Evaluation of a Yeast Hydrolysate from a Novel Strain of Saccharomyces cerevisiae for Mycotoxin Mitigation using In Vitro and In Vivo Models

Abstract
:1. Introduction
2. Results
2.1. In Vitro Adsorption of Mycotoxins by Yeast Products
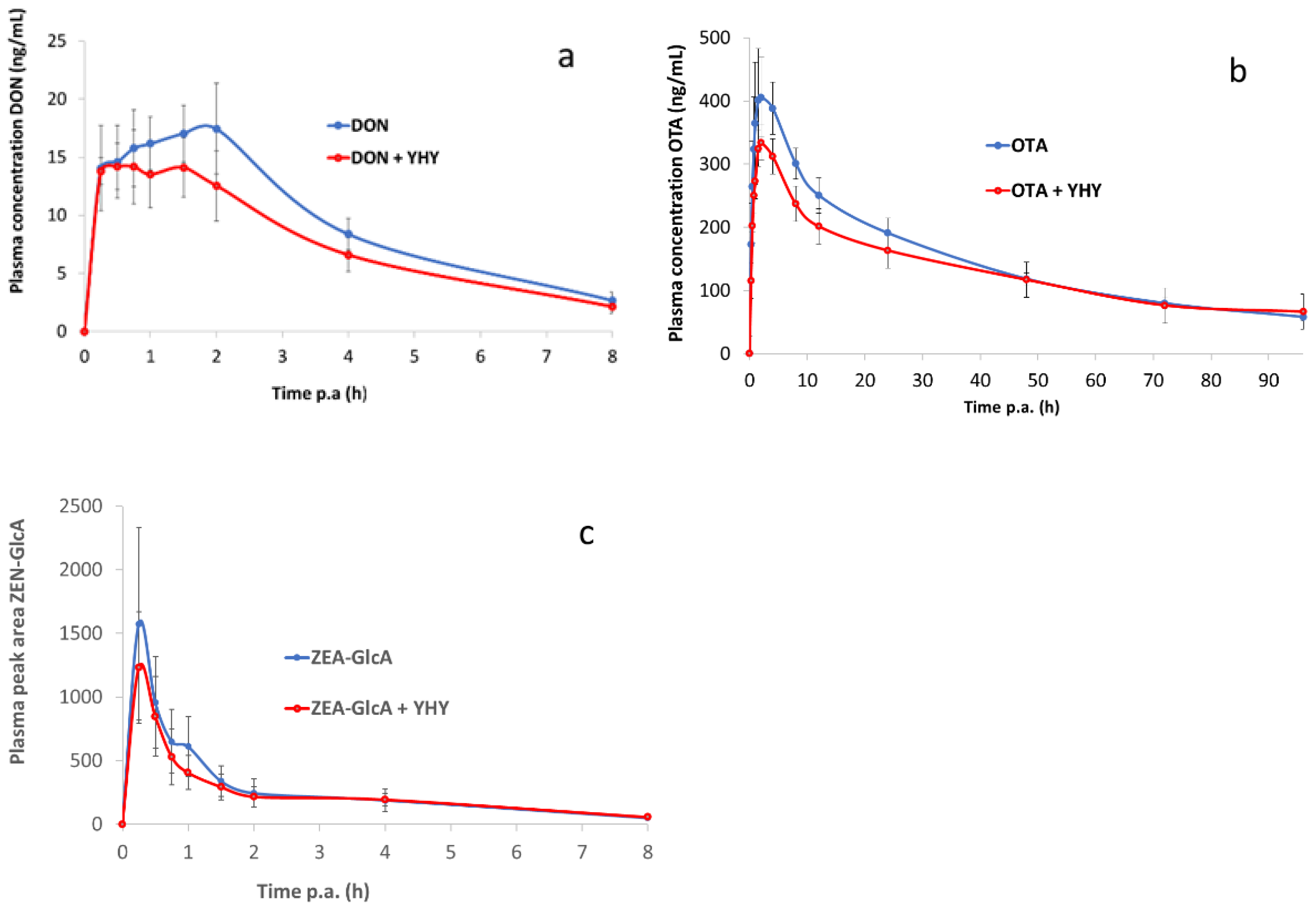
2.2. Toxicokinetic Study
3. Discussion
4. Materials and Methods
4.1. Preparation of the Yeast Products
4.2. In Vitro Assessment of pH-Dependent Adsorption/Desorption
4.3. Quantification of Mycotoxinsin Supernatants of Adsorption Tests
4.4. Animal Study
4.5. Quantification of Mycotoxins in Plasma
4.6. Toxicokinetic Analysis
4.7. Effect of the Mycotoxin Binder on Oral Absorption of the Mycotoxins
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pierron, A.; Alassane-Kpembi, I.; Oswald, I.P. Impact of mycotoxins on immune response and consequences for pig health. Anim. Nutr. 2016, 2, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Vidal, A.; Mengelers, M.; Yang, S.; De Saeger, S.; De Boevre, M. Mycotoxin biomarkers: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1127–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schatzmayer, G.; Streit, E. Global occurrence of mycotoxins in food and feed chain: Facts and figures. World Mycotoxin J. 2013, 6, 213–222. [Google Scholar] [CrossRef]
- Guerre, P. Worldwide mycotoxins exposure in pig and poultry feed formulations. Toxins 2016, 8, 350. [Google Scholar] [CrossRef]
- Alizadeh, A.; Braber, S.; Akbari, P.; Garssen, J.; Fink-Gremmels, J. Deoxynivalenol impairs weight gain and affects markers of gut health after low-dose, short-term exposure of growing pigs. Toxins 2015, 7, 2071–2095. [Google Scholar] [CrossRef]
- Fink-Gremmels, J.; Malekinejad, H. Clinical effects and biochemical mechanisms associated with exposure to the mycoestrogen zearalenone. Anim. Feed Technol. 2007, 137, 326–341. [Google Scholar] [CrossRef]
- Voss, K.A.; Smith, G.W.; Haschek, W.M. Fumonisins: Toxicokinetics, mechanism of action and toxicity. Anim. Feed Sci. Technol. 2007, 137, 299–325. [Google Scholar] [CrossRef]
- Piotrowska, M.; Masek, A. Saccharomyces cerevisiae cell wall components as tools for ochratoxin A decontamination. Toxins 2015, 7, 1151–1162. [Google Scholar] [CrossRef] [Green Version]
- Jouany, J.-P.; Yiannikouris, A.; Bertin, G. The chemical bonds between mycotoxins and cell wall components of Saccharomyces cerevisiae have been identified. Arch. Zootech. 2005, 8, 26–50. [Google Scholar]
- Ringot, D.; Lerzy, B.; Bonhoure, J.P.; Auclair, E.; Oriol, E.; Larondelle, Y. Effect of temperature on in vitro ochratoxin bisorption onto yeast cell wall derivatives. Process Biochem. 2005, 40, 3008–3016. [Google Scholar] [CrossRef]
- Yiannikouris, A.; François, J.; Poughon, L.; Dussap, C.-G.; Bertin, G.; Jeminet, G.; Jouany, J.-P. Alkali extraction of beta-D-glucans from Saccharomyces cerevisiae cell wall and study of their adsorptive properties toward zearalenone. J. Agric. Food Chem. 2004, 52, 3666–3673. [Google Scholar] [CrossRef]
- Yiannikouris, A.; André, G.; Poughon, L.; François, J.; Dussap, C.-G.; Jeminet, G.; Bertin, G.; Jouany, J.-P. Chemical and conformational study of the interactions involved in mycotoxin complexation with β-D-glucans. Biomacromolecules 2006, 7, 1147–1155. [Google Scholar] [CrossRef]
- Faucet-Marquis, V.; Joannis-Cassan, C.; Hadjeba-Medjdoub, K.; Ballet, N.; Pfohl-Leszkowicz, A. Development of an in vitro method for the prediction of mycotoxin binding on yeast-based products: Case of aflatoxin B1, zearalenone and ochratoxin A. Appl. Microbiol. Biotechnol. 2014, 98, 7583–7596. [Google Scholar] [CrossRef] [Green Version]
- Shetty, P.H.; Jespersen, L. Saccharomyces cerevisiae and lactic acid bacteria as potential mycotoxin decontaminating agents. Trends Food Sci. Technol. 2006, 17, 48–55. [Google Scholar] [CrossRef]
- Gambacorta, L.; Pinton, P.; Avantaggiato, G.; Oswald, I.P.; Solfrizzo, M. Grape pomace, an agricultural byproduct reducing mycotoxin absorption: In vivo assessment in pig using urinary biomarkers. J. Food Chem. 2016, 64, 6762–6771. [Google Scholar] [CrossRef]
- EFSA. Review of Mycotoxin-Detoxifying agents Used as Feed Additives: Mode of Action, Efficacy and Feed/Food Safety. 8 December 2009. Available online: http://www.efsa.europa.eu/en/scdocs/scdoc/22e.htm (accessed on 21 October 2021).
- EFSA. Statement on the establishment of guidelines for the assessment of additives from the functional group ‘substances for reduction of the contamination of feed by mycotoxins’. EFSA J. 2010, 8, 1693. [Google Scholar] [CrossRef]
- European Commission. COMMISSION RECOMMENDATION of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding. Off. J. Eur. Union 2006, L229, 7–9. [Google Scholar]
- De Mil, T.; Devreese, M.; De Saeger, S.; Eeckhout, M.; De Backer, P.; Croubels, S. Influence of mycotoxin binders on the oral bioavailability of doxycycline in pigs. J. Agric. Food Chem. 2016, 64, 2120–2126. [Google Scholar] [CrossRef] [Green Version]
- De Mil, T.; Devreese, M.; Maes, A.; De Saeger, S.; De Backer, P.; Croubels, S. Influence of mycotoxin binders on the oral bioavailability of tylosin, doxycycline, diclazuril, and salinomycin in fed broiler chickens. Poult. Sci. 2017, 96, 2137–2144. [Google Scholar] [CrossRef]
- Sabater-Vilarand, M.; Malekinejad, H.; Selman, M.H.J.; van der Doelen, M.A.M.; Fink-Gremmels, J. In vitro assessment of adsorbents aiming to prevent deoxynivalenol and zearalenone mycotoxicoses. Mycopathologia 2007, 163, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Cavret, S.; Laurent, N.; Videmann, B.; Mazallou, M.; Lecoeur, S. Assessment of deoxynivalenol (DON) adsorbents and characterization of their efficacy using complementary in vitro tests. Food Addit. Contam. 2010, 27, 43–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devreese, M.; Antonissen, G.; De Backer, P.; Croubles, S. Efficacy of active carbon towards the absorption of deoxynivalenol in pigs. Toxins 2014, 5, 2998–3004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwerdt, G.; Freudinger, R.; Silbernagagl, S.; Gekle, M. Ochratoxin A-binding proteins in rat organs and plasma and in different cell lines of the kidney. Toxicology 1999, 135, 1–10. [Google Scholar] [CrossRef]
- Kollar, R.; Reinhold, B.B.; Petrakova, E.; Yeh, H.J.C.; Ashwell, G.; Drgonova, J.; Kapteyn, J.C.; Klis, F.M.; Cabib, E. Architecture of the yeast cell wall: β(1,6)-D-glucan interconnects mannoprotein, β(1,3)-D-glucan, and chitin. J. Biol. Chem. 1997, 272, 17762–17775. [Google Scholar] [CrossRef] [Green Version]
- Manners, D.J.; Masson, A.J.; Patterson, J.C. The structure of a β(1,3)-D-glucan from yeast cell walls. Biochem. J. 1973, 135, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Yiannikouris, A.; François, J.; Poughon, L.; Dussap, C.-G.; Bertin, G.; Jeminet, G.; Jouany, J.-P. Adsorption of zearalenone by β-D-glucans in the Saccharomyces cerevisiae cell wall. J. Food Prot. 2004, 67, 1195–1200. [Google Scholar] [CrossRef]
- Freimund, S.; Sauter, M.; Rys, P. Efficient adsorption of the mycotoxins zearalenone and T-2 toxin on a modified yeast glucan. J. Environ. Sci. Health B Pestic. Food Contam. Agric. Wastes 2003, 38, 243–255. [Google Scholar] [CrossRef]
- Kolawole, O.; Meneely, J.; Greer, B.; Chevallier, O.; Jones, D.S.; Connoly, L.; Elliot, C. Comparative in vitro assessment of a range of commercial feed additives with multiple mycotoxin binding claims. Toxins 2019, 11, 659. [Google Scholar] [CrossRef] [Green Version]
- Nathanail, A.V.; Gibson, B.; Han, L.; Peltonen, K.; Ollilainen, V.; Jestoi, M.; Laitila, A. The lager yeast Saccharomyces pastorianus removes and transforms trichothecene mycotoxins during fermentation of brewer’s wort. Food Chem. 2016, 203, 448–455. [Google Scholar] [CrossRef]
- Grenier, B.; Applegate, T.J. Modulation of intestinal functions following mycotoxin ingestion: Meta-analysis of published experiments in animals. Toxins 2013, 5, 396–430. [Google Scholar] [CrossRef] [Green Version]
- Catteuw, A.; Devreese, M.; De Baere, S.; Antonissen, G.; Ivanova, L.; Uhlig, S.; Martens, A.; De Saeger, S.; De Boevre, M.; Croubels, S. Investigation of age-related differences in toxicokinetic processes of deoxynivalenol and dexoxynivalenol-3-glucoside in weaned piglets. Arch. Toxicol. 2020, 94, 417–425. [Google Scholar] [CrossRef]
- Broekaert, N.; Devreese, M.; De Mil, T.; Fraeyman, S.; Antonissen, G.; De Baere, S.; De Backer, P.; Vermeulen, A.; Croubels, S. Oral bioavailability, hydrolysis, and comparative toxicokinetics of 3-acetyldeoxynivalenol and 15-acetyldeoxynivalenol in broiler chickens and pigs. J. Agric. Food Chem. 2015, 63, 8734–8742. [Google Scholar] [CrossRef]
- Broekaert, N.; Devreese, M.; van Bergen, T.; Schauvliege, S.; De Boevre, M.; De Saeger, S.; Vanhaecke, L.; Berthiller, F.; Michlmayr, H.; Malachová, A.; et al. In vivo contribution of deoxynivalenol-3-β-D-glucoside to deoxynivalenol exposure in broiler chickens and pigs: Oral bioavailability, hydrolysis and toxicokinetics. Arch. Toxicol. 2017, 91, 699–712. [Google Scholar] [CrossRef] [Green Version]
- Serviento, A.M.; Brossard, L.; Renaudeau, D. An acute challenge with a deoxynivalenol-contaminated diet has short- and long-term effects on performance and feeding behaviour in finishing pigs. J. Anim. Sci. 2018, 96, 5209–5221. [Google Scholar] [CrossRef]
- Friend, D.W.; Trenholm, H.L.; Elliot, J.I.; Thompson, B.K.; Hartin, K.E. Effect of feeding vomitoxin-contaminated wheat to pigs. Can. J. Anim. Sci. 1982, 62, 1211–1222. [Google Scholar] [CrossRef]
- Devreese, M.; Osselaere, A.; Goossens, J.; Vandenbroucke, V.; De Baere, S.; Eeckhout, M.; De Baeker, P.; Croubels, S. New bolus models for in vivo efficacy testing of mycotoxin-detoxifying agents in relation to EFSA guidelines, assessed using deoxynivalenol in broiler chickens. Food Addit. Contam. Part A 2012, 29, 1101–1107. [Google Scholar] [CrossRef] [Green Version]
- Goossens, J.; Vandenbroucke, V.; Pasmans, F.; De Baere, S.; Devreese, M.; Osselaere, A.; Verbrugghe, E.; Haesebrouck, F.; De Saeger, S.; Eeckhout, M.; et al. Influence of mycotoxin detoxifying agents on oral bioavailability of commonly used antibiotics in pigs. Toxins 2012, 4, 281–295. [Google Scholar] [CrossRef] [Green Version]
- Osselaere, A.; Devreese, M.; Watteyn, A.; Vandenbroucke, V.; Goossens, J.; Haetekiet, V.; Eeckhout, M.; De Saeger, S.; De Baere, S.; De Backer, P.; et al. Efficacy and safety testing of mycotoxin-detoxifying agents in broilers following EFSA guidelines. Poult. Sci. 2012, 91, 2046–2054. [Google Scholar] [CrossRef]
- Alassane-Kpembi, M.; Canlet, C.; Tremblay-Franco, M.; Jourdan, F.; Chalzaviel, M.; Pinton, P.; Cossalter, A.M.; Achard, C.; Castex, M.; Combes, S.; et al. 1H-NMR metabolomics response to a realistic diet contamination with the mycotoxin deoxynivalenol: Effect of probiotics supplementation. Food Chem. Toxicol. 2020, 138, 111222. [Google Scholar] [CrossRef]
- Kovalsky, P.; Kos, G.; Nährer, K.; Schwab, C.; Jenkins, T.; Schatzmayer, G.; Sulyok, M.; Krska, R. Co-occurrence of regulated, masked and emerging mycotoxins in finished feed and maize-An extensive survey. Toxins 2016, 8, 363. [Google Scholar] [CrossRef] [Green Version]
- Lauwers, M.; De Baere, S.; Letor, B.; Rychlik, M.; Croubels, S.; Devreese, M. Multi LC-MS/MS and LC-HRMS methods for determination of 24 mycotoxins including major phase I and phase II biomarker metabolites in biological matrices from pigs and broiler chickens. Toxins 2019, 11, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| DON | |||||||||
| pH 3.0 | pH 5.0 | pH 8.5 | |||||||
| Treatment | ng/mL | % of NC | p Value | (ng/mL) | % of NC | p Value | ng/mL | % of NC | p Value |
| NC | 990 ± 26 | --- | --- | 992 ± 30 | --- | --- | 1012 ± 70 | --- | --- |
| YHY | 540 ± 5 | 45.5 ± 1.0 | <0.0001 | 538 ± 7 | 45.8 ± 0.8 | <0.0001 | 543 ± 20 | 46.7 ± 1.8 | <0.0001 |
| YA1 | 887 ± 36 | 10.3 ± 1.7 | 0.3133 | 890 ± 5 | 10.0 ± 2.1 | 0.3345 | 903 ± 92 | 11.4 ± 2.1 | 0.2551 |
| YA2 | 876 ± 45 | 11.4 ± 2.4 | 0.2049 | 878 ± 26 | 11.3 ± 1.9 | 0.2049 | 963 ± 90 | 4.8 ± 3.6 | 0.9802 |
| OTA | |||||||||
| NC | 32 ± 6 | --- | --- | 57 ± 8 | --- | --- | 68 ± 6 | ---- | --- |
| YHY | 31 ± 3 | 9.3 ± 8.1 | 1.0000 | 52 ± 3 | 8.3 ± 7.6 | 0.9943 | 60 ± 5 | 12.0 ± 3.6 | 0.8283 |
| YA1 | 25 ± 5 | 21.0 ± 7.6 | 0.9525 | 50 ± 9 | 14.0 ± 13.5 | 0.9525 | 66 ± 5 | 8.3 ± 7.6 | 0.9998 |
| YA2 | 27 ± 8 | 16.3 ± 14.8 | 0.9943 | 52 ± 6 | 8.3 ± 7.6 | 0.9943 | 65 ± 5 | 5.0 ± 4.4 | 0.9998 |
| ZEA | |||||||||
| NC | 275 ± 6 | --- | --- | 280 ± 5 | --- | --- | 376 ± 38 | --- | --- |
| YHY | 264 ± 13 | 4.0 ± 3.5 | 0.9998 | 247 ± 11 | 11.7 ± 2.0 | 0.6119 | 328 ± 45 | 13.0 ± 2.7 | 0.1427 |
| YA1 | 125 ± 9 | 54.3 ± 4.0 | <0.0001 | 120 ± 10 | 57.0 ± 3.1 | <0.0001 | 188 ± 8 | 49.7 ± 5.5 | <0.0001 |
| YA2 | 109 ± 5 | 60.3 ± 0.6 | <0.0001 | 107 ± 16 | 62.0 ± 6.2 | <0.0001 | 133 ± 8 | 64.7 ± 3.2 | <0.0001 |
| Toxicokinetic Parameters | DON | DON + YHY | p Value |
|---|---|---|---|
| AUC0–8 h (h.ng/mL) | 74.57 ± 8.39 | 60.03 ± 12.58 | 0.462 |
| AUC0–2 h (h.ng/mL) | 30.03 ± 3.87 | 25.76 ± 4.58 | 0.561 |
| Cmax (ng/mL) | 20.17 ± 4.22 | 15.47 ± 1.83 | 0.306 |
| Tmax (h) | 1.13 ± 0.71 | 0.88 ± 0.29 | 0.626 |
| T1/2 el (h) | 2.15 ± 0.10 | 2.39 ± 0.25 | 0.479 |
| ke (1/h) | 0.32 ± 0.01 | 0.30 ± 0.03 | 0.377 |
| Relative F AUC0–8 h (%) | / | 80.50 | |
| Relative F AUC0–2 h (%) | / | 85.79 |
| Toxicokinetic Parameters | OTA | OTA + YHY | p Value |
|---|---|---|---|
| AUC0–96 h (h.µg/mL) | 14.13 ± 1.48 | 12.28 ± 2.06 | 0.135 |
| AUC0–4 h (h.µg/mL) | 1.42 ± 0.24 | 1.14 ± 0.25 | 0.288 |
| Cmax (ng/mL) | 426.23 ± 85.10 | 353.83 ± 64.21 | 0.076 |
| Tmax (h) | 2.42 ± 1.06 | 2.04 ± 0.65 | 0.853 |
| T1/2 el (h) | 45.12 ± 6.61 | 50.69 ± 12.40 | 0.464 |
| ke (1/h) | 0.016 ± 0.002 | 0.015 ± 0.004 | 0.674 |
| Relative F AUC0–96 h (%) | / | 86.86 | |
| Relative F AUC0–4 h (%) | / | 79.86 |
| Toxicokinetic Parameters | ZEA | ZEA + YHY | p Value |
|---|---|---|---|
| AUC0–8 h (h.peak area/mL) | 2168.20 ± 494.87 | 1917.16 ± 444.98 | 0.985 |
| AUC0–0.5 h (h.peak area/mL) | 505.92 ± 227.89 | 407.48 ± 141.70 | 0.765 |
| Cmax (peak area/mL) | 1627.15 ± 737.12 | 1302.53 ± 485.21 | 0.308 |
| Tmax (h) | 0.33 ± 0.14 | 0.29 ± 0.07 | 0.482 |
| T1/2 el (h) | 3.60 ± 2.00 | 2.93 ± 0.85 | 0.523 |
| ke (1/h) | 0.27 ± 0.12 | 0.27 ± 0.05 | 0.533 |
| Relative F AUC0–8 h (%) | / | 88.42 | |
| Relative F AUC0–0.5 h (%) | / | 80.54 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bruinenberg, P.G.; Castex, M. Evaluation of a Yeast Hydrolysate from a Novel Strain of Saccharomyces cerevisiae for Mycotoxin Mitigation using In Vitro and In Vivo Models. Toxins 2022, 14, 7. https://doi.org/10.3390/toxins14010007
Bruinenberg PG, Castex M. Evaluation of a Yeast Hydrolysate from a Novel Strain of Saccharomyces cerevisiae for Mycotoxin Mitigation using In Vitro and In Vivo Models. Toxins. 2022; 14(1):7. https://doi.org/10.3390/toxins14010007
Chicago/Turabian StyleBruinenberg, Paul Gerard, and Mathieu Castex. 2022. "Evaluation of a Yeast Hydrolysate from a Novel Strain of Saccharomyces cerevisiae for Mycotoxin Mitigation using In Vitro and In Vivo Models" Toxins 14, no. 1: 7. https://doi.org/10.3390/toxins14010007
APA StyleBruinenberg, P. G., & Castex, M. (2022). Evaluation of a Yeast Hydrolysate from a Novel Strain of Saccharomyces cerevisiae for Mycotoxin Mitigation using In Vitro and In Vivo Models. Toxins, 14(1), 7. https://doi.org/10.3390/toxins14010007