Genetic Interaction of Global Regulators AflatfA and AflatfB Mediating Development, Stress Response and Aflatoxins B1 Production in Aspergillus flavus


Abstract
:1. Introduction
2. Results
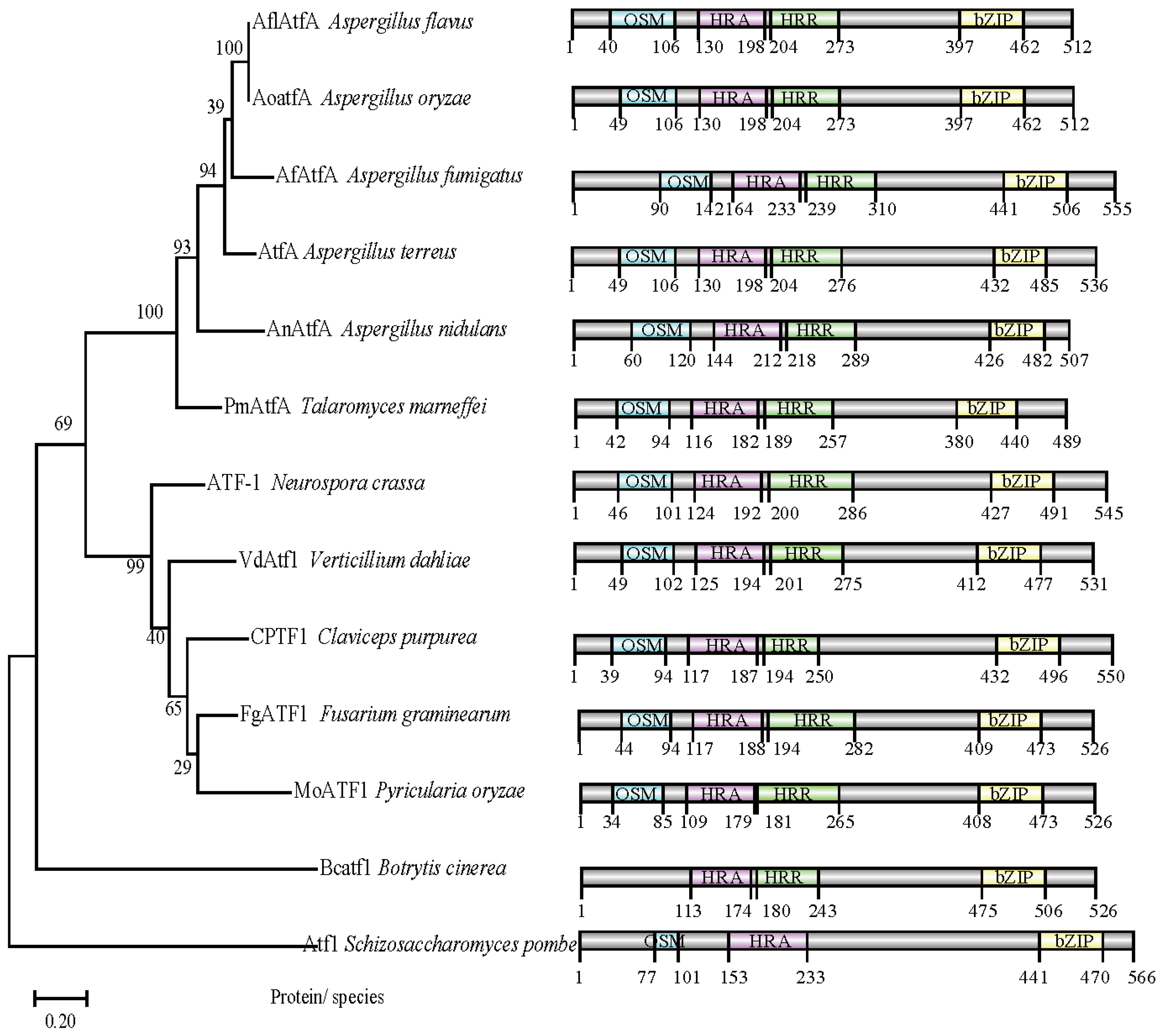
2.1. AflatfA and AflatfB Are ATF/CREB Homologous Genes in A. flavus
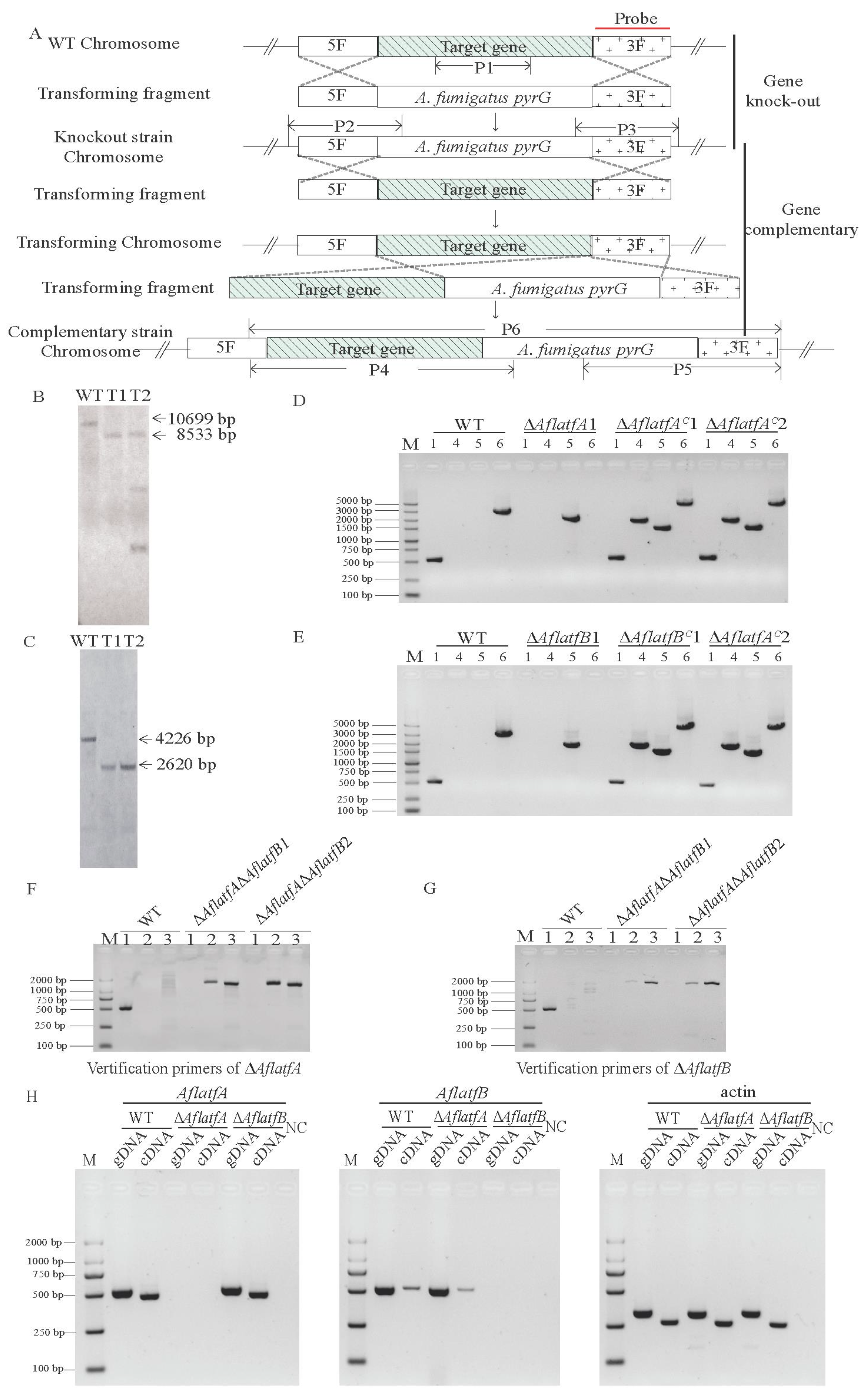
2.2. The Generation of Mutants and Expression of AflatfB Was Impacted by AflatfA
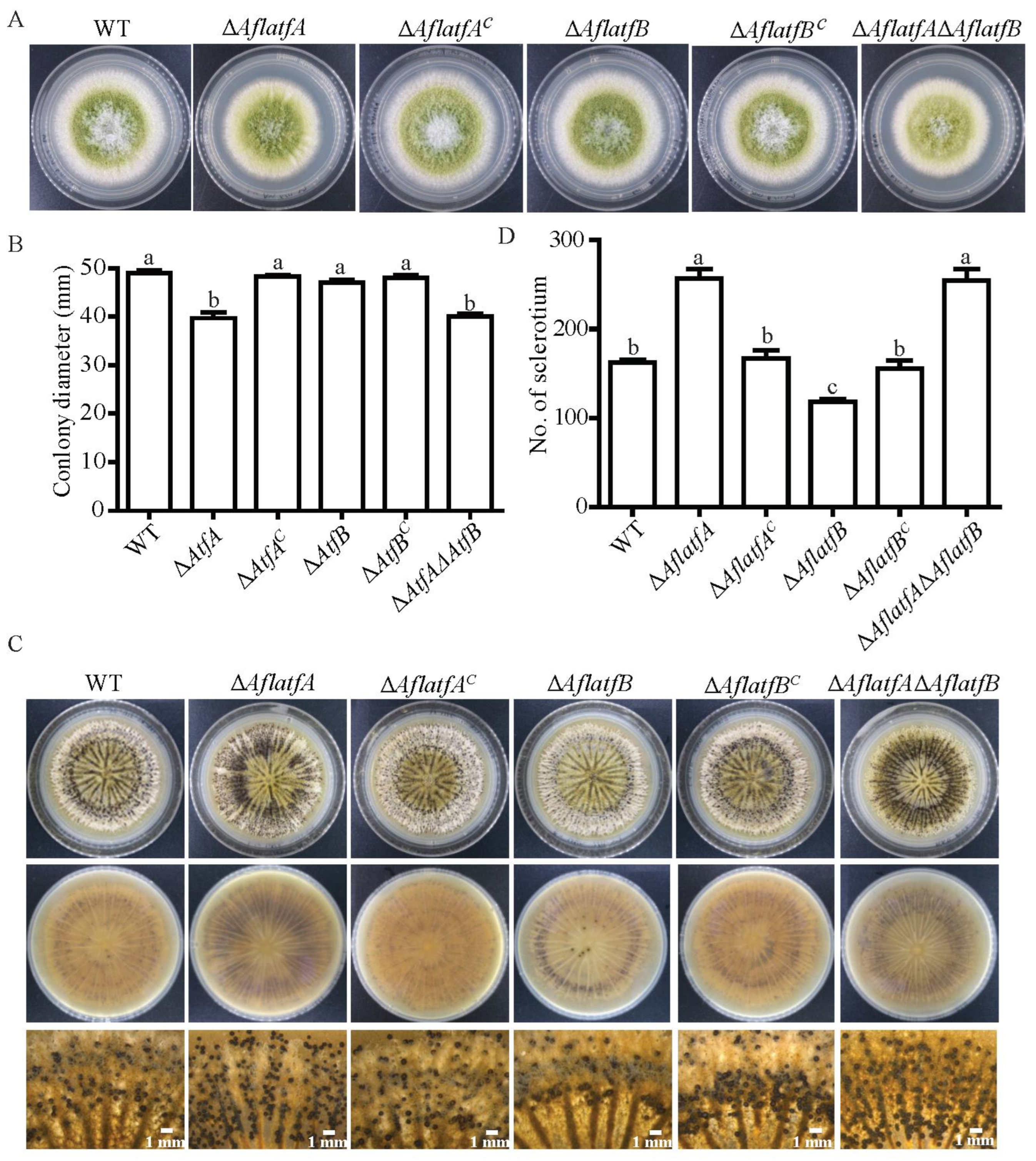
2.3. The Impact of AflatfA and AflatfB on Vegetative Growth and Sclerotia Formation of A. flavus
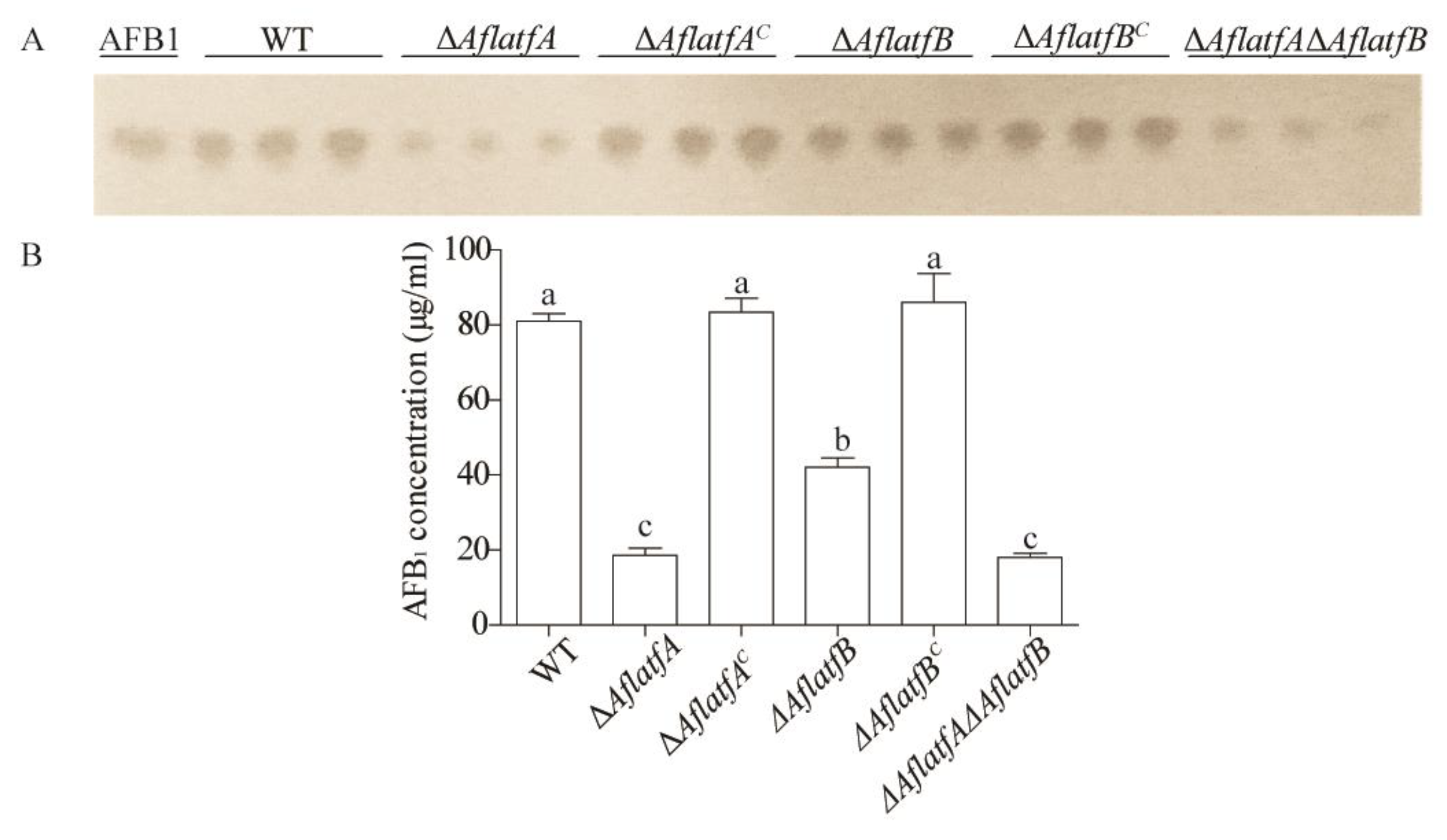
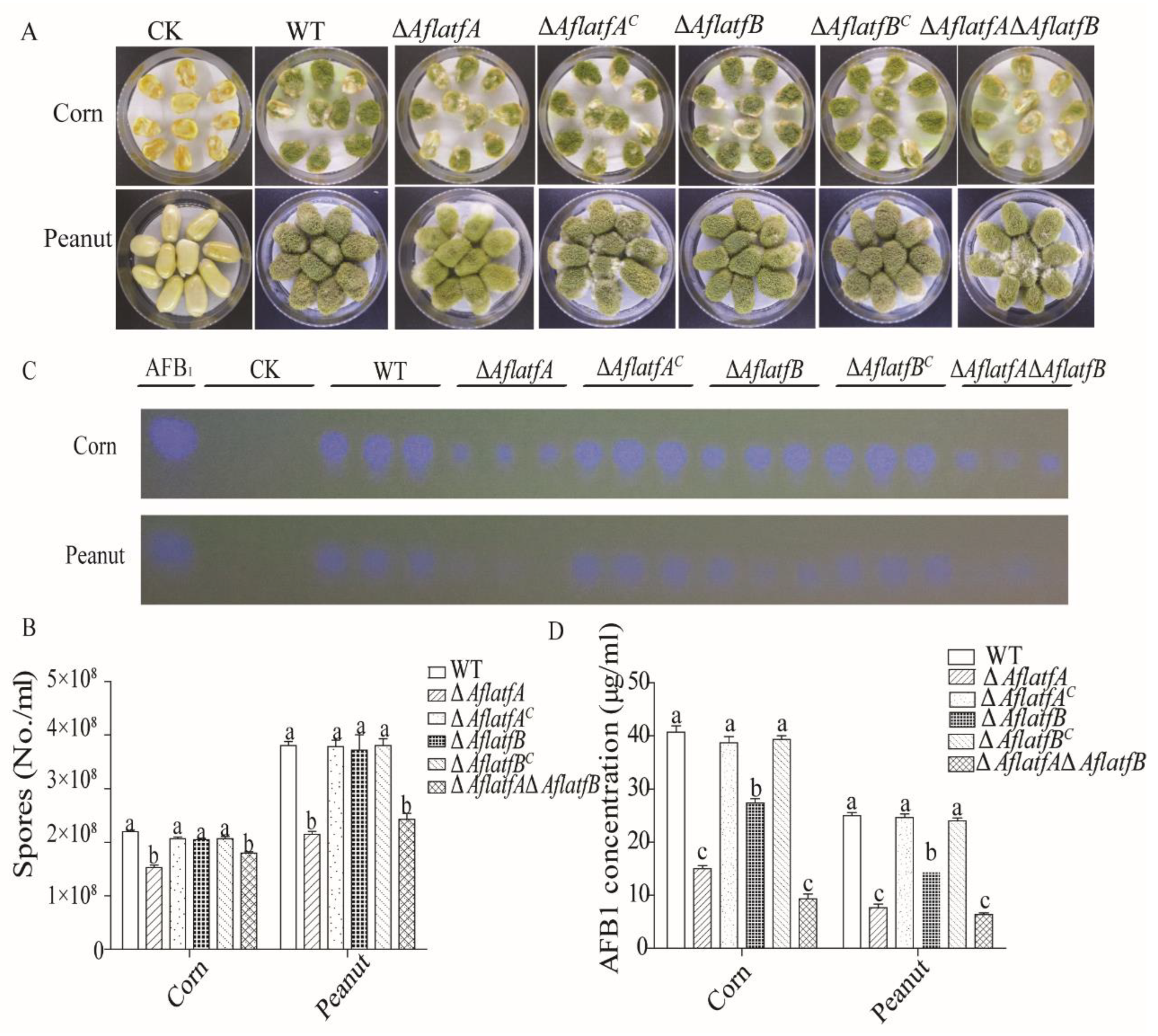
2.4. The Decline of AFB1 Yield from ΔAflatfA, ΔAflatfB and ΔAflatfAΔAflatfB Strains on Artificial Medium YES
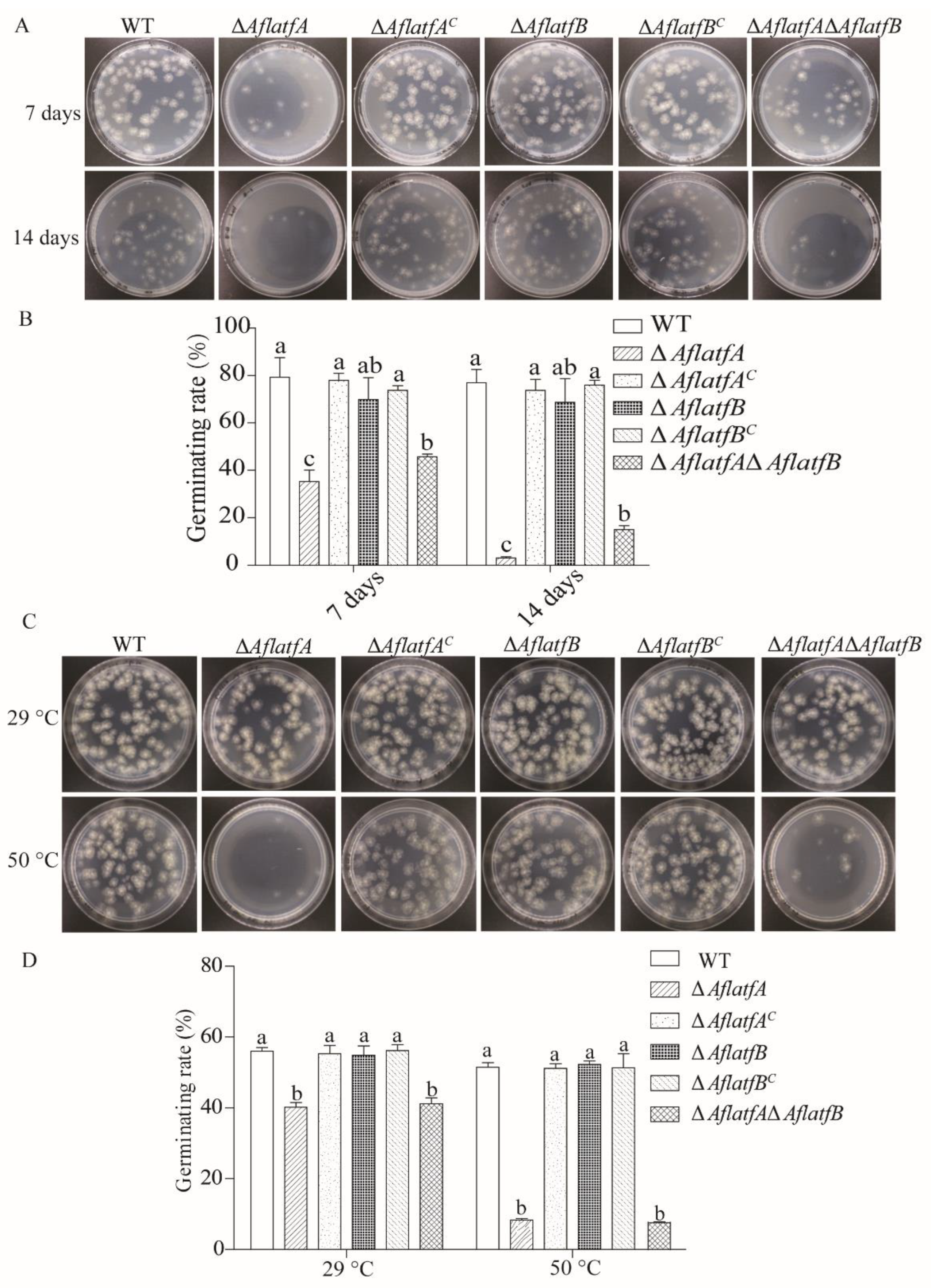
2.5. The ΔAflatfA Strain Has a Defect in the Conidia Response to Temperature Stress
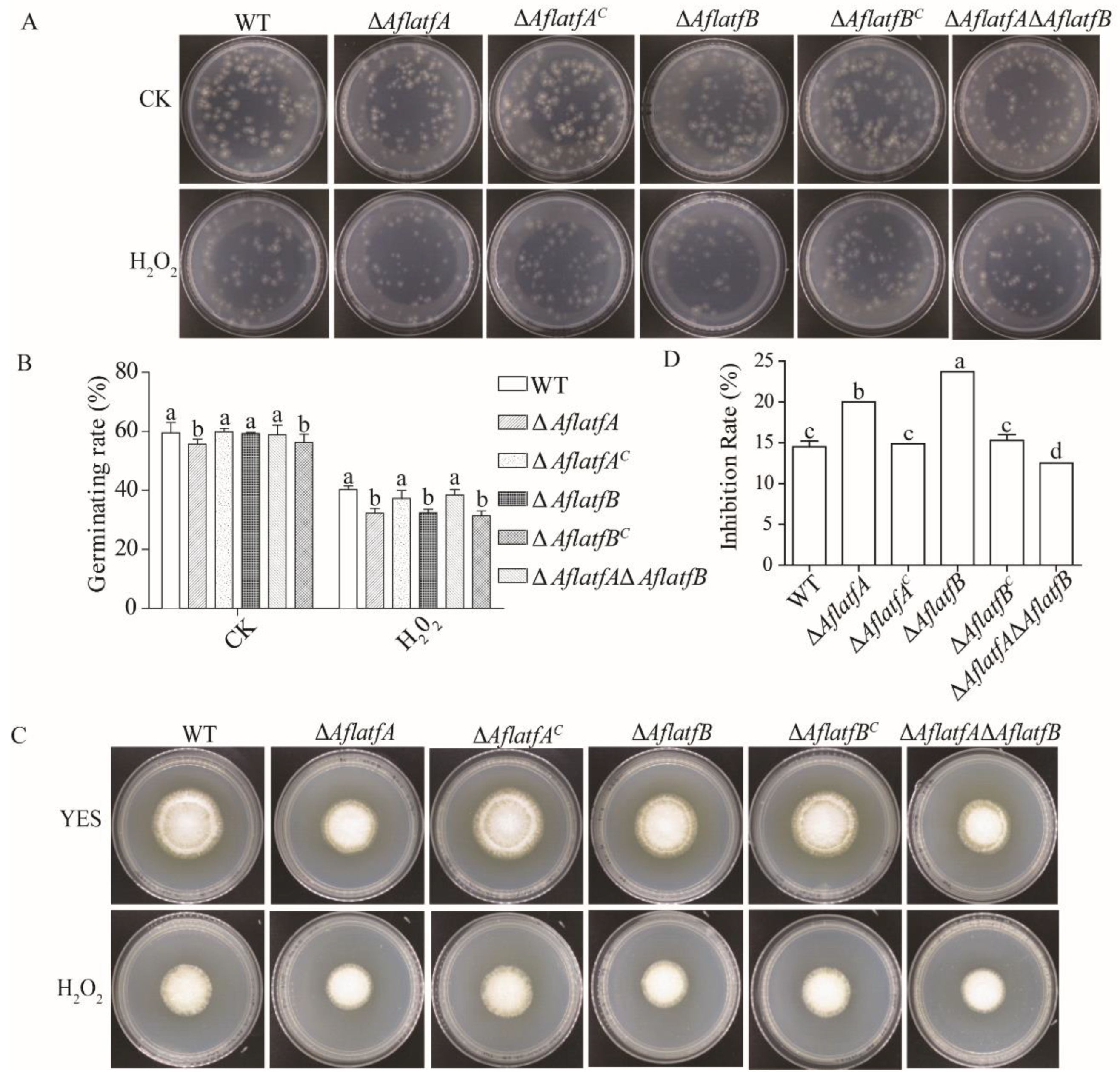
2.6. Deletion of AflatfA and AflatfB Increases Sensitivity to Oxidative Stress
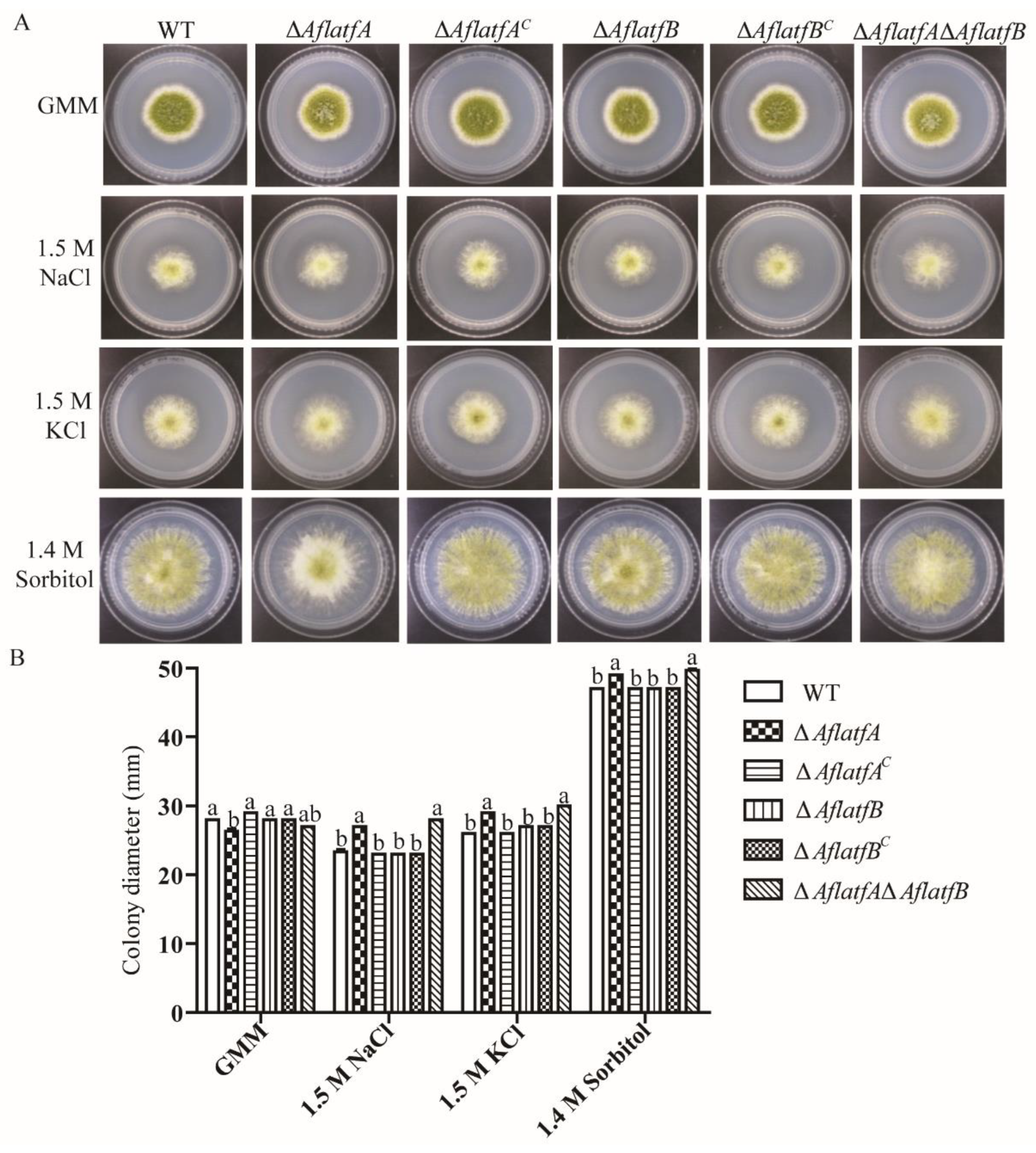
2.7. AflatfA but AflatfB Contributes to Osmotic Stress Response
2.8. Effect of AflatfA and AflatfB on Pathogenicity of A. flavus
3. Discussion
4. Materials and Methods
4.1. Fungal Strains and Culturing Conditions
4.2. Identification, Gene Composition and Phylogentic Analysis of AflatfA and AflatfB
4.3. RNA Isolation and Reverse-Transcription PCR of AflatfA and AflatfB
4.4. Gene Deletion and Complementation of AflatfA and AflatfB
4.5. Observation of Developmental Phenotype
4.6. Oxidative Stress Bioassay
4.7. Osmotic Stress Bioassay
4.8. Temperature Treatment Assay
4.9. Aflatoxin B1 Production Analysis
4.10. Virulence Assays on Corn and Peanut
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Brakhage, A.A. Regulation of fungal secondary metabolism. Nat. Rev. Microbiol. 2013, 11, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Leiter, É.; Emri, T.; Pákozdi, K.; Hornok, L.; Pócsi, I. The impact of bZIP Atf1 ortholog global regulators in fungi. Appl. Microbiol. Biotechnol. 2021, 105, 5769–5783. [Google Scholar] [CrossRef] [PubMed]
- Amoutzias, G.D.; Veron, A.S.; Weiner, J.; Robinson-Rechavi, M.; Bornberg-Bauer, E.; Oliver, S.G.; Robertson, D.L. One billion years of bZIP transcription factor evolution: Conservation and change in dimerization and DNA-binding site specificity. Mol. Biol. Evol. 2007, 24, 827–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, M.G.; Samuels, M.; Takeda, T.; Toone, W.M.; Shieh, J.C.; Toda, T.; Millar, J.B.; Jones, N. The Atf1 transcription factor is a target for the Sty1 stress-activated MAP kinase pathway in fission yeast. Genes Dev. 1996, 10, 2289–2301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiozaki, K.; Russell, P. Conjugation, meiosis, and the osmotic stress response are regulated by Spc1 kinase through Atf1 transcription factor in fission yeast. Genes Dev. 1996, 10, 2276–2288. [Google Scholar] [CrossRef] [Green Version]
- Takeda, T.; Toda, T.; Kominami, K.; Kohnosu, A.; Yanagida, M.; Jones, N. Schizosaccharomyces pombe atf1+ encodes a transcription factor required for sexual development and entry into stationary phase. EMBO J. 1995, 14, 6193–6208. [Google Scholar] [CrossRef]
- Watanabe, Y.; Yamamoto, M. Schizosaccharomyces pombe pcr1+ encodes a CREB/ATF protein involved in regulation of gene expression for sexual development. Mol. Cell. Biol. 1996, 16, 704–711. [Google Scholar] [CrossRef] [Green Version]
- Kon, N.M.; Krawchuk, D.; Warren, B.G.; Smith, G.R.; Wahls, W.P. Transcription factor Mts1/Mts2 (Atf1/Pcr1, Gad7/Pcr1) activates the M26 meiotic recombination hotspot in Schizosaccharomyces pombe. Proc. Natl. Acad. Sci. USA 1997, 94, 13765–13770. [Google Scholar] [CrossRef] [Green Version]
- Koda, W.; Senmatsu, S.; Abe, T.; Hoffman, C.S.; Hirota, K. Reciprocal stabilization of transcription factor binding integrates two signaling pathways to regulate fission yeast fbp1 transcription. Nucleic Acids Res. 2021, 49, 9809–9820. [Google Scholar] [CrossRef]
- Nickels, J.F.; Della-Rosa, M.E.; Goyeneche, I.M.; Charlton, S.J.; Sneppen, K.; Thon, G. The transcription factor Atf1 lowers the transition barrier for nucleosome-mediated establishment of heterochromatin. Cell Rep. 2022, 39, 110828. [Google Scholar] [CrossRef]
- Sansó, M.; Gogol, M.; Ayté, J.; Seidel, C.; Hidalgo, E. Transcription factors Pcr1 and Atf1 have distinct roles in stress- and Sty1- dependent gene regulation. Eukaryot. Cell 2008, 7, 826–835. [Google Scholar] [CrossRef] [Green Version]
- Bandyopadhyay, S.; Ghosh, P.M.; Basu, S.; Paul, M.; Alam, S.B.; Das, E.; Sundaram, G. Antagonistic regulation of cyclin expression by the bZIP transcription factors Pcr1 and Atf1 during G2/M transition. FEMS Microbiol. Lett. 2017, 364, 14. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, C.L.; Jones, N.; Wilkinson, C.R. Stress-induced phosphorylation of S. pombe Atf1 abrogates its interaction with F box protein Fbh1. Curr. Biol. 2009, 19, 1907–1911. [Google Scholar] [PubMed] [Green Version]
- García, P.; Paulo, E.; Gao, J.; Wahls, W.P.; Ayté, J.; Lowy, E.; Hidalog, E. Binding of the transcription factor Atf1 to promoters serves as a barrier to phase nucleosome arrays and avoid cryptic transcription. Nucleic Acids Res. 2014, 42, 10351–10359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagiwara, D.; Takahashi, H.; Kusuya, Y.; Kawamoto, S.; Kamei, K.; Gonoi, T. Comparative transcriptome analysis revealing dormant conidia and germination associated genes in Aspergillus species: An essential role for AtfA in conidial dormancy. BMC Genom. 2016, 17, 358. [Google Scholar] [CrossRef] [Green Version]
- Sakamot, K.; Iwashita, K.; Yamada, O.; Kobayashi, K.; Mizuno, A.; Akita, O.; Mikami, S.; Shimoi, H.; Gomi, K. Aspergillus oryzae atfA controls conidial germination and stress tolerance. Fungal Genet. Biol. 2009, 46, 887–897. [Google Scholar] [CrossRef]
- Temme, N.; Oeser, B.; Massaroli, M.; Heller, J.; Simon, A.; Collado, I.G.; Viaud, M.; Tudzynski, P. BcAtf1, a global regulator, controls various differentiation processes and phytotoxin production in Botrytis cinerea. Mol. Plant Pathol. 2012, 13, 704–718. [Google Scholar] [CrossRef]
- Guo, M.; Guo, W.; Chen, Y.; Dong, S.; Zhang, X.; Zhang, X.; Zhang, H.; Song, W.; Wang, W.; Wang, Q.; et al. The basic leucine zipper transcription factor Moatf1 mediates oxidative stress responses and is necessary for full virulence of the rice blast fungus Magnaporthe oryzae. Mol. Plant Microbe Interact. 2010, 23, 1053–1068. [Google Scholar] [CrossRef] [Green Version]
- Szabó, Z.; Pákozdi, K.; Murvai, K.; Pusztahelyi, T.; Kecskeméti, Á.; Gáspár, A.; Logrieco, A.F.; Emri, T.; Ádám, A.L.; Leiter, É.; et al. FvatfA regulates growth, stress tolerance as well as mycotoxin and pigment productions in Fusarium verticillioides. Appl. Microbiol. Biotechnol. 2020, 104, 7879–7899. [Google Scholar] [CrossRef]
- Colot, H.V.; Park, G.; Turner, G.E.; Ringelberg, C.; Crew, C.M.; Litvnkova, L.; Weiss, R.L.; Borkovich, K.A.; Duanlap, J.C. A high-throughput gene knockout procedure for Neurospora reveals functions for multiple transcription factors. Proc. Natl. Acad. Sci. USA 2006, 103, 10352–10357. [Google Scholar] [CrossRef]
- Nguyen, T.V.; Kröger, C.; Bönnighausen, J.; Schäfer, W.; Jörg, B. The ATF/CREB transcription factor Atf1 is essential for full virulence, deoxynivalenol production, and stress tolerance in the cereal pathogen Fusarium graminearum. Mol. Plant Microbe Interact. 2013, 26, 1378–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, C.; Zhang, S.; Zhang, Q.; Tao, Y.; Wang, C.; Xu, J. FgSKN7 and FgATF1 have overlapping functions in ascosporogenesis, pathogenesis and stress responses in Fusarium graminearum. Environ. Microbiol. 2015, 17, 1245–1260. [Google Scholar] [CrossRef]
- Fang, Y.; Xiong, D.; Tian, L.; Tang, C.; Wang, Y.; Tian, C. Functional characterization of two bZIP transcription factors in Verticillium dahliae. Gene 2017, 626, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Nathues, E.; Joshi, S.; Tenberge, K.B.; von den Driesch, M.; Oeser, B.; Bäumer, N.; Mihlan, M.; Tudzynski, P. CPTF1, a CREB-like transcription factor, is involved in the oxidative stress response in the phytopathogen Claviceps purpurea and modulates ROS level in its host Secale cereale. Mol. Plant Microbe Interact. 2004, 17, 383–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balázs, A.; Pócsi, I.; Hamari, Z.; Leiter, É.; Emri, T.; Miskei, M.; Oláh, J.; Tóth, V.; Hegedus, N.; Prade, R.A.; et al. AtfA bZIP-type transcription factor regulates oxidative and osmotic stress responses in Aspergillus nidulans. Mol. Genet Genom. 2010, 283, 289–303. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, D.; Asano, Y.; Yamashino, T.; Mizuno, T. Characterization of bZip-type transcription factor AtfA with reference to stress responses of conidia of Aspergillus nidulans. Biosci. Biotechnol. Biochem. 2008, 72, 2756–2760. [Google Scholar] [CrossRef] [Green Version]
- Hagiwara, D.; Suzuki, S.; Kamei, K.; Gonoi, T.; Kawamoto, S. The role of AtfA and HOG MAPK pathway in stress tolerance in conidia of Aspergillus fumigatus. Fungal Genet. Biol. 2014, 73, 138–149. [Google Scholar] [CrossRef]
- Lara-Rojas, F.; Sanchez, O.; Kawasaki, L.; Aguirre, J. Aspergillus nidulans transcription factor AtfA interacts with the MAPK SakA to regulate general stress responses, development and spore functions. Mol. Microbiol. 2011, 80, 436–454. [Google Scholar] [CrossRef] [Green Version]
- Emri, T.; Szarvas, V.; Orosz, E.; Antal, K.; Park, H.; Han, K.H.; Yu, J.H.; Pócsi, I. Core oxidative stress response in Aspergillus nidulans. BMC Genom. 2015, 16, 478. [Google Scholar] [CrossRef] [Green Version]
- Silva, L.P.; Horta, M.A.C.; Goldman, G.H. Genetic interactions between Aspergillus fumigatus basic leucine zipper (bZIP) transcription factors AtfA, AtfB, AtfC and AtfD. Front. Fungal Biol. 2021, 2, 632048. [Google Scholar] [CrossRef]
- Nimmanee, P.; Woo, P.C.Y.; Vanittanakom, P.; Youngchim, S.; Vanittanakom, N. Functional analysis of atfA gene to stress response in pathogenic thermal dimorphic fungus Penicillium marneffei. PLoS ONE 2014, 9, e111200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, X.; Guo, L.; Yang, L.; Huang, J. Foatf1, a bZIP transcription factor of Fusarium oxysporum f. sp. cubense is involved in pathogenesis by regulating the oxidative stress responses of Cavendish banana (Musa spp.). Physiol. Mol. Plant Pathol. 2013, 84, 76–85. [Google Scholar]
- Hagiwara, D.; Asano, Y.; Marui, J.; Yoshimi, A.; Mizuno, T.; Abe, K. Transcriptional profiling for Aspergillus nidulans HogA MAPK signaling pathway in response to fludioxonil and osmotic stress. Fungal Genet. Biol. 2009, 46, 868–878. [Google Scholar] [CrossRef]
- Yamashita, K.; Shiozawa, A.; Watanabe, S.; Fukumori, F.; Kimura, M.; Fujimura, M. ATF-1 transcription factor regulates the expression of ccg-1 and cat-1 genes in response to fludioxonil under OS-2 MAP kinase in Neurospora crassa. Fungal Genet. Biol. 2008, 45, 1562–1569. [Google Scholar] [CrossRef] [PubMed]
- Valero, C.; Colabardini, A.C.; Chiaratto, J.; Pardeshi, L.; de Castro, P.A.; Filho, J.A.F.; Silva, L.P.; Rocha, M.C.; Malavazi, I.; Costa, J.H.; et al. Aspergillus fumigatus transcription factors involved in the caspofungin paradoxical effect. mBio 2020, 11, e00816-20. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.P.; Alves de Castro, P.; Dos Reis, T.F.; Paziani, M.H.; Von Zeska Kress, M.R.; Riaño-Pachón, D.M.; Hagiwara, D.; Ries, L.N.A.; Brown, N.A.; Goldman, G.H. Genome-wide transcriptome analysis of Aspergillus fumigatus exposed to osmotic stress reveals regulators of osmotic and cell wall stresses that are SakAHOG1 and MpkC dependent. Cell. Microbiol. 2017, 19, e12681. [Google Scholar] [CrossRef] [PubMed]
- Emri, T.; Gila, B.; Antal, K.; Fekete, F.; Moon, H.; Yu, J.-H.; Pócsi, I. AtfA-independent adaptation to the toxic heavy metal cadmium an Aspergillus nidulans. Microorganisms 2021, 9, 1433. [Google Scholar] [CrossRef]
- Tang, C.; Li, T.; Klosterman, S.J.; Tian, C.; Wang, Y. The bZIP transcription factor VdAtf1 regulates virulence by mediating nitrogen metabolism in Verticillium dahliae. New Phytol. 2020, 226, 1461–1479. [Google Scholar] [CrossRef]
- Jung, M.-G.; Kim, S.S.; Yu, J.-H.; Shin, K.S. Characterization of gprK encoding a putative hybrid G-protein-coupled receptor in Aspergillus fumigatus. PLoS ONE 2016, 11, e0161312. [Google Scholar] [CrossRef] [Green Version]
- Gressler, M.; Meyer, F.; Heine, D.; Hortschansky, P.; Hertweck, C.; Brock, M. Phytotoxin production in Aspergillus terreus is regulated by independent environmental signals. eLife 2015, 4, e07861. [Google Scholar] [CrossRef]
- Pérez-Arques, C.; Navarro-Mendoza, M.I.; Murcia, L.; Lax, C.; Martínez-García, P.; Heitman, J.; Nicolás, F.E.; Garre, V. Mucor circinelloides thrives inside the phagosome through an Atf-mediated germination pathway. mBio 2019, 10, e02765-18. [Google Scholar] [CrossRef]
- Missall, T.A.; Lodge, J.K. Function of the thioredoxin proteins in during stress or virulence and regulation by putative transcriptional modulators. Mol. Microbiol. 2005, 57, 847–858. [Google Scholar] [CrossRef]
- Sakamoto, K.; Arima, T.; Iwashita, K.; Yamada, O.; Gomi, K.; Akita, O. Aspergillus oryzae atfB encodes a transcription factor required for stress tolerance in conidia. Fungal Genet. Biol. 2008, 45, 922–932. [Google Scholar] [CrossRef] [PubMed]
- Roze, L.V.; Miller, M.J.; Rarick, M.; Mahanti, N.; Linz, J.E. A novel cAMP-response element, CRE1, modulates expression of nor-1 in Aspergillus parasiticus. J. Biol. Chem. 2004, 279, 27428–27439. [Google Scholar] [CrossRef] [Green Version]
- Roze, L.V.; Chanda, A.; Wee, J.; Awad, D.; Linz, J.E. Stress-related transcription factor AtfB integrates secondary metabolism with oxidative stress response in Aspergilli. J. Biol. Chem. 2011, 286, 35137–35148. [Google Scholar] [CrossRef] [Green Version]
- Baidya, S.; Duran, R.M.; Lohmar, J.M.; Harris-Coward, P.Y.; Cary, J.W.; Hong, S.Y.; Roze, L.V.; Linz, J.E.; Calvo, A.M. VeA is associated with the response to oxidative stress in the aflatoxin producer Aspergillus flavus. Eukaryot. Cell 2014, 13, 1095–1103. [Google Scholar] [CrossRef] [Green Version]
- Wee, J.; Hong, S.Y.; Roze, L.V.; Day, D.M.; Chanda, A.; Linz, J.E. The fungal bZIP transcription factor AtfB controls virulence-associated processes in Aspergillus parasiticus. Toxins 2017, 9, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pleadin, J.; Frece, J.; Markov, K. Mycotoxins in food and feed. Adv. Food Nutr. Res. 2019, 89, 297–345. [Google Scholar] [PubMed]
- Dey, D.K.; Kang, J.I.; Bajpai, V.K.; Kim, K.; Lee, H.; Sonwal, S.; Simal-Gandara, J.; Xiao, J.; Ali, S.; Huh, Y.S.; et al. Mycotoxins in food and feed: Toxicity, preventive challenges, and advanced detection techniques for associated diseases. Crit. Rev. Food Sci. 2022, 21, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Hatabayashi, H.; Arai, H.; Kitamoto, H.K.; Yabe, K. Function of the cypX and moxY genes in aflatoxin biosynthesis in Aspergillus parasiticus. Appl. Environ. Microbiol. 2005, 71, 3192–3198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickova, D.; Ostry, V.; Toman, J.; Malir, F. Aflatoxins: History, significant milestones, recent data on their toxicity and ways to mitigation. Toxins 2021, 13, 399. [Google Scholar] [CrossRef] [PubMed]
- Amaike, S.; Keller, N.P. Aspergillus flavus. Annu. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef] [PubMed]
- Caceres, I.; Khoury, A.A.; Khoury, R.E.; Lorber, S.; Oswald, I.P.; Khoury, A.E.; Atoui, A.; Puel, O.; Bailly, J.D. Aflatoxin biosynthesis and genetic regulation: A review. Toxins 2019, 12, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caceres, I.; Khoury, E.R.; Bailly, S.; Oswald, I.P.; Puel, O.; Bailly, J.-D. Piperine inhibits aflatoxin B1 production in Aspergillus flavus by modulating fungal oxidative stress response. Fungal Genet. Biol. 2017, 107, 77–85. [Google Scholar] [CrossRef]
- Xu, D.; Peng, S.; Guo, R.; Yao, L.; Mo, H.; Li, H.; Song, H.; Hu, L. EGCG alleviates oxidative stress and inhibits aflatoxin B1 biosynthesis via MAPK signaling pathway. Toxins 2021, 13, 693. [Google Scholar] [CrossRef]
- Ma, L.; Li, X.; Ma, X.; Yu, Q.; Yu, X.; Liu, Y.; Nie, C.; Zhang, Y.; Xing, F. The regulatory mechanism of water activities on aflatoxins biosynthesis and conidia development, and transcription factor AtfB is involved in this regulation. Toxins 2021, 13, 431. [Google Scholar] [CrossRef]
- Li, X.; Ren, Y.; Jing, J.; Jiang, Y.; Yang, Q.; Luo, S.; Xing, F. The inhibitory mechanism of methyl jasmonate on Aspergillus flavus growth and aflatoxin biosynthesis and two novel transcription factors are involved in this action. Foo. Res. Int. 2021, 140, 110051. [Google Scholar] [CrossRef]
- Zhao, Q.; Pei, H.; Zhao, X.; Zhao, K.; Yu, M.; Han, G.; Fan, J.; Tao, F. Systematic characterization of bZIP transcription factors required for development and aflatoxins generation by high-throughput gene knockout in Aspergillus flavus. J. Fungi 2022, 8, 356. [Google Scholar] [CrossRef]
- Kim, M.S.; Ko, Y.-J.; Maeng, S.; Floyd, A.; Heitman, J.; Bahn, Y.-S. Comparative transcriptome analysis of the CO2 sensing pathway via differential expression of carbonic anhydrase in Cryptococcus neoformans. Genetics 2010, 185, 1207–1219. [Google Scholar] [CrossRef] [Green Version]
- Chang, P.K.; Scharfenstein, L.L.; Wei, Q.; Bhatnagar, D. Development and refinement of high-efficiency gene-targeting system for Aspergillus flavus. J. Microbiol. Methods. 2010, 81, 240–246. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Szewczyk, E.; Nayak, T.; Oakley, C.E.; Edgerton, H.; Xiong, Y.; Taheri-Talesh, N.; Osmani, S.A.; Oakley, B.R. Fusion PCR and gene targeting in Aspergillus nidulans. Nat. Protoc. 2006, 1, 3111–3120. [Google Scholar] [CrossRef]
- Chang, P.K.; Ehrlich, K.C.; Yu, J.J.; Bhatnagar, D.; Cleveland, T.E. Increased expression of Aspergillus parasiticus AflR, encoding a sequence-specific DNA-binding protein, relieves nitrate inhibition of aflatoxin biosynthesis. Appl. Environ. Microbiol. 1995, 61, 2372–2377. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Description | Reference |
|---|---|---|
| Recipient strain | PTSΔku70ΔpyrG | [60] |
| wild type (WT) | PTSΔku70ΔpyrG::AfpyrG | [60] |
| ΔAflatfA | Δku70ΔpyrGΔAflatfA::pyrG | This study |
| ΔAflatfB | Δku70ΔpyrGΔAflatfB::pyrG | This study |
| ΔAflatfAC | Δku70ΔpyrGΔAflatfA::pyrG, AflatfA::pyrG | This study |
| ΔAflatfBC | Δku70ΔpyrGΔAflatfB::pyrG, AflatfB::pyrG | This study |
| ΔAflatfAΔAflatfB | Δku70ΔpyrGΔAflatfA::pyrGΔAflatfB::ptrA | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Zha, W.; Yao, B.; Yang, L.; Wang, S. Genetic Interaction of Global Regulators AflatfA and AflatfB Mediating Development, Stress Response and Aflatoxins B1 Production in Aspergillus flavus. Toxins 2022, 14, 857. https://doi.org/10.3390/toxins14120857
Wang X, Zha W, Yao B, Yang L, Wang S. Genetic Interaction of Global Regulators AflatfA and AflatfB Mediating Development, Stress Response and Aflatoxins B1 Production in Aspergillus flavus. Toxins. 2022; 14(12):857. https://doi.org/10.3390/toxins14120857
Chicago/Turabian StyleWang, Xiuna, Wenjie Zha, Bin Yao, Lan Yang, and Shihua Wang. 2022. "Genetic Interaction of Global Regulators AflatfA and AflatfB Mediating Development, Stress Response and Aflatoxins B1 Production in Aspergillus flavus" Toxins 14, no. 12: 857. https://doi.org/10.3390/toxins14120857
APA StyleWang, X., Zha, W., Yao, B., Yang, L., & Wang, S. (2022). Genetic Interaction of Global Regulators AflatfA and AflatfB Mediating Development, Stress Response and Aflatoxins B1 Production in Aspergillus flavus. Toxins, 14(12), 857. https://doi.org/10.3390/toxins14120857