.F.; Reineke, A.; Williams, A.K.; Gordon, K.H. Insect resistance to Bacillus thuringiensis toxin Cry2Ab is conferred by mutations in an ABC transporter subfamily A protein. PLoS Genet. 2015, 11. [Google Scholar] [CrossRef] [PubMed]
- Storer, N.P.; Babcock, J.M.; Schlenz, M.; Meade, T.; Thompson, G.D.; Bing, J.W.; Huckaba, R.M. Discovery and characterization of field resistance to Bt maize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico. J. Econ. Entomol. 2010, 103, 1031–1038. [Google Scholar] [CrossRef]
- Monnerat, R.; Martins, E.; Macedo, C.; Queiroz, P.; Praça, L.; Soares, C.M.; Moreira, H.; Grisi, I.; Silva, J.; Soberón, M.; et al. Evidence of field-evolved resistance of Spodoptera frugiperda to Bt corn expressing Cry1F in Brazil that is still sensitive to modified Bt toxins. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, R.; Hasler, J.; Meagher, R.; Nagoshi, R.; Hietala, L.; Huang, F.; Narva, K.; Jurat-Fuentes, J.L. Mechanism and DNA/based detection of field-evolved resistance to transgenic Bt corn in fall armyworm (Spodoptera frugiperda). Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Boaventura, D.; Ulrich, J.; Lueke, B.; Bolzan, A.; Okuma, D.; Gutbrod, O.; Geibel, S.; Zeng, Q.; Dourado, P.M.; Martinelli, S.; et al. Molecular characterization of Cry1F resistance in fall armyworm, Spodoptera frugiperda from Brazil. Insect Biochem. Mol. Biol. 2020, 116. [Google Scholar] [CrossRef]
- Bernardi, O.; Bernardi, D.; Horikoshi, R.J.; Okuma, D.M.; Miraldo, L.L.; Fatoretto, J.; Medeiros, F.C.; Burd, T.; Omoto, C. Selection and characterization of resistance to the Vip3Aa20 protein from Bacillus thuringiensis in Spodoptera frugiperda. Pest Manag. Sci. 2016, 72, 1794–1802. [Google Scholar] [CrossRef]
- Kahn, T.W.; Chakroun, M.; Williams, J.; Walsh, T.; James, B.; Monserrate, J.; Ferré, J. Efficacy and resistance management potential of a modified Vip3C protein for control of Spodoptera frugiperda in Maize. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Flagel, L.; Lee, Y.W.; Wanjugi, H.; Swarup, S.; Brown, A.; Wang, J.; Kraft, E.; Greenplate, J.; Simmons, J.; Adams, N.; et al. Mutational disruption of the ABCC2 gene in fall armyworm, Spodoptera frugiperda, confers resistance to the Cry1Fa and Cry1A.105 insecticidal proteins. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, I.; Ocelotl, J.; Jorge Sánchez, J.; Lima, C.; Martins, E.; Rosales-Juárez, A.; Aguilar-Medel, S.; Abad, A.; Dong, H.; Monnerat, R.; et al. Enhancement of Bacillus thuringiensis Cry1Ab and Cry1Fa toxicity to Spodoptera frugiperda by domain III mutations indicates two limiting steps in toxicity as defined by receptor binding and protein stability. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Chen, Z.; Yang, Y.; Xiao, Y.; Liu, C.; Ma, Y.; Soberón, M.; Bravo, A.; Yang, Y.; Liu, K. A single amino acid polymorphism in ABCC2 loop 1 is responsible for differential toxicity of Bacillus thuringiensis Cry1Ac toxin in different Spodoptera (Noctuidae) species. Insect Biochem. Mol. Biol. 2018, 100, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yu, L.; Wu, Y. Disruption of a cadherin gene associated with resistance to Cry1Ac endotoxin of Bacillus thuringiensis in Helicoverpa armigera. Appl. Environ. Microbiol. 2005, 71, 948–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, L.; Hou, L.; Zhang, B.; Liu, L.; Li, B.; Deng, P.; Ma, W.; Wang, X.; Fabrick, J.A.; Chen, L.; et al. Cadherin is involved in the action of Bacillus thuringiensis toxins Cry1Ac and Cry2Aa in the beet armyworm, Spodoptera exigua. J. Invertebr. Pathol. 2015, 127, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Adegawa, S.; Nakama, Y.; Endo, H.; Shinkawa, N.; Kikuta, S.; Sato, R. The domain II loops of Bacillus thuringiensis Cry1Aa form an overlapping interaction site for two Bombyx mori larvae functional receptors, ABC transporter C2 and cadherin-like receptor. Biochim. Biophys. Acta Proteins Proteom. 2017, 1865, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Endo, H.; Adegawa, S.; Kikuta, S.; Sato, R. The intracellular region of silkworm cadherin-like protein is not necessary to mediate the toxicity of Bacillus thuringiensis Cry1Aa and Cry1Ab toxins. Insect Biochem. Mol. Biol. 2018, 94, 36–41. [Google Scholar] [CrossRef]
- Wang, S.; Kain, W.; Wang, P. Bacillus thuringiensis Cry1A toxins exert toxicity by multiple pathways in insects. Insect Biochem. Mol. Biol. 2018, 102, 59–66. [Google Scholar] [CrossRef]
- Jurat-Fuentes, J.L.; Adang, M.J. The Heliothis virescens cadherin protein expressed in Drosophila S2 cells functions as receptor for Bacillus thuringiensis Cry1A but not Cry1Fa toxins. Biochemistry 2006, 45, 9688–9695. [Google Scholar] [CrossRef]
- Badran, A.H.; Guzov, V.M.; Huai, Q.; Kemp, M.M.; Vishwanath, P.; Kain, W.; Nance, A.M.; Evdokimov, A.; Moshiri, F.; Turner, K.H.; et al. Continuous evolution of Bacillus thuringiensis toxins overcomes insect resistance. Nature 2016, 533, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Davis, T.R.; Wickham, T.J.; McKenna, K.A.; Granados, R.R.; Shuler, M.L.; Wood, H.A. Comparative recombinant production of eight insect lines. In Vitro Cell. Dev. Biol. Anim. 1993, 29A, 388–390. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Shen, B.; Zhang, C.; Huang, X.; Zhang, Y. sgRNAcas9: A software package for designing CRISPR sgRNA and evaluating potential off-target cleavage sites. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Tao, J.; Li, Q.; Cheng, Y.; Sun, X.; Wu, K.; Xiao, Y. Genome editing of the SfABCC2 gene confers resistance to Cry1F toxin from Bacillus thuringiensis in Spodoptera frugiperda. J. Integr. Agric. 2019, 18, 2–7. [Google Scholar]
- Li, J.; Ma, Y.; Yuan, W.; Xiao, Y.; Liu, C.; Wang, J.; Peng, J.; Peng, R.; Soberón, M.; Bravo, A.; et al. FOXA transcriptional factor modulates insect susceptibility to Bacillus thuringiensis Cry1Ac toxin by regulating the expression of toxin-receptor ABCC2 and ABCC3 genes. Insect Biochem. Mol. Biol. 2017, 88, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Cushing, N.L.; Johnson, M.W. Diamondback moth (Lepidoptera: Plutellidae) resistance to insecticides in Hawaii: Intra-island variation and cross-resistance. J. Econ. Entomol. 1987, 80, 1091–1099. [Google Scholar] [CrossRef]
- Xie, R.; Zhuang, M.; Ross, L.S.; Gomez, I.; Oltean, D.I.; Bravo, A.; Soberón, M.; Gill, S.S. Single amino acid mutations in the cadherin receptor from Heliothis virescens affect its toxin binding ability to Cry1A toxins. J. Biol. Chem. 2005, 280, 8416–8425. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain. | Cry1Ab LC50 in μg/cm2 (95% of FL) | Cry1Fa LC50 in μg/cm2 (95% of FL) | Vip3Aa LC50 in μg/cm2 (95% of FL) |
|---|---|---|---|
| Cad-KO | 1.103 (0.798–1.453) | 0.05 (0.037–0.069) | 0.035 (0.026–0.047) |
| DH19-S | 1.797 (1.311–2.458) | 0.054 (0.041–0.073) | 0.033 (0.025–0.044) |
| Toxin | Putative Receptor | EC50 (µg/mL) | 95% FL | Slope ± SE | χ2 | df |
|---|---|---|---|---|---|---|
| Cry1Ab | SfCAD-GFP | — * | — | — | — | — |
| Cry1Ab | SfABCC2-GFP | 0.06a ** | 0.04–0.08 | 1.55 ± 0.08 | 7.20 | 3 |
| Cry1Fa | SfCAD-GFP | — | — | — | — | — |
| Cry1Fa | SfABCC2-GFP | 0.23b | 0.19–0.27 | 2.18 ± 0.09 | 6.77 | 3 |
| Fragment | Forward Primer (5′–3′) | Reverse Primer (5′–3′) | Vector |
|---|---|---|---|
| His-HaTBR | CCGGAATTCTACGATTCGTGCTACGGACGGT | CCCAAGCTTCAGGTACACCTTCACTTCCGT-3 | pET22b (Novagen, Madison, WI, USA) |
| His-SeTBR | CCGGAATTCTGTTATCCGAGCTACTGATGG | CCCAAGCTTCATGAAGATTGTCACTTCAGCTCGATC | pET22b |
| His-SlTBR | CCGGAATTCTGTTATTCGTGCCACGGATGGT | CCCAAGCTTCATGTAGATTATAACTTTTGCTCG | pET22b |
| His-SfTBR | CCGGAATTCTGGAGGCGGTGGAGGCGGTGTTATTCGGGCCACGGACGGCG | CCCAAGCTTCATGTAAATTGACACTTTTGCTCGATCACTCGC | pET22b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Jin, M.; Yang, Y.; Liu, L.; Yang, Y.; Gómez, I.; Bravo, A.; Soberón, M.; Xiao, Y.; Liu, K. The Cadherin Protein Is Not Involved in Susceptibility to Bacillus thuringiensis Cry1Ab or Cry1Fa Toxins in Spodoptera frugiperda. Toxins 2020, 12, 375. https://doi.org/10.3390/toxins12060375
Zhang J, Jin M, Yang Y, Liu L, Yang Y, Gómez I, Bravo A, Soberón M, Xiao Y, Liu K. The Cadherin Protein Is Not Involved in Susceptibility to Bacillus thuringiensis Cry1Ab or Cry1Fa Toxins in Spodoptera frugiperda. Toxins. 2020; 12(6):375. https://doi.org/10.3390/toxins12060375
Chicago/Turabian StyleZhang, Jianfeng, Minghui Jin, Yanchao Yang, Leilei Liu, Yongbo Yang, Isabel Gómez, Alejandra Bravo, Mario Soberón, Yutao Xiao, and Kaiyu Liu. 2020. "The Cadherin Protein Is Not Involved in Susceptibility to Bacillus thuringiensis Cry1Ab or Cry1Fa Toxins in Spodoptera frugiperda" Toxins 12, no. 6: 375. https://doi.org/10.3390/toxins12060375
APA StyleZhang, J., Jin, M., Yang, Y., Liu, L., Yang, Y., Gómez, I., Bravo, A., Soberón, M., Xiao, Y., & Liu, K. (2020). The Cadherin Protein Is Not Involved in Susceptibility to Bacillus thuringiensis Cry1Ab or Cry1Fa Toxins in Spodoptera frugiperda. Toxins, 12(6), 375. https://doi.org/10.3390/toxins12060375