Effect of Temperature during Drying and Storage of Dried Figs on Growth, Gene Expression and Aflatoxin Production

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
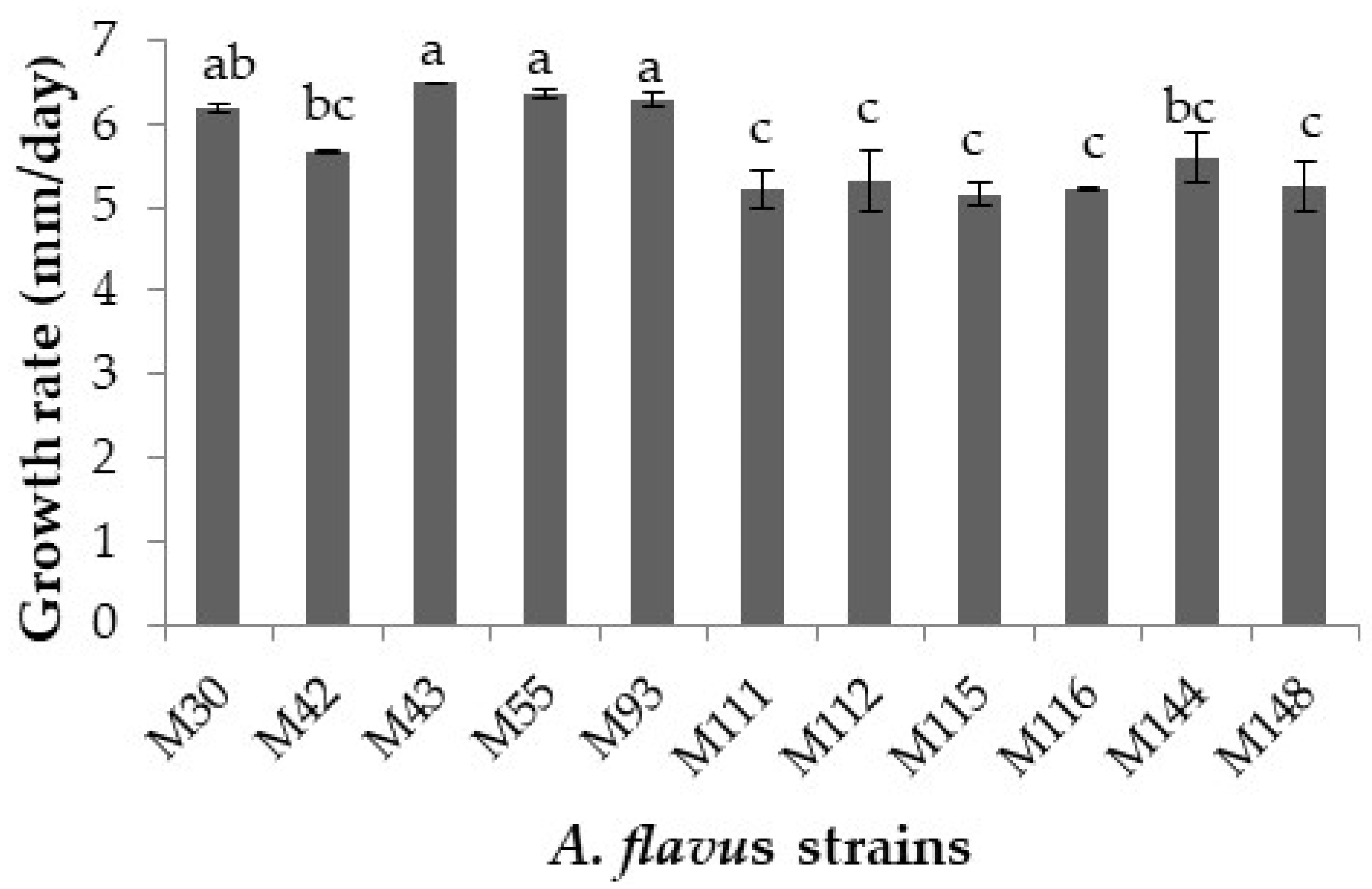
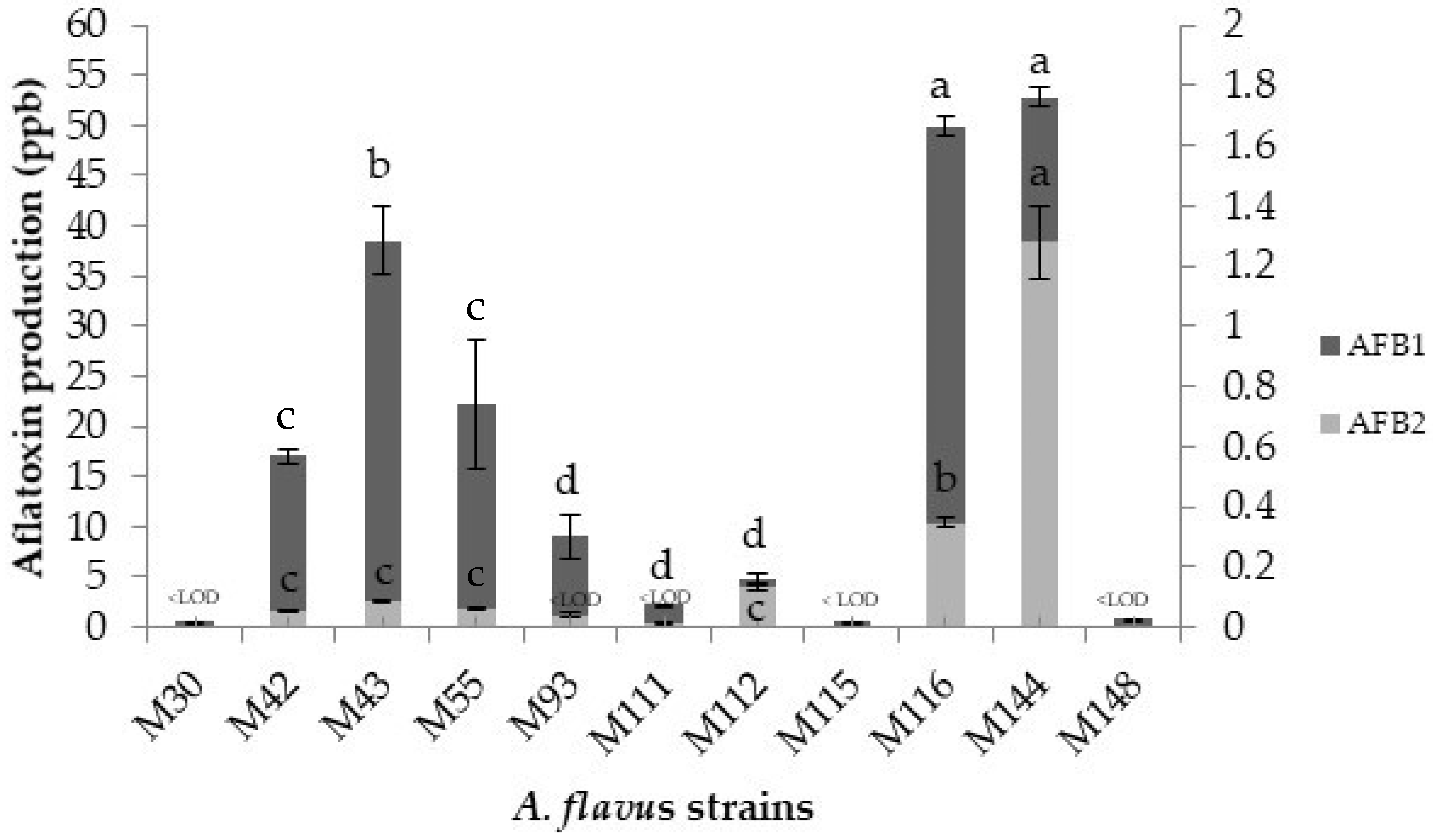
2.1. Selection of Two Aflatoxigenic Strains: Initial Screening
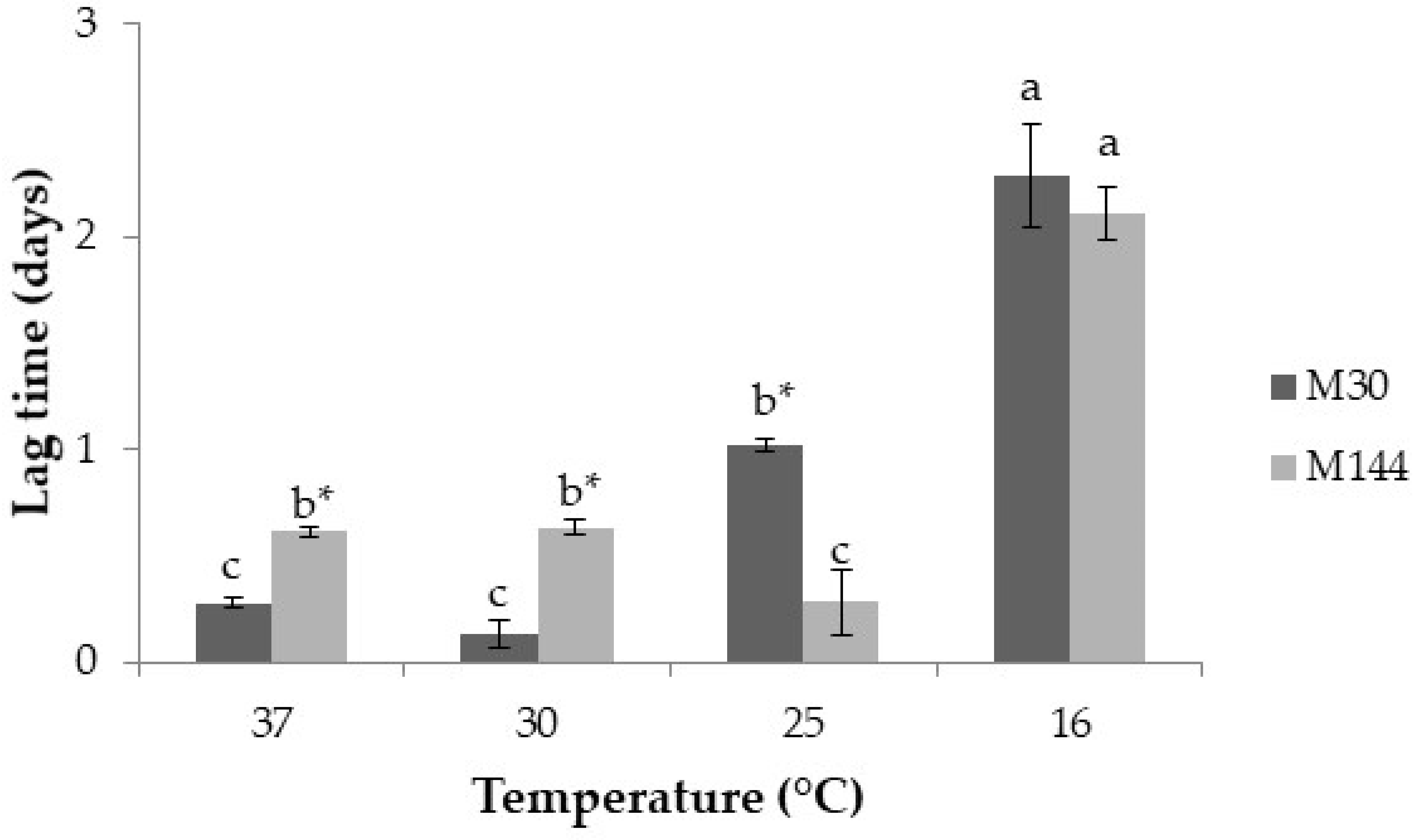
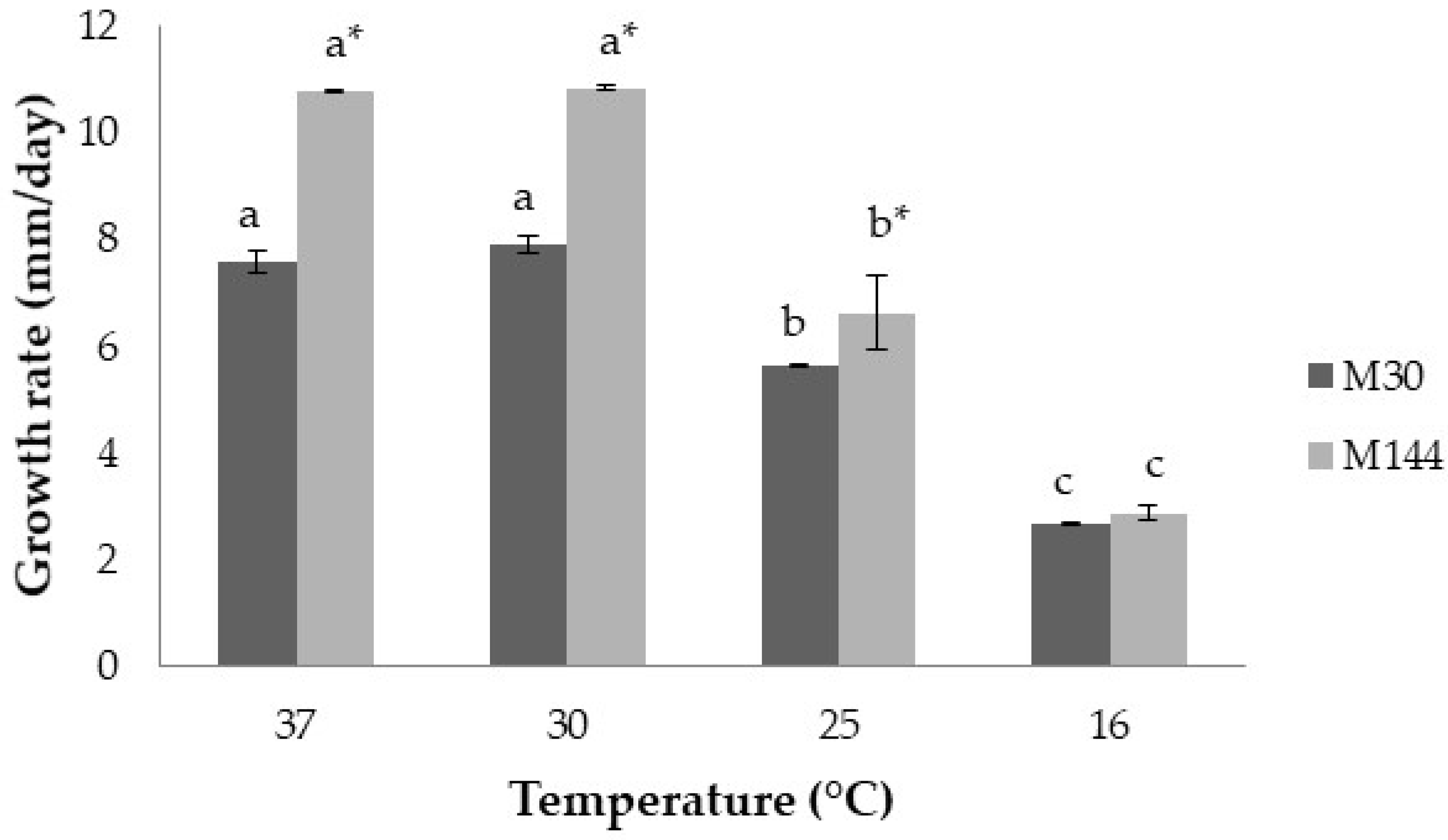
2.2. Effect of Temperature on Lag Times, Growth Rates, Mycotoxin Production and Aflatoxin-Related Gene Expression
2.2.1. Lag Times Prior to Growth
2.2.2. Growth
2.2.3. Aflatoxin Production
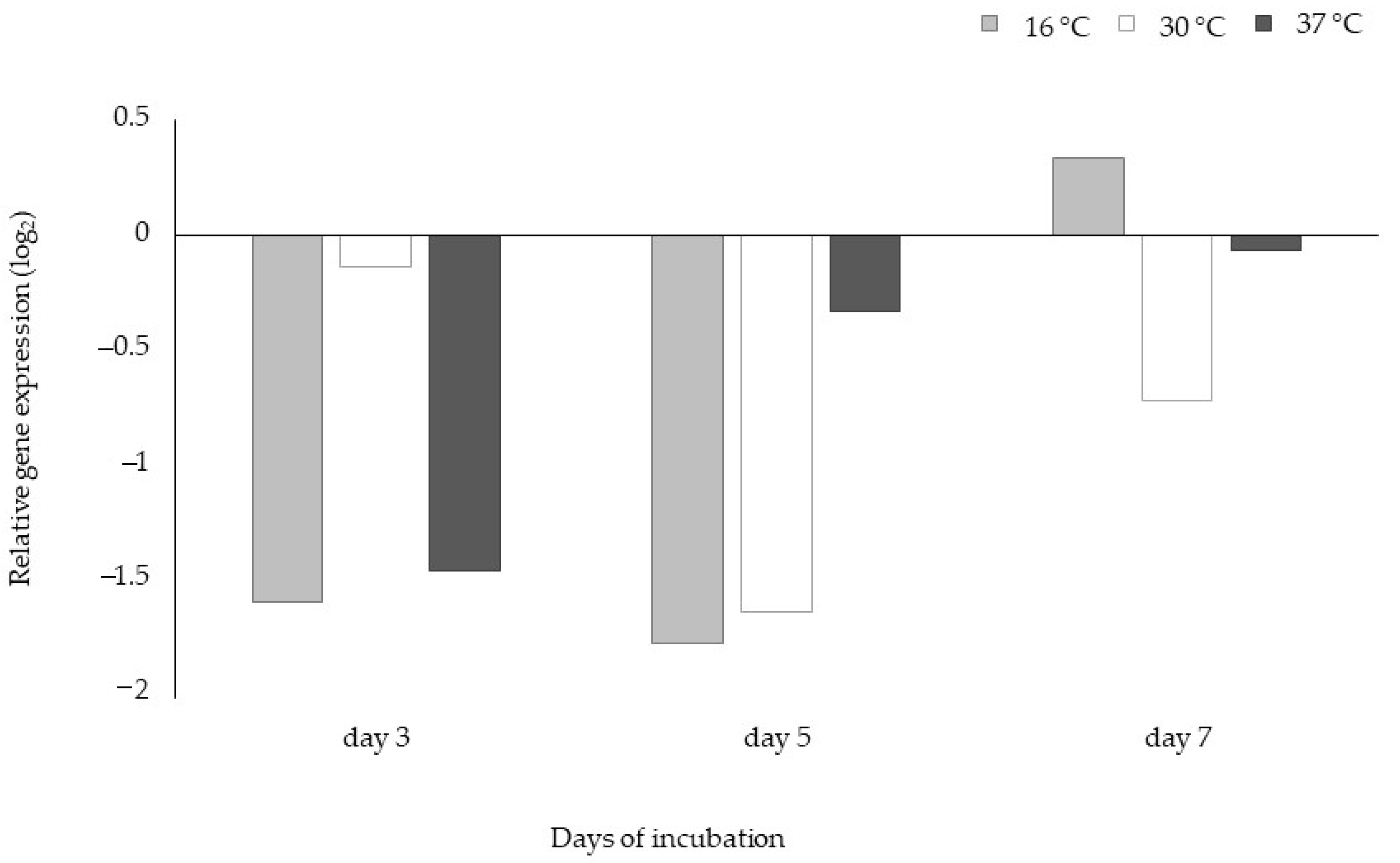
2.2.4. Gene Expression Studies
3. Discussion
4. Conclusions
5. Material and Methods
5.1. Mould Strains
5.2. Culture Medium Preparation
5.3. Inoculum, Inoculation, and Experimental Settings
5.4. Lag Time Prior to Growth and Growth Assessment
5.5. Gene Expression Analysis
5.5.1. Sampling and Sample Preparation
5.5.2. RNA Extraction
5.5.3. RT-qPCR Reactions and Relative Quantification
- Primers
- 2.
- cDNA synthesis
- 3.
- Real-time PCR reactions
- 4.
- Relative gene expression
5.6. Mycotoxin Analysis
5.6.1. Sampling and Sample Preparation
5.6.2. Aflatoxin Extraction and Quantification
5.7. Statistical Análisis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arvaniti, O.S.; Samaras, Y.; Gatidou, G.; Thomaidis, N.S.; Stasinakis, A.S. Review on fresh and dried figs: Chemical analysis and occurrence of phytochemical compounds, antioxidant capacity and health effects. Food Res. Int. 2019, 119, 244–267. [Google Scholar] [CrossRef]
- Ouchemoukh, S.; Hachoud, S.; Boudraham, H.; Mokrani, A.; Louaileche, H. Antioxidant activities of some dried fruits consumed in Algeria. LWT 2012, 49, 329–332. [Google Scholar] [CrossRef]
- Vallejo, F.; Marín, J.G.; Tomás-Barberán, F.A. Phenolic compound content of fresh and dried figs (Ficus carica L.). Food Chem. 2012, 130, 485–492. [Google Scholar] [CrossRef]
- FAOSTAT. Available online: http://www.fao.org/faostat/es/ (accessed on 20 December 2020).
- Encuesta sobre Superficies y Rendimientos Cultivos (ESYRCE). Available online: https://www.mapa.gob.es/es/estadistica/temas/estadisticas-agrarias/agricultura/esyrce/ (accessed on 20 December 2020).
- Mat Desa, W.N.; Mohammad, M.; Fudholi, A. Review of drying technology of fig. Trends Food Sci. Technol. 2019, 88, 93–103. [Google Scholar] [CrossRef]
- Gilbert, J.; Senyuva, H. Fungal and mycotoxin contamination of dried figs-a review. Mycotoxins 2008, 58, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Nuroğlu, E.; Öz, E.; Bakırdere, S.; Bursalıoğlu, E.O.; Kavanoz, H.B.; İçelli, O. Evaluation of magnetic field assisted sun drying of food samples on drying time and mycotoxin production. Innov. Food Sci. Emerg. Technol. 2019, 52, 237–243. [Google Scholar] [CrossRef]
- Trucksess, M.W.; Scott, P.M. Mycotoxins in botanicals and dried fruits: A review. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2008, 25, 181–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turkoz Bakirci, G. Investigation of aflatoxins levels in commercial dried figs from western Turkey. Int. Food Res. J. 2020, 27, 245–251. [Google Scholar]
- Dilek Heperkan, Z.; Moretti, A.; Daskaya Dikmen, C.; Heperkan, D.; Moretti, A.; Dikmen, C.D.; Logrieco, A.F. Toxigenic fungi and mycotoxin associated with figs in the Mediterranean area mycotoxins and biosynthetic genes View project prevention of Cronobacter sakazakii in baby foods View project Toxigenic fungi and mycotoxin associated with figs in the Mediterran. Phytopathol. Mediterr. 2012, 51, 119–130. [Google Scholar]
- Javanmard, M. Occurrence of Mould Counts and Aspergillus Species in Iranian Dried Figs at Different Stages of Production and Processing. J. Agric. Sci. Technol. 2010, 12, 331–338. [Google Scholar]
- López, P.; Venema, D.; Mol, H.; Spanjer, M.; de Stoppelaar, J.; Pfeiffer, E.; de Nijs, M. Alternaria toxins and conjugates in selected foods in the Netherlands. Food Control 2016, 69, 153–159. [Google Scholar] [CrossRef]
- López, P.; Venema, D.; de Rijk, T.; de Kok, A.; Scholten, J.M.; Mol, H.G.J.; de Nijs, M. Occurrence of Alternaria toxins in food products in The Netherlands. Food Control 2016, 60, 196–204. [Google Scholar] [CrossRef]
- Aldars-García, L.; Marín, S.; Sanchis, V.; Magan, N.; Medina, A. Assessment of intraspecies variability in fungal growth initiation of Aspergillus flavus and aflatoxin B1 production under static and changing temperature levels using different initial conidial inoculum levels. Int. J. Food Microbiol. 2018, 272, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Hadi, A.; Schmidt-Heydt, M.; Parra, R.; Geisen, R.; Magan, N. A systems approach to model the relationship between aflatoxin gene cluster expression, environmental factors, growth and toxin production by Aspergillus flavus. J. R. Soc. Interface 2012, 9, 757–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, P.C.; Sylla, A.; Gong, Y.Y.; Diallo, M.S.; Sutcliffe, A.E.; Hall, A.J.; Wild, C.P. Reduction in exposure to carcinogenic aflatoxins by postharvest intervention measures in west Africa: A community-based intervention study. Lancet 2005, 365, 1950–1956. [Google Scholar] [CrossRef]
- Devreese, M.; De Baere, S.; De Backer, P.; Croubels, S. Quantitative determination of the Fusarium mycotoxins beauvericin, enniatin A, A1, B and B1 in pig plasma using high performance liquid chromatography–tandem mass spectrometry. Talanta 2013, 106, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Food and Feed Safety Alerts (RASFF). Available online: https://ec.europa.eu/food/safety/rasff_ (accessed on 20 December 2020).
- Zohri, A.A.; Abdel-Gawad, K.M. Survey of mycoflora and mycotoxins of some dried fruits in Egypt. J. Basic Microbiol. 1993, 33, 279–288. [Google Scholar] [CrossRef]
- Wang, Y.J.; Nie, J.Y.; Yan, Z.; Li, Z.X.; Cheng, Y.; Farooq, S. Multi-mycotoxin exposure and risk assessments for Chinese consumption of nuts and dried fruits. J. Integr. Agric. 2018, 17, 1676–1690. [Google Scholar] [CrossRef]
- Ioannou-Kakouri, E.; Aletrari, M.; Christou, E.; Ralli, A.; Koliou, A. Occurrence of mycotoxins in local food in Cyprus. COST Action 2001, 835, 13–18. [Google Scholar]
- International Agency for Research on Cancer (IARC). Available online: https://www.iarc.fr/ (accessed on 20 December 2020).
- Diario Oficial de la Unión Europea. Reglamento (UE) No 1058/2012 de la Comisión. Available online: https://www.boe.es/doue/2012/313/L00014-00015.pdf (accessed on 20 December 2020).
- Drusch, S.; Aumann, J. Mycotoxins in Fruits: Microbiology, Occurrence, and Changes during Fruit Processing. Adv. Food Nutr. Res. 2005, 50, 33–78. [Google Scholar] [CrossRef]
- Molyneux, R.J.; Mahoney, N.; Kim, J.H.; Campbell, B.C. Mycotoxins in edible tree nuts. Int. J. Food Microbiol. 2007, 119, 72–78. [Google Scholar] [CrossRef]
- Özlüoymak, Ö.B. Development of a UV-based imaging system for Real-Time detection and separation of dried figs contaminated with aflatoxins. J. Agric. Sci. 2014, 20, 302–316. [Google Scholar] [CrossRef]
- Ait Mimoune, N.; Arroyo-Manzanares, N.; Gámiz-Gracia, L.; García-Campaña, A.M.; Bouti, K.; Sabaou, N.; Riba, A. Aspergillus section Flavi and aflatoxins in dried figs and nuts in Algeria. Food Addit. Contam. Part B Surveill. 2018, 11, 119–125. [Google Scholar] [CrossRef]
- Peromingo, B.; Rodríguez, A.; Bernáldez, V.; Delgado, J.; Rodríguez, M. Effect of temperature and water activity on growth and aflatoxin production by Aspergillus flavus and Aspergillus parasiticus on cured meat model systems. Meat Sci. 2016, 122, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Casquete, R.; Benito, M.J.; Córdoba, M.D.G.; Ruiz-Moyano, S.; Martín, A. The growth and aflatoxin production of Aspergillus flavus strains on a cheese model system are influenced by physicochemical factors. J. Dairy Sci. 2017, 100, 6987–6996. [Google Scholar] [CrossRef]
- Bernáldez, V.; Córdoba, J.J.; Magan, N.; Peromingo, B.; Rodríguez, A. The influence of ecophysiological factors on growth, aflR gene expression and aflatoxin B1 production by a type strain of Aspergillus flavus. LWT Food Sci. Technol. 2017, 83, 283–291. [Google Scholar] [CrossRef] [Green Version]
- Schmidt-Heydt, M.; Abdel-Hadi, A.; Magan, N.; Geisen, R. Complex regulation of the aflatoxin biosynthesis gene cluster of Aspergillus flavus in relation to various combinations of water activity and temperature. Int. J. Food Microbiol. 2009, 135, 231–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbas, H.K.; Wikinson, J.R.; Zablotowicz, R.M.; Accinelli, C.; Abel, C.A.; Bruns, H.A.; Weaver, M.A. Ecology of Aspergillus flavus, regulation of aatoxin production, and management strategies to reduce aatoxin contamination of corn. Toxin Rev. 2009, 28, 142–153. [Google Scholar] [CrossRef]
- Mousa, W.; Ghazali, F.M.; Jinap, S.; Ghazali, H.M.; Radu, S. Modelling the effect of water activity and temperature on growth rate and aflatoxin production by two isolates of Aspergillus flavus on paddy. J. Appl. Microbiol. 2011, 111, 1262–1274. [Google Scholar] [CrossRef]
- Mohale, S.; Magan, N.; Medina, A. Comparison of growth, nutritional utilisation patterns, and niche overlap indices of toxigenic and atoxigenic Aspergillus flavus strains. Fungal Biol. 2013, 117, 650–659. [Google Scholar] [CrossRef]
- Pitt, J.I.; Miscamble, B.F. Water Relations of Aspergillus flavus and Closely Related Species. J. Food Prot. 1995, 58, 86–90. [Google Scholar] [CrossRef]
- Sanchis, V.; Magan, N. Environmental conditions affecting mycotoxins. Mycotoxins Food 2004, 174–189. [Google Scholar] [CrossRef]
- Geisen, R.; Touhami, N.; Schmidt-Heydt, M. Mycotoxins as adaptation factors to food related environments. Curr. Opin. Food Sci. 2017, 17, 1–8. [Google Scholar] [CrossRef]
- Delgado, J.; Rodríguez, A.; García, A.; Núñez, F.; Asensio, M. Inhibitory Effect of PgAFP and Protective Cultures on Aspergillus parasiticus Growth and Aflatoxins Production on Dry-Fermented Sausage and Cheese. Microorganisms 2018, 6, 69. [Google Scholar] [CrossRef] [Green Version]
- Gourama, H.; Bullerman, L.B. Aspergillus flavus and Aspergillus parasiticus: Aflatoxigenic Fungi of Concern in Foods and Feeds. J Food Prot. 1995, 58, 1395–1404. [Google Scholar] [CrossRef]
- Martín Castaño, S.; Medina, A.; Magan, N. Comparison of dry matter losses and aflatoxin B1 contamination of paddy and brown rice stored naturally or after inoculation with Aspergillus flavus at different environmental conditions. J. Stored Prod. Res. 2017, 73, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Chang, P.K. The Aspergillus parasiticus protein AFLJ interacts with the aflatoxin pathway-specific regulator AFLR. Mol. Genet. Genom. 2003, 268, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Peromingo, B.; Rodríguez, M.; Delgado, J.; Andrade, M.J.; Rodríguez, A. Gene expression as a good indicator of aflatoxin contamination in dry-cured ham. Food Microbiol. 2017, 67, 31–40. [Google Scholar] [CrossRef]
- Accinelli, C.; Abbas, H.K.; Zablotowicz, R.M.; Wikinson, J.R. Aspergillus flavus aflatoxin occurrence and expression of aflatoxin biosynthesis genes in soil. Can J. Microbiol. 2008, 54, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Moyano, S.; Benito, M.J.; Martín, A.; Aranda, E.; Hernández, A.; Córdoba, M.G. Characterization of molds isolated from smoked paprika by PCR-RFLP and micellar electrokinetic capillary electrophoresis. Food Microbiol. 2009, 26, 776–782. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.J.; Taylor, J. 38-Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Rodríguez, A.; Medina, Á.; Córdoba, J.J.; Magan, N. The influence of salt (NaCl) on ochratoxin A biosynthetic genes, growth and ochratoxin A production by three strains of Penicillium nordicum on a dry-cured ham-based medium. Int. J. Food Microbiol. 2014, 178, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.; Rodríguez, M.; Luque, M.I.; Martín, A.; Córdoba, J.J. Real-time PCR assays for detection and quantification of aflatoxin producing molds in foods. Food Microbiol. 2012, 31, 89–99. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aflatoxin | Days of Incubation | 37 °C | 30 °C | 25 °C | 16 °C |
|---|---|---|---|---|---|
| B1 | 12 | <LOD 2 | 0.25 ± 0.35a3 | 60.63 ± 7.70a1 | 10.15 ± 1.56a2 |
| 7 | <LOD | 0.03 ± 0.01c3 | 58.39 ± 1.93a1 | 0.10 ± 0.07b2 | |
| 5 | <LOD | 0.12 ± 0.04b3 | 2.68 ± 0.51b1 | 0.03 ± 0.01b2 | |
| 3 | <LOD | 0.02 ± 0.01c2 | 1.26 ± 0.83b1 | <LOD | |
| B2 | 12 | <LOD | <LOD | 0.02 ± 0.01b | <LOD |
| 7 | 0.10 ± 0.01a2 | 0.06 ± 0.00 2 | 0.15 ± 0.06a1 | 0.13 ± 0.01 1 | |
| 5 | 0.02 ± 0.01b | <LOD | <LOD | <LOD | |
| 3 | <LOD | <LOD | <LOD | <LOD |
| Isolate Code | Geographical Area | Origin 1 | Moisture Content (%) |
|---|---|---|---|
| A. flavus M30 | South of Extremadura | Field | 16.78 |
| A. flavus M42 | South of Extremadura | Field | 16.78 |
| A. flavus M43 | South of Extremadura | Field | 16.39 |
| A. flavus M55 | South of Extremadura | Field | 16.78 |
| A. flavus M93 | South of Extremadura | Field | 20.46 |
| A. flavus M111 | South of Extremadura | Field | 20.46 |
| A. flavus M112 | South of Extremadura | Field | 19.01 |
| A. flavus M115 | South of Extremadura | Industry | 27.62 |
| A. flavus M116 | South of Extremadura | Industry | 27.62 |
| A. flavus M144 | North of Extremadura | Field | 36.20 |
| A. flavus M148 | South of Extremadura | Field | 16.39 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galván, A.I.; Rodríguez, A.; Martín, A.; Serradilla, M.J.; Martínez-Dorado, A.; Córdoba, M.d.G. Effect of Temperature during Drying and Storage of Dried Figs on Growth, Gene Expression and Aflatoxin Production. Toxins 2021, 13, 134. https://doi.org/10.3390/toxins13020134
Galván AI, Rodríguez A, Martín A, Serradilla MJ, Martínez-Dorado A, Córdoba MdG. Effect of Temperature during Drying and Storage of Dried Figs on Growth, Gene Expression and Aflatoxin Production. Toxins. 2021; 13(2):134. https://doi.org/10.3390/toxins13020134
Chicago/Turabian StyleGalván, Ana Isabel, Alicia Rodríguez, Alberto Martín, Manuel Joaquín Serradilla, Ana Martínez-Dorado, and María de Guía Córdoba. 2021. "Effect of Temperature during Drying and Storage of Dried Figs on Growth, Gene Expression and Aflatoxin Production" Toxins 13, no. 2: 134. https://doi.org/10.3390/toxins13020134
APA StyleGalván, A. I., Rodríguez, A., Martín, A., Serradilla, M. J., Martínez-Dorado, A., & Córdoba, M. d. G. (2021). Effect of Temperature during Drying and Storage of Dried Figs on Growth, Gene Expression and Aflatoxin Production. Toxins, 13(2), 134. https://doi.org/10.3390/toxins13020134