Adaptation of Staphylococcus aureus in a Medium Mimicking a Diabetic Foot Environment

 , , , and
, , , and 
Abstract
:1. Introduction
2. Results
2.1. Phenotypic Effects on S. aureus after a Prolonged Culture in a Medium Mimicking a DFU Environment
2.1.1. Addition of 10% Glucose
2.1.2. Addition of Antibiotics
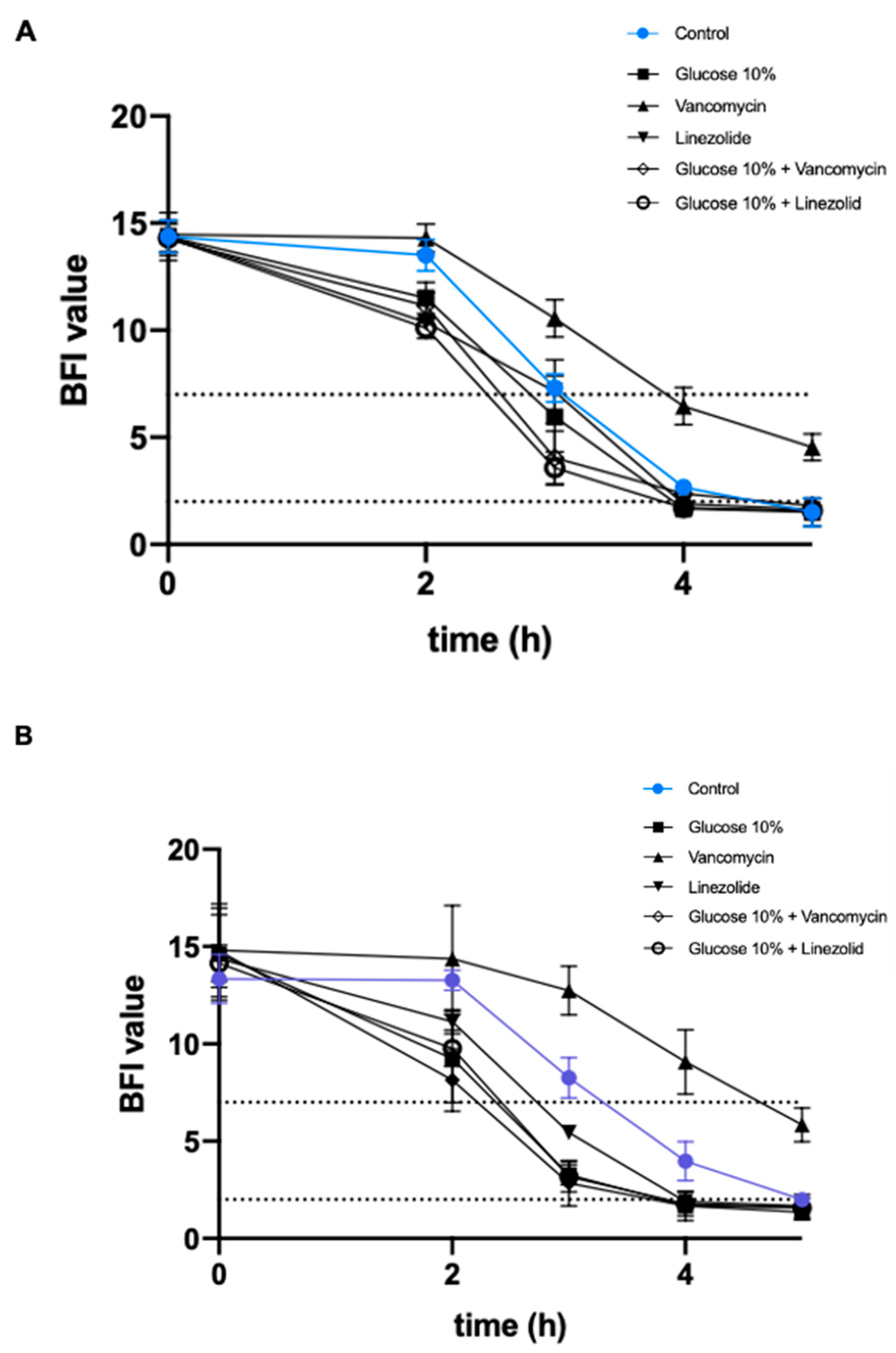
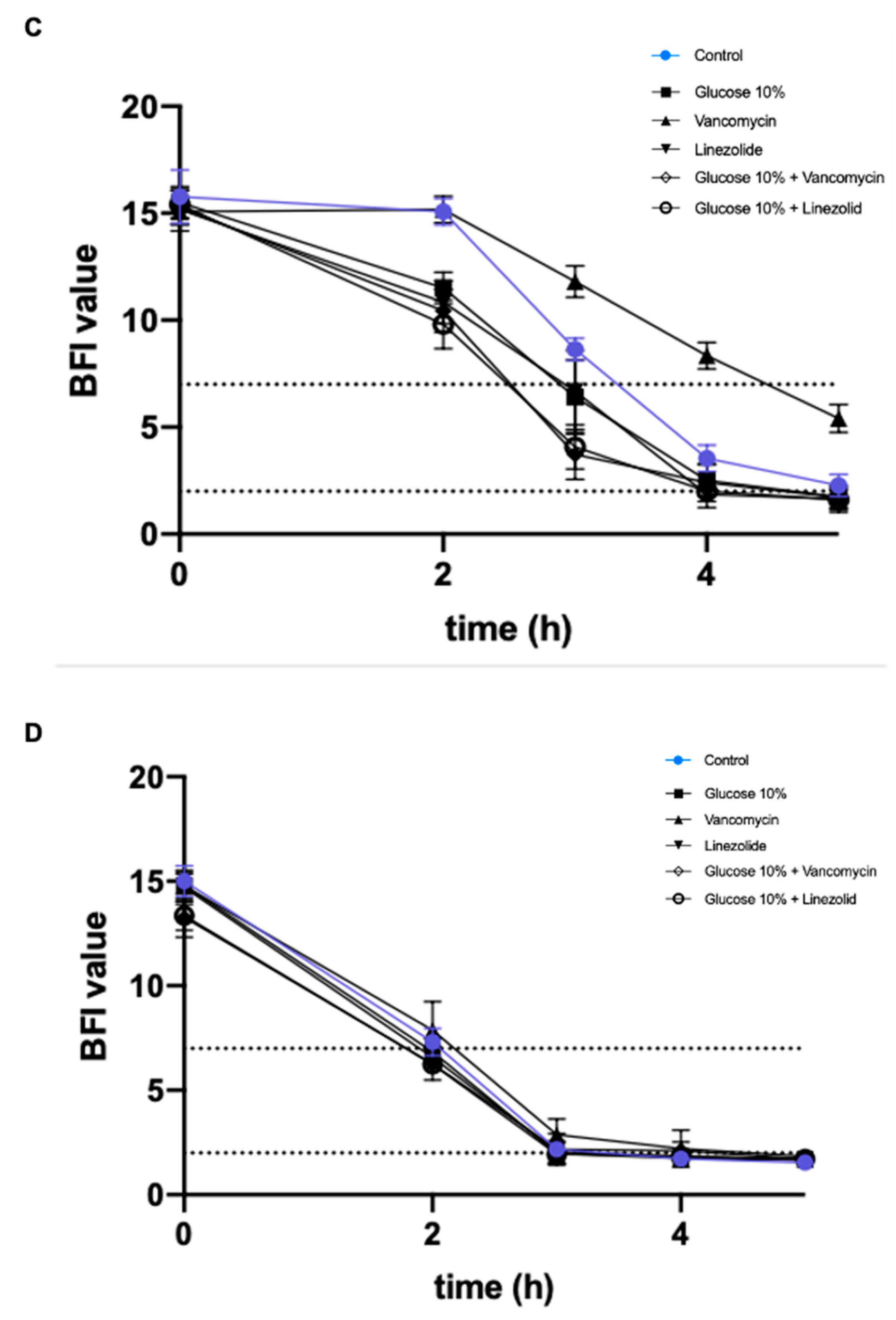
2.2. Effect of a Prolonged Culture in a Medium Mimicking DFU Environment on S. aureus Virulence
2.2.1. Addition of 10% Glucose
2.2.2. Addition of Antibiotics
2.3. Effect of a Prolonged Culture in a Medium Mimicking DFU Environment on Kinetics of S. aureus Biofilm Formation
2.3.1. Addition of 10% Glucose
2.3.2. Addition of Antibiotics
2.4. Effect of a Prolonged Culture in a Medium Mimicking DFU Environment on S. aureus Genes’ Expression
2.4.1. Addition of 10% Glucose
2.4.2. Addition of Antibiotics
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Bacterial Strains and Growth Conditions
5.2. Nematode Killing Assay
5.3. Feeding Behavior Assays
5.4. Biofilm Formation
5.5. qRT-PCR Assays
5.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Armstrong, D.G.; Boulton, A.J.; Bus, S.A. Diabetic foot ulcers and their recurrence. N. Engl. J. Med. 2017, 376, 2367–2375. [Google Scholar] [CrossRef]
- Lecube, A.; Pachón, G.; Petriz, J.; Hernández, C.; Simó, R. Phagocytic activity is impaired in type 2 diabetes mellitus and in-creases after metabolic improvement. PLoS ONE 2011, 6, e23366. [Google Scholar] [CrossRef]
- Tesfaye, S.; Boulton, A.J.; Dyck, P.J.; Freeman, R.; Horowitz, M.; Kempler, P.; Lauria, G.; Malik, R.A.; Spallone, V.; Vinik, A.; et al. Diabetic neuropathies: Update on definitions, diagnostic criteria, estimation of severity, and treatments. Diabetes Care 2010, 33, 2285–2293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chawla, R.; Chawla, A.; Jaggi, S. Microvasular and macrovascular complications in diabetes mellitus: Distinct or continuum? Indian J. Endocrinol. Metab. 2016, 20, 546–551. [Google Scholar] [CrossRef]
- Bakker, K.; Apelqvist, J.; Lipsky, B.A.; Van Netten, J.J.; Schaper, N.C.; International Working Group on the Diabetic Foot (IWGDF). The 2015 IWGDF guidance documents on prevention and management of foot problems in diabetes: Development of an evidence-based global consensus. Diabetes Metab. Res. Rev. 2016, 32, 2–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunyach-Remy, C.; Essebe, C.N.; Sotto, A.; Lavigne, J.-P. Staphylococcus aureus toxins and diabetic foot ulcers: Role in pathogenesis and interest in diagnosis. Toxins 2016, 8, 209. [Google Scholar] [CrossRef] [Green Version]
- Pouget, C.; Dunyach-Remy, C.; Pantel, A.; Schuldiner, S.; Sotto, A.; Lavigne, J.-P. Biofilms in diabetic foot ulcers: Significance and clinical relevance. Microorganisms 2020, 8, 1580. [Google Scholar] [CrossRef]
- Chakraborty, S.P.; Mahapatra, S.K.; Sahu, S.K.; Chattopadhyay, S.; Pramanik, P.; Roy, S. Nitric oxide mediated Staphylococcus aureus pathogenesis and protective role of nanoconjugated vancomycin. Asian Pac. J. Trop. Biomed. 2011, 1, 102–109. [Google Scholar] [CrossRef] [Green Version]
- Josse, J.; Laurent, F.; Diot, A. Staphylococcal adhesion and host cell invasion: Fibronectin-binding and other mechanisms. Front. Microbiol. 2017, 8, 2433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falugi, F.; Kim, H.K.; Missiakas, D.M.; Schneewind, O. Role of protein A in the evasion of host adaptive immune responses by Staphylococcus aureus. mBio 2013, 4, e00575-13. [Google Scholar] [CrossRef] [Green Version]
- Bisognano, C.; Vaudaux, P.; Rohner, P.; Lew, D.P.; Hooper, D.C. Induction of fibronectin-binding proteins and increased adhesion of quinolone-resistant Staphylococcus aureus by subinhibitory levels of ciprofloxacin. Antimicrob. Agents Chemother. 2000, 44, 1428–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harraghy, N.; Kormanec, J.; Wolz, C.; Homerova, D.; Goerke, C.; Ohlsen, K.; Qazi, S.; Hill, P.; Herrmann, M. Sae is essential for expression of the staphylococcal adhesins Eap and Emp. Microbiology 2005, 151, 1789–1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeed, K.; Gould, I.; Esposito, S.; Ahmad-Saeed, N.; Ahmed, S.S.; Alp, E.; Bal, A.M.; Bassetti, M.; Bonnet, E.; Chan, M.; et al. Panton–Valentine leukocidin-positive Staphylococcus aureus: A position statement from the international society of chemotherapy. Int. J. Antimicrob. Agents 2018, 51, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Courjon, J.; Munro, P.; Benito, Y.; Visvikis, O.; Bouchiat, C.; Boyer, L.; Doye, A.; Lepidi, H.; Ghigo, E.; Lavigne, J.-P.; et al. EDIN-B promotes the translocation of Staphylococcus aureus to the bloodstream in the course of pneumonia. Toxins 2015, 7, 4131–4142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franke, G.C.; Böckenholt, A.; Sugai, M.; Rohde, H.; Aepfelbacher, M. Epidemiology, variable genetic organization and regu-lation of the EDIN-B toxin in Staphylococcus aureus from bacteraemic patients. Microbiology 2010, 156, 860–872. [Google Scholar] [CrossRef] [Green Version]
- Grumann, D.; Nübel, U.; Bröker, B.M. Staphylococcus aureus toxins—Their functions and genetics. Infect. Genet. Evol. 2014, 21, 583–592. [Google Scholar] [CrossRef] [Green Version]
- Tuchscherr, L.; Löffler, B.; Proctor, R.A. Persistence of Staphylococcus aureus: Multiple metabolic pathways impact the expression of virulence factors in small-colony variants (SCVs). Front. Microbiol. 2020, 11, 1028. [Google Scholar] [CrossRef]
- Heravi, F.S.; Zakrzewski, M.; Vickery, K.; Malone, M.; Hu, H. Metatranscriptomic analysis reveals active bacterial communities in diabetic foot infections. Front. Microbiol. 2020, 11, 1688. [Google Scholar] [CrossRef]
- Richard, J.-L.; Lavigne, J.-P.; Sotto, A. Diabetes and foot infection: More than double trouble. Diabetes Metab. Res. Rev. 2012, 28, 46–53. [Google Scholar] [CrossRef]
- Dumitrescu, O.; Boisset, S.; Badiou, C.; Bes, M.; Benito, Y.; Reverdy, M.E.; Vandenesch, F.; Etienne, J.; Lina, G. Effect of anti-biotics on Staphylococcus aureus producing panton-valentine leukocidin. Antimicrob. Agents Chemother. 2007, 51, 1515–1519. [Google Scholar] [CrossRef] [Green Version]
- Otto, M.P.; Martin, E.; Badiou, C.; Lebrun, S.; Bes, M.; Vandenesch, F.; Etienne, J.; Lina, G.; Dumitrescu, O. Effects of subin-hibitory concentrations of antibiotics on virulence factor expression by community-acquired methicillin-resistant Staphylococcus aureus. J. Antimicrob. Chemother. 2013, 68, 1524–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, D.J.; Gould, I.M. Does vancomycin have a future in the treatment of skin infections? Curr. Opin. Infect. Dis. 2014, 27, 146–154. [Google Scholar] [CrossRef]
- Mauriello, C.T.; Hair, P.S.; Rohn, R.D.; Rister, N.S.; Krishna, N.K.; Cunnion, K.M. Hyperglycemia inhibits complement-mediated immunological control of S. aureus in a rat model of peritonitis. J. Diabetes Res. 2014, 2014, 762051. [Google Scholar] [CrossRef] [Green Version]
- Hodille, E.; Rose, W.; Diep, B.A.; Goutelle, S.; Lina, G.; Dumitrescu, O. The role of antibiotics in modulating virulence in Straphylococcus aureus. Clin. Microbiol. Rev. 2017, 30, 887–917. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Wijenayaka, A.R.; Solomon, L.B.; Pederson, S.M.; Findlay, D.M.; Kidd, S.P.; Atkins, G.J. Novel insights into Staphy-lococcus aureus deep bone infections: The involvement of osteocytes. mBio 2018, 8, e00415-18. [Google Scholar]
- Bui, L.M.; Turnidge, J.D.; Kidd, S.P. The induction of Staphylococcus aureus biofilm formation or small colony variants is a strain-specific response to host-generated chemical stresses. Microbes Infect. 2015, 17, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Bui, L.M.G.; Hoffmann, P.; Turnidge, J.D.; Zilm, P.S.; Kidd, S.P. Prolonged growth of a clinical Staphylococcus aureus strain selects for a stable small-colony-variant cell type. Infect. Immun. 2014, 83, 470–481. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Zilm, P.S.; Kidd, S.P. Novel Research Models for Staphylococcus aureus small colony variants (SCV) development: Co-pathogenesis and Growth Rate. Front. Microbiol. 2020, 11, 321. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Dowd, S.E.; Smith, E.; Rhoads, D.D.; Wolcott, R.D. In vitro multispecies Lubbock chronic wound biofilm model. Wound Repair Regen. 2008, 16, 805–813. [Google Scholar] [CrossRef]
- DeLeon, S.; Clinton, A.; Fowler, H.; Everett, J.; Horswill, A.R.; Rumbaugh, K.P. Synergistic interactions of Pseudomonas aeruginosa and Staphylococcus aureus in an in vitro wound model. Infect. Immun. 2014, 82, 4718–4728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sotto, A.; Lina, G.; Richard, J.-L.; Combescure, C.; Bourg, G.; Vidal, L.; Jourdan, N.; Etienne, J.; Lavigne, J.-P. Virulence Potential of Staphylococcus aureus strains isolated from diabetic foot ulcers: A new paradigm. Diabetes Care 2008, 31, 2318–2324. [Google Scholar] [CrossRef] [Green Version]
- Sotto, A.; Richard, J.-L.; Messad, N.; Molinari, N.; Jourdan, N.; Schuldiner, S.; Sultan, A.; Carrière, C.; Canivet, B.; Landraud, L.; et al. Distinguishing colonization from infection with Staphylococcus aureus in diabetic foot ulcers with miniaturized oligonucleotide arrays: A French multicenter study. Diabetes Care 2012, 35, 617–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Ponsero, A.J.; Armstrong, D.G.; Lipsky, B.A.; Hurwitz, B.L. The dynamic wound microbiome. BMC Med. 2020, 18, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Spichler, A.; Hurwitz, B.L.; Armstrong, D.G.; Lipsky, B.A. Microbiology of diabetic foot infections: From Louis Pasteur to ‘crime scene investigation’. BMC Med. 2015, 13, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowd, S.E.; Wolcott, R.D.; Sun, Y.; McKeehan, T.; Smith, E.; Rhoads, D. Polymicrobial nature of chronic diabetic foot ulcer biofilm infections determined using bacterial tag encoded flx amplicon pyrosequencing (bTEFAP). PLoS ONE 2008, 3, e3326. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.G.; Moura, J.; Carvalho, E.; Empadinhas, N. Microbiota of chronic diabetic wounds: Ecology, impact, and potential for innovative treatment strategies. Front. Microbiol. 2017, 8, 1791. [Google Scholar] [CrossRef]
- Hibbing, M.E.; Fuqua, C.; Parsek, M.R.; Peterson, S.B. Bacterial competition: Surviving and thriving in the microbial jungle. Nat. Rev. Genet. 2009, 8, 15–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalan, L.R.; Meisel, J.S.; Loesche, M.A.; Horwinski, J.; Soaita, I.; Chen, X.; Uberoi, A.; Gardner, S.E.; Grice, E.A. Strain-and Species-Level variation in the microbiome of diabetic wounds is associated with clinical outcomes and therapeutic efficacy. Cell Host Microbe 2019, 25, 641–655.e5. [Google Scholar] [CrossRef]
- James, G.A.; Swogger, E.; Wolcott, R.; Pulcini, E.D.; Secor, P.; Sestrich, J.; Costerton, J.W.; Stewart, P.S. Biofilms in chronic wounds. Wound Repair Regen. 2008, 16, 37–44. [Google Scholar] [CrossRef]
- Malone, M.; Bjarnsholt, T.; McBain, A.; James, G.; Stoodley, P.; Leaper, D.; Tachi, M.; Schultz, G.; Swanson, T.; Wolcott, R. The prevalence of biofilms in chronic wounds: A systematic review and meta-analysis of published data. J. Wound Care 2017, 26, 20–25. [Google Scholar] [CrossRef] [Green Version]
- Clinton, A.; Carter, T. Chronic wound biofilms: Pathogenesis and potential therapies. Lab. Med. 2015, 46, 277–284. [Google Scholar] [CrossRef] [Green Version]
- Regassa, L.B.; Couch, J.L.; Betley, M.J. Steady-state staphylococcal enterotoxin type C mRNA is affected by a product of the accessory gene regulator (AGR) and by glucose. Infect. Immun. 1991, 59, 955–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotz, F. Staphylococcus and biofilms. Mol. Microbiol. 2002, 43, 1367–1378. [Google Scholar] [CrossRef] [PubMed]
- Waldrop, R.; McLaren, A.; Calara, F.; McLemore, R. Biofilm growth has a threshold response to glucose in vitro. Clin. Orthop. Relat. Res. 2014, 472, 3305–3310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bischoff, M.; Wonnenberg, B.; Nippe, N.; Nyffenegger-Jann, N.J.; Voss, M.; Beisswenger, C.; Sunderkötter, C.; Molle, V.; Dinh, Q.T.; Lammert, F.; et al. CcpA affects infectivity of Staphylococcus aureus in a hyperglycemic environment. Front. Cell. Infect. Microbiol. 2017, 7, 172. [Google Scholar] [CrossRef] [PubMed]
- You, Y.; Xue, T.; Cao, L.; Zhao, L.; Sun, H.; Sun, B. Staphylococcus aureus glucose-induced biofilm accessory proteins, GbaAB, influence biofilm formation in a PIA-dependent manner. Int. J. Med. Microbiol. 2014, 304, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Cohen, T.S.; Takahashi, V.; Bonnell, J.; Tovchigrechko, A.; Chaerkady, R.; Yu, W.; Jones-Nelson, O.; Lee, Y.; Raja, R.; Hess, S.; et al. Staphylococcus aureus drives expansion of low-density neutrophils in diabetic mice. J. Clin. Investig. 2019, 129, 2133–2144. [Google Scholar] [CrossRef] [PubMed]
- Shaji, B.V.; Shaji, S.; Haritha, V.H.; Pramod, S.; Anie, Y. Calcium depletion at high glucose concentration promotes vesicle-mediated NET release in response to Staphylococcus aureus. Mol. Immunol. 2020, 124, 211–217. [Google Scholar] [CrossRef]
- Rasigade, J.-P.; Moulay, A.; Lhoste, Y.; Tristan, A.; Bes, M.; Vandenesch, F.; Etienne, J.; Lina, G.; Laurent, F.; Dumitrescu, O. Impact of sub-inhibitory antibiotics on fibronectin-mediated host cell adhesion and invasion by Staphylococcus aureus. BMC Microbiol. 2011, 11, 263. [Google Scholar] [CrossRef] [Green Version]
- Häffner, N.; Bär, J.; Haunreiter, V.D.; Shambat, S.M.; Seidl, K.; Crosby, H.A.; Horswill, A.R.; Zinkernagel, A.S. Intracellular environment and AGR System affect colony size heterogeneity of Staphylococcus aureus. Front. Microbiol. 2020, 11, 1415. [Google Scholar] [CrossRef]
- Gemmell, C.G.; Ford, C.W. Virulence factor expression by Gram-positive cocci exposed to sub-inhibitory concentrations of linezolid. J. Antimicrob. Chemother. 2002, 50, 665–672. [Google Scholar] [CrossRef] [Green Version]
- Bernardo, K.; Pakulat, N.; Fleer, S.; Schnaith, A.; Utermöhlen, O.; Krut, O.; Müller, S.; Krönke, M. Subinhibitory concentrations of linezolid reduce Staphylococcus aureus virulence factor expression. Antimicrob. Agents Chemother. 2004, 48, 546–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pichereau, S.; Pantrangi, M.; Couet, W.; Badiou, C.; Lina, G.; Shukla, S.K.; Rose, W.E. Simulated antibiotic exposures in an in vitro hollow-fiber infection model influence toxin gene expression and production in community-associated methicillin-resistant Staphylococcus aureus Strain MW2. Antimicrob. Agents Chemother. 2011, 56, 140–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyle-Vavra, S.; Daum, R.S. Community-acquired methicillin-resistant Staphylococcus aureus: The role of Panton–Valentine leukocidin. Lab. Investig. 2006, 87, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Vandenesch, F.; Lina, G.; Henry, T. Staphylococcus aureus hemolysins, bi-component leukocidins, and cytolytic peptides: A redundant arsenal of membrane-damaging virulence factors? Front. Cell. Infect. Microbiol. 2012, 2, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shallcross, L.J.; Fragaszy, E.; Johnson, A.M.; Hayward, A.C. The role of the Panton-Valentine leucocidin toxin in staphylococcal disease: A systematic review and meta-analysis. Lancet Infect. Dis. 2013, 13, 43–54. [Google Scholar] [CrossRef] [Green Version]
- Gillet, Y.; Issartel, B.; Vanhems, P.; Fournet, J.-C.; Lina, G.; Bes, M.; Vandenesch, F.; Piémont, Y.; Brousse, N.; Floret, D.; et al. Association between Staphylococcus aureus strains carrying gene for Panton-Valentine leukocidin and highly lethal necrotising pneumonia in young immunocompetent patients. Lancet 2002, 359, 753–759. [Google Scholar] [CrossRef]
- Senneville, E.; Brière, M.; Neut, C.; Messad, N.; Lina, G.; Richard, J.-L.; Sotto, A.; Lavigne, J.-P. First report of the predominance of clonal complex 398 Staphylococcus aureus strains in osteomyelitis complicating diabetic foot ulcers: A national French study. Clin. Microbiol. Infect. 2014, 20, O274–O277. [Google Scholar] [CrossRef] [Green Version]
- Boquet, P.; Lemichez, E. Bacterial virulence factors targeting Rho GTPases: Parasitism or symbiosis? Trends Cell Biol. 2003, 13, 238–246. [Google Scholar] [CrossRef]
- Boyer, L.; Doye, A.; Rolando, M.; Flatau, G.; Munro, P.; Gounon, P.; Clément, R.; Pulcini, C.; Popoff, M.R.; Mettouchi, A.; et al. Induction of transient macroapertures in endothelial cells through RhoA inhibition by Staphylococcus aureus factors. J. Cell Biol. 2006, 173, 809–819. [Google Scholar] [CrossRef]
- Lemichez, E.; Lecuit, M.; Nassif, X.; Bourdoulous, S. Breaking the wall: Targeting of the endothelium by pathogenic bacteria. Nat. Rev. Genet. 2009, 8, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Rolando, M.; Munro, P.; Stefani, C.; Auberger, P.; Flatau, G.; Lemichez, E. Injection of Staphylococcus aureus EDIN by the Bacillus anthracis protective antigen machinery induces vascular permeability. Infect. Immun. 2009, 77, 3596–3601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munro, P.; Benchetrit, M.; Nahori, M.-A.; Stefani, C.; Clément, R.; Michiels, J.-F.; Landraud, L.; Dussurget, O.; Lemichez, E. The Staphylococcus aureus epidermal cell differentiation inhibitor toxin promotes formation of infection foci in a mouse model of bacteremia. Infect. Immun. 2010, 78, 3404–3411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messad, N.; Landraud, L.; Canivet, B.; Lina, G.; Richard, J.-L.; Sotto, A.; Lavigne, J.-P.; Lemichez, E. Distribution of edin in Staphylococcus aureus isolated from diabetic foot ulcers. Clin. Microbiol. Infect. 2013, 19, 875–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 10.0. 2020. Available online: http://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_10.0_Breakpoint_Tables.pdf (accessed on 8 February 2021).
- Essebe, C.N.; Visvikis, O.; Fines-Guyon, M.; Vergne, A.; Cattoir, V.; Lecoustumier, A.; Lemichez, E.; Sotto, A.; Lavigne, J.-P.; Dunyach-Remy, C. Decrease of Staphylococcus aureus virulence by Helcococcus kunzii in a Caenorhabditis elegans model. Front. Cell. Infect. Microbiol. 2017, 7, 77. [Google Scholar] [CrossRef] [Green Version]
- Garsin, D.A.; Sifri, C.D.; Mylonakis, E.; Qin, X.; Singh, K.V.; Murray, B.E.; Calderwood, S.B.; Ausubel, F.M. A simple model host for identifying Gram-positive virulence factors. Proc. Natl. Acad. Sci. USA 2001, 98, 10892–10897. [Google Scholar] [CrossRef] [Green Version]
- Chavant, P.; Gaillard-Martinie, B.; Talon, R.; Hebraud, M.; Bernardi, T. A new device for rapid evaluation of biofilm formation potential by bacteria. J. Microbiol. Methods 2007, 68, 605–612. [Google Scholar] [CrossRef]
- Qiu, J.; Feng, H.; Xiang, H.; Wang, D.; Xia, L.; Jiang, Y.; Song, K.; Lu, J.; Yu, L.; Deng, X. Influence of subinhibitory concen-trations of licochalcone A on the secretion of enterotoxins A and B by Staphylococcus aureus. FEMS Microbiol. Lett. 2010, 307, 135–141. [Google Scholar] [CrossRef] [Green Version]
- Dumitrescu, O.; Choudhury, P.; Boisset, S.; Badiou, C.; Bes, M.; Benito, Y.; Wolz, C.; Vandenesch, F.; Etienne, J.; Cheung, A.L.; et al. Beta-lactams interfering with PBP1 induce Panton-Valentine leukocidin expression by triggering sarA and rot global regulators of Staphylococcus aureus. Antimicrob. Agents Chemother. 2011, 55, 3261–3271. [Google Scholar] [CrossRef] [Green Version]
- Munro, P.; Clément, R.; Lavigne, J.P.; Pulcini, C.; Lemichez, E.; Landraud, L. High prevalence of edin-C encodign RhoA-targeting toxin in clinical isolates of Staphylococcus aureus. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 30, 965–972. [Google Scholar] [CrossRef] [Green Version]
- Atshan, S.S.; Shamsudin, M.N.; Karunanidhi, A.; Van Belkum, A.; Lung, L.T.T.; Sekawi, Z.; Nathan, J.J.; Ling, K.H.; Seng, J.S.C.; Ali, A.M.; et al. Quantitative PCR analysis of genes expressed during biofilm development of methicillin resistant Staphylococcus aureus (MRSA). Infect. Genet. Evol. 2013, 18, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Labandeira-Rey, M.; Couzon, F.; Boisset, S.; Brown, E.L.; Bes, M.; Benito, Y.; Barbu, E.M.; Vazquez, V.; Höök, M.; Etienne, J.; et al. Staphylococcus aureus Panton-valentine leukocidin causes necrotizing pneumonia. Science 2007, 315, 1130–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garzoni, C.; Francois, P.; Huyghe, A.; Couzinet, S.; Tapparel, C.; Charbonnier, Y.; Renzoni, A.; Lucchini, S.; Lew, D.P.; Vaudaux, P.; et al. A global view of Staphylococcus aureus whole genome expression upon internalization in human epithelial cells. BMC Genom. 2007, 8, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Characteristics | References |
|---|---|---|
| NSA739 | S. aureus strain isolated from DFI (Grade 3), PVL-, edin-, agr2, ST8 | [31] |
| NSA1385 | S. aureus strain isolated from colonized DFU (Grade 1), PVL-, edin-, agr1, ST8 | [31] |
| NSA1077 | S. aureus strain isolated from DFI (Grade 3), PVL+, edinB+, agr1, ST152 | [32] |
| NSA7475 | S. aureus strain isolated from DFI (Grade 3), PVL-, edinB+, agr1, ST25 | [32] |
| Strains | In Vitro WLM Alone | WLM + 10% Glucose | WLM + Vancomycin | WLM + Linezolid | WLM + 10% Glucose + Vancomycin | WLM + 10% Glucose + Linezolid | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| β-hemol 1 | SCV 2 | β-hemol | SCV | β-hemol | SCV | β-hemol | SCV | β-hemol | SCV | β-hemol | SCV | |
| NSA739 | 99% ± 2 | 2% ± 2 | 12% ± 5 | 22% ± 2 | 93% ± 5 | 2% ± 2 | 28% ± 4 | 5% ± 2 | 5% ± 2 | 30% ± 7 | 13% ± 6 | 18%± 7 |
| NSA1077 | 98% ± 3 | 2% ± 2 | 5% ± 5 | 11% ± 2 | 91% ± 7 | 2% ± 3 | 55% ± 5 | 20% ± 5 | 8% ± 4 | 18% ± 5 | 10% ± 10 | 10%± 5 |
| NSA7475 | 98% ± 2 | 1% ± 2 | 65% ± 3 | 24% ± 2 | 88% ± 7 | 4% ± 2 | 48% ± 2 | 15% ± 4 | 50% ± 5 | 15% ± 3 | 37% ± 8 | 17%± 8 |
| NSA1385 | 99% ± 2 | 0% ± 2 | 93%± 7 | 4% ± 2 | 100% ± 2 | 0% ± 2 | 75% ± 5 | 7% ± 2 | 94% ± 2 | 2% ± 2 | 91% ± 5 | 3% ± 3 |
| Strains | Length of Preculture | Without Pre-Culture in WLM | WLM Alone | WLM + 10% Glucose | WLM + Vancomycin | WLM + Linezolid | WLM + 10% Glucose + Vancomycin | WLM + 10% Glucose + Linezolid |
|---|---|---|---|---|---|---|---|---|
| NSA739 24 h | 24 h | 1.7 ± 0.3 | 3.5 ± 0.2 | 3.5 ± 0.3 | 3.4 ± 0.2 | 3.8 ± 0.3 | 3.9 ± 0.2 | 5.00 ± 0.4 |
| NSA739 16-week | 16 weeks | NA | 3.9 ± 0.2 | 4.9 ± 0.2 | 3.2 ± 0.3 | 4.9 ± 0.2 | 5.0 ± 0.2 | 4.9 ± 0.2 |
| NSA1077 24 h | 24 h | 2.2 ± 0.2 | 3.9 ± 0.2 | 4.2 ± 0.3 | 4.1 ± 0.1 | 4.3 ± 0.2 | 4.4 ± 0.1 | 4.9 ± 0.2 |
| NSA1077 16-week | 16 weeks | NA | 4.4 ± 0.1 | 5.4 ± 0.4 | 3.7 ± 0.2 | 5.4 ± 0.4 | 5.5 ± 0.3 | 5.5 ± 0.4 |
| NSA7475 24 h | 24 h | 2.3 ± 0.3 | 3.8 ± 0.2 | 3.9 ± 0.3 | 3.7 ± 0.2 | 3.5 ± 0.2 | 3.8 ± 0.2 | 5.2 ± 0.4 |
| NSA7475 16-week | 16 weeks | NA | 4.3 ± 0.3 | 5.3 ± 0.4 | 3.5 ± 0.2 | 4.7 ± 0.3 | 5.4 ± 0.4 | 5.0 ± 0.2 |
| 1385 24 h | 24 h | 4.3 ± 0.3 | 4.8 ± 0.3 | 4.6 ± 0.3 | 4.3 ± 0.4 | 5.1 ± 0.2 | 4.8 ± 0.3 | 4.4 ± 0.2 |
| 1385 16-week | 16 weeks | NA | 5.0 ± 0.4 | 5.2 ± 0.2 | 5.1 ± 0.2 | 5.1 ± 0.3 | 4.9 ± 0.3 | 4.9 ± 0.2 |
| OP50 (Control strain) | - | 7.7 ± 0.2 | NA | NA | NA | NA | NA | NA |
| Effect on Expression of Virulence Factor 1 | ||||
|---|---|---|---|---|
| PVL | EDIN | Alpha-Hemolysin | Protein A | |
| Short exposure in WLM added to | ||||
| Glucose | ↑↑ | ↑↑ | - | ↓ |
| Vancomycin | - | - | - | - |
| Linezolid | ↑↑ | ↓ | ↓ | ↓ |
| Long exposure in WLM added to | ||||
| Glucose | ↓ | ↓ | ↓ | ↑ |
| Vancomycin | - | ↓↓ | ↑ | ↓ |
| Linezolid | ↓↓ | ↓↓ | ↓ | ↑ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pouget, C.; Gustave, C.-A.; Ngba-Essebe, C.; Laurent, F.; Lemichez, E.; Tristan, A.; Sotto, A.; Dunyach-Rémy, C.; Lavigne, J.-P. Adaptation of Staphylococcus aureus in a Medium Mimicking a Diabetic Foot Environment. Toxins 2021, 13, 230. https://doi.org/10.3390/toxins13030230
Pouget C, Gustave C-A, Ngba-Essebe C, Laurent F, Lemichez E, Tristan A, Sotto A, Dunyach-Rémy C, Lavigne J-P. Adaptation of Staphylococcus aureus in a Medium Mimicking a Diabetic Foot Environment. Toxins. 2021; 13(3):230. https://doi.org/10.3390/toxins13030230
Chicago/Turabian StylePouget, Cassandra, Claude-Alexandre Gustave, Christelle Ngba-Essebe, Frédéric Laurent, Emmanuel Lemichez, Anne Tristan, Albert Sotto, Catherine Dunyach-Rémy, and Jean-Philippe Lavigne. 2021. "Adaptation of Staphylococcus aureus in a Medium Mimicking a Diabetic Foot Environment" Toxins 13, no. 3: 230. https://doi.org/10.3390/toxins13030230
APA StylePouget, C., Gustave, C.-A., Ngba-Essebe, C., Laurent, F., Lemichez, E., Tristan, A., Sotto, A., Dunyach-Rémy, C., & Lavigne, J.-P. (2021). Adaptation of Staphylococcus aureus in a Medium Mimicking a Diabetic Foot Environment. Toxins, 13(3), 230. https://doi.org/10.3390/toxins13030230